
Gene Silencing of Sarco/Endoplasmic Reticulum Ca2+-ATPase and NADPH–Cytochrome P450 Reductase as a Novel Approach to Leptinotarsa decemlineata Management

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
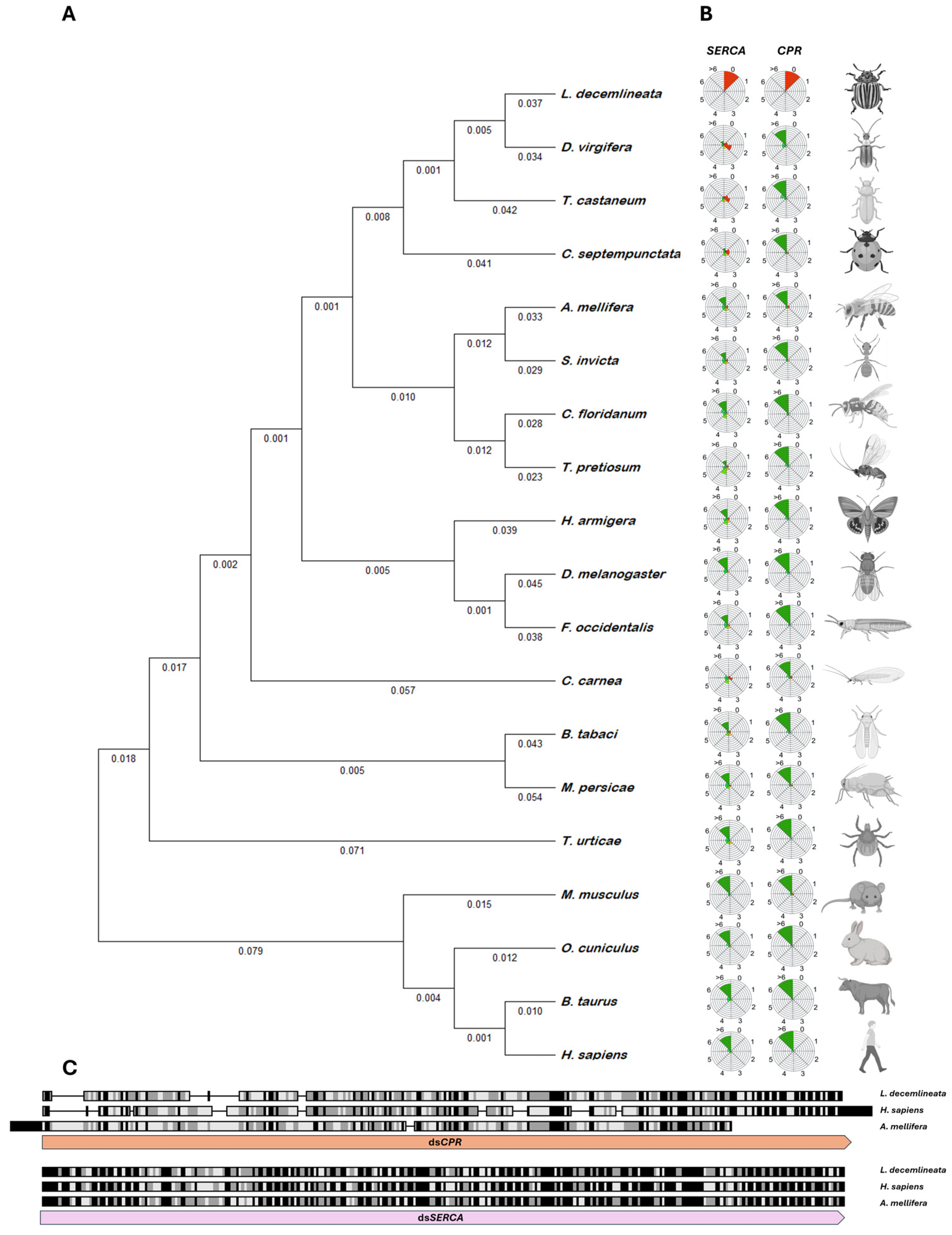
2.1. Selection of Target Genes
2.2. In Silico Analysis of RNAi Effectiveness and Off-Target Effects
2.3. Synthesis and Cloning of SERCA and CPR Fragments
2.4. In Vitro dsRNA Synthesis
2.5. Insect Growth Conditions
2.6. RNA Extraction
2.7. Quantitative Real-Time PCR (qPCR)
2.8. Statistical Analysis
3. Results
3.1. In Vitro Synthesis of dsRNA Targeting SERCA and CPR Sequences
3.2. Effects of Larval Feeding with dsRNA Targeting SERCA and CPR Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | d0 | d1 | d2 | d3 | d4 | d5 | d6 | d7 |
| Control | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| dsSERCA | 100 | 92 | 86 | 73 | 23 | — | — | — |
| dsCPR | 100 | 96 | 92 | 85 | 84 | 81 | 80 | 72 |
| Treatment | d3 | d5 | d7 |
| Control | 13.57 | 17.09 | 27.39 |
| dsCPR | 7.87 | 11.58 | 11.28 |
| dsSERCA | 7.29 | 8.47 | — |
| Treatment | Standard error | ||
| d3 | d5 | d7 | |
| Control | 0.89 | 0.55 | 1.38 |
| dsCPR | 0.08 | 0.46 | 0.24 |
| dsSERCA | 0.26 | 0.22 | — |
| Treatment | Post-hoc Tukey | ||
| d3 | d5 | d7 | |
| Control | A | A | A |
| dsCPR | B | B | B |
| dsSERCA | B | C | — |
| Treatment | d3 | d5 | d7 |
| Control | 1.00 | 1.00 | 1.00 |
| dsCPR | 0.95 | 0.83 | 0.52 |
| dsSERCA | 0.44 | 0.72 | — |
| Treatment | Standard error | ||
| d3 | d5 | d7 | |
| Control | 0.00 | 0.00 | 0.00 |
| dsCPR | 0.03 | 0.09 | 0.13 |
| dsSERCA | 0.07 | 0.10 | — |
| Treatment | Post-hoc Tukey | ||
| d3 | d5 | d7 | |
| Control | A | A | A |
| dsCPR | A | AB | B |
| dsSERCA | B | B | — |
References
- FAOSTAT. Food and Agriculture Organization of the United Nations: Crops and Livestock Products. Available online: https://www.fao.org/faostat/es/#data/QCL (accessed on 25 February 2025).
