
Cover Crops Optimize Soil Fertility and Soybean Productivity in the Cerrado of MATOPIBA, Brazil

 ,
,  , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
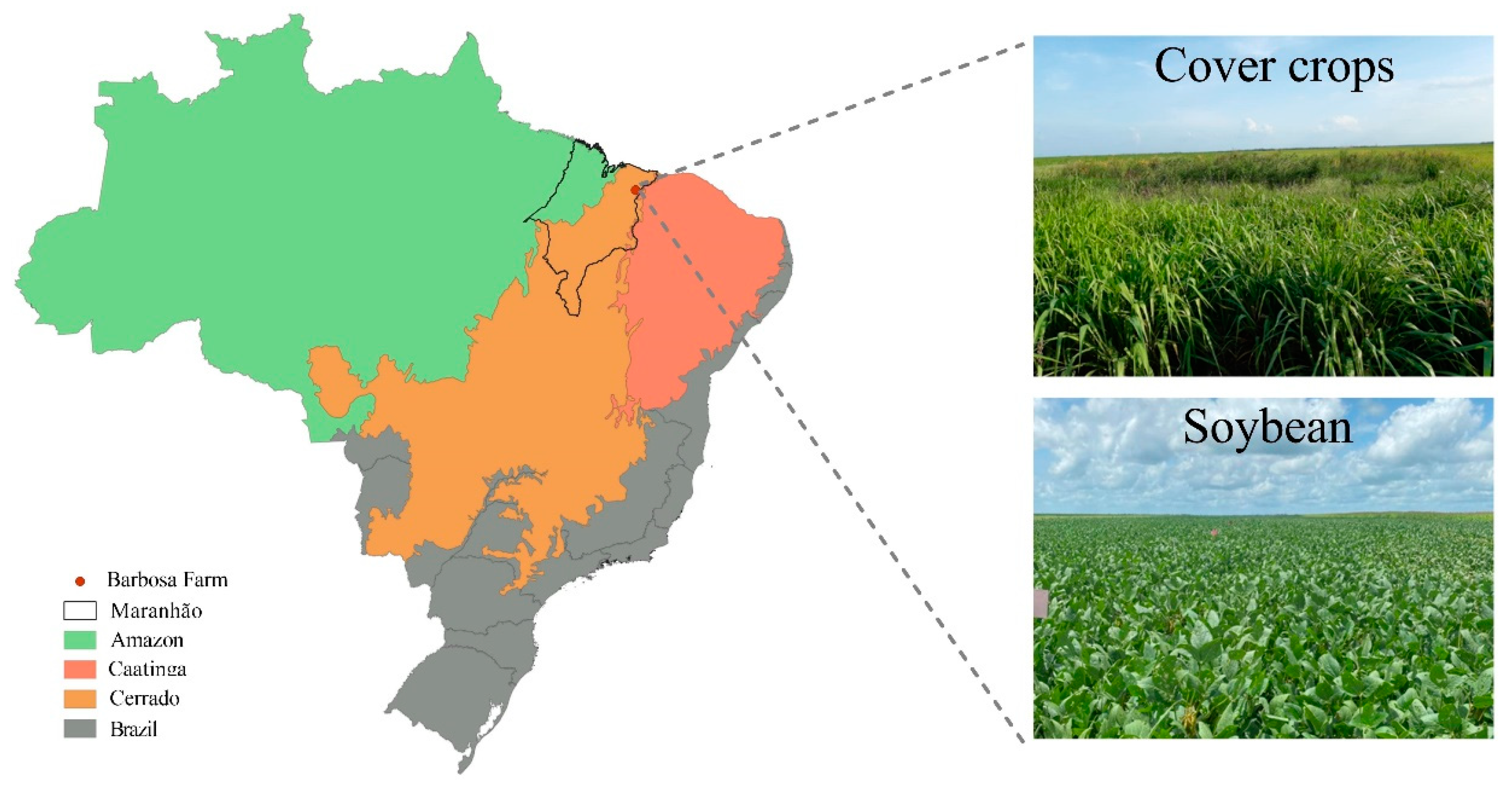
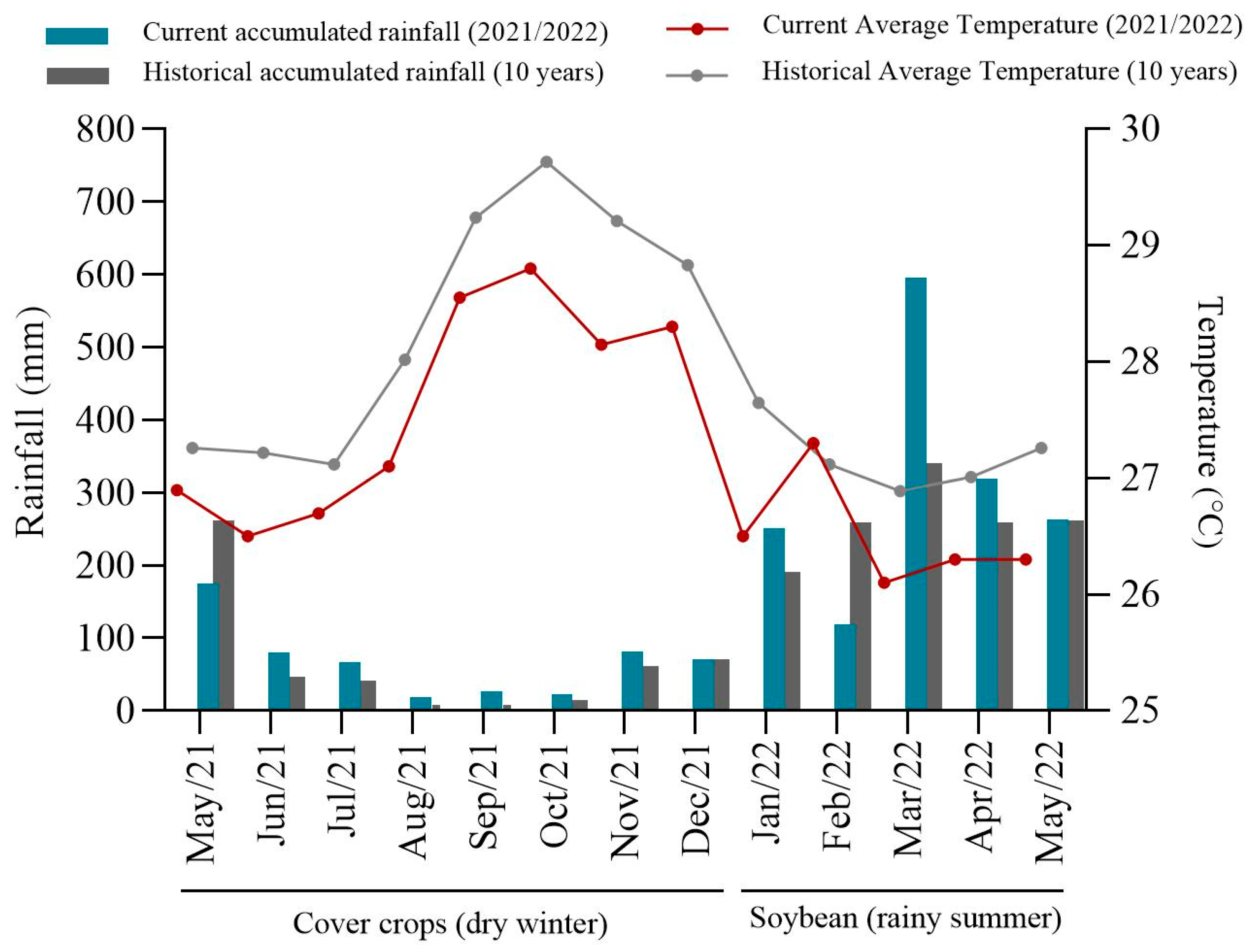
2.1. Site Details and Experimental Design
