
Melatonin Elicitation Differentially Enhances Flavanone and Its Endogenous Content in Lemon Tissues Through Preharvest and Postharvest Applications




Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material, Experimental Design, and Storage Conditions
2.2. Preparation of Tissues and Juice from Lemon Fruit
2.2.1. Freeze-Dried Samples: Leaf, Flavedo and Albedo
2.2.2. Fresh Juice
2.3. Qualitative and Quantitative Analysis of Phenolic Compounds in the Juice
2.4. Extraction and Quantification of Endogenous Melatonin in the Leaf and Fruit Tissues
2.5. Statistical Analysis
3. Results and Discussion
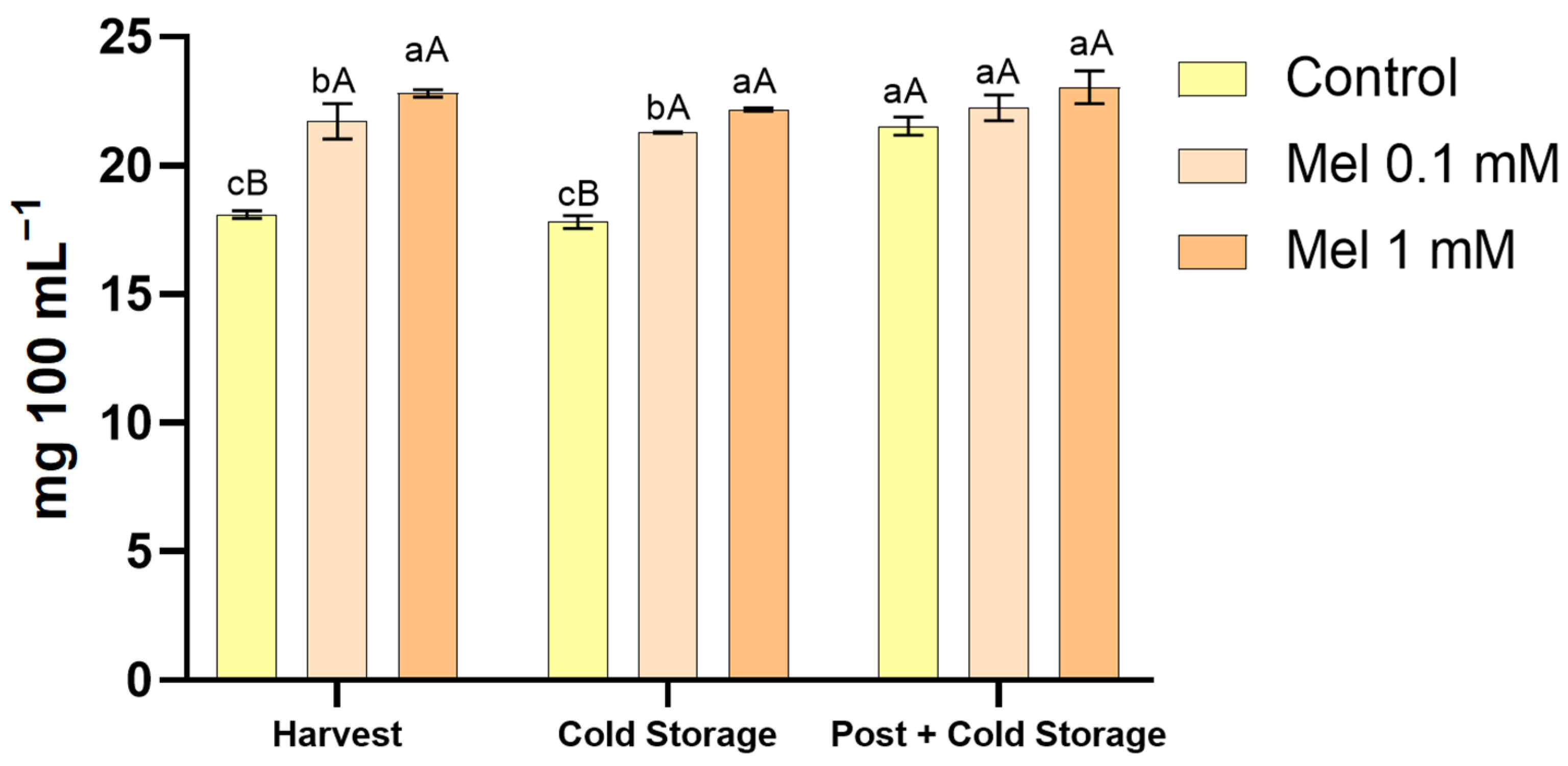
3.1. Effect of Melatonin on Flavanones Concentration in the Juice of Lemon
3.2. Melatonin Distribution in the Leaf and Tissues of Lemon
4. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirwan, D. Global health: Current issues, future trends and foreign policy. Clin. Med. 2009, 9, 247–253. [Google Scholar] [CrossRef]
- Malik, V.S.; Hu, F.B.; Schulze, M.B. Intake of sugar-sweetened beverages and weight gain: A systematic review. Am. J. Clin. Nutr. 2006, 84, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Organización Mundial de la Salud. Evaluación de la capacidad nacional para la prevención y el control de enfermedades no transmisibles. Inf. Encuesta Mund. 2021, 2023, 1–122. [Google Scholar]
- Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Redondo-Flórez, L.; López-Mora, C.; Yáñez-Sepúlveda, R.; Tornero-Aguilera, J.F. New Insights and Potential Therapeutic Interventions in Metabolic Diseases. Int. J. Mol. Sci. 2023, 24, 10672. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Aune, D.; Giovannucci, E.; Boffetta, P.; Fadnes, L.T.; Keum, N.N.; Norat, T.; Greenwood, D.C.; Riboli, E.; Vatten, L.J.; Tonstad, S. Fruit and vegetable intake and the risk of cardiovascular disease, total cancer and all-cause mortality-a systematic review and dose-response meta-analysis of prospective studies. Int. J. Epidemiol. 2017, 46, 1029–1056. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/ (accessed on 16 April 2025).
- Ministerio de Agricultura, Pesca y Alimentación. Available online: https://www.mapa.gob.es/ (accessed on 16 April 2025).
- Home. Available online: https://www.usda.gov/ (accessed on 16 April 2025).
- Tripoli, E.; La Guardia, M.; Giammanco, S.; Di Majo, D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Taher, M.; Shukry, N.A.S.B.; Susanti, D.; Saleh, W.M.N.H.W.; Syukri, Y. Citrus flavonoids in preventing cardiovascular diseases. Plant-Deriv. Bioact. Chem. Mode Action 2020, 1, 495–508. [Google Scholar] [CrossRef]
- Manthey, J.A.; Guthrie, N. Antiproliferative activities of citrus flavonoids against six human cancer cell lines. J. Agric. Food Chem. 2002, 50, 5837–5843. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; García, P.C.; López-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Lin, X.; Wei, Q.; Yang, X.; Zhang, Y.; Chen, J. Melatonin treatment delays postharvest senescence and maintains the organoleptic quality of ‘Newhall’ navel orange (Citrus sinensis (L.) Osbeck) by inhibiting respiration and enhancing antioxidant capacity. Sci. Hortic. 2021, 286, 110236. [Google Scholar] [CrossRef]
- Shah, H.M.S.; Singh, Z.; Hasan, M.U.; Kaur, J.; Afrifa-Yamoah, E.; Woodward, A. Melatonin application suppresses oxidative stress and maintains fruit quality of cold stored ‘Esperanza’ raspberries by regulating antioxidant system. Postharvest Biol. Technol. 2024, 207, 112597. [Google Scholar] [CrossRef]
- Aboryia, M.S.; Lo’Ay, A.A.; Omar, A.S.M. Reduction of chilling injury of “Washington” navel orange fruits by melatonin treatments during cold storage. Folia Hortic. 2021, 33, 343–353. [Google Scholar] [CrossRef]
- Badiche-El Hilali, F.; Valverde, J.M.; García-Pastor, M.E.; Serrano, M.; Castillo, S.; Valero, D. Melatonin Postharvest Treatment in Leafy ‘Fino’ Lemon Maintains Quality and Bioactive Compounds. Foods 2023, 12, 2979. [Google Scholar] [CrossRef]
- Badiche-El Hilali, F.; García-Pastor, M.E.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Melatonin as an Efficient and Eco-Friendly Tool to Increase Yield and to Maintain Quality Attributes during Lemon Storage. Int. J. Mol. Sci. 2024, 25, 10025. [Google Scholar] [CrossRef]
- Carrión-Antolí, A.; Lorente-Mento, J.M.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Effects of melatonin treatment on sweet cherry tree yield and fruit quality. Agronomy 2022, 12, 3. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Serrano, M.; Guillén, F.; Zapata, P.J.; Valero, D. Preharvest or a combination of preharvest and postharvest treatments with methyl jasmonate reduced chilling injury, by maintaining higher unsaturated fatty acids, and increased aril colour and phenolics content in pomegranate. Postharvest Biol. Technol. 2020, 167, 111226. [Google Scholar] [CrossRef]
- Agulló, V.; Domínguez-Perles, R.; Moreno, D.A.; Zafrilla, P.; García-Viguera, C. Alternative sweeteners modify the urinary excretion of flavanones metabolites ingested through a new maqui-berry beverage. Foods 2020, 9, 41. [Google Scholar] [CrossRef]
- Fernández-Pachõn, M.S.; Medina, S.; Herrero-Martín, G.; Cerrillo, I.; Berná, G.; Escudero-Lõpez, B.; Ferreres, F.; Martín, F.; García-Parrilla, M.C.; Gil-Izquierdo, A. Alcoholic fermentation induces melatonin synthesis in orange juice. J. Pineal Res. 2014, 56, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Shahsavar, A. The Effect of Foliar Application of Melatonin on Changes in Secondary Metabolite Contents in Two Citrus Species Under Drought Stress Conditions. Front. Plant Sci. 2021, 12, 692735. [Google Scholar] [CrossRef] [PubMed]
- Rastgoo, N.; Rastegar, S.; Rohani, A. Optimization of melatonin treatment using response surface methodology to enhance postharvest quality of lemon fruit during cold storage. J. Food Meas. Charact. 2024, 18, 2814–2833. [Google Scholar] [CrossRef]
- Liang, D.; Shen, Y.; Ni, Z.; Wang, Q.; Lei, Z.; Xu, N.; Deng, Q.; Lin, L.; Wang, J.; Lv, X.; et al. Exogenous melatonin application delays senescence of kiwifruit leaves by regulating the antioxidant capacity and biosynthesis of flavonoids. Front. Plant Sci. 2018, 9, 360644. [Google Scholar] [CrossRef]
- Khatam, A.S.; Rastegar, S.; Jahromi, A.A.; Khankahdani, H.H.; Bagherian, S.A.A. Biochemical and physiological mechanism induced by melatonin in Mexican lime (Citrus aurantifolia Swingle) plants: Cold and freezing stress. Acta Physiol. Plant. 2023, 45, 98. [Google Scholar] [CrossRef]
- Huynh, T.D.; Kha, C.T.; Nguyen, V.A. Changes in quality parameters and bioactive components of seedless lime fruit (Citrus latifolia) during cold storage. Food Res. 2024, 8, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Recent Advances of Polyphenol Oxidases in Plants. Molecules 2023, 28, 2158. [Google Scholar] [CrossRef]
- Al-Qurashi, A.D.; Awad, M.A. Effect of exogenous melatonin and chitosan treatments on quality and biochemical changes of ‘Balady Banzahir’ limed during shelf life. J. Anim. Plant Sci. 2023, 33, 310–319. [Google Scholar] [CrossRef]
- Hosseini, M.S.; Samsampour, D.; Zahedi, S.M.; Zamanian, K.; Rahman, M.M.; Mostofa, M.G.; Tran, L.S.P. Melatonin alleviates drought impact on growth and essential oil yield of lemon verbena by enhancing antioxidant responses, mineral balance, and abscisic acid content. Physiol. Plant. 2021, 172, 1363–1375. [Google Scholar] [CrossRef] [PubMed]
- Hasnat, H.; Shompa, S.A.; Islam, M.M.; Alam, S.; Richi, F.T.; Emon, N.U.; Ashrafi, S.; Ahmed, N.U.; Chowdhury, M.N.R.; Fatema, N.; et al. Flavonoids: A treasure house of prospective pharmacological potentials. Heliyon 2024, 10, e27533. [Google Scholar] [CrossRef]
- Badiche-El Hilali, F.; Medeiros-Fonseca, B.; Silva, J.; Silvestre-Ferreira, A.C.; Pires, M.J.; Gil da Costa, R.M.; Peixoto, F.; Oliveira, P.A.; Valero, D. The Effect of Lemon Juice (Citrus limon L.) Treated with Melatonin on the Health Status and Treatment of K14HPV16 Mice. Antioxidants 2024, 13, 588. [Google Scholar] [CrossRef]
- Gao, H.; Chen, F.; Wang, S. Hesperidin reduces systolic blood pressure in diabetic patients and has no effect on blood pressure in healthy individuals: A systematic review and meta-analysis. Phytother. Res. 2024, 38, 3706–3719. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Hesperetin protects hippocampal neurons from the neurotoxicity of Aflatoxin B1 in mice. Ecotoxicol. Environ. Saf. 2024, 269, 115782. [Google Scholar] [CrossRef] [PubMed]
- Rastgoo, N.; Rastegar, S.; Rohani, A. Maintaining quality of Lisbon lemon (Citrus limon) in cold storage using natural elicitors. J. Hortic. Postharvest Res. 2024, 7, 99–114. [Google Scholar] [CrossRef]
- Jiki, Z.; Lecour, S.; Nduhirabandi, F. Cardiovascular Benefits of Dietary Melatonin: A Myth or a Reality? Front. Physiol. 2018, 9, 528. [Google Scholar] [CrossRef]
- Tobeiha, M.; Jafari, A.; Fadaei, S.; Mirazimi, S.M.A.; Dashti, F.; Amiri, A.; Khan, H.; Asemi, Z.; Reiter, R.J.; Hamblin, M.R.; et al. Evidence for the Benefits of Melatonin in Cardiovascular Disease. Front. Cardiovasc. Med. 2022, 9, 888319. [Google Scholar] [CrossRef]
- Raygan, F.; Ostadmohammadi, V.; Bahmani, F.; Reiter, R.J.; Asemi, Z. Melatonin administration lowers biomarkers of oxidative stress and cardio-metabolic risk in type 2 diabetic patients with coronary heart disease: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 191–196. [Google Scholar] [CrossRef]
- Ostadmohammadi, V.; Soleimani, A.; Bahmani, F.; Aghadavod, E.; Ramezani, R.; Reiter, R.J.; Mansournia, M.A.; Banikazemi, Z.; Soleimani, M.; Zaroudi, M.; et al. The Effects of Melatonin Supplementation on Parameters of Mental Health, Glycemic Control, Markers of Cardiometabolic Risk, and Oxidative Stress in Diabetic Hemodialysis Patients: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Ren. Nutr. 2020, 30, 242–250. [Google Scholar] [CrossRef]
- Treister-Goltzman, Y.; Peleg, R. Melatonin and the health of menopausal women: A systematic review. J. Pineal Res. 2021, 71, e12743. [Google Scholar] [CrossRef]
- Delpino, F.M.; Figueiredo, L.M.; Nunes, B.P. Effects of melatonin supplementation on diabetes: A systematic review and meta-analysis of randomized clinical trials. Clin. Nutr. 2021, 40, 4595–4605. [Google Scholar] [CrossRef]
- Lin, Y.; Fan, L.; Xia, X.; Wang, Z.; Yin, Y.; Cheng, Y.; Li, Z. Melatonin decreases resistance to postharvest green mold on citrus fruit by scavenging defense-related reactive oxygen species. Postharvest Biol. Technol. 2019, 153, 21–30. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, X.; Yang, Q.; Zhao, Q. Exogenous melatonin delays postharvest fruit senescence and maintains the quality of sweet cherries. Food Chem. 2019, 301, 125311. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of melatonin treatment on the postharvest quality of strawberry fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Yan, R.; Li, S.; Cheng, Y.; Kebbeh, M.; Huan, C.; Zheng, X. Melatonin treatment maintains the quality of cherry tomato by regulating endogenous melatonin and ascorbate-glutathione cycle during room temperature. J. Food Biochem. 2022, 46, e14285. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Liu, L.; Wang, L.; Li, B.; Jin, C.; Lin, X. Melatonin: A master regulator of plant development and stress responses. J. Integr. Plant Biol. 2021, 63, 126–145. [Google Scholar] [CrossRef]
- Li, N.; Zhai, K.; Yin, Q.; Gu, Q.; Zhang, X.; Melencion, M.G.; Chen, Z. Crosstalk between melatonin and reactive oxygen species in fruits and vegetables post-harvest preservation: An update. Front. Nutr. 2023, 10, 1143511. [Google Scholar] [CrossRef]
- Xie, J.; Qin, Z.; Pan, J.; Li, J.; Li, X.; Khoo, H.E.; Dong, X. Melatonin treatment improves postharvest quality and regulates reactive oxygen species metabolism in “Feizixiao” litchi based on principal component analysis. Front. Plant Sci. 2022, 13, 965345. [Google Scholar] [CrossRef]
- Cortés-Montaña, D.; Bernalte-García, M.J.; Palomino-Vasco, M.; Serradilla, M.J.; Velardo-Micharet, B. Effect of preharvest melatonin applications at dusk on quality and bioactive compounds content of early sweet cherries. J. Sci. Food Agric. 2024, 104, 1583–1590. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef]
- Abbas, T.; Ahmad, I.; Nawaz, R.; Nazim, M.; Gatasheh, M.K.; Alamri, A.M.; Muneeb, A. Physiological responses and antioxidant properties of Citrus reticulata under different abiotic stresses mitigated by endogenous melatonin. Sci. Hortic. 2023, 322, 112442. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Compound | Retention Time | Precursor Ion [M-H] (m/z) | Product Ion (m/z) | Fragmentation (V) | CE (V) | Polarity |
|---|---|---|---|---|---|---|
| Melatonin | 0.74 | 233 | 174.10 | 90 | 0 | Positive |
| Melatonin Isomer 1 | 1.88 | 233 | 174.10 | 90 | 0 | Positive |
| Melatonin Isomer 2 | 2.15 | 233 | 174.10 | 90 | 0 | Positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agulló, V.; García-Pastor, M.E.; Valero, D. Melatonin Elicitation Differentially Enhances Flavanone and Its Endogenous Content in Lemon Tissues Through Preharvest and Postharvest Applications. Agronomy 2025, 15, 1233. https://doi.org/10.3390/agronomy15051233
Agulló V, García-Pastor ME, Valero D. Melatonin Elicitation Differentially Enhances Flavanone and Its Endogenous Content in Lemon Tissues Through Preharvest and Postharvest Applications. Agronomy. 2025; 15(5):1233. https://doi.org/10.3390/agronomy15051233
Chicago/Turabian StyleAgulló, Vicente, María Emma García-Pastor, and Daniel Valero. 2025. "Melatonin Elicitation Differentially Enhances Flavanone and Its Endogenous Content in Lemon Tissues Through Preharvest and Postharvest Applications" Agronomy 15, no. 5: 1233. https://doi.org/10.3390/agronomy15051233
APA StyleAgulló, V., García-Pastor, M. E., & Valero, D. (2025). Melatonin Elicitation Differentially Enhances Flavanone and Its Endogenous Content in Lemon Tissues Through Preharvest and Postharvest Applications. Agronomy, 15(5), 1233. https://doi.org/10.3390/agronomy15051233