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Skryabin, K. Do Russia and Eastern Europe need GM plants? New Biotechnol. 2010, 27, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Ferro, D.N.; Logan, J.A.; Voss, R.H.; Elkinton, J.S. Colorado Potato Beetle (Coleoptera: Chrysomelidae) temperature-dependent growth and feeding rates. Environ. Entomol. 1985, 14, 343–348. [Google Scholar] [CrossRef]
- Weber, D. Colorado Beetle: Pest on the move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Alyokhin, A.; Benkovskayab, G.; Udalovc, M. Colorado Potato Beetle. In Insect Pests of Potato, Global Perspectives on Biology and Management, 2nd ed.; Alyokhin, A., Rondon, S.I., Gao, Y., Eds.; Elsevier: London, UK, 2022; Volume 1, p. 503. [Google Scholar]
- Alvarez, J.M.; Srinivasan, R.; Cervantes, F.A. Occurrence of the Carabid Beetle, Pterostichus melanarius (Illiger), in potato ecosystems of Idaho and its predatory potential on the Colorado Potato Beetle and aphids. Am. J. Potato Res. 2013, 90, 83–92. [Google Scholar] [CrossRef]
- Sablon, L.; Haubruge, E.; Verheggen, F.J. Consumption of immature stages of Colorado Potato Beetle by Chrysoperla carnea (Neuroptera: Chrysopidae) larvae in the laboratory. Am. J. Potato Res. 2013, 90, 51–57. [Google Scholar] [CrossRef]
- Speese, J.; Sterrett, S.B. Crop rotation reduces the cost of Colorado Potato Beetle control in potatoes. HortTechnology 1998, 8, 229–234. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Delayed efficacy of Beauveria bassiana foliar spray applications against Colorado Potato Beetle: Impacts of number and timing of applications on larval and next-generation adult populations. Biol. Control 2015, 83, 51–67. [Google Scholar] [CrossRef]
- Chen, Y.H.; Cohen, Z.P.; Bueno, E.M.; Christensen, B.M.; Schoville, S.D. Rapid evolution of insecticide resistance in the Colorado Potato Beetle, Leptinotarsa decemlineata. Curr. Opin. Insect Sci. 2023, 55, 101000. [Google Scholar] [CrossRef]
- Pélissié, B.; Chen, Y.H.; Cohen, Z.P.; Crossley, M.S.; Hawthorne, D.J.; Izzo, V.; Schoville, S.D. Genome resequencing reveals rapid, repeated evolution in the Colorado Potato Beetle. Mol. Biol. Evol. 2022, 39, msac016. [Google Scholar] [CrossRef] [PubMed]
- Gaddelapati, S.C.; Kalsi, M.; Roy, A.; Palli, S.R. Cap ‘n’ collar C regulates genes responsible for imidacloprid resistance in the Colorado Potato Beetle, Leptinotarsa decemlineata. Insect Biochem. Mol. Biol. 2018, 99, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.-J.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado Potato Beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef]
- Zhu, F.; Moural, T.W.; Nelson, D.R.; Palli, S.R. A specialist herbivore pest adaptation to xenobiotics through up-regulation of multiple Cytochrome P450s. Sci. Rep. 2016, 6, 20421. [Google Scholar] [CrossRef]
- APRD. Arthropod Pesticide Resistance Database: Leptinotarsa decemlineata. Available online: https://www.pesticideresistance.org (accessed on 25 February 2025).
- FAO. Food and Agriculture Organization of the United Nations Statistical Databases: Pesticides Indicators. Available online: https://www.fao.org/faostat/en/#data/RP (accessed on 4 February 2025).
- Barathi, S.; Sabapathi, N.; Kandasamy, S.; Lee, J. Present status of insecticide impacts and eco-friendly approaches for remediation-a review. Environ. Res. 2024, 240, 117432. [Google Scholar] [CrossRef] [PubMed]
- Eddleston, M. Poisoning by pesticides. Medicine 2020, 48, 214–217. [Google Scholar] [CrossRef]
- Christiaens, O.; Whyard, S.; Vélez, A.M.; Smagghe, G. Double-stranded RNA technology to control insect pests: Current status and challenges. Front. Plant Sci. 2020, 11, 451. [Google Scholar] [CrossRef]
- Svoboda, P. Key mechanistic principles and considerations concerning RNA interference. Front. Plant Sci. 2020, 11, 1237. [Google Scholar] [CrossRef]
- Gordon, K.H.J.; Waterhouse, P.M. RNAi for insect-proof plants. Nat. Biotechnol. 2007, 25, 1231–1232. [Google Scholar] [CrossRef]
- Eakteiman, G.; Moses-Koch, R.; Moshitzky, P.; Mestre-Rincon, N.; Vassão, D.G.; Luck, K.; Sertchook, R.; Malka, O.; Morin, S. Targeting detoxification genes by phloem-mediated RNAi: A new approach for controlling phloem-feeding insect pests. Insect Biochem. Mol. Biol. 2018, 100, 10–21. [Google Scholar] [CrossRef]
- Xu, H.-X.; Qian, L.-X.; Wang, X.-W.; Shao, R.-X.; Hong, Y.; Liu, S.-S.; Wang, X.-W. A salivary effector enables whitefly to feed on host plants by eliciting salicylic acid-signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.H.; Lee, S.H.; Park, Y.-D. Development of mite (Tetranychus urticae)-resistant transgenic Chinese cabbage using plant-mediated RNA interference. Hortic. Environ. Biotechnol. 2020, 61, 305–315. [Google Scholar] [CrossRef]
- Saini, R.P.; Raman, V.; Dhandapani, G.; Malhotra, E.V.; Sreevathsa, R.; Kumar, P.A.; Sharma, T.R.; Pattanayak, D. Silencing of HaAce1 gene by host-delivered artificial microRNA disrupts growth and development of Helicoverpa armigera. PLoS ONE 2018, 13, e0194150. [Google Scholar] [CrossRef] [PubMed]
- Bally, J.; Fishilevich, E.; Doran, R.L.; Lee, K.; de Campos, S.B.; German, M.A.; Narva, K.E.; Waterhouse, P.M. Plin-amiR, a pre-microRNA-based technology for controlling herbivorous insect pests. Plant Biotechnol. J. 2020, 18, 1925–1932. [Google Scholar] [CrossRef]
- Rana, S.; Rajurkar, A.B.; Kumar, K.; Mohankumar, S. Comparative analysis of Chitin Synthase A dsRNA mediated RNA interference for management of crop pests of different families of Lepidoptera. Front. Plant Sci. 2020, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Liu, Z.; Chen, J.; Sun, G.; Jiang, Y.; Li, M.; Xiong, L.; Chen, S.; Zhou, Y.; Asad, M.; et al. Silencing arginine kinase/integrin β1 subunit by transgenic plant expressing dsRNA inhibits the development and survival of Plutella xylostella. Pest Manag. Sci. 2020, 76, 1761–1771. [Google Scholar] [CrossRef]
- Camargo, R.A.; Barbosa, G.O.; Possignolo, I.P.; Peres, L.E.; Lam, E.; Lima, J.E.; Figueira, A.; Marques-Souza, H. RNA interference as a gene silencing tool to control Tuta absoluta in tomato (Solanum lycopersicum). PeerJ 2016, 4, e2673. [Google Scholar] [CrossRef]
- Murtaza, S.; Tabassum, B.; Tariq, M.; Riaz, S.; Yousaf, I.; Jabbar, B.; Khan, A.; Samuel, A.O.; Zameer, M.; Nasir, I.A. Silencing a Myzus persicae macrophage inhibitory factor by plant-mediated RNAi induces enhanced aphid mortality coupled with boosted RNAi efficacy in transgenic potato lines. Mol. Biotechnol. 2022, 64, 1152–1163. [Google Scholar] [CrossRef]
- Rauf, I.; Asif, M.; Amin, I.; Naqvi, R.Z.; Umer, N.; Mansoor, S.; Jander, G. Silencing cathepsin L expression reduces Myzus persicae protein content and the nutritional value as prey for Coccinella septempunctata. Insect Mol. Biol. 2019, 28, 785–797. [Google Scholar] [CrossRef]
- Niu, X.; Kassa, A.; Hu, X.; Robeson, J.; McMahon, M.; Richtman, N.M.; Steimel, J.P.; Kernodle, B.M.; Crane, V.C.; Sandahl, G. Control of western corn rootworm (Diabrotica virgifera virgifera) reproduction through plant-mediated RNA interference. Sci. Rep. 2017, 7, 12591. [Google Scholar] [CrossRef]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W.; et al. Characterizing the mechanism of action of double-stranded RNA activity against Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yang, B.; Zhang, A.; Ding, D.; Wang, G. Plant-mediated RNAi for controlling Apolygus lucorum. Front. Plant Sci. 2019, 10, 64. [Google Scholar] [CrossRef]
- Liang, S.; Luo, J.; Alariqi, M.; Xu, Z.; Wang, A.; Zafar, M.N.; Ren, J.; Wang, F.; Liu, X.; Xin, Y. Silencing of a LIM gene in cotton exhibits enhanced resistance against Apolygus lucorum. J. Cell. Physiol. 2021, 236, 5921–5936. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, J.; Francis, F.; Chen, J. Molecular characterization and gene silencing of Laccase 1 in the grain aphid, Sitobion avenae. Arch. Insect Biochem. Physiol. 2018, 97, e21446. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Q.; Li, X.; Sun, Y.; Wang, H.; Xia, L. Double-stranded RNA in the biological control of grain aphid (Sitobion avenae F.). Funct. Integr. Genom. 2015, 15, 211–223. [Google Scholar] [CrossRef]
- Zhu, J.; Dong, Y.-C.; Li, P.; Niu, C.-Y. The effect of silencing 20E biosynthesis relative genes by feeding bacterially expressed dsRNA on the larval development of Chilo suppressalis. Sci. Rep. 2016, 6, 28697. [Google Scholar] [CrossRef]
- Mao, C.; Zhu, X.; Wang, P.; Sun, Y.; Huang, R.; Zhao, M.; Hull, J.J.; Lin, Y.; Zhou, F.; Chen, H. Transgenic double-stranded RNA rice, a potential strategy for controlling striped stem borer (Chilo suppressalis). Pest Manag. Sci. 2022, 78, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Saberi, E.; Mondal, M.; Paredes-Montero, J.R.; Nawaz, K.; Brown, J.K.; Qureshi, J.A. Optimal dsRNA concentration for RNA interference in Asian Citrus Psyllid. Insects 2024, 15, 58. [Google Scholar] [CrossRef]
- Killiny, N.; Hajeri, S.; Tiwari, S.; Gowda, S.; Stelinski, L.L. Double-stranded RNA uptake through topical application, mediates silencing of five CYP4 Genes and suppresses insecticide resistance in Diaphorina citri. PLoS ONE 2014, 9, e110536. [Google Scholar] [CrossRef]
- Julian-Chávez, B.; Siqueiros-Cendón, T.S.; Torres-Castillo, J.A.; Sinagawa-García, S.R.; Abraham-Juárez, M.J.; González-Barriga, C.D.; Rascón-Cruz, Q.; Siañez-Estrada, L.I.; Arévalo-Gallegos, S.; Espinoza-Sánchez, E.A. Silencing ACE1 gene with dsRNA of different lengths impairs larval development in Leptinotarsa decemlineata. Insects 2024, 15, 1000. [Google Scholar] [CrossRef]
- Zhu, F.; Xu, J.; Palli, R.; Ferguson, J.; Palli, S.R. Ingested RNA interference for managing the populations of the Colorado Potato Beetle, Leptinotarsa decemlineata. Pest Manag. Sci. 2011, 67, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Petek, M.; Coll, A.; Ferenc, R.; Razinger, J.; Gruden, K. Validating the potential of double-stranded RNA targeting Colorado Potato Beetle mesh gene in laboratory and field trials. Front. Plant Sci. 2020, 11, 1250. [Google Scholar] [CrossRef]
- Shen, C.-H.; Jin, L.; Fu, K.-Y.; Guo, W.-C.; Li, G.-Q. RNA interference targeting Ras GTPase gene Ran causes larval and adult lethality in Leptinotarsa decemlineata. Pest Manag. Sci. 2022, 78, 3849–3858. [Google Scholar] [CrossRef]
- Xu, Q.-Y.; Deng, P.; Mu, L.-L.; Fu, K.-Y.; Guo, W.-C.; Li, G.-Q. Silencing Taiman impairs larval development in Leptinotarsa decemlineata. Pestic. Biochem. Physiol. 2019, 160, 30–39. [Google Scholar] [CrossRef]
- Xu, Q.-Y.; Meng, Q.-W.; Deng, P.; Guo, W.-C.; Li, G.-Q. Leptinotarsa hormone receptor 4 (HR4) tunes ecdysteroidogenesis and mediates 20-hydroxyecdysone signaling during larval-pupal metamorphosis. Insect Biochem. Mol. Biol. 2018, 94, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Aksoy, E.; Çalışkan, M.E.; Bakhsh, A. Transgenic potato lines expressing hairpin RNAi construct of molting-associated EcR gene exhibit enhanced resistance against Colorado Potato Beetle (Leptinotarsa decemlineata, Say). Transgenic Res. 2019, 28, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Campuzano, C.; Martínez-Ramírez, A.C.; Contreras, E.; Rausell, C.; Real, M.D. Prohibitin, an essential protein for Colorado Potato Beetle larval viability, is relevant to Bacillus thuringiensis Cry3Aa toxicity. Pestic. Biochem. Physiol. 2013, 107, 299–308. [Google Scholar] [CrossRef]
- Liu, X.-P.; Fu, K.-Y.; Lü, F.-G.; Meng, Q.-W.; Guo, W.-C.; Li, G.-Q. Involvement of FTZ-F1 in the regulation of pupation in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2014, 55, 51–60. [Google Scholar] [CrossRef]
- Kong, Y.; Liu, X.-P.; Wan, P.-J.; Shi, X.-Q.; Guo, W.-C.; Li, G.-Q. The P450 enzyme Shade mediates the hydroxylation of ecdysone to 20-hydroxyecdysone in the Colorado Potato Beetle, Leptinotarsa decemlineata. Insect Mol. Biol. 2014, 23, 632–643. [Google Scholar] [CrossRef]
- Zhou, L.-T.; Jia, S.; Wan, P.-J.; Kong, Y.; Guo, W.-C.; Ahmat, T.; Li, G.-Q. RNA interference of a putative S-adenosyl-L-homocysteine hydrolase gene affects larval performance in Leptinotarsa decemlineata (Say). J. Insect Physiol. 2013, 59, 1049–1056. [Google Scholar] [CrossRef]
- Fu, K.-Y.; Guo, W.-C.; Ahmat, T.; Li, G.-Q. Knockdown of a nutrient amino acid transporter gene LdNAT1 reduces free neutral amino acid contents and impairs Leptinotarsa decemlineata pupation. Sci. Rep. 2015, 5, 18124. [Google Scholar] [CrossRef]
- Guo, W.-C.; Liu, X.-P.; Fu, K.-Y.; Shi, J.-F.; Lü, F.-G.; Li, G.-Q. Functions of nuclear receptor HR3 during larval-pupal molting in Leptinotarsa decemlineata (Say) revealed by in vivo RNA interference. Insect Biochem. Mol. Biol. 2015, 63, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.-J.; Fu, K.-Y.; Lü, F.-G.; Guo, W.-C.; Li, G.-Q. Knockdown of a putative alanine aminotransferase gene affects amino acid content and flight capacity in the Colorado Potato Beetle Leptinotarsa decemlineata. Amino Acids 2015, 47, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-F.; Fu, J.; Mu, L.-L.; Guo, W.-C.; Li, G.-Q. Two Leptinotarsa uridine diphosphate N-acetylglucosamine pyrophosphorylases are specialized for chitin synthesis in larval epidermal cuticle and midgut peritrophic matrix. Insect Biochem. Mol. Biol. 2016, 68, 1–12. [Google Scholar] [CrossRef]
- Shi, J.-F.; Xu, Q.-Y.; Sun, Q.-K.; Meng, Q.-W.; Mu, L.-L.; Guo, W.-C.; Li, G.-Q. Physiological roles of trehalose in Leptinotarsa larvae revealed by RNA interference of trehalose-6-phosphate synthase and trehalase genes. Insect Biochem. Mol. Biol. 2016, 77, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.-Y.; Li, Q.; Zhou, L.-T.; Meng, Q.-W.; Lü, F.-G.; Guo, W.-C.; Li, G.-Q. Knockdown of juvenile hormone acid methyl transferase severely affects the performance of Leptinotarsa decemlineata (Say) larvae and adults. Pest Manag. Sci. 2016, 72, 1231–1241. [Google Scholar] [CrossRef]
- Lü, F.-G.; Fu, K.-Y.; Guo, W.-C.; Li, G.-Q. Characterization of two juvenile hormone epoxide hydrolases by RNA interference in the Colorado potato beetle. Gene 2015, 570, 264–271. [Google Scholar] [CrossRef]
- Shi, J.F.; Mu, L.L.; Chen, X.; Guo, W.C.; Li, G.Q. RNA interference of chitin synthase genes inhibits chitin biosynthesis and affects larval performance in Leptinotarsa decemlineata (Say). Int. J. Biol. Sci. 2016, 12, 1319–1331. [Google Scholar] [CrossRef]
- Wan, P.-J.; Fu, K.-Y.; Lü, F.-G.; Wang, X.-X.; Guo, W.-C.; Li, G.-Q. Knocking down a putative -pyrroline-5-carboxylate dehydrogenase gene by RNA interference inhibits flight and causes adult lethality in the Colorado Potato Beetle Leptinotarsa decemlineata (Say). Pest Manag. Sci. 2015, 71, 1387–1396. [Google Scholar] [CrossRef]
- Guo, W.-C.; Liu, X.-P.; Fu, K.-Y.; Shi, J.-F.; Lü, F.-G.; Li, G.-Q. Nuclear receptor ecdysone-induced protein 75 is required for larval–pupal metamorphosis in the Colorado Potato Beetle Leptinotarsa decemlineata (Say). Insect Mol. Biol. 2016, 25, 44–57. [Google Scholar] [CrossRef]
- EPA. U.S. Environmental Protection Agency, Registers Innovative Tool to Control Corn Rootworm. Available online: https://www.epa.gov/ (accessed on 13 November 2024).
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef] [PubMed]
- ISAAA. GM Approval Database. Available online: https://www.isaaa.org/gmapprovaldatabase/ (accessed on 25 March 2025).
- EPA. U.S. Environmental Protection Agency: EPA Registers Novel Pesticide Technology for Potato Crops. Available online: https://www.epa.gov/pesticides/epa-registers-novel-pesticide-technology-potato-crops (accessed on 26 February 2025).
- Rodrigues, T.B.; Mishra, S.K.; Sridharan, K.; Barnes, E.R.; Alyokhin, A.; Tuttle, R.; Kokulapalan, W.; Garby, D.; Skizim, N.J.; Tang, Y.-W.; et al. First sprayable double-stranded RNA-based biopesticide product targets proteasome subunit beta type-5 in Colorado Potato Beetle (Leptinotarsa decemlineata). Front. Plant Sci. 2021, 12, 141530. [Google Scholar] [CrossRef]
- Dietz-Pfeilstetter, A.; Mendelsohn, M.; Gathmann, A.; Klinkenbuß, D. Considerations and regulatory approaches in the USA and in the EU for dsRNA-based externally applied pesticides for plant protection. Front. Plant Sci. 2021, 12, 682387. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, K.; Taning, C.N.T.; Van Daele, L.; Van Damme, E.J.M.; Dubruel, P.; Smagghe, G. RNAi-based biocontrol products: Market status, regulatory aspects, and risk assessment. Front. Insect Sci. 2022, 1, 818037. [Google Scholar] [CrossRef] [PubMed]
- Verdonckt, T.W.; Vanden, B.J. Methods for the cost-effective production of bacteria-derived double-stranded RNA for in vitro knockdown studies. Front. Physiol. 2022, 13, 836106. [Google Scholar] [CrossRef]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S.; Huvenne, H.; Smagghe, G. Delivery of dsRNA for RNAi in insects: An overview and future directions. Insect Sci. 2013, 20, 4–14. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef]
- Christiaens, O.; Swevers, L.; Smagghe, G. DsRNA degradation in the pea aphid (Acyrthosiphon pisum) associated with lack of response in RNAi feeding and injection assay. Peptides 2014, 53, 307–314. [Google Scholar] [CrossRef]
- Shukla, J.N.; Megha, K.; Amit, S.; E. Narva, K.; Elane, F.; Satnam, S.; Kanakachari, M.; Palli, S.R. Reduced stability and intracellular transport of dsRNA contribute to poor RNAi response in lepidopteran insects. RNA Biol. 2016, 13, 656–669. [Google Scholar] [CrossRef]
- Lu, A.Y.H.; Junk, K.W.; Coon, M.J. Resolution of the Cytochrome P-450-containing ω-hydroxylation system of liver microsomes into three components. J. Biol. Chem. 1969, 244, 3714–3721. [Google Scholar] [CrossRef]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; Le Goff, G.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, G.J.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef]
- Feyereisen, R. Origin and evolution of the CYP4G subfamily in insects, cytochrome P450 enzymes involved in cuticular hydrocarbon synthesis. Mol. Phylogenet. Evol. 2020, 143, 106695. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Sams, S.; Moural, T.; Haynes, K.F.; Potter, M.F.; Palli, S.R. RNA Interference of NADPH-Cytochrome P450 Reductase results in reduced insecticide resistance in the bed bug, Cimex lectularius. PLoS ONE 2012, 7, e31037. [Google Scholar] [CrossRef]
- Rewitz, K.F.; O’Connor, M.B.; Gilbert, L.I. Molecular evolution of the insect Halloween family of cytochrome P450s: Phylogeny, gene organization and functional conservation. Insect Biochem. Mol. Biol. 2007, 37, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Maïbèche-Coisne, M.; Nikonov, A.A.; Ishida, Y.; Jacquin-Joly, E.; Leal, W.S. Pheromone anosmia in a scarab beetle induced by in vivo inhibition of a pheromone-degrading enzyme. Proc. Natl. Acad. Sci. USA 2004, 101, 11459–11464. [Google Scholar] [CrossRef]
- Sandstrom, P.; Welch, W.H.; Blomquist, G.J.; Tittiger, C. Functional expression of a bark beetle cytochrome P450 that hydroxylates myrcene to ipsdienol. Insect Biochem. Mol. Biol. 2006, 36, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Lycett, G.J.; McLaughlin, L.A.; Ranson, H.; Hemingway, J.; Kafatos, F.C.; Loukeris, T.G.; Paine, M.J.I. Anopheles gambiae P450 reductase is highly expressed in oenocytes and in vivo knockdown increases permethrin susceptibility. Insect Mol. Biol. 2006, 15, 321–327. [Google Scholar] [CrossRef]
- Huang, Y.; Lu, X.P.; Wang, L.L.; Wei, D.; Feng, Z.J.; Zhang, Q.; Xiao, L.F.; Dou, W.; Wang, J.J. Functional characterization of NADPH-cytochrome P450 reductase from Bactrocera dorsalis: Possible involvement in susceptibility to malathion. Sci. Rep. 2015, 5, 18394. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Nissen, P. P-Type ATPases. Annu. Rev. 2011, 40, 243–266. [Google Scholar] [CrossRef]
- Berchtold, M.W.; Brinkmeier, H.; Müntener, M. Calcium Ion in skeletal muscle: Its crucial role for muscle function, plasticity, and disease. Physiol. Rev. 2000, 80, 1215–1265. [Google Scholar] [CrossRef]
- Primeau, J.O.; Armanious, G.P.; Fisher, M.L.E.; Young, H.S. The SarcoEndoplasmic Reticulum Calcium ATPase. In Membrane Protein Complexes: Structure and Function; Harris, J.R., Boekema, E.J., Eds.; Springer: Singapore, 2018; pp. 229–258. [Google Scholar] [CrossRef]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Endo, M. Calcium-induced calcium release in skeletal muscle. Physiol. Rev. 2009, 89, 1153–1176. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Jassar, O.; Ghanim, M. The plant pathogenic bacterium Candidatus Liberibacter solanacearum induces calcium-regulated autophagy in midgut cells of its insect vector Bactericera trigonica. Microbiol. Spectr. 2023, 11, e0130123. [Google Scholar] [CrossRef]
- Sanyal, S.; Consoulas, C.; Kuromi, H.; Basole, A.; Mukai, L.; Kidokoro, Y.; Krishnan, K.S.; Ramaswami, M. Analysis of conditional paralytic mutants in Drosophila sarco-endoplasmic reticulum calcium ATPase reveals novel mechanisms for regulating membrane excitability. Genetics 2005, 169, 737–750. [Google Scholar] [CrossRef]
- Jayakumar, S.; Hasan, G. Neuronal calcium signaling in metabolic regulation and adaptation to nutrient stress. Front. Neural Circuits 2018, 12, 25. [Google Scholar] [CrossRef]
- Denmeade, S.R.; Isaacs, J.T. The SERCA pump as a therapeutic target: Making a “smart bomb” for prostate cancer. Cancer Biol. Ther. 2005, 4, 14–22. [Google Scholar] [CrossRef]
- Xu, H.; Van Remmen, H. The SarcoEndoplasmic Reticulum Calcium ATPase (SERCA) pump: A potential target for intervention in aging and skeletal muscle pathologies. Skelet. Muscle 2021, 11, 25. [Google Scholar] [CrossRef]
- Zhao, B.; Lucas, K.J.; Saha, T.T.; Ha, J.; Ling, L.; Kokoza, V.A.; Roy, S.; Raikhel, A.S. MicroRNA-275 targets sarco/endoplasmic reticulum Ca2+ adenosine triphosphatase (SERCA) to control key functions in the mosquito gut. PLoS Genet. Genet. 2017, 13, e1006943. [Google Scholar] [CrossRef]
- He, W.; Xu, W.; Xu, L.; Fu, K.; Guo, W.; Bock, R.; Zhang, J. Length-dependent accumulation of double-stranded RNAs in plastids affects RNA interference efficiency in the Colorado Potato Beetle. J. Exp. Bot. 2020, 71, 2670–2677. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Guo, W.-C.; Fu, K.-Y.; Yang, S.; Li, X.-X.; Li, G.-Q. Instar-dependent systemic RNA interference response in Leptinotarsa decemlineata larvae. Pestic. Biochem. Physiol. 2015, 123, 64–73. [Google Scholar] [CrossRef] [PubMed]
- UN. United Nations. Department of Economic and Social Affairs. Population Division. 2022 Revision of World Population Prospects. Available online: https://population.un.org/wpp/ (accessed on 20 March 2025).
- FAO. Food and Agriculture Organization of the United Nations. Plant Health 2020. Available online: https://www.fao.org/plant-health-2020/en/ (accessed on 25 February 2025).
- Rizzo, D.M.; Lichtveld, M.; Mazet, J.A.K.; Togami, E.; Miller, S.A. Plant health and its effects on food safety and security in a One Health framework: Four case studies. One Health Outlook 2021, 3, 6. [Google Scholar] [CrossRef]
- Hemathilake, D.M.K.S.; Gunathilake, D.M.C.C. Chapter 31—Agricultural productivity and food supply to meet increased demands. In Future Foods; Bhat, R., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 539–553. [Google Scholar] [CrossRef]
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations. Feeding the World, Eradicating Hunger. Available online: https://www.fao.org/fileadmin/templates/wsfs/Summit/WSFS_Issues_papers/WSFS_Background_paper_Feeding_the_world.pdf (accessed on 25 February 2025).
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Hens, L.; Quynh, L.X. Environmental Space. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Ray, D.K.; Ramankutty, N.; Mueller, N.D.; West, P.C.; Foley, J.A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 2012, 3, 1293. [Google Scholar] [CrossRef]
- Pimentel, D. Techniques for Reducing Pesticide Use: Economic and Environmental Benefits; John Wiley and Sons: New York, NY, USA, 1997. [Google Scholar]
- Paoletti, M.G.; Pimentel, D. Environmental risks of pesticides versus genetic engineering for agricultural pest control. J. Agric. Environ. Ethics 2000, 12, 279–303. [Google Scholar] [CrossRef]
- Grapputo, A.; Boman, S.; LindstrÖM, L.; Lyytinen, A.; Mappes, J. The voyage of an invasive species across continents: Genetic diversity of North American and European Colorado Potato Beetle populations. Mol. Ecol. 2005, 14, 4207–4219. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, R.A. The Colorado Potato Beetle: 125 years of mismanagement. Bull. Entomol. Soc. Am. 1987, 33, 142–150. [Google Scholar] [CrossRef]
- Khajuria, C.; Ivashuta, S.; Wiggins, E.; Flagel, L.; Moar, W.; Pleau, M.; Miller, K.; Zhang, Y.; Ramaseshadri, P.; Jiang, C.; et al. Development and characterization of the first dsRNA-resistant insect population from western corn rootworm, Diabrotica virgifera virgifera LeConte. PLoS ONE 2018, 13, e0197059. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Dee, J.; Moar, W.; Dufner-Beattie, J.; Baum, J.; Dias, N.P.; Alyokhin, A.; Buzza, A.; Rondon, S.I.; Clough, M.; et al. Selection for high levels of resistance to double-stranded RNA (dsRNA) in Colorado potato beetle (Leptinotarsa decemlineata Say) using non-transgenic foliar delivery. Sci. Rep. 2021, 11, 6523. [Google Scholar] [CrossRef]
- Pallis, S.; Alyokhin, A.; Manley, B.; Rodrigues, T.B.; Barnes, E.; Narva, K. Baseline susceptibility to a novel dsRNA-based insecticide across US populations of Colorado Potato Beetle. Agriculture 2023, 13, 2283. [Google Scholar] [CrossRef]
- Sanyal, S.; Jennings, T.; Dowse, H.; Ramaswami, M. Conditional mutations in SERCA, the Sarco-endoplasmic reticulum Ca2+-ATPase, alter heart rate and rhythmicity in Drosophila. J. Comp. Physiol. B 2006, 176, 253–263. [Google Scholar] [CrossRef]
- Zhu, W.; Duan, Y.; Chen, J.; Merzendorfer, H.; Zou, X.; Yang, Q. SERCA interacts with chitin synthase and participates in cuticular chitin biogenesis in Drosophila. Insect Biochem. Mol. Biol. 2022, 145, 103783. [Google Scholar] [CrossRef]
- Chen, X.E.; Zhang, Y. Identification and characterization of NADPH-dependent cytochrome P450 reductase gene and cytochrome b5 gene from Plutella xylostella: Possible involvement in resistance to beta-cypermethrin. Gene 2015, 558, 208–214. [Google Scholar] [CrossRef]
- Huang, Y.; Liao, M.; Yang, Q.; Shi, S.; Xiao, J.; Cao, H. Knockdown of NADPH-cytochrome P450 reductase and CYP6MS1 increases the susceptibility of Sitophilus zeamais to terpinen-4-ol. Pestic. Biochem. Physiol. 2020, 162, 15–22. [Google Scholar] [CrossRef]
- Liu, N.; Li, M.; Gong, Y.; Liu, F.; Li, T. Cytochrome P450s—Their expression, regulation, and role in insecticide resistance. Pestic. Biochem. Physiol. 2015, 120, 77–81. [Google Scholar] [CrossRef]
- Tang, Q.; Li, X.; He, Y.; Ma, K. RNA interference of NADPH-cytochrome P450 reductase increases the susceptibility of Aphis gossypii Glover to sulfoxaflor. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 274, 109745. [Google Scholar] [CrossRef] [PubMed]
- Adesanya, A.W.; Cardenas, A.; Lavine, M.D.; Walsh, D.B.; Lavine, L.C.; Zhu, F. RNA interference of NADPH-cytochrome P450 reductase increases susceptibilities to multiple acaricides in Tetranychus urticae. Pestic. Biochem. Physiol. 2020, 165, 104550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Wang, L.; Yao, J.; Guo, H.; Fang, J. Knockdown of NADPH-cytochrome P450 reductase results in reduced resistance to buprofezin in the small brown planthopper, Laodelphax striatellus (fallén). Pestic. Biochem. Physiol. 2016, 127, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, J.; Liu, J.; Li, Y.; Liu, X.; Wu, H.; Ma, E.; Zhang, J. Knockdown of NADPH-cytochrome P450 reductase increases the susceptibility to carbaryl in the migratory locust, Locusta migratoria. Chemosphere 2017, 188, 517–524. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, J.; Shen, G.; Xu, Z.; Wei, P.; Zhang, Y.; Xu, Q.; He, L. Silencing NADPH-cytochrome P450 reductase results in reduced acaricide resistance in Tetranychus cinnabarinus (Boisduval). Sci. Rep. 2015, 5, 15581. [Google Scholar] [CrossRef]
- Liu, S.; Liang, Q.-M.; Zhou, W.-W.; Jiang, Y.-D.; Zhu, Q.-Z.; Yu, H.; Zhang, C.-X.; Gurr, G.M.; Zhu, Z.-R. RNA interference of NADPH–cytochrome P450 reductase of the rice brown planthopper, Nilaparvata lugens, increases susceptibility to insecticides. Pest Manag. Sci. 2015, 71, 32–39. [Google Scholar] [CrossRef]
- Kitzmann, P.; Schwirz, J.; Schmitt-Engel, C.; Bucher, G. RNAi phenotypes are influenced by the genetic background of the injected strain. BMC Genom. 2013, 14, 5. [Google Scholar] [CrossRef]
- Mehlhorn, S.; Hunnekuhl, V.S.; Geibel, S.; Nauen, R.; Bucher, G. Establishing RNAi for basic research and pest control and identification of the most efficient target genes for pest control: A brief guide. Front. Zool. 2021, 18, 60. [Google Scholar] [CrossRef]
- Cagliari, D.; Dias, N.P.; dos Santos, E.Á.; Rickes, L.N.; Kremer, F.S.; Farias, J.R.; Lenz, G.; Galdeano, D.M.; Garcia, F.R.M.; Smagghe, G.; et al. First transcriptome of the Neotropical pest Euschistus heros (Hemiptera: Pentatomidae) with dissection of its siRNA machinery. Sci. Rep. 2020, 10, 4856. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Wang, Y.-C.; Zhang, X.-X.; Iqbal, J.; Du, Y.-Z. RNA interference of genes encoding the Vacuolar-ATPase in Liriomyza trifoliis. Insects 2021, 12, 41. [Google Scholar] [CrossRef]
- Adeyinka, O.S.; Nasir, I.A.; Riaz, S.; Yousaf, I.; Toufiq, N.; Okiki, A.P.; Tabassum, B. A Protective dsRNA is crucial for optimum RNAi gene silencing in Chilo partellus. Int. J. Agric. Biol. 2021, 25, 1238–1248. [Google Scholar] [CrossRef]
- Westerhout, E.M.; Berkhout, B. A systematic analysis of the effect of target RNA structure on RNA interference. Nucleic Acids Res. 2007, 35, 4322–4330. [Google Scholar] [CrossRef]
- Cao, M.; Gatehouse, J.A.; Fitches, E.C. A Systematic study of RNAi effects and dsRNA stability in Tribolium castaneum and Acyrthosiphon pisum, following injection and ingestion of analogous dsRNAs. Int. J. Mol. Sci. 2018, 19, 1079. [Google Scholar] [CrossRef]
- Elbashir, S.M.; Harborth, J.; Weber, K.; Tuschl, T. Analysis of gene function in somatic mammalian cells using small interfering RNAs. Methods 2002, 26, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Teimoori-Toolabi, L.; Hashemi, S.; Azadmanesh, K.; Eghbalpour, F.; Safavifar, F.; Khorramizadeh, M.R. Silencing the wild-type and mutant K-ras increases the resistance to 5-flurouracil in HCT-116 as a colorectal cancer cell line. Anti-Cancer Drugs 2015, 26, 187–196. [Google Scholar] [CrossRef]
- Fakhr, E.; Zare, F.; Teimoori-Toolabi, L. Precise and efficient siRNA design: A key point in competent gene silencing. Cancer Gene Ther. 2016, 23, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, J.; Lenartowicz, E.; Soszynska-Jozwiak, M.; Szutkowska, B.; Kierzek, R.; Kierzek, E. RNA secondary structure motifs of the influenza A virus as targets for siRNA-mediated RNA interference. Mol. Ther. Nucleic Acids 2020, 19, 627–642. [Google Scholar] [CrossRef]
- Luo, K.Q.; Chang, D.C. The gene-silencing efficiency of siRNA is strongly dependent on the local structure of mRNA at the targeted region. Biochem. Biophys. Res. Commun. 2004, 318, 303–310. [Google Scholar] [CrossRef]
- Shao, Y.; Chan, C.Y.; Maliyekkel, A.; Lawrence, C.E.; Roninson, I.B.; Ding, Y. Effect of target secondary structure on RNAi efficiency. RNA 2007, 13, 1631–1640. [Google Scholar] [CrossRef]
- Lu, Z.J.; Mathews, D.H. Efficient siRNA selection using hybridization thermodynamics. Nucleic Acids Res. 2007, 36, 640–647. [Google Scholar] [CrossRef]

| Gene | Sequence (5′-3′) |
|---|---|
| First amplicon | |
| SERCA | Fw—5′TAATACGACTCACTATAGGATGGAGGACGGTCACATTAAAA3′ |
| Rv—5′ATTAAAAGAATTACGAAGGGTT3′ | |
| CPR | Fw—5′TAATACGACTCACTATAGGATGTCCGAGAGTGAAGCAAAAG3′ |
| Rv—5′TCTTCTCCTGTTCCAGTTTGAC3′ | |
| Second amplicon | |
| SERCA | Fw—5′ATGGAGGACGGTCACATTAAAA3′ |
| Rv—5′TAATACGACTCACTATAGGATTAAAAGAATTACGAAGGGTT3′ | |
| CPR | Fw—5′ATGTCCGAGAGTGAAGCAAAAG3′ |
| Rv—5′TAATACGACTCACTATAGGTCTTCTCCTGTTCCAGTTTGAC3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendoza-Alatorre, M.; Siqueiros-Cendón, T.S.; Torres-Castillo, J.A.; Sinagawa-García, S.R.; Iglesias-Figueroa, B.F.; Abraham-Juárez, M.J.; González-Barriga, C.D.; Rascón-Cruz, Q.; Siañez-Estrada, L.I.; Espinoza-Sánchez, E.A. Gene Silencing of Sarco/Endoplasmic Reticulum Ca2+-ATPase and NADPH–Cytochrome P450 Reductase as a Novel Approach to Leptinotarsa decemlineata Management. Agronomy 2025, 15, 1151. https://doi.org/10.3390/agronomy15051151
Mendoza-Alatorre M, Siqueiros-Cendón TS, Torres-Castillo JA, Sinagawa-García SR, Iglesias-Figueroa BF, Abraham-Juárez MJ, González-Barriga CD, Rascón-Cruz Q, Siañez-Estrada LI, Espinoza-Sánchez EA. Gene Silencing of Sarco/Endoplasmic Reticulum Ca2+-ATPase and NADPH–Cytochrome P450 Reductase as a Novel Approach to Leptinotarsa decemlineata Management. Agronomy. 2025; 15(5):1151. https://doi.org/10.3390/agronomy15051151
Chicago/Turabian StyleMendoza-Alatorre, Maribel, Tania Samanta Siqueiros-Cendón, Jorge Ariel Torres-Castillo, Sugey Ramona Sinagawa-García, Blanca Flor Iglesias-Figueroa, María Jazmín Abraham-Juárez, Carmen Daniela González-Barriga, Quintín Rascón-Cruz, Luis Ignacio Siañez-Estrada, and Edward Alexander Espinoza-Sánchez. 2025. "Gene Silencing of Sarco/Endoplasmic Reticulum Ca2+-ATPase and NADPH–Cytochrome P450 Reductase as a Novel Approach to Leptinotarsa decemlineata Management" Agronomy 15, no. 5: 1151. https://doi.org/10.3390/agronomy15051151
APA StyleMendoza-Alatorre, M., Siqueiros-Cendón, T. S., Torres-Castillo, J. A., Sinagawa-García, S. R., Iglesias-Figueroa, B. F., Abraham-Juárez, M. J., González-Barriga, C. D., Rascón-Cruz, Q., Siañez-Estrada, L. I., & Espinoza-Sánchez, E. A. (2025). Gene Silencing of Sarco/Endoplasmic Reticulum Ca2+-ATPase and NADPH–Cytochrome P450 Reductase as a Novel Approach to Leptinotarsa decemlineata Management. Agronomy, 15(5), 1151. https://doi.org/10.3390/agronomy15051151