
Foliar Iron and Zinc Modulate the Qualitative and Nutritional Status of Sideritis cypria with Diverse Rates of Phosphorus in Hydroponic Cultivation

 ,
, 

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Site
2.2. Leaf Photochemistry Features
2.3. Plant Tissue Nutrient Content
2.4. Total Phenols, Total Flavonoids, Antioxidant Capacity
2.5. Stress Indicators
2.6. In Vitro Antibacterial Activity of Ethanolic Extract
2.6.1. Preparation of Ethanolic Extracts and Bacterial Cultures
2.6.2. Disc Diffusion and Microdilution Method
2.7. Statistical Analysis
3. Results
3.1. Plant Growth Parameters
3.2. Leaf Photochemistry Features
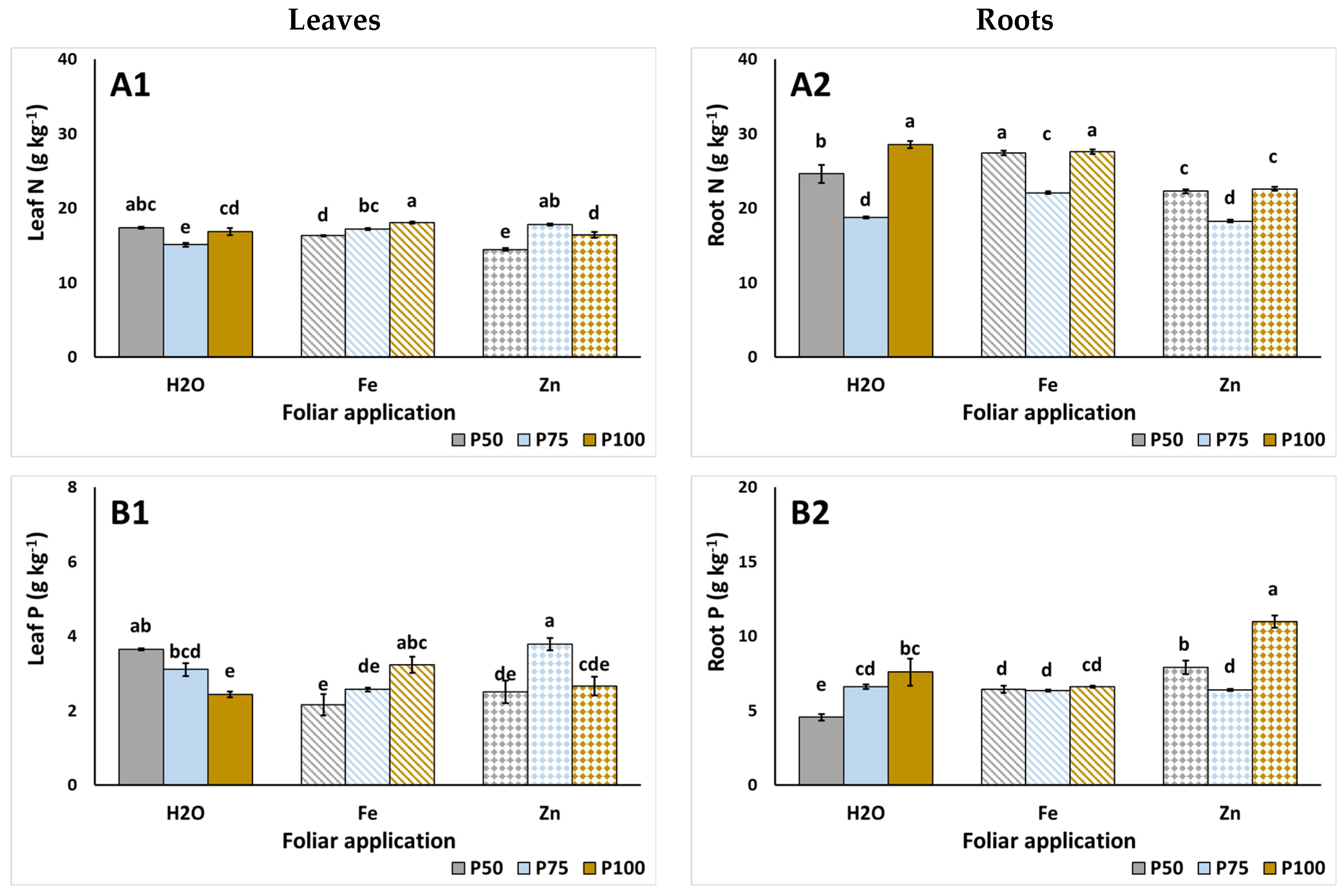
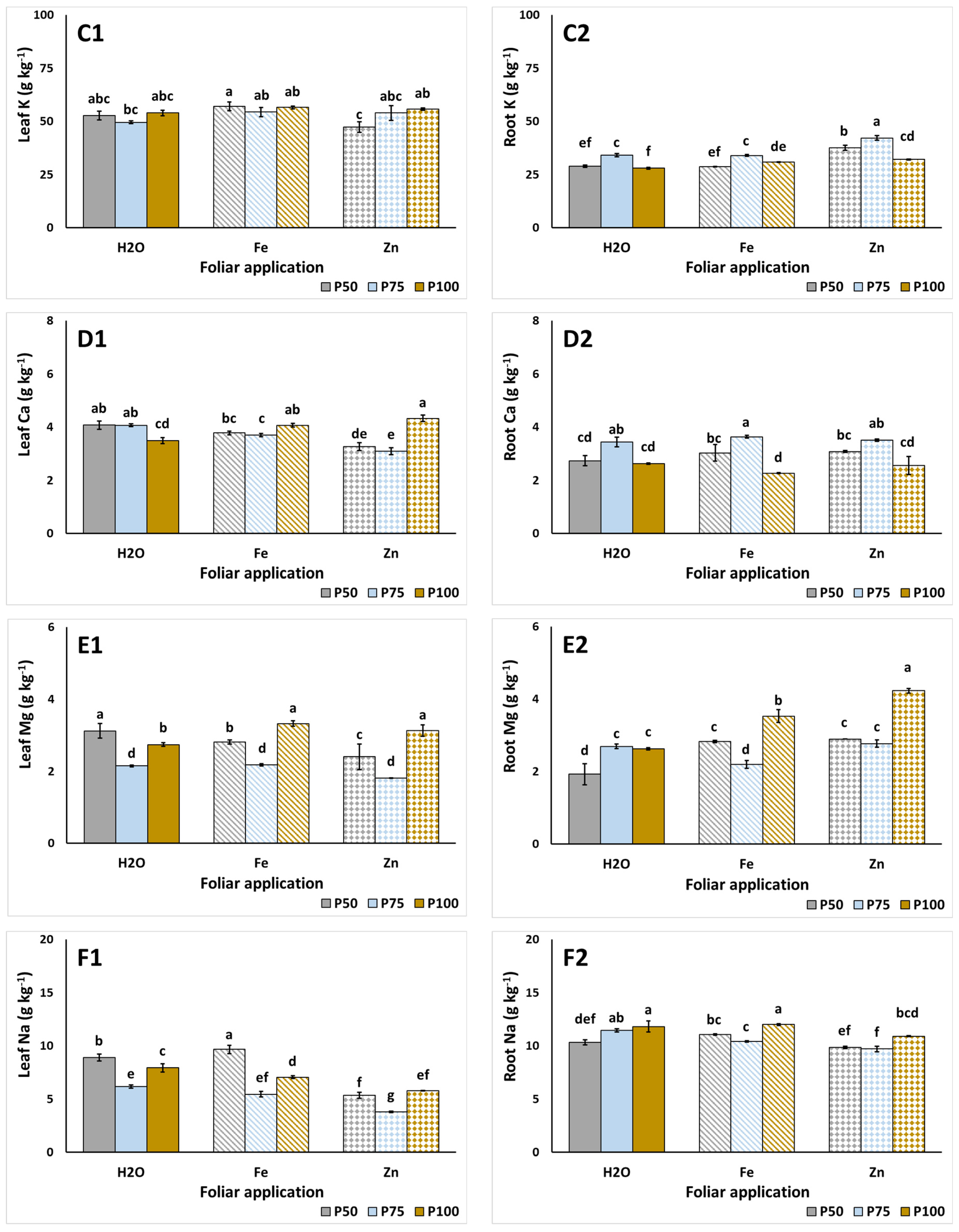
3.3. Plant Nutrient Content
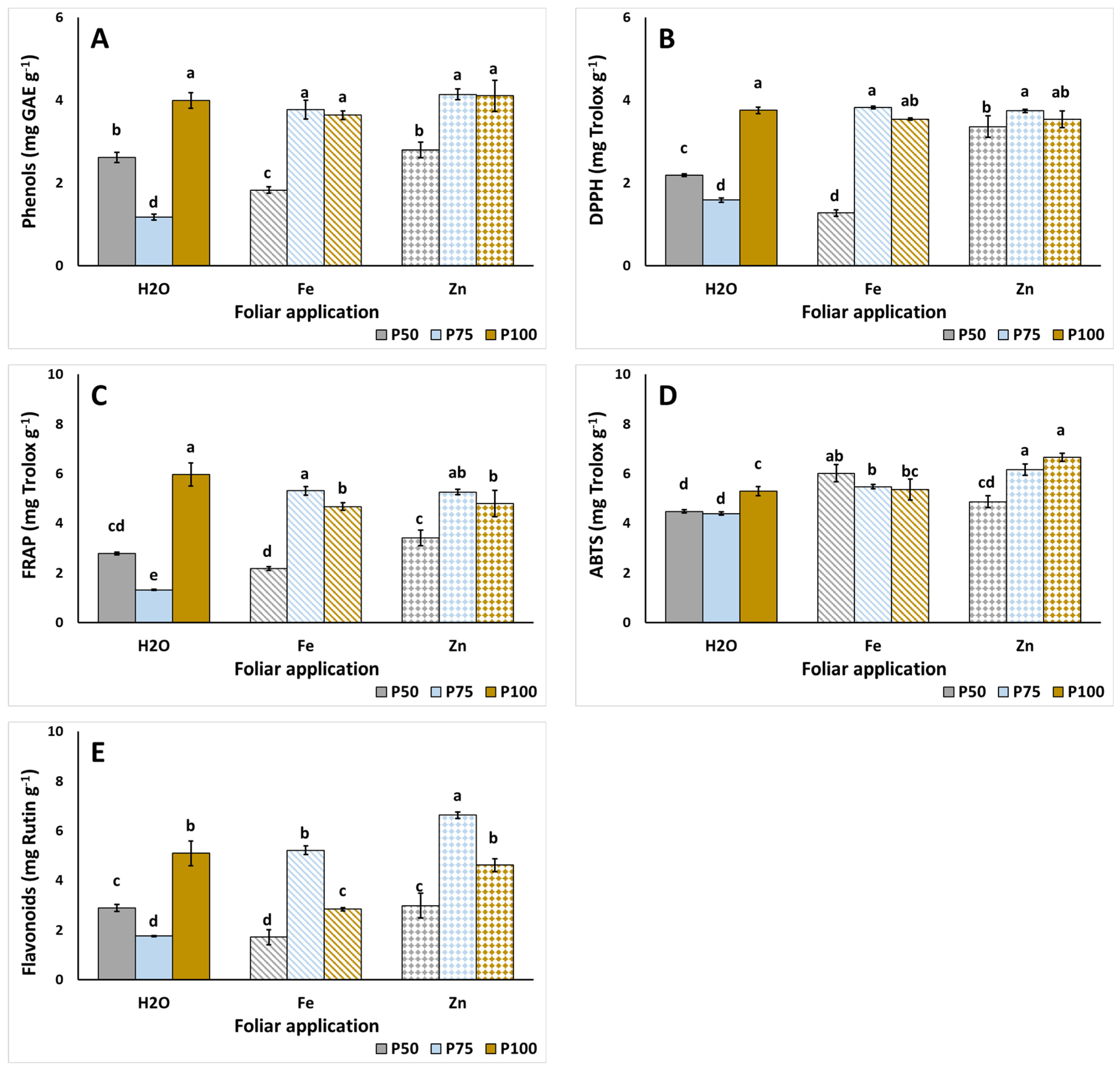
3.4. Total Phenolics Content, Antioxidant Activity, and Flavonoids Content
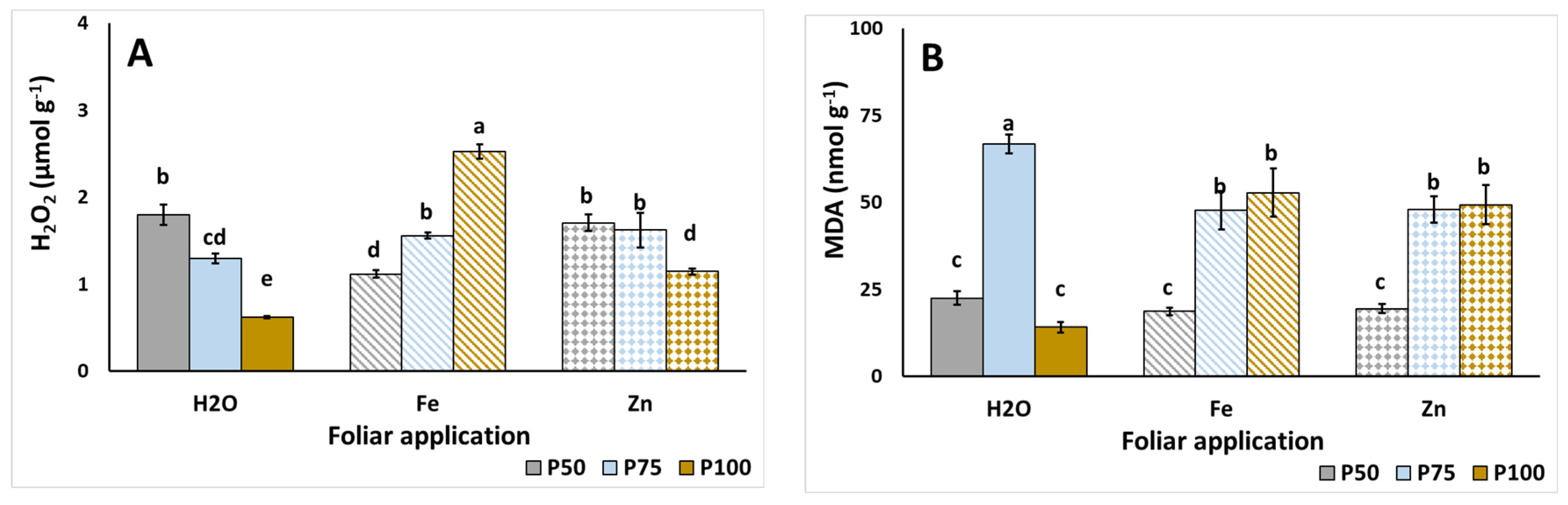
3.5. Stress Indicators
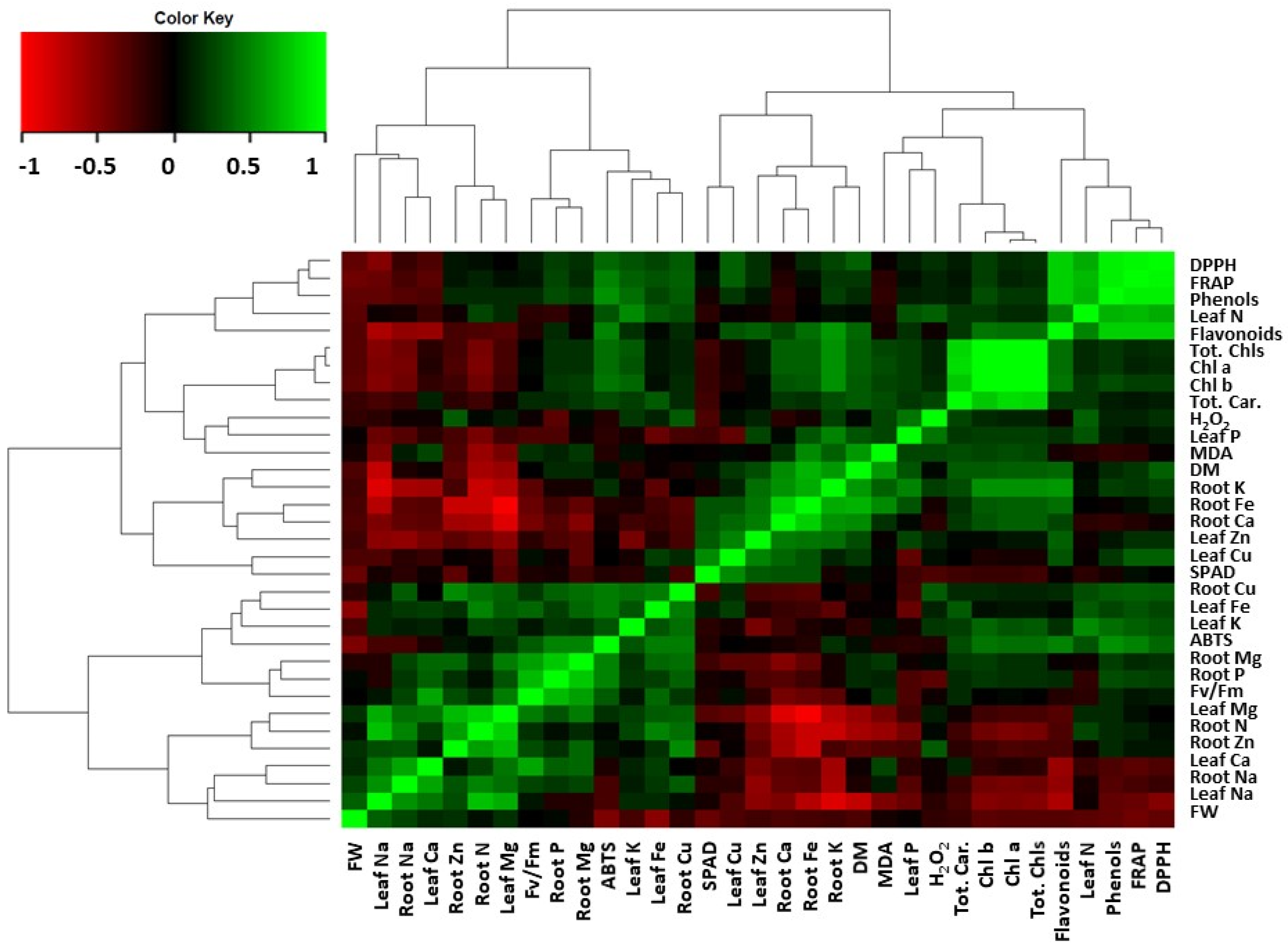
3.6. Correlation Matrix
3.7. Antibacterial Activity of Ethanolic Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aneva, I.; Zhelev, P.; Kozuharova, E.; Danova, K.; Nabavi, S.F.; Behzad, S. Genus Sideritis, section Empedoclia in southeastern Europe and Turkey—Studies in ethnopharmacology and recent progress of biological activities. DARU J. Pharm. Sci. 2019, 27, 407–421. [Google Scholar] [CrossRef]
- González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Sideritis spp.: Uses, chemical composition and pharmacological activities—A review. J. Ethnopharmacol. 2011, 135, 209–225. [Google Scholar] [CrossRef]
- Christodoulou, C.S.; Griffiths, G.H.; Vogiatzakis, I.N. Systematic Conservation Planning in a Mediterranean island context: The example of Cyprus. Glob. Ecol. Conserv. 2021, 32, e01907. [Google Scholar] [CrossRef]
- Yildirim, E.; Turan, M.; Ekinci, M.; Ercisli, S.; Ozturk, H.I.; Aydin, M.; Ilhan, E.; Vicas, S.I.; Iancu, C.V.; Gitea, D.; et al. Composition of Anthocyanins, Specific Sugars, and Organic Acids in Wild Edible Aromatic and Medicinal Vegetables. Horticulturae 2025, 11, 145. [Google Scholar] [CrossRef]
- Hanoğlu, D.Y.; Hanoğlu, A.; Güvenir, M.; Süer, K.; Demirci, B.; Başer, K.H.C.; Yavuz, D.Ö. Chemical composition and antimicrobial activity of the essential oil of Sideritis cypria Post endemic in Northern Cyprus. J. Essent. Oil Res. 2017, 29, 228–232. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Tzortzakis, N. Nitrogen, phosphorus, and potassium requirements to improve Sideritis cypria growth, nutrient and water use efficiency in hydroponic cultivation. Heliyon 2025, 11, e40755. [Google Scholar] [CrossRef]
- Atherton, H.R.; Li, P. Hydroponic Cultivation of Medicinal Plants—Plant Organs and Hydroponic Systems: Techniques and Trends. Horticulturae 2023, 9, 349. [Google Scholar] [CrossRef]