2.2. Conducting and Evaluating Cover Crops
2.3. Conducting and Evaluating Soybeans
2.4. Statistical Analysis
3. Results
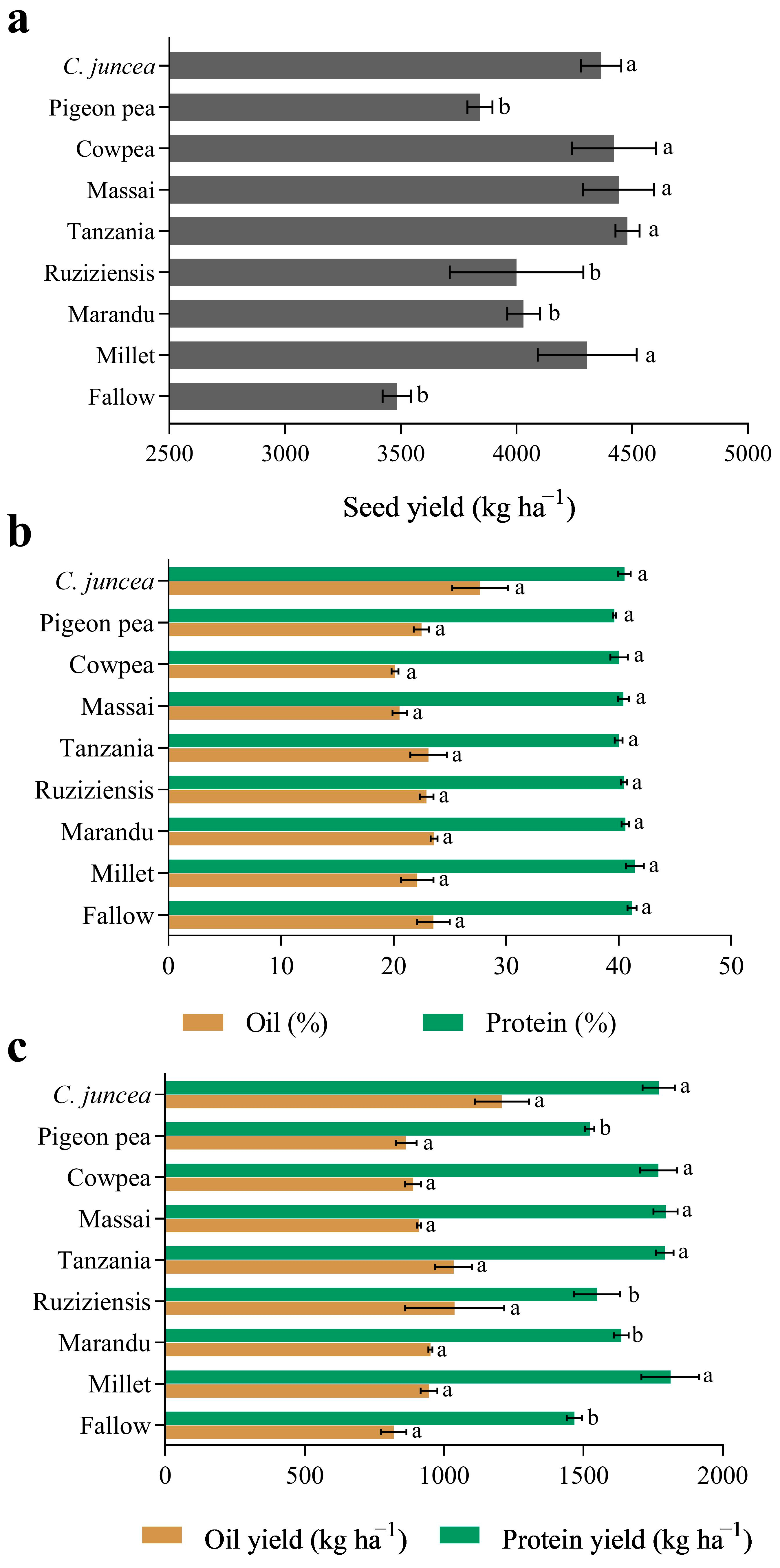
3.1. Quantity and Quality of Cover Crop Biomass
3.2. Productivity and Composition of Soybean Seeds
3.3. Soil Biological Properties
3.4. Soil Chemical Properties
3.5. Determination of Nutritional Status of Soybeans
3.6. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USDA. United States Department of Agriculture. World Agricultural Production. 2023. Available online: https://apps.fas.usda.gov/psdonline/circulars/production.pdf.ANO (accessed on 1 November 2024).
