
Arbuscular Mycorrhizal Fungi Mitigate Lead Toxicity in Maize by Restructuring Rhizosphere Microbiome and Enhancing Antioxidant Defense Mechanisms

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Materials and Plot Experiment
2.1.1. Soil Preparation and Pb Contamination
2.1.2. AMF Inoculum Source and Application
2.2. Plant Cultivation and Sampling
2.2.1. Plant Growth and Biomass Measurement
2.2.2. Antioxidant Enzyme Activity and Pb Quantification
2.3. Rhizosphere Microbiome Analysis
2.3.1. Sampling and Preparation of Rhizosphere Soil
2.3.2. Diversity and Structure of the Microbial Community
2.4. Data Analysis
3. Results
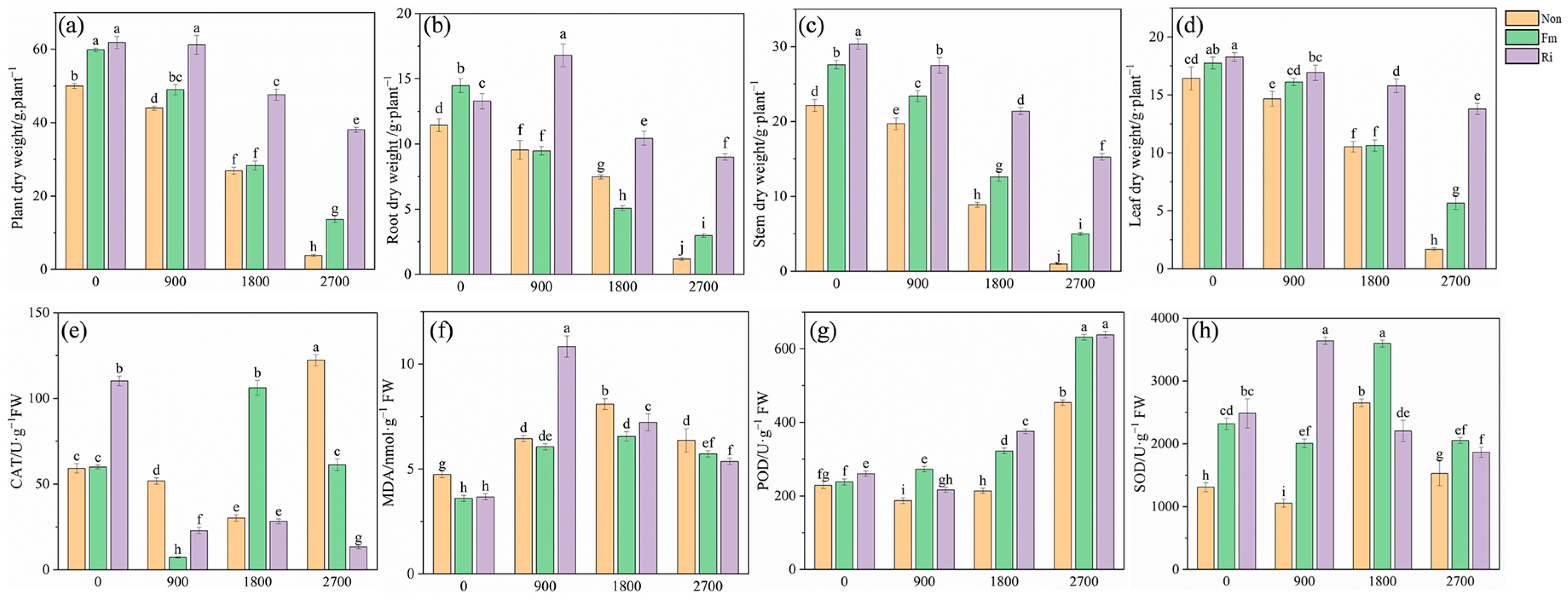
3.1. Plant Biomass and Physicochemical Properties
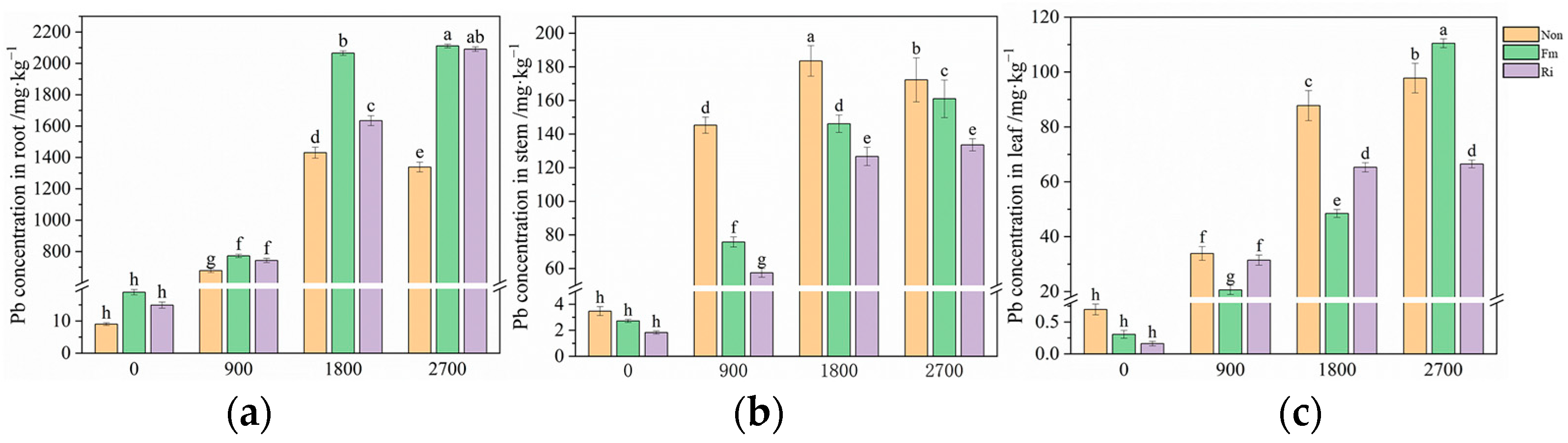
3.2. Pb Uptake and Translocation in Maize Under Different Pb Treatments
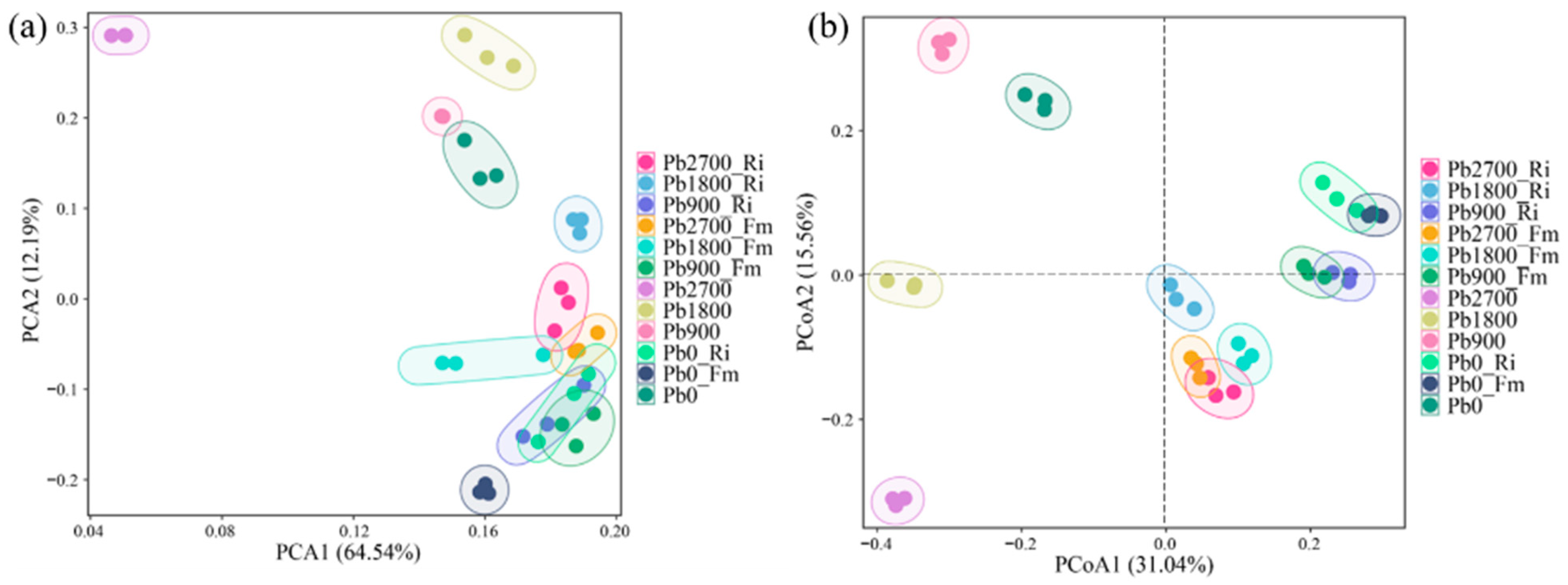
3.3. Diversity of Microbial Communities in the Rhizosphere Soils
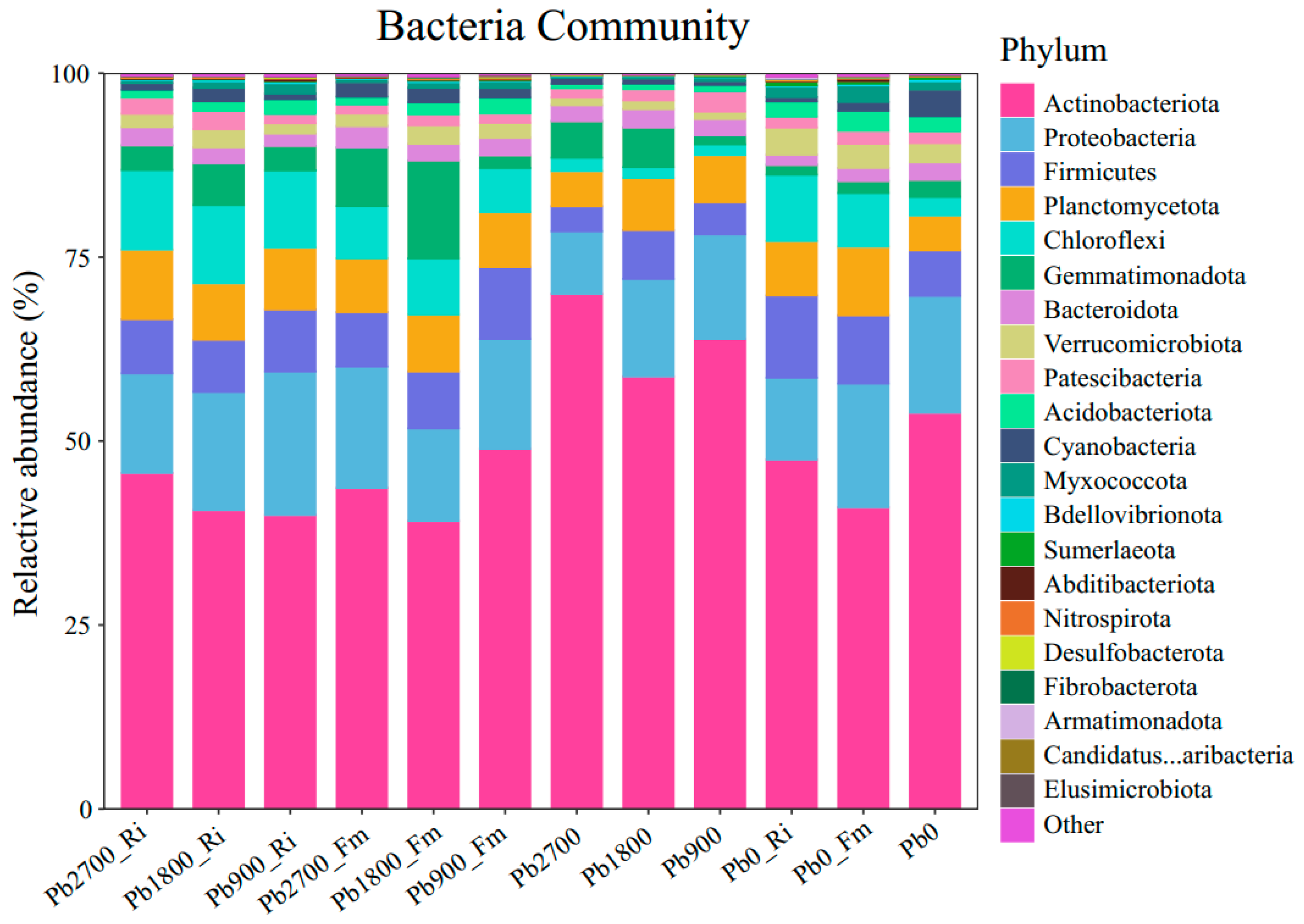
3.4. Microbial Community Composition in the Rhizosphere Soils
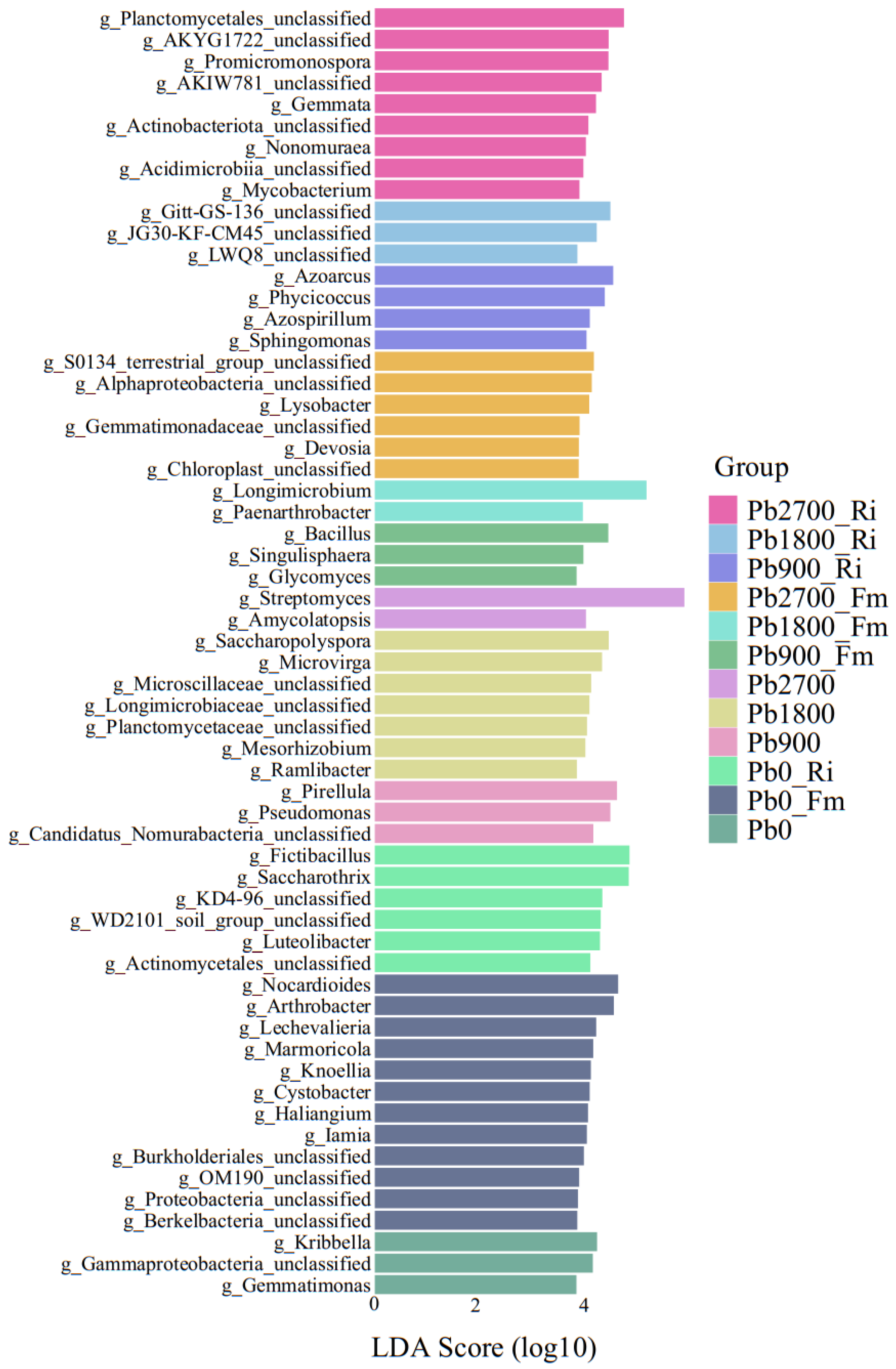
3.5. Analysis of Differences in Microbial Community
4. Discussion
4.1. Implications of AMF Inoculation for Maize Resistance to Pb Stress
4.2. Effects of AMF on Microbial Diversity and Composition in the Rhizosphere Soil of Maize
4.3. Impact of Pb Stress and AMF on Key Rhizosphere Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, D.; Yu, W.; Hao, Z.; Qiu, M.; Cui, J.; Tang, Y.; Teng, X.; Liu, Y.; Liu, H. Molecular mechanism of selenium against lead-induced apoptosis in chicken brainstem relating to heat shock protein, selenoproteins, and inflammatory cytokines. Ecotoxicol. Environ. Saf. 2024, 272, 116028. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Teng, Y.; Lu, S.; Wang, Y.; Wang, J. Contamination features and health risk of soil heavy metals in China. Sci. Total Environ. 2015, 512–513, 143–153. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Dubey, S.; Shri, M.; Gupta, A.; Rani, V.; Chakrabarty, D. Toxicity and detoxification of heavy metals during plant growth and metabolism. Environ. Chem. Lett. 2018, 16, 1169–1192. [Google Scholar] [CrossRef]
- Soares, T.; Dias, D.; Oliveira, A.M.S.; Ribeiro, D.M.; Dias, L. Exogenous brassinosteroids increase lead stress tolerance in seed germination and seedling growth of Brassica juncea L. Ecotoxicol. Environ. Saf. 2020, 193, 110296. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, L.; Wang, C.; Liu, P.; Ma, L.; Zou, C.; Pan, G.; Shen, Y. Combined genome-wide association study and gene co-expression network analysis identified ZmAKINβγ1 involved in lead tolerance and accumulation in maize seedlings. Int. J. Biol. Macromol. 2023, 226, 1374–1386. [Google Scholar] [CrossRef]
- Kanwar, V.S.; Sharma, A.; Srivastav, A.L.; Rani, L. Phytoremediation of toxic metals present in soil and water environment: A critical review. Environ. Sci. Pollut. Res. 2020, 27, 44835–44860. [Google Scholar] [CrossRef]
- Wei, Z.; Hao, Z.; Li, X.; Guan, Z.; Cai, Y.; Liao, X. The effects of phytoremediation on soil bacterial communities in an abandoned mine site of rare earth elements. Sci. Total Environ. 2019, 670, 950–960. [Google Scholar] [CrossRef]
- Jin, Y.; Luan, Y.; Ning, Y.; Wang, L. Effects and mechanisms of microbial remediation of heavy metals in soil: A critical review. Appl. Sci. 2018, 8, 1336. [Google Scholar] [CrossRef]
- Zhu, H.; Hu, L.; Wang, Y.; Mei, P.; Zhou, F.; Rozhkova, T.; Li, C. Effects of Streptomyces sp. HU2014 inoculation on wheat growth and rhizosphere microbial diversity under hexavalent chromium stress. Ecotoxicol. Environ. Saf. 2024, 276, 116313. [Google Scholar] [CrossRef]
- Khalid, M.; Saeed, U.-R.; Hassani, D.; Hayat, K.; Pei, Z.; Nan, H. Advances in fungal-assisted phytoremediation of heavy metals: A review. Pedosphere 2021, 31, 475–495. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, Y.; Han, X.; Chiu, T.-Y.; Ghosh, A.; Chen, H.; Tang, M. The roles of arbuscular mycorrhizal fungi (AMF) in phytoremediation and tree-herb interactions in Pb contaminated soil. Sci. Rep. 2016, 6, 20469. [Google Scholar] [CrossRef] [PubMed]
- Arora, K.; Sharma, S.; Monti, A. Bio-remediation of Pb and Cd polluted soils by switchgrass: A case study in India. Int. J. Phytoremediat. 2016, 18, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Shi, Z.; Lu, S.; Wang, F. AMF Inoculation Alleviates Molybdenum Toxicity to Maize by Protecting Leaf Performance. J. Fungi 2023, 9, 479. [Google Scholar] [CrossRef]
- Wipf, D.; Krajinski, F.; van Tuinen, D.; Recorbet, G.; Courty, P.E. Trading on the arbuscular mycorrhiza market: From arbuscules to common mycorrhizal networks. New Phytol. 2019, 223, 1127–1142. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, B.; Xu, J.; Li, Z.; Tang, Z.; Wu, X. Heavy metal domestication enhances beneficial effects of arbuscular mycorrhizal fungi on lead (Pb) phytoremediation efficiency of Bidens parviflora through improving plant growth and root Pb accumulation. Environ. Sci. Pollut. Res. 2022, 29, 32988–33001. [Google Scholar] [CrossRef]
- Wang, H.-R.; Zhao, X.-Y.; Zhang, J.-M.; Lu, C.; Feng, F.-J. Arbuscular mycorrhizal fungus regulates cadmium accumulation, migration, transport, and tolerance in Medicago sativa. J. Hazard. Mater. 2022, 435, 129077. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, Z.; Yang, X.; Sheng, L.; Mao, H.; Zhu, S. Metagenomics reveal arbuscular mycorrhizal fungi altering functional gene expression of rhizosphere microbial community to enhance Iris tectorum’s resistance to Cr stress. Sci. Total Environ. 2023, 895, 164970. [Google Scholar] [CrossRef]
- Zhao, W.; Zhu, S.; Yang, X.; Xia, G.; Wang, B.; Gu, B. Arbuscular mycorrhizal fungi alter rhizosphere bacterial community characteristics to improve Cr tolerance of Acorus calamus. Ecotoxicol. Environ. Saf. 2023, 253, 114652. [Google Scholar]
- Chen, X.W.; Wu, L.; Luo, N.; Mo, C.H.; Wong, M.H.; Li, H. Arbuscular mycorrhizal fungi and the associated bacterial community influence the uptake of cadmium in rice. Geoderma 2019, 337, 749–757. [Google Scholar] [CrossRef]
- Hamedi, J.; Dehhaghi, M.; Mohammdipanah, F. Isolation of extremely heavy metal resistant strains of rare actinomycetes from high metal content soils in Iran. Int. J. Environ. Res. 2015, 9, 475–480. [Google Scholar]
- Mitra, A.; Chatterjee, S.; Kataki, S.; Rastogi, R.P.; Gupta, D.K. Bacterial tolerance strategies against lead toxicity and their relevance in bioremediation application. Environ. Sci. Pollut. Res. 2021, 28, 14271–14284. [Google Scholar] [CrossRef] [PubMed]
- George, S.E.; Wan, Y. Microbial functionalities and immobilization of environmental lead: Biogeochemical and molecular mechanisms and implications for bioremediation. J. Hazard. Mater. 2023, 457, 131738. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, Y.; Chen, J.; Lin, H.; Zhao, M.; Peng, H.; Liu, L.; Yuan, G.; Zhang, S.; Zhang, Z.; et al. Genome expression profile analysis reveals important transcripts in maize roots responding to the stress of heavy metal Pb. Physiol. Plant. 2013, 147, 270–282. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, R.; Kertesz, M.A.; Zhang, F.; Feng, G. Arbuscular mycorrhizal fungal hyphae mediating acidification can promote phytate mineralization in the hyphosphere of maize (Zea mays L.). Soil Biol. Biochem. 2013, 65, 69–74. [Google Scholar] [CrossRef]
- Wang, F.; Jing, X.; Adams, C.A.; Shi, Z.; Sun, Y. Decreased ZnO nanoparticle phytotoxicity to maize by arbuscular mycorrhizal fungus and organic phosphorus. Environ. Sci. Pollut. Res. 2018, 25, 23736–23747. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Mu, M.; Wang, Z.; Chen, Z.; Wu, Y.; Nie, W.; Zhao, S.; Yin, X.; Teng, X. Physiological characteristics, rhizosphere soil properties, and root-related microbial communities of Trifolium repens L. in response to Pb toxicity. Sci. Total Environ. 2024, 907, 167871. [Google Scholar] [CrossRef]
- Wang, T.; Sun, H.; Jiang, C.; Mao, H.; Zhang, Y. Immobilization of Cd in soil and changes of soil microbial community by bioaugmentation of UV-mutated Bacillus subtilis 38 assisted by biostimulation. Eur. J. Soil Biol. 2014, 65, 62–69. [Google Scholar] [CrossRef]
- Li, Q.; Xing, Y.; Huang, B.; Chen, X.; Ji, L.; Fu, X.; Li, T.; Wang, J.; Chen, G.; Zhang, Q. Rhizospheric mechanisms of Bacillus subtilis bioaugmentation-assisted phytostabilization of cadmium-contaminated soil. Sci. Total Environ. 2022, 825, 154136. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, B.; Ma, X.; Jin, X.; Chen, S.; Wang, P.; Zhongrong, G.; Wu, X.; Zhang, H. Combining transcriptome and metabolome analyses to reveal the response of maize roots to Pb stress. Plant Physiol. Biochem. 2024, 217, 109265. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Lounes-Hadj Sahraoui, A. Arbuscular mycorrhizal fungal responses to abiotic stresses: A review. Phytochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef]
- Wood, J.L.; Tang, C.; Franks, A.E. Microbial associated plant growth and heavy metal accumulation to improve phytoextraction of contaminated soils. Soil Biol. Biochem. 2016, 103, 131–137. [Google Scholar] [CrossRef]
- Wang, X.; Fang, L.; Beiyuan, J.; Cui, Y.; Peng, Q.; Zhu, S.; Wang, M.; Zhang, X. Improvement of alfalfa resistance against Cd stress through rhizobia and arbuscular mycorrhiza fungi co-inoculation in Cd-contaminated soil. Environ. Pollut. 2021, 277, 116758. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, T.; Zhou, J.; Peng, X. Effects of Arbuscular Mycorrhizal Fungi on Robinia pseudoacacia L. Growing on Soils Contaminated with Heavy Metals. J. Fungi 2023, 9, 684. [Google Scholar] [CrossRef]
- Jia, Q.; Sun, J.; Gan, Q.; Shi, N.-N.; Fu, S. Zea mays cultivation, biochar, and arbuscular mycorrhizal fungal inoculation influenced lead immobilization. Microbiol. Spectr. 2024, 12, e03427-23. [Google Scholar] [CrossRef]
- Fatima, R.A.; Ahmad, M. Certain antioxidant enzymes of Allium cepa as biomarkers for the detection of toxic heavy metals in wastewater. Sci. Total Environ. 2005, 346, 256–273. [Google Scholar] [CrossRef]
- Fang, L.; Ju, W.; Yang, C.; Jin, X.; Liu, D.; Li, M.; Yu, J.; Zhao, W.; Zhang, C. Exogenous application of signaling molecules to enhance the resistance of legume-rhizobium symbiosis in Pb/Cd-contaminated soils. Environ. Pollut. 2020, 265, 114744. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Ma, X.; Jin, X.; Wu, X.; Zhang, H.; Guan, Z.; Fu, Z.; Chen, S.; Wang, P. Transcriptomics Combined with Physiology and Metabolomics Reveals the Mechanism of Tolerance to Lead Toxicity in Maize Seedling. Physiol. Plant. 2024, 176, e14547. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.; Yu, Z.; Shen, G.; Cheng, H.; Tao, S. Composition and diversity of soil microbial communities in the alpine wetland and alpine forest ecosystems on the Tibetan Plateau. Sci. Total Environ. 2020, 747, 141358. [Google Scholar] [CrossRef]
- Luo, C.; Deng, Y.; Liang, J.; Zhu, S.; Wei, Z.; Guo, X.; Luo, X. Exogenous rare earth element-yttrium deteriorated soil microbial community structure. J. Rare Earth. 2018, 36, 430–439. [Google Scholar] [CrossRef]
- Hao, L.; Zhang, Z.; Hao, B.; Diao, F.; Zhang, J.; Bao, Z.; Guo, W. Arbuscular mycorrhizal fungi alter microbiome structure of rhizosphere soil to enhance maize tolerance to La. Ecotoxicol. Environ. Saf. 2021, 212, 111996. [Google Scholar] [CrossRef]
- Hillebrand, H.; Matthiessen, B. Biodiversity in a complex world: Consolidation and progress in functional biodiversity research. Ecol. Lett. 2009, 12, 1405–1419. [Google Scholar] [CrossRef]
- Banerjee, S.; Schlaeppi, K.; Van Der Heijden, M.G. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Hu, J.; Wei, Z.; Friman, V.-P.; Gu, S.-h.; Wang, X.f.; Eisenhauer, N.; Yang, T.j.; Ma, J.; Shen, Q.r.; Xu, Y.c. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. mBio 2016, 7, 10–1128. [Google Scholar] [CrossRef]
- Quadros, P.D.; Zhalnina, K.; Davis-Richardson, A.G.; Drew, J.C.; Menezes, F.B.; Camargo, F.A.d.O.; Triplett, E.W. Coal mining practices reduce the microbial biomass, richness and diversity of soil. Appl. Soil Ecol. 2016, 98, 195–203. [Google Scholar] [CrossRef]
- Zhu, Y.-M.; Wei, D.-Z. Biosorption of Mycobacterium phlei to the Heavy Metal Ions Pb2+, Zn2+, Ni2+ and Cu2+. J. Northeast. Univ. Nat. Sci. 2003, 24, 91–93. [Google Scholar]
- Erardi, F.; Failla, M.; Falkinham, J., 3rd. Accumulation and transport of cadmium by tolerant and susceptible strains of Mycobacterium scrofulaceum. Antimicrob. Agents Chemother. 1989, 33, 350–355. [Google Scholar] [CrossRef]
- Laborda, P.; Ling, J.; Chen, X.; Liu, F. ACC deaminase from Lysobacter gummosus OH17 can promote root growth in Oryza sativa Nipponbare plants. J. Agric. Food Chem. 2018, 66, 3675–3682. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Guo, P.; Zhang, Z.; Cui, X.; Hao, B.; Guo, W. Arbuscular mycorrhizal fungi facilitate Astragalus adsurgens growth and stress tolerance in cadmium and lead contaminated saline soil by regulating rhizosphere bacterial community. Appl. Soil Ecol. 2023, 187, 104842. [Google Scholar] [CrossRef]
- Gan, M.; Zhou, Y.; Huang, D.; He, P.; Tang, B.; Cai, Y.; Zhu, J. The enhanced effect of key microorganisms in chromium contaminated soil in Cr(VI) reduction. Chemosphere 2024, 362, 142682. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pb Treatment/mg·kg−1 | Inoculation | Chao1 | Observed_Species | ACE | Simpson | Shannon |
|---|---|---|---|---|---|---|
| 0 | Non | 1181.88 ± 54.85 c | 1177.67 ± 54.60 d | 1185.69 ± 55.39 c | 0.97 ± 0.00 a | 7.33 ± 0.15 d |
| Fm | 1574.50 ± 273.03 a | 1573.00 ± 271.55 a | 1577.55 ± 275.82 a | 0.99 ± 0.00 a | 8.81 ± 0.13 a | |
| Ri | 1367.47 ± 57.64 abc | 1364.00 ± 57.11 abcd | 1371.45 ± 57.96 abc | 0.98 ± 0.00 a | 7.96 ± 0.18 c | |
| 900 | Non | 840.11 ± 104.37 d | 837.00 ± 102.31 e | 842.73 ± 105.45 d | 0.75 ± 0.02 b | 4.89 ± 0.19 f |
| Fm | 1401.30 ± 120.36 abc | 1397.67 ± 119.07 abc | 1404.00 ± 121.34 abc | 0.98 ± 0.01 a | 8.22 ± 0.13 bc | |
| Ri | 1534.11 ± 12.08 a | 1531.00 ± 11.27 a | 1537.97 ± 12.8 a | 0.99 ± 0.00 a | 8.64 ± 0.18 ab | |
| 1800 | Non | 960.29 ± 15.72 d | 958.33 ± 14.36 e | 963.50 ± 16.84 d | 0.96 ± 0.01 a | 6.69 ± 0.10 e |
| Fm | 1448.35 ± 25.27 ab | 1444.67 ± 24.91 ab | 1451.35 ± 24.77 ab | 0.98 ± 0.00 a | 7.93 ± 0.10 c | |
| Ri | 1389.45 ± 47.51 abc | 1384.67 ± 46.32 abcd | 1393.93 ± 48.11 abc | 0.98 ± 0.01 a | 7.93 ± 0.20 c | |
| 2700 | Non | 952.45 ± 205.52 d | 946.33 ± 204.29 e | 956.92 ± 206.26 d | 0.76 ± 0.08 b | 5.00 ± 0.84 f |
| Fm | 1297.37 ± 24.72 bc | 1294.33 ± 26.27 bcd | 1301.09 ± 24.94 bc | 0.98 ± 0.00 a | 7.87 ± 0.09 c | |
| Ri | 1198.34 ± 111.74 c | 1193.67 ± 109.04 cd | 1201.57 ± 112.32 c | 0.98 ± 0.01 a | 7.90 ± 0.27 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhao, B.; Zheng, Y.; Li, M.; Zhang, H.; Wang, P.; Chen, S.; Jin, X.; Wu, X. Arbuscular Mycorrhizal Fungi Mitigate Lead Toxicity in Maize by Restructuring Rhizosphere Microbiome and Enhancing Antioxidant Defense Mechanisms. Agronomy 2025, 15, 1310. https://doi.org/10.3390/agronomy15061310
Zhang X, Zhao B, Zheng Y, Li M, Zhang H, Wang P, Chen S, Jin X, Wu X. Arbuscular Mycorrhizal Fungi Mitigate Lead Toxicity in Maize by Restructuring Rhizosphere Microbiome and Enhancing Antioxidant Defense Mechanisms. Agronomy. 2025; 15(6):1310. https://doi.org/10.3390/agronomy15061310
Chicago/Turabian StyleZhang, Xiaoxiang, Bin Zhao, Yan Zheng, Min Li, Huaisheng Zhang, Pingxi Wang, Shilin Chen, Xining Jin, and Xiangyuan Wu. 2025. "Arbuscular Mycorrhizal Fungi Mitigate Lead Toxicity in Maize by Restructuring Rhizosphere Microbiome and Enhancing Antioxidant Defense Mechanisms" Agronomy 15, no. 6: 1310. https://doi.org/10.3390/agronomy15061310
APA StyleZhang, X., Zhao, B., Zheng, Y., Li, M., Zhang, H., Wang, P., Chen, S., Jin, X., & Wu, X. (2025). Arbuscular Mycorrhizal Fungi Mitigate Lead Toxicity in Maize by Restructuring Rhizosphere Microbiome and Enhancing Antioxidant Defense Mechanisms. Agronomy, 15(6), 1310. https://doi.org/10.3390/agronomy15061310