- Nafiu, M.O.; Hamid, A.A.; Muritala, H.F.; Adeyemi, S.B. Preparation, Standardization, and Quality Control of Medicinal Plants in Africa. In Medicinal Spices and Vegetables from Africa; Elsevier: Amsterdam, The Netherlands, 2017; pp. 171–204. [Google Scholar]
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crops Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Peng, L.-C.; Ng, L.-T. Impacts of Nitrogen and Phosphorus Fertilization on Biomass, Polyphenol Contents, and Essential Oil Yield and Composition of Vitex negundo Linn. Agriculture 2022, 12, 859. [Google Scholar] [CrossRef]
- Ceccanti, C.; Brizzi, A.; Landi, M.; Incrocci, L.; Pardossi, A.; Guidi, L. Evaluation of Major Minerals and Trace Elements in Wild and Domesticated Edible Herbs Traditionally Used in the Mediterranean Area. Biol. Trace Elem. Res. 2021, 199, 3553–3561. [Google Scholar] [CrossRef]
- Korkmaz, K.; Akgün, M.; Özcan, M.M.; Özkutlu, F.; Kara, Ş.M. Interaction effects of phosphorus (P) and zinc (Zn) on dry matter, concentration and uptake of P and Zn in chia. J. Plant Nutr. 2021, 44, 755–764. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus Nutrition: Plant Growth in Response to Deficiency and Excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Fujita, M., Oku, H., Nahar, K., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2018; pp. 171–190. [Google Scholar] [CrossRef]
- White, P.J.; Hammond, J.P. Phosphorus nutrition of terrestrial plants. In The Ecophysiology of Plant-Phosphorus Interactions; Springer: Dordrecht, The Netherlands, 2008; pp. 51–81. [Google Scholar]
- Yang, X.; Liu, C.; Liang, C.; Wang, T.; Tian, J. The Phosphorus-Iron Nexus: Decoding the Nutrients Interaction in Soil and Plant. Int. J. Mol. Sci. 2024, 25, 6992. [Google Scholar] [CrossRef]
- Bouain, N.; Shahzad, Z.; Rouached, A.; Khan, G.A.; Berthomieu, P.; Abdelly, C.; Poirier, Y.; Rouached, H. Phosphate and zinc transport and signalling in plants: Toward a better understanding of their homeostasis interaction. J. Exp. Bot. 2014, 65, 5725–5741. [Google Scholar] [CrossRef]
- Xie, X.; Hu, W.; Fan, X.; Chen, H.; Tang, M. Interactions Between Phosphorus, Zinc, and Iron Homeostasis in Nonmycorrhizal and Mycorrhizal Plants. Front. Plant Sci. 2019, 10, 1172. [Google Scholar] [CrossRef]
- Fan, X.; Zhou, X.; Chen, H.; Tang, M.; Xie, X. Cross-Talks Between Macro- and Micronutrient Uptake and Signaling in Plants. Front. Plant Sci. 2021, 12, 663477. [Google Scholar] [CrossRef]
- Rotaru, V.; Sinclair, T.R. Influence of Plant Phosphorus and Iron Concentrations on Growth of Soybean. J. Plant Nutr. 2009, 32, 1513–1526. [Google Scholar] [CrossRef]
- Sharma, S.; Malhotra, H.; Borah, P.; Meena, M.K.; Bindraban, P.; Chandra, S.; Pande, V.; Pandey, R. Foliar application of organic and inorganic iron formulation induces differential detoxification response to improve growth and biofortification in soybean. Plant Physiol. Rep. 2019, 24, 119–128. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Physiological limits to zinc biofortification of edible crops. Front. Plant Sci. 2011, 2, 80. [Google Scholar] [CrossRef]
- Abdoli, S.; Ghassemi-Golezani, K.; Alizadeh-Salteh, S. Responses of ajowan (Trachyspermum ammi L.) to exogenous salicylic acid and iron oxide nanoparticles under salt stress. Environ. Sci. Pollut. Res. 2020, 27, 36939–36953. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. Role of Iron in Plant Growth and Metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef]
- Sattar, A.; Wang, X.; Ul-Allah, S.; Sher, A.; Ijaz, M.; Irfan, M.; Abbas, T.; Hussain, S.; Nawaz, F.; Al-Hashimi, A.; et al. Foliar application of zinc improves morpho-physiological and antioxidant defense mechanisms, and agronomic grain biofortification of wheat (Triticum aestivum L.) under water stress. Saudi J. Biol. Sci. 2022, 29, 1699–1706. [Google Scholar] [CrossRef]
- Nekoukhou, M.; Fallah, S.; Abbasi-Surki, A.; Pokhrel, L.R.; Rostamnejadi, A. Improved efficacy of foliar application of zinc oxide nanoparticles on zinc biofortification, primary productivity and secondary metabolite production in dragonhead. J. Clean. Prod. 2022, 379, 134803. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Neofytou, G.; Chrysargyris, A. Nitrogen Fertilization Coupled with Zinc Foliar Applications Modulate the Production, Quality, and Stress Response of Sideritis cypria Plants Grown Hydroponically Under Excess Copper Concentrations. Plants 2025, 14, 691. [Google Scholar] [CrossRef]
- Marinou, E.; Chrysargyris, A.; Tzortzakis, N. Use of sawdust, coco soil and pumice in hydroponically grown strawberry. Plant Soil Environ. 2013, 59, 452–459. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; De Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Lall, N.; Chrysargyris, A.; Lambrechts, I.; Fibrich, B.; Blom Van Staden, A.; Twilley, D.; de Canha, M.N.; Oosthuizen, C.B.; Bodiba, D.; Tzortzakis, N. Sideritis perfoliata (Subsp. Perfoliata) Nutritive Value and Its Potential Medicinal Properties. Antioxidants 2019, 8, 521. [Google Scholar]
- Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Antioxidant and antibacterial activities, mineral and essential oil composition of spearmint (Mentha spicata L.) affected by the potassium levels. Ind. Crops Prod. 2017, 103, 202–212. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Change 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Yang, M.; He, W.; Li, Y.; Qiu, J.; Liu, F.; Li, R.; Qiu, Y. Effects of low levels of nitrogen or phosphorus provided in hydroponic culture on brown planthopper feeding and survival. Int. J. Pest Manag. 2021, 67, 89–98. [Google Scholar] [CrossRef]
- Dordas, C. Dry matter, nitrogen and phosphorus accumulation, partitioning and remobilization as affected by N and P fertilization and source–sink relations. Eur. J. Agron. 2009, 30, 129–139. [Google Scholar] [CrossRef]
- Fageria, V.D. Nutrient interactions in crop plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Fageria, N.K. Influence of micronutrients on dry matter yield and interaction with other nutrients in annual crops. Pesqui. Agropecuária Bras. 2002, 37, 1765–1772. [Google Scholar] [CrossRef]
- Roosta, H.R.; Mohsenian, Y. Effects of foliar spray of different Fe sources on pepper (Capsicum annum L.) plants in aquaponic system. Sci. Hortic. 2012, 146, 182–191. [Google Scholar] [CrossRef]
- Lambers, H. Phosphorus Acquisition and Utilization in Plants. Annu. Rev. Plant Biol. 2022, 73, 17–42. [Google Scholar] [CrossRef]
- Lefever, K.; Laubscher, C.P.; Ndakidemi, P.A.; Nchu, F. Effects of pH and Phosphorus Concentrations on the Chlorophyll Responses of Salvia chamelaeagnea (Lamiaceae) Grown in Hydroponics. In Chlorophyll; InTech: Houston, TX, USA, 2017. [Google Scholar]
- Siedliska, A.; Baranowski, P.; Pastuszka-Woźniak, J.; Zubik, M.; Krzyszczak, J. Identification of plant leaf phosphorus content at different growth stages based on hyperspectral reflectance. BMC Plant Biol. 2021, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- El Naqma, K.; Elawady, R.; Ramadan, M.; Elsherpiny, M. Improving Soil Phosphorus Availability and Its Influence on Faba Bean performance: Exploring Mineral, Bio and Organic Fertilization with Foliar application of Iron and Zinc. Egypt. J. Soil Sci. 2024, 64, 619–630. [Google Scholar] [CrossRef]
- Mousavi, S.R. Zinc in Crop Production and Interaction with Phosphorus. Aust. J. Basic Appl. Sci. 2011, 5, 1503–1509. [Google Scholar]
- Derakhshani, Z.; Hassani, A.; Sadaghiani, M.H.R.; Hassanpouraghdam, M.B.; Khalifani, B.H.; Dalkani, M. Effect of Zinc Application on Growth and Some Biochemical Characteristics of Costmary (Chrysanthemum balsamita L.). Commun. Soil Sci. Plant Anal. 2011, 42, 2493–2503. [Google Scholar] [CrossRef]
- Asle-Mohammadi, Z.; Kharazmi, M.; Sheikhi, H.; Mohammadkhani, N.; Nicola, S. Foliar Application of Fe, Zn, and Mn as a Practical Strategy to Alleviate the Soil Cu Toxicity and Stimulate the Physiological and Biochemical Properties of Peppermint (Mentha piperita L.). J. Soil Sci. Plant Nutr. 2024, 24, 371–388. [Google Scholar] [CrossRef]
- Ranade-Malvi, U. Interaction of micronutrients with major nutrients with special reference to potassium. Karnataka J. Agric. Sci 2011, 24, 106–109. [Google Scholar]
- Du, M.; Zhang, W.; Gao, J.; Liu, M.; Zhou, Y.; He, D.; Zhao, Y.; Liu, S. Improvement of Root Characteristics Due to Nitrogen, Phosphorus, and Potassium Interactions Increases Rice (Oryza sativa L.) Yield and Nitrogen Use Efficiency. Agronomy 2021, 12, 23. [Google Scholar] [CrossRef]
- Kumar, S.S.; Kumar, S.S.; Mohapatra, T. Interaction Between Macro- and Micro-Nutrients in Plants. Front. Plant Sci. 2021, 12, 753. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Qin, L.; Bao, L.; Li, Y.; Li, X. Critical nutrient thresholds needed to control eutrophication and synergistic interactions between phosphorus and different nitrogen sources. Environ. Sci. Pollut. Res. 2016, 23, 21008–21019. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Wu, W. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63, 34–52. [Google Scholar] [CrossRef]
- Moshtagh, S.; Aminpanah, H. Effects of Phosphorus Rate and Iron Foliar Application on Green Bean (Phaseolus vulgaris L.) Growth and Yield. Agric. Conspec. Sci. 2016, 80, 139–146. [Google Scholar]
- Santos, E.F.; Pongrac, P.; Reis, A.R.; Rabêlo, F.H.S.; Azevedo, R.A.; White, P.J.; Lavres, J. Unravelling homeostasis effects of phosphorus and zinc nutrition by leaf photochemistry and metabolic adjustment in cotton plants. Sci. Rep. 2021, 11, 13746. [Google Scholar] [CrossRef]
- Mardamootoo, T.; Du Preez, C.C.; Barnard, J.H. Phosphorus management issues for crop production: A review. Afr. J. Agric. Res. 2021, 17, 939–952. [Google Scholar]
- Guardiola-Márquez, C.E.; del Martínez-Ballesta, M.C.; García-Sánchez, C.V.; Bojorquez-Rodríguez, E.M.; Jacobo-Velázquez, D.A.; Martínez-Hernández, G.B. Effects of foliar and root application of Zn and Fe bio-nanofertilizers on the glucosinolate, Zn and Fe contents of Pak choi (Brassica rapa Subsp. Chinensis) grown under hydroponic and pot cultivation. Sci. Hortic. 2024, 338, 113654. [Google Scholar] [CrossRef]
- Lytra, K.; Tomou, E.; Chrysargyris, A.; Drouza, C.C.; Skaltsa, H.; Tzortzakis, N. Traditionally Used Sideritis cypria Post: Phytochemistry, Nutritional Content, Bioactive Compounds of Cultivated Populations. Front. Pharmacol. 2020, 11, 650. [Google Scholar] [CrossRef]
- Broschat, T.K.; Moore, K.K. Phytotoxicity of several iron fertilizers and their effects on Fe, Mn, Zn, Cu, and P content of African marigolds and zonal geraniums. HortScience 2004, 39, 595–598. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Tzortzakis, N. Iron and Zinc Foliar Spraying Affected Sideritis cypria Post. Growth, Mineral Content and Antioxidant Properties. Plants 2025, 14, 840. [Google Scholar] [CrossRef]
- Sahin, S.; Yucel, H.; Saglam, N.; Aydın, M.; Cakmak, P.; Gebologlu, N. Foliar Applications Of Ca, Zn, And Urea On Crispy Lettuce In Soilless Culture. Soil-Water J. 2013, 2, 23–30. [Google Scholar]
- Mahmoud, A.W.M.; Ayad, A.A.; Abdel-Aziz, H.S.M.; Williams, L.L.; El-Shazoly, R.M.; Abdel-Wahab, A.; Abdeldaym, E.A. Foliar Application of Different Iron Sources Improves Morpho-Physiological Traits and Nutritional Quality of Broad Bean Grown in Sandy Soil. Plants 2022, 11, 2599. [Google Scholar] [CrossRef] [PubMed]
- Nell, M.; Vötsch, M.; Vierheilig, H.; Steinkellner, S.; Zitterl-Eglseer, K.; Franz, C.; Novak, J. Effect of phosphorus uptake on growth and secondary metabolites of garden sage (Salvia officinalis L.). J. Sci. Food Agric. 2009, 89, 1090–1096. [Google Scholar] [CrossRef]
- Elsafy, M.; Tia, N.A.J.; Sir Elkhatim, K.A.; Othman, M.H.; Hassan, A.B.; Rahmatov, M.; Abdelhalim, T.S. Unveiling the influences of P fertilization on bioactive compounds and antioxidant activity in grains of four sorghum cultivars. PLoS ONE 2024, 19, e0311756. [Google Scholar] [CrossRef]
- Moradbeygi, H.; Jamei, R.; Heidari, R.; Darvishzadeh, R. Investigating the enzymatic and non-enzymatic antioxidant defense by applying iron oxide nanoparticles in Dracocephalum moldavica L. plant under salinity stress. Sci. Hortic. 2020, 272, 109537. [Google Scholar] [CrossRef]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef]
- Lytra, K.; Tomou, E.; Chrysargyris, A.; Christofi, M.; Miltiadous, P.; Tzortzakis, N.; Skaltsa, H. Bio-Guided Investigation of Sideritis cypria Methanol Extract Driven by in Vitro Antioxidant and Cytotoxic Assays. Chem. Biodivers. 2021, 18, e2000966. [Google Scholar] [CrossRef]
- Panayi, T.; Sarigiannis, Y.; Mourelatou, E.; Hapeshis, E.; Papaneophytou, C. Anti-quorum-sensing potential of ethanolic extracts of aromatic plants from the flora of Cyprus. Plants 2022, 11, 2632. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Petrovic, J.D.; Tomou, E.; Kyriakou, K.; Xylia, P.; Kotsoni, A.; Gkretsi, V.; Miltiadous, P.; Skaltsa, H.; Sokovi, M.D.; et al. Phytochemical Profiles and Biological Activities of Plant Extracts from Aromatic Plants Cultivated in Cyprus. Biology 2024, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Çarıkçı, S.; Kılıç, T.; Gören, A.C.; Dirmenci, T.; Alim Toraman, G.Ö.; Topçu, G. Chemical profile of the Anatolian Sideritis species with bioactivity studies. Pharm. Biol. 2023, 61, 1484–1511. [Google Scholar] [CrossRef] [PubMed]
- Szerement, J.; Szatanik-Kloc, A.; Mokrzycki, J.; Mierzwa-Hersztek, M. Agronomic Biofortification with Se, Zn, and Fe: An Effective Strategy to Enhance Crop Nutritional Quality and Stress Defense—A Review; Springer International Publishing: Berlin/Heidelberg, Germany, 2022; Volume 22, ISBN 0123456789. [Google Scholar]
- Akinboye, A.O.; Adeyemo, R.O.; Karzis, J.; Petzer, I.M.; McGaw, L.J. Susceptibility patterns of Escherichia coli and streptococcal isolates from bovine mastitis cases to antibiotics and selected South African plant extracts with known antibacterial activities. S. Afr. J. Bot. 2024, 166, 14–26. [Google Scholar] [CrossRef]
- Lahmar, A.; Bedoui, A.; Mokdad-Bzeouich, I.; Dhaouifi, Z.; Kalboussi, Z.; Cheraif, I.; Ghedira, K.; Chekir-Ghedira, L. Reversal of resistance in bacteria underlies synergistic effect of essential oils with conventional antibiotics. Microb. Pathog. 2017, 106, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Saci, S.; Msela, A.; Saoudi, B.; Sebbane, H.; Trabelsi, L.; Alam, M.; Ernst, B.; Benguerba, Y.; Houali, K. Assessment of antibacterial activity, modes of action, and synergistic effects of Origanum vulgare hydroethanolic extract with antibiotics against avian pathogenic Escherichia coli. Fitoterapia 2024, 177, 106055. [Google Scholar] [CrossRef]
- Farhat, G.; Cheng, L.; Al-Dujaili, E.A.S.; Zubko, M. Antimicrobial Potential of Pomegranate and Lemon Extracts Alone or in Combination with Antibiotics against Pathogens. Int. J. Mol. Sci. 2024, 25, 6943. [Google Scholar] [CrossRef]
- Ciriello, M.; Formisano, L.; Kyriacou, M.; Soteriou, G.A.; Graziani, G.; De Pascale, S.; Rouphael, Y. Zinc biofortification of hydroponically grown basil: Stress physiological responses and impact on antioxidant secondary metabolites of genotypic variants. Front. Plant Sci. 2022, 13, 1049004. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foliar Application | P Levels | FW | DM |
|---|---|---|---|
| H2O | P50 | 40.64 ± 4.95 | 17.00 ± 0.30 cd |
| P75 | 39.10 ± 5.68 | 19.79 ± 0.65 a | |
| P100 | 42.44 ± 3.70 | 18.17 ± 0.22 bc | |
| Fe | P50 | 33.57 ± 1.66 | 16.37 ± 0.68 d |
| P75 | 29.47 ± 3.86 | 19.63 ± 0.17 a | |
| P100 | 34.87 ± 3.98 | 18.69 ± 0.42 ab | |
| Zn | P50 | 40.87 ± 2.82 | 18.74 ± 0.15 ab |
| P75 | 31.65 ± 2.89 | 20.06 ± 0.42 a | |
| P100 | 35.93 ± 2.56 | 19.36 ± 0.12 ab |
| Foliar Application | P Levels | SPAD | Fv/Fm | Chl a | Chl b | Total Chl | Total Car |
|---|---|---|---|---|---|---|---|
| H2O | P50 | 45.15 ± 2.06 ab | 0.82 ± 0.01 bc | 0.60 ± 0.05 ab | 0.23 ± 0.03 abc | 0.83 ± 0.09 ab | 0.123 ± 0.009 a |
| P75 | 48.23 ± 0.77 ab | 0.81 ± 0.01 bc | 0.56 ± 0.04 ab | 0.22 ± 0.02 bc | 0.78 ± 0.06 abc | 0.105 ± 0.009 a | |
| P100 | 49.80 ± 3.24 a | 0.80 ± 0.00 cd | 0.41 ± 0.02 c | 0.16 ± 0.01 c | 0.57 ± 0.03 c | 0.073 ± 0.007 b | |
| Fe | P50 | 46.48 ± 2.02 ab | 0.81 ± 0.00 bc | 0.60 ± 0.03 ab | 0.23 ± 0.01 abc | 0.84 ± 0.04 ab | 0.113 ± 0.009 a |
| P75 | 52.16 ± 2.53 a | 0.83 ± 0.01 bc | 0.63 ± 0.03 ab | 0.25 ± 0.01 abc | 0.87 ± 0.05 ab | 0.123 ± 0.009 a | |
| P100 | 42.08 ± 2.15 b | 0.83 ± 0.02 b | 0.55 ± 0.08 bc | 0.22 ± 0.04 abc | 0.77 ± 0.12 bc | 0.117 ± 0.019 a | |
| Zn | P50 | 45.73 ± 1.47 ab | 0.83 ± 0.00 b | 0.65 ± 0.07 ab | 0.27 ± 0.04 ab | 0.93 ± 0.11 ab | 0.113 ± 0.007 a |
| P75 | 47.08 ± 2.68 ab | 0.78 ± 0.01 d | 0.73 ± 0.05 a | 0.31 ± 0.03 a | 1.03 ± 0.08 a | 0.123 ± 0.005 a | |
| P100 | 46.17 ± 1.46 ab | 0.90 ± 0.01 a | 0.69 ± 0.04 ab | 0.29 ± 0.03 ab | 0.98 ± 0.06 ab | 0.130 ± 0.000 a |
| Foliar Application | H2O | Fe | Zn | Ampicillin | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacterium | P Levels | P50 | P75 | P100 | P50 | P75 | P100 | P50 | P75 | P100 | |
| E. coli | DIZ | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 7.50 ± 0.50 a |
| MIC | 0.50 ± 0.00 a | 0.50 ± 0.00 a | 0.25 ± 0.00 b | 0.50 ± 0.00 a | 0.50 ± 0.00 a | 0.25 ± 0.00 b | 0.25 ± 0.00 b | 0.25 ± 0.00 b | 0.12 ± 0.00 c | 0.0500 ± 0.0000 d | |
| IC50 | 8.81 ± 0.41 abc | 9.15 ± 1.40 abc | 10.68 ± 1.01 ab | 3.96 ± 0.44 de | 13.14 ± 0.87 a | 10.08 ± 1.71 abc | 5.45 ± 0.00 cd | 7.69 ± 2.54 bcd | 7.71 ± 2.73 bcd | 0.1108 ± 0.0115 e | |
| S. enterica | DIZ | 7.50 ± 0.50 | 7.50 ± 0.50 | 7.50 ± 0.50 | 7.50 ± 0.50 | 7.50 ± 0.50 | 9.00 ± 0.00 | 7.50 ± 0.50 | 7.50 ± 0.50 | 8.00 ± 0.00 | 8.50 ± 0.50 |
| MIC | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 0.50 ± 0.00 b | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 0.25 ± 0.00 c | 0.50 ± 0.00 b | 0.50 ± 0.00 b | 0.12 ± 0.00 d | 0.0062 ± 0.0000 e | |
| IC50 | 4.46 ± 0.63 abc | 3.10 ± 0.09 cd | 5.29 ± 0.38 a | 3.74 ± 0.15 bc | 3.34 ± 0.05 bc | 3.68 ± 0.16 bc | 3.31 ± 0.17 bc | 1.84 ± 0.06d | 4.64 ± 1.11 ab | 0.0121 ± 0.0004 e | |
| S. aureus | DIZ | 6.00 ± 0.00 b | 9.50 ± 1.50 a | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 9.50 ± 0.50 a |
| MIC | 0.25 ± 0.00 a | 0.12 ± 0.00 b | 0.12 ± 0.00 b | 0.25 ± 0.00 a | 0.25 ± 0.00 a | 0.12 ± 0.00 b | 0.12 ± 0.00 b | 0.25 ± 0.00 a | 0.06 ± 0.00 c | 0.0016 ± 0.0000 d | |
| IC50 | 3.26 ± 0.20 a | 2.96 ± 0.38 a | 2.87 ± 0.04 a | 3.18 ± 0.03 a | 4.09 ± 0.43 ab | 2.89 ± 0.44 b | 2.97 ± 0.40 b | 3.14 ± 0.39 b | 4.63 ± 1.00 a | 0.0034 ± 0.0003 c | |
| L. monocytogenes | DIZ | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 6.00 ± 0.00 b | 7.00 ± 0.00 a |
| MIC | 0.25 ± 0.00 a | 0.12 ± 0.00 b | 0.12 ± 0.00 b | 0.25 ± 0.00 a | 0.12 ± 0.00 b | 0.12 ± 0.00 b | 0.25 ± 0.00 a | 0.25 ± 0.00 a | 0.12 ± 0.00 b | 0.0008 ± 0.0000 c | |
| IC50 | 0.33 ± 0.03 bc | 0.24 ± 0.03 bc | 0.43 ± 0.03 bc | 0.35 ± 0.02 bc | 0.39 ± 0.00 bc | 0.52 ± 0.02b | 0.34 ± 0.00 bc | 0.39 ± 0.09 bc | 0.96 ± 0.18 a | 0.0019 ± 0.0000 c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neofytou, G.; Chrysargyris, A.; Xylia, P.; Botsaris, G.; Tzortzakis, N. Foliar Iron and Zinc Modulate the Qualitative and Nutritional Status of Sideritis cypria with Diverse Rates of Phosphorus in Hydroponic Cultivation. Agronomy 2025, 15, 1178. https://doi.org/10.3390/agronomy15051178
Neofytou G, Chrysargyris A, Xylia P, Botsaris G, Tzortzakis N. Foliar Iron and Zinc Modulate the Qualitative and Nutritional Status of Sideritis cypria with Diverse Rates of Phosphorus in Hydroponic Cultivation. Agronomy. 2025; 15(5):1178. https://doi.org/10.3390/agronomy15051178
Chicago/Turabian StyleNeofytou, Giannis, Antonios Chrysargyris, Panayiota Xylia, George Botsaris, and Nikolaos Tzortzakis. 2025. "Foliar Iron and Zinc Modulate the Qualitative and Nutritional Status of Sideritis cypria with Diverse Rates of Phosphorus in Hydroponic Cultivation" Agronomy 15, no. 5: 1178. https://doi.org/10.3390/agronomy15051178
APA StyleNeofytou, G., Chrysargyris, A., Xylia, P., Botsaris, G., & Tzortzakis, N. (2025). Foliar Iron and Zinc Modulate the Qualitative and Nutritional Status of Sideritis cypria with Diverse Rates of Phosphorus in Hydroponic Cultivation. Agronomy, 15(5), 1178. https://doi.org/10.3390/agronomy15051178