- Araújo, M.L.S.; Sano, E.E.; Bolfe, É.L.; Santos, J.R.N.; Santos, J.S.; Silva, F.B. Spatiotemporal dynamics of soybean crop in the Matopiba region, Brazil (1990–2015). Land Use Policy 2019, 80, 57–67. [Google Scholar] [CrossRef]
- Araújo, M.L.S.D.; Rufino, I.A.A.; Silva, F.B.; Brito, H.C.D.; Santos, J.R.N. The relationship between climate, agriculture and land cover in Matopiba, Brazil (1985–2020). Sustainability 2024, 16, 2670. [Google Scholar] [CrossRef]
- Reis, L.; Silva, C.M.S.E.; Bezerra, B.; Mutti, P.; Spyrides, M.H.; Silva, P.; Magalhães, T.; Ferreira, R.; Rodrigues, D.; Andrade, L. Influence of climate variability on soybean yield in Matopiba, Brazil. Atmosphere 2020, 11, 1130. [Google Scholar] [CrossRef]
- Pompeu, J. Legal deforestation can jeopardize plant diversity conservation in an agricultural frontier in the brazilian Cerrado: A spatial explicit contribution to Santana and Simon (2022). Biodivers. Conserv. 2022, 31, 2899–2903. [Google Scholar] [CrossRef]
- Nascente, A.S.; Stone, L.F. Cover crops as affecting soil chemical and physical properties and development of upland rice and soybean cultivated in rotation. Rice Sci. 2018, 25, 340–349. [Google Scholar] [CrossRef]
- Ghimire, R.; Ghimire, B.; Mesbah, A.O.; Sainju, U.M.; Idowu, O.J. Soil health response of cover crops in winter wheat–fallow system. Agron. J. 2019, 111, 2108–2115. [Google Scholar] [CrossRef]
- Jian, J.; Du, X.; Reiter, M.S.; Stewart, R.D. A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biol. Biochem. 2020, 143, 107735. [Google Scholar] [CrossRef]
- Martínez-García, L.B.; Korthals, G.; Brussaard, L.; Jørgensen, H.B.; De Deyn, G.B. Organic management and cover crop species steer soil microbial community structure and functionality along with soil organic matter properties. Agric. Ecosyst. Environ. 2018, 263, 7–17. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Hallama, M.; Pekrun, C.; Lambers, H.; Kandeler, E. Hidden miners–the roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2019, 434, 7–45. [Google Scholar] [CrossRef]
- Hallama, M.; Pekrun, C.; Pilz, S.; Jarosch, K.A.; Frąc, M.; Uksa, M.; Marhan, S.; Kandeler, E. Interactions between cover crops and soil microorganisms increase phosphorus availability in conservation agriculture. Plant Soil 2021, 463, 307–328. [Google Scholar] [CrossRef]
- Romdhane, S.; Spor, A.; Busset, H.; Falchetto, L.; Martin, J.; Bizouard, F.; Bru, D.; Breuil, M.; Philippot, L.; Cordeau, S. Cover crop management practices rather than composition of cover crop mixtures affect bacterial communities in no-till agroecosystems. Front. Microbiol. 2019, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Balbinot Junior, A.A.; Debiasi, H.; Franchini, J.C.; Oliveira, M.A.; Coelho, A.E.; Moraes, M.T. Soybean yield, seed protein and oil concentration, and soil fertility affected by off-season crops. Eur. J. Agron. 2024, 153, 127039. [Google Scholar] [CrossRef]
- Krenchinski, F.H.; Cesco, V.J.S.; Rodrigues, D.M.; Albrecht, L.P.; Wobeto, K.S.; Albrecht, A.J.P. Agronomic performance of soybean grown in succession to winter cover crops. Pes. Agropec. Bras. 2018, 53, 909–917. [Google Scholar] [CrossRef]
- Theurl, M.C.; Lauk, C.; Kalt, G.; Mayer, A.; Kaltenegger, K.; Morais, T.G.; Teixeira, R.F.M.; Domingos, T.; Winiwarter, W.; Erb, K.; et al. Food systems in a zero-deforestation world: Dietary change is more important than intensification for climate targets in 2050. Sci. Total Environ. 2020, 735, 139353. [Google Scholar] [CrossRef]
- Castro, G.S.; Crusciol, C.A.; Calonego, J.C.; Rosolem, C.A. Management impacts on soil organic matter of tropical soils. Vadose Zone J. 2015, 14, vzj2014-07. [Google Scholar] [CrossRef]
- Silva, J.F.D.; Gontijo Neto, M.M.; Silva, G.F.D.; Borghi, E.; Calonego, J.C. Soil organic matter and aggregate stability in soybean, maize and Urochloa production systems in a very clayey soil of the brazilian Savanna. Agronomy 2022, 12, 1652. [Google Scholar] [CrossRef]
- Aparecido, L.E.O.; Meneses, K.C.; Lorençone, P.A.; Lorençone, J.A.; Moraes, J.R.D.S.C.D.; Rolim, G.S. Climate classification by Thornthwaite (1948) humidity index in future scenarios for Maranhão State, Brazil. Environ. Dev. Sustain. 2022, 25, 855–878. [Google Scholar] [CrossRef]
- Sousa, D.C.; Rosa, J.D.; Medeiros, J.C.; Boechat, C.L.; Nóbrega, R.S.A.; Souza, H.A.; Sagrilo, E. Microbial indicators of soil quality and soybean yield in agricultural production system using cover crops under no-tillage. Aust. J. Crop Sci. 2023, 17, 507–513. [Google Scholar] [CrossRef]
- Bublitz, L.R.; Gurgel, A.L.C.; Mauri, A.C.; Queiroz, V.C.; Lima, K.S.; Campelo, I.B.R.; Araújo, M.J.; Dias-Silva, T.P.; Barros, J.S.; Aguiar, I.O.M.; et al. Panicum maximum cultivars for use in integrated agricultural production systems in Cerrado biome soils. Grassland Sci. 2024, 70, 121–129. [Google Scholar] [CrossRef]
- Tanaka, K.S.; Crusciol, C.A.; Soratto, R.P.; Momesso, L.; Costa, C.H.; Franzluebbers, A.J.; Oliveira Júnior, A.; Calonego, J.C. Nutrients released by Urochloa cover crops prior to soybean. Nutr. Cycl. Agroecosyst. 2019, 113, 267–281. [Google Scholar] [CrossRef]
- Aiosa, M.L.; Neely, C.B.; Morgan, C.L.; Jessup, R.W.; Corriher-Olson, V.A.; Somenahally, A.C.; Norman, K.D.; Smith, G.R.; Rouquette Júnior, F.M. Cowpeas as a summer cover crop for forage rye. Agrosyst. Geosci. Environ. 2020, 3, e20057. [Google Scholar] [CrossRef]
- Rebonatti, M.D.; Cordeiro, C.F.S.; Volf, M.R.; Silva, P.C.G.; Tiritan, C.S. Effects of silage crops between crop seasons on soybean grain yield and soil fertility in tropical sandy soils. Eur. J. Agron. 2023, 143, 126685. [Google Scholar] [CrossRef]
- Atakoun, A.M.; Tovihoudji, P.G.; Diogo, R.V.; Yemadje, P.L.; Balarabe, O.; Akponikpè, P.I.; Sekloka, E.; Hougni, A.; Tittonell, P. Evaluation of cover crop contributions to conservation agriculture in northern Benin. Field Crops Res. 2023, 303, 109118. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.D.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araujo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos, 3rd ed.; Embrapa Solos: Brasília, Brazil, 2018; p. 356. [Google Scholar]
- USDA–NRCS. Keys to Soil Taxonomy, 12th ed.; USDA: Washington DC, USA, 2014; 142p. [Google Scholar]
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo, 3rd ed.; Embrapa Cerrado: Brasília, Brazil, 2017; p. 574. [Google Scholar]
- Miyazawa, M.; Pavan, M.A.; Muraoka, T.; Carmo, C.A.F.S.; Melo, W.J. Análise química de tecido vegetal. In Manual de Análises Químicas de Solos, Plantas e Fertilizantes; Embrapa: Brasília, Brazil, 2009; pp. 191–234. [Google Scholar]
- Van Soest, P.J.; Robertison, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Tedesco, M.; Gianello, G.; Bissani, C.; Bohnen, H.; Volkweiss, S. Análises de Solo, Plantas e Outros Materiais; Universidade Federal do Rio Grande do Sul: Rio Grande do Sul, Brazil, 1995; p. 174. [Google Scholar]
- Ferreira, A.S.; Camargo, F.A.O.; Vidor, C. Utilização de microondas na avaliação da biomassa microbiana do solo. Rev. Bras. Ciênc. Solo 1999, 23, 991–996. [Google Scholar] [CrossRef]
- Alef, K. Estimation of soil respiration. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 464–467. [Google Scholar]
- Silva, E.E.; Azevedo, P.H.S.; De-Polli, H. Determinação do Carbono da Biomassa Microbiana do Solo (BMS-C); Embrapa Agrobiologia: Rio de Janeiro, Brazil, 2007; p. 6. [Google Scholar]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Aust. J. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- Schnurer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl Environ. Microb. 1982, 43, 1256–1261. [Google Scholar] [CrossRef]
- Chen, W.; Hoitink, H.A.J.; Madden, L.V. Microbial activity and biomass in container media for predicting suppressiveness to damping-off caused by Pythiumultimum. Phytopathology 1988, 78, 1447–1450. [Google Scholar] [CrossRef]
- Casida, L.E.J.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Bitton, G.; Ben, K. Biochemical tests for toxicity screening. In Toxicity Testing Using Microorganisms; Bitton, G., Dutka, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 1986; pp. 27–55. [Google Scholar]
- Silva, D.; Queiroz, A.C. Análise de Alimentos: Métodos Químicos e Biológicos; Universidade Federal de Viçosa: Viçosa, Brazil, 2006; p. 235. [Google Scholar]
- Cafaro La Menza, N.; Monzón, J.P.; Specht, J.E.; Grassini, P. Is soybean yield limited by nitrogen supply? Field Crop Res. 2017, 213, 204–212. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.; González, L.; Tablada, M.; Robledo, C.W. InfoStat; Universidