

Identification and Characterization of Alternaria Species Causing Early Blight on Tomato in Kazakhstan

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Morphological Characterization
2.3. DNA Extraction and PCR Amplification
2.4. Pathogenicity Tests
2.5. Phylogenetic Analysis
2.6. Data Analyses
3. Results
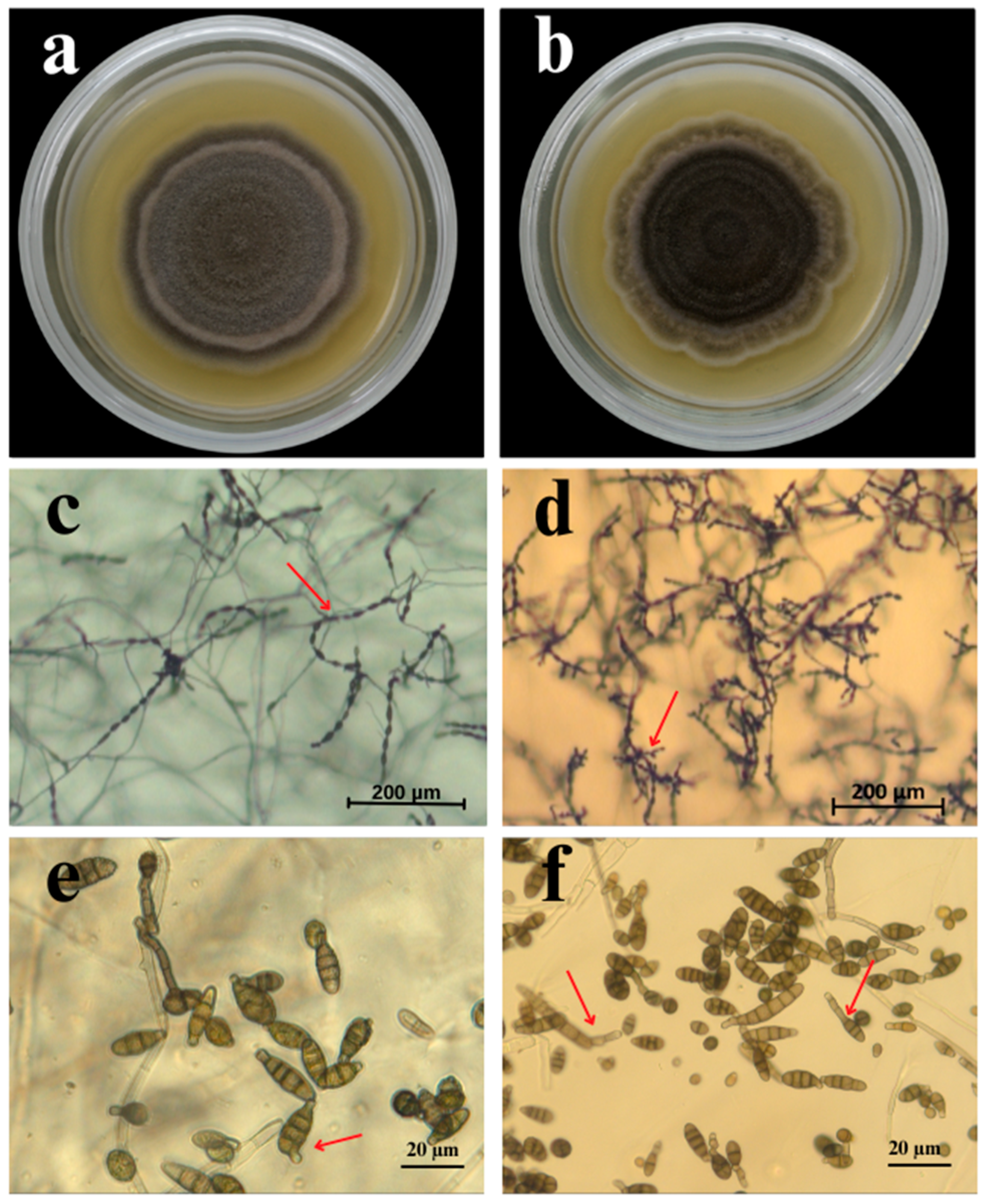
3.1. Morphological Identification of Alternaria Species Associated with Early Blight on Tomato
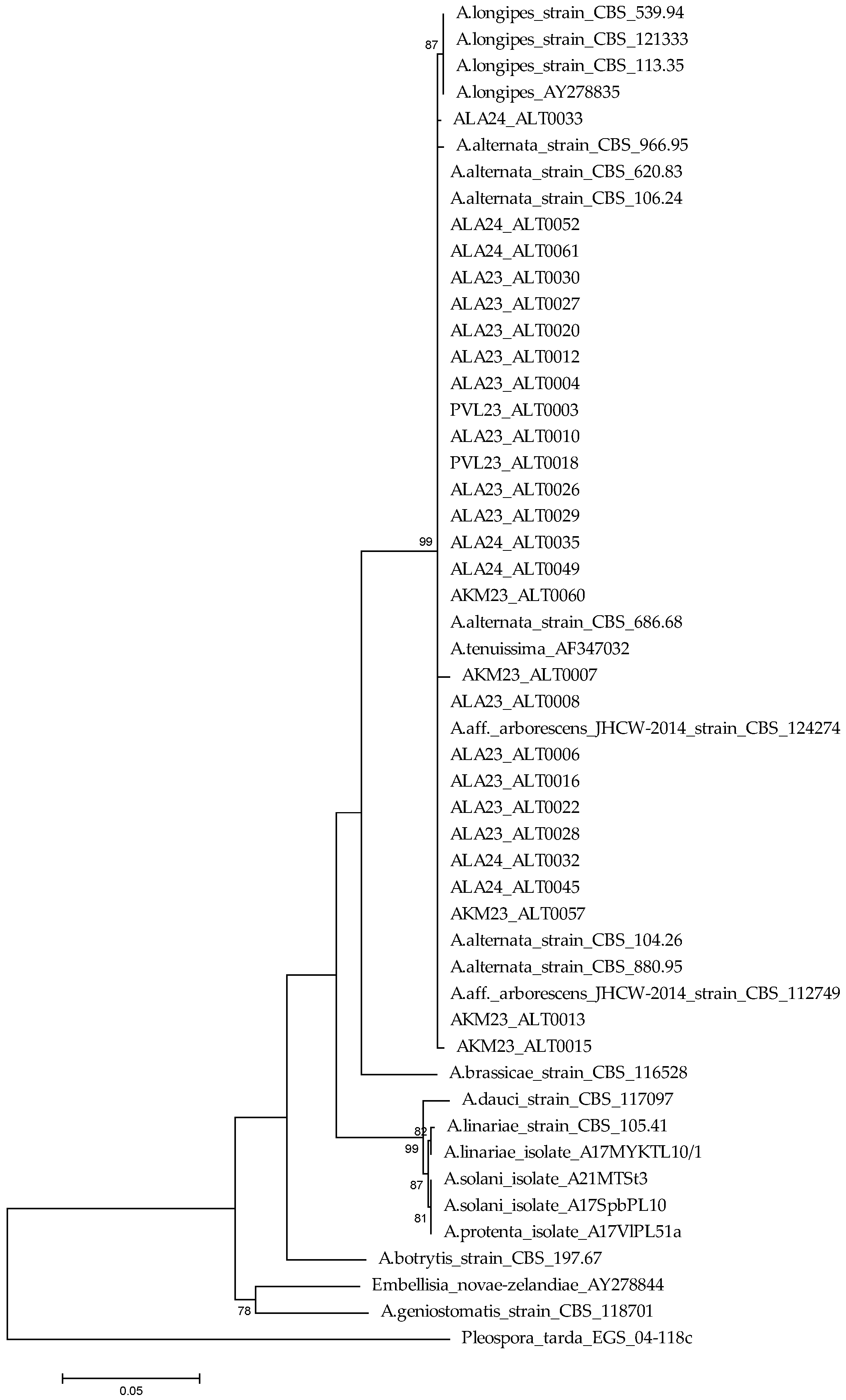
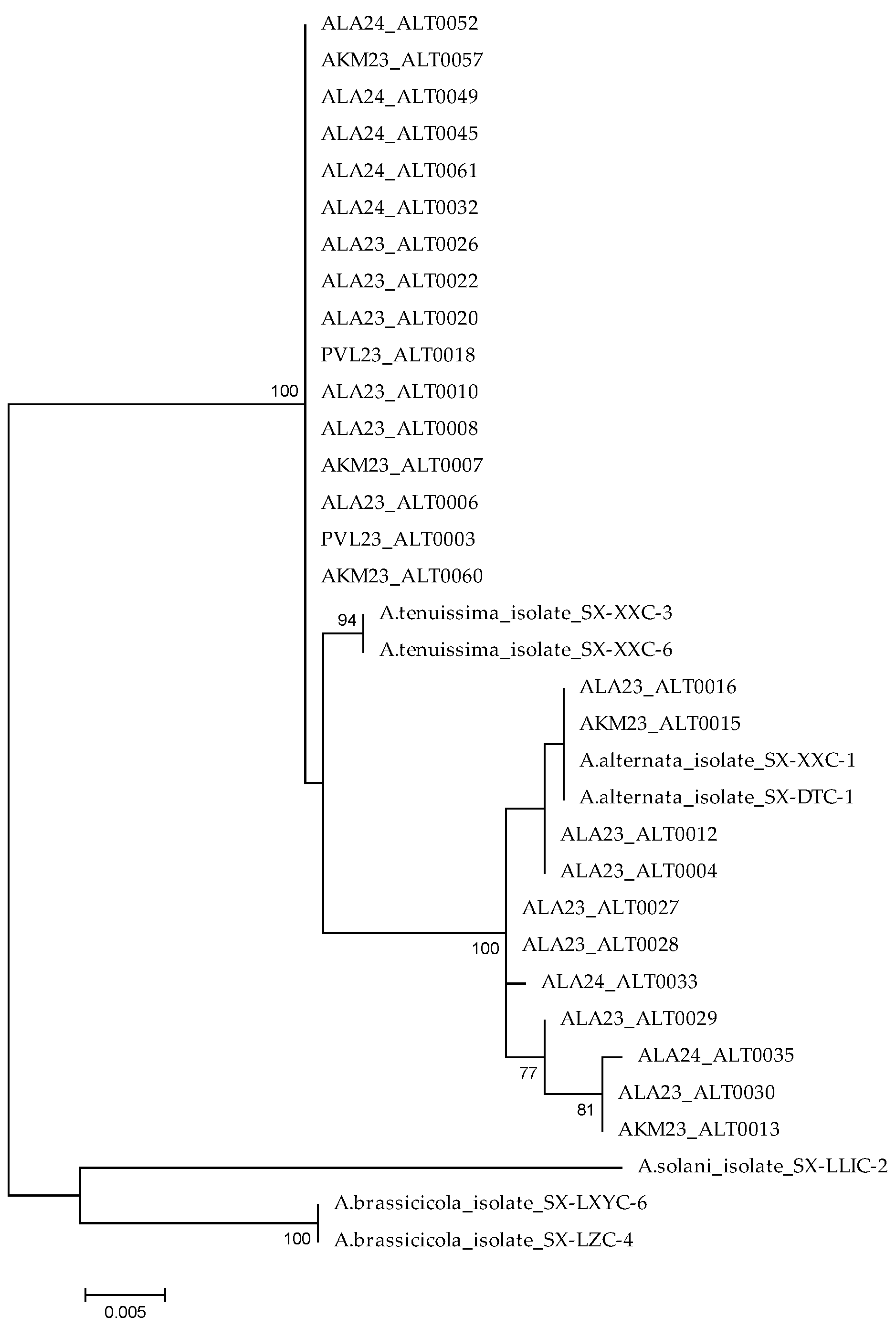
3.2. DNA Sequencing and Phylogenetic Analysis
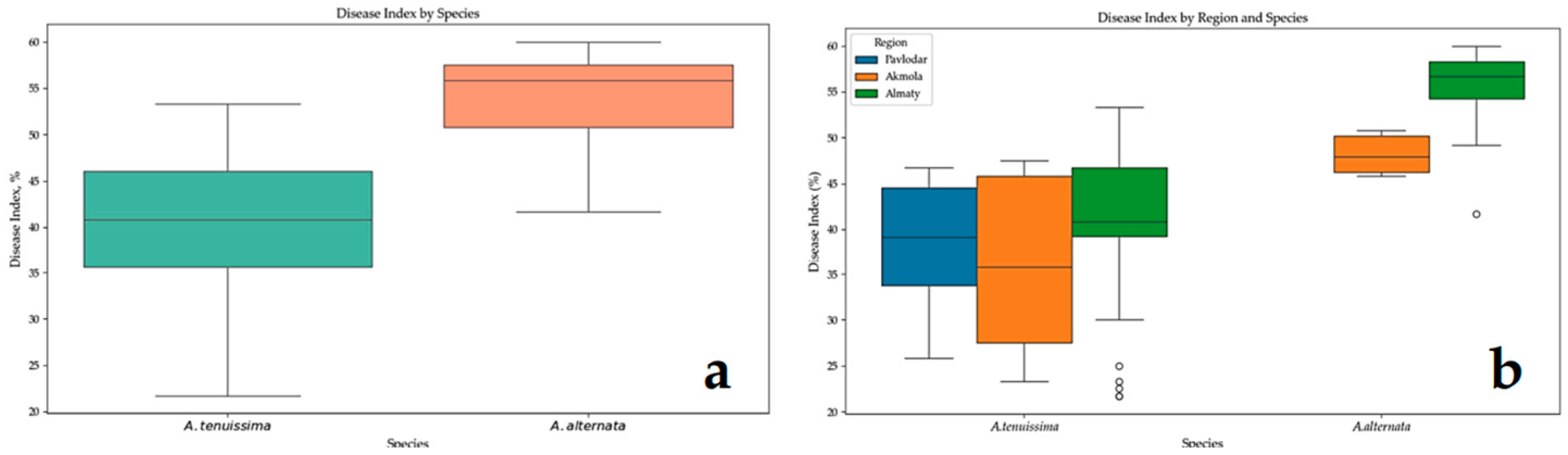
3.3. Pathogenicity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ITS | Internal transcribed spacer |
| PCA | Potato–carrot agar |
| DI | Disease index |
| ANOVA | Analysis of variance |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| H3 | Histone H3 |
| DS | Disease severity |
References
- FAOSTAT Database. Available online: http://faostat.fao.org/site/339/default.aspx (accessed on 20 September 2024).
- Böhm, F.; Edge, R.; Truscott, G. Interactions of dietary carotenoids with activated (singlet) oxygen and free radicals: Potential effects for human health. Mol. Nutr. Food Res. 2012, 56, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Van Breemen, R.B.; Pajkovic, N. Multitargeted therapy of cancer by lycopene. Cancer Lett. 2008, 269, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, P.; Singh, P.; Kethiravan, D.; Ramathani, I.; Ramakrishnan, N. Late Blight in Tomato: Insights into the pathogenesis of the Aggressive Pathogen Phytophthora infestans and Future Research Priorities. Planta 2021, 253, 119. [Google Scholar] [CrossRef]
- Ali, M.; Hussain, S.; Nadeem, A.; Ullah, S.; Yasin, M. Studies on biological management of Fusarium Wilt of tomato. Gesunde Pflanz. 2023, 75, 1475–1483. [Google Scholar] [CrossRef]
- Adhikari, P.; Oh, Y.; Panthee, D.R. Current status of early blight resistance in tomato: An update. Int. J. Mol. Sci. 2017, 18, 2019. [Google Scholar] [CrossRef]
- Pandey, A.; Paudel, R.; Adhikari, T.B.; Panthee, D.R.; Louws, F.J. Septoria Leaf spot of tomatoes: Historical insights, present challenges, and future prospects. Horticulturae 2024, 10, 1299. [Google Scholar] [CrossRef]
- Gannibal, P.B. Monitoring of Alternaria Diseases in Agricultural Crops and Identification of Alternaria Fungi: A Methodological Guide; Levitin, M.M., Ed.; GNU VIZR of the Russian Academy of Agricultural Sciences: St. Petersburg, Russia, 2011; p. 70. [Google Scholar]
- Kokaeva, L.Y.; Belosokhov, A.F.; Doeva, L.Y.; Skolotneva, E.S.; Elansky, S.N. Distribution of Alternaria species on blighted potato and tomato leaves in Russia. J. Plant Dis. Prot. 2017, 125, 205–212. [Google Scholar] [CrossRef]
- Salotti, I.; Giorni, P.; Battilani, P. Biology, ecology, and epidemiology of Alternaria species affecting tomato: Ground information for the development of a predictive model. Front. Plant Sci. 2024, 15, 1430965. [Google Scholar] [CrossRef]
- Habib, W.; Masiello, M.; El Ghorayeb, R.; Gerges, E.; Susca, A.; Meca, G.; Moretti, A. Mycotoxin profile and phylogeny of pathogenic Alternaria species isolated from symptomatic tomato plants in Lebanon. Toxins 2021, 13, 513. [Google Scholar] [CrossRef]
- López, P.; Venema, D.; de Rijk, T.; de Kok, A.; Scholten, J.M.; Mol, H.G.J.; de Nijs, M. Occurrence of Alternaria toxins in Food Products in the Netherlands. Food Control 2016, 60, 196–204. [Google Scholar] [CrossRef]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis, and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Pinto, V.E.F.; Patriarca, A. Alternaria species and Their Associated Mycotoxins. Mycotoxigenic Fungi. Methods Mol. Biol. 2017, 1542, 13–32. [Google Scholar] [CrossRef]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Dongo, A.; Pryor, B.M. Secondary metabolite profiling of Alternaria dauci, A. porri, A. solani, and A. tomatophila. Mycol. Res. 2008, 112, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Maldonado Haro, M.L.; Cabrera, G.; Fernández Pinto, V.; Patriarca, A. Alternaria toxins in tomato products from the Argentinean market. Food Control 2023, 147, 109607. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An overview. J. Food Qual. 2017, 1, 1569748. [Google Scholar]
- Bessadat, N.; Berruyer, R.; Hamon, B.; Bataille-Simoneau, N.; Benichou, S.; Kihal, M.; Henni, D.E.; Simoneau, P. Alternaria species associated with early blight epidemics on tomato and other Solanaceae crops in Northwestern Algeria. Eur. J. Plant Pathol. 2017, 148, 181–197. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria: An Identification Manual; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Woudenberg, J.H.; Truter, M.; Groenewald, J.Z.; Crous, P.W. Large-spored Alternaria pathogens in section Porri disentangled. Stud. Mycol. 2014, 79, 1–47. [Google Scholar] [CrossRef]
- Zhao, L.; Cheng, H.; Liu, H.-F.; Gao, G.-Y.; Zhang, Y.; Li, Z.-N.; Deng, J.-X. Pathogenicity and diversity of large-spored Alternaria associated with three solanaceous vegetables (Solanum tuberosum, S. lycopersicum and S. melongena) in China. Plant Pathol. 2023, 72, 376–391. [Google Scholar] [CrossRef]
- Rodrigues, T.T.M.S.; Berbee, M.L.; Simmons, E.G.; Cardoso, C.R.; Reis, A.; Maffia, L.A.; Mizubuti, E.S.G. First report of Alternaria tomatophila and A. grandis causing early blight on tomato and potato in Brazil. New Dis. Rep. 2010, 22, 28. [Google Scholar] [CrossRef]
- Bessadat, N.; Hamon, B.; Henni, D.E.; Simoneau, P. First report of tomato early blight caused by Alternaria grandis in Algeria. Plant Dis. 2016, 100, 533. [Google Scholar] [CrossRef]
- Alizadeh-Moghaddam, G.; Rezayatmand, Z.; Nasr-Esfahani, M.; Khozaei, M. Bio-genetic analysis of resistance in tomato to early blight disease, Alternaria alternata. Phytochemistry 2020, 179, 112486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y. Alternaria . In Flora Fungorum Sinicorum; Zhao, J.D., Zhang, X.Q., Eds.; Beijing Science Press: Beijing, China, 2003; Volume 16, pp. 1–283. [Google Scholar]
- Andersen, B.; Krøger, E.; Roberts, R.G. Chemical and Morphological Segregation of Alternaria arborescens, A. infectoria, and A. tenuissima Species-Groups. Mycol. Res. 2002, 106, 170–182. [Google Scholar] [CrossRef]
- Oviedo, M.S.; Sturm, M.E.; Reynoso, M.M.; Chulze, S.N.; Ramirez, M.L. Toxigenic profile and AFLP variability of Alternaria alternata and Alternaria infectoria occurring on wheat. Braz. J. Microbiol. 2013, 44, 447–455. [Google Scholar] [CrossRef]
- Kokaeva, L.Y.; Yarmeeva, M.M.; Kokaeva, Z.G.; Chudinova, E.M.; Balabko, P.N.; Elansky, S.N. Phylogenetic study of Alternaria potato and tomato pathogens in Russia. Diversity 2022, 14, 685. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Dugan, F.M.; Pryor, B.M. Characterization of Alternaria isolates from the Infectoria species-group and a new taxon from Arrhenatherum, Pseudoalternaria arrhenatheria sp. nov. Mycol. Progress. 2014, 13, 257–276. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; De Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria Section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef]
- Wang, F.; Saito, S.; Michailides, T.J.; Xiao, C.L. Fungicide resistance in Alternaria alternata from blueberry in California and its impact on control of Alternaria rot. Plant Dis. 2022, 106, 1446–1453. [Google Scholar] [CrossRef]
- Abylaeva, U.A.; Sardar, A.A.; Tursunova, A.K.; Turbekova, S.M.; Abisheva, G.D. Isolation and identification of pathogenic fungi isolated from solanum lycopersicum (tomato) in the conditions of the Almaty region. Microbiol. Virol. 2023, 3, 243–260. [Google Scholar]
- Ismailova, E.T.; Cadanov, A.K.; Shemshura, O.N.; Seitbattalova, A.I.; Daugalieva, C.T.; Kaptagai, R.J. Morphological and molecular genetic characteristics of causative agents of the main fungal diseases of tomatoes growing in Almaty region. News of the National Academy of Sciences of the Republic of Kazakhstan, Series of Biological and Medical; Institute of Plant Biology and Biotechnology: Almaty, Kazakhstan, 2017; Volume 5, pp. 75–81. [Google Scholar]
- Huang, F.; Fu, Y.; Nie, D.; Stewart, J.E.; Peever, T.L.; Li, H. Identification of a novel phylogenetic lineage of Alternaria alternata causing citrus brown spot in China. Fungal Biol. 2015, 119, 320–330. [Google Scholar] [CrossRef] [PubMed]
- White, T.J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. Ph.D. Thesis, Stanford University, Stanford, CA, USA, 1990. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Pryor, B.M.; Michailides, T.J. Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with Alternaria Late Blight of pistachio. Phytopathology 2002, 92, 406–416. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, X.; Liu, S.; Hu, K.; Wu, X. Characterization of Alternaria species associated with black spot of strawberry in Beijing Municipality of China. Can. J. Plant Pathol. 2020, 42, 235–242. [Google Scholar] [CrossRef]
- Allex, C.F.; Shavlik, J.W.; Blattner, F.R. Neural network input representations that produce accurate consensus sequences from DNA fragment assemblies. Bioinformatics 1999, 15, 723–728. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Ramezani, Y.; Taheri, P.; Mamarabadi, M. Identification of Alternaria spp. Associated with tomato early blight in Iran and investigating some of their virulence factors. J. Plant Pathol. 2019, 101, 647–659. [Google Scholar] [CrossRef]
- Elansky, S.N.; Pobedinskaya, M.A.; Kokaeva, L.; Statsyuk, N.; Alexandrova, A. Molecular identification of the species composition of Russian isolates of pathogens causing early blight of potato and tomato. PPO-Spec. Rep. 2012, 15, 151–156. [Google Scholar]
- Shafique, M.S.; Amrao, L.; Saeed, S.; Ahmed, M.Z.; Ghuffar, S.; Anwaar, H.A.; Sheikh, U.A.A.; Khan, M.A.; Qadir, A.; Abdullah, A. Occurrence of leaf spot caused by Alternaria alternata on eggplant (Solanum melongena) in Pakistan. Plant Dis. 2021, 105, 1224. [Google Scholar] [CrossRef]
- Al-Nadabi, H.H.; Maharachchikumbura, S.S.N.; Agrama, H.; Al-Azri, M.; Nasehi, A.; Al-Sadi, A.M. Molecular characterization and pathogenicity of Alternaria species on wheat and date palms in Oman. Eur. J. Plant Pathol. 2018, 152, 577–588. [Google Scholar] [CrossRef]
- Ramjegathesh, R.; Ebenezar, E.G. Morphological and physiological characters of Alternaria alternata causing leaf blight disease of onion. J. Plant Pathol. 2012, 34, 34–44. [Google Scholar]
- Ismail, A.M.; Elshewy, E.S.; El-Ganainy, S.M.; Magistà, D.; Hamouda, A.F.; Alhudaib, K.A.; Ebrahim, W.; Almaghasla, M.I. Mycotoxins from Tomato Pathogenic Alternaria alternata and Their Combined Cytotoxic Effects on Human Cell Lines and Male Albino Rats. J. Fungi 2023, 9, 282. [Google Scholar] [CrossRef]
- Zwickel, T.; Kahl, S.M.; Klaffke, H.; Rychlik, M.; Müller, M.E.H. Spotlight on the Underdogs—An Analysis of Underrepresented Alternaria Mycotoxins Formed Depending on Varying Substrate, Time, and Temperature Conditions. Toxins 2016, 8, 344. [Google Scholar] [CrossRef]
- Nottensteiner, M.; Absmeier, C.; Zellner, M. QoI Fungicide Resistance Mutations in Alternaria solani and Alternaria alternata are Fully Established in Potato Growing Areas in Bavaria and Dual Resistance against SDHI Fungicides is Upcoming. Gesunde Pflanz. 2019, 71, 155–164. [Google Scholar] [CrossRef]
- Masiello, M.; Somma, S.; Susca, A.; Ghionna, V.; Logrieco, A.F.; Franzoni, M.; Ravaglia, S.; Meca, G.; Moretti, A. Molecular identification and mycotoxin production by Alternaria species occurring on durum wheat, showing black point symptoms. Toxins 2020, 12, 275. [Google Scholar] [CrossRef]
- Sun, X.; Wang, C.; Gao, X.; Wu, X.; Fu, Y. Characterization of Alternaria species associated with black spot of strawberry in Dandong, China. Agronomy 2023, 13, 1014. [Google Scholar] [CrossRef]
- Armitage, A.D.; Barbara, D.J.; Harrison, R.J.; Lane, C.R.; Sreenivasaprasad, S.; Woodhall, J.W.; Clarkson, J.P. Discrete lineages within Alternaria alternata species Group: Identification using new highly variable loci and support from morphological characters. Fungal Biol. 2015, 119, 994–1006. [Google Scholar] [CrossRef]
- Shi, X.; Zeng, K.; Wang, X.; Liang, Z.; Wu, X. Characterization of Alternaria species causing leaf spot on Chinese cabbage in Shanxi province of China. J. Plant Pathol. 2021, 103, 283–293. [Google Scholar] [CrossRef]
- Ding, S.; Meinholz, K.; Cleveland, K.; Jordan, S.A.; Gevens, A.J. Diversity and virulence of Alternaria spp. causing potato early blight and brown spot in Wisconsin. Phytopathology 2019, 109, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Gou, Y.N.; Aung, S.L.L.; Htun, A.A.; Huang, C.X.; Deng, J.X. Alternaria species in section Alternaria associated with Iris plants in China. Front. Microbiol. 2022, 13, 1036950. [Google Scholar] [CrossRef] [PubMed]
- Aung, S.L.L.; Wu, L.; Yang, T.; Fu, Y.; Wu, Y.; Luo, C. Morphology and Molecular Characterization of a Fungus from the Alternaria alternata Species Complex Causing Black Spots on Pyrus sinkiangensis. Mycobiology 2020, 48, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.; Gupta, A.K.; Sharma, J.N.; Sharma, M. Characterization of Alternaria blotch disease of apple in Himachal Pradesh, India: Insights on morphology, pathogenicity, and molecular features. Mol. Biol. Rep. 2024, 51, 687. [Google Scholar] [CrossRef]
- Rotondo, F.; Collina, M.; Brunelli, A.; Pryor, B.M. Comparison of Alternaria spp. Collected in Italy from apple with A. mali and other AM-toxin producing strains. Phytopathology 2012, 102, 1130–1142. [Google Scholar] [CrossRef]
- Kang, J.C.; Crous, P.W.; Mchau, G.R.A.; Serdani, M.; Song, S.M. Phylogenetic Analysis of Alternaria spp. Associated with Apple Core Rot and Citrus Black Rot in South Africa. Mycol. Res. 2002, 106, 1151–1162. [Google Scholar] [CrossRef]
- Ma, G.P.; Bao, S.W.; Zhao, J.; Sui, Y.; Wu, X.H. Morphological and molecular characterization of Alternaria species causing leaf blight on watermelon in China. Plant Dis. 2021, 105, 60–70. [Google Scholar] [CrossRef]
- RSE “Kazhydromet”. Archive of Average Meteorological Indicators by Region. Available online: https://kazhydromet.kz (accessed on 5 April 2025).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Number of Alternaria Isolates | |
|---|---|---|
| A. alternata | A. tenuissima | |
| Akmola | 5(2) * | 9(3) |
| Pavlodar | 0(0) | 3(2) |
| Almaty | 23(9) | 21(11) |
| Total | 28(11) | 33(16) |
| Ratio | 46% | 54% |
| Region | Disease Incidence (%) | Disease Index (%) | ||
|---|---|---|---|---|
| A. alternata | A. tenuissima | A. alternata | A. tenuissima | |
| Akmola | 91.7 ± 8.1 * | 84.8 ± 9.2 | 48.2 ± 2.3 | 36.2 ± 9.2 |
| Pavlodar | ND | 75.0 ± 19.9 | ND | 38.2 ± 8.0 |
| Almaty | 86.4 ± 7.3 | 81.5 ± 13.4 | 55.7 ± 3.9 | 40.1 ± 8.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yessimseitova, A.; Abdrakhmanova, A.; Tokbergenova, Z.; Abdullaeva, B.; Muranets, A.; Nurtaza, A.; Kakimzhanova, A. Identification and Characterization of Alternaria Species Causing Early Blight on Tomato in Kazakhstan. Agronomy 2025, 15, 1251. https://doi.org/10.3390/agronomy15051251
Yessimseitova A, Abdrakhmanova A, Tokbergenova Z, Abdullaeva B, Muranets A, Nurtaza A, Kakimzhanova A. Identification and Characterization of Alternaria Species Causing Early Blight on Tomato in Kazakhstan. Agronomy. 2025; 15(5):1251. https://doi.org/10.3390/agronomy15051251
Chicago/Turabian StyleYessimseitova, Assel, Aisha Abdrakhmanova, Zhursinkul Tokbergenova, Barchinay Abdullaeva, Anna Muranets, Aidana Nurtaza, and Almagul Kakimzhanova. 2025. "Identification and Characterization of Alternaria Species Causing Early Blight on Tomato in Kazakhstan" Agronomy 15, no. 5: 1251. https://doi.org/10.3390/agronomy15051251
APA StyleYessimseitova, A., Abdrakhmanova, A., Tokbergenova, Z., Abdullaeva, B., Muranets, A., Nurtaza, A., & Kakimzhanova, A. (2025). Identification and Characterization of Alternaria Species Causing Early Blight on Tomato in Kazakhstan. Agronomy, 15(5), 1251. https://doi.org/10.3390/agronomy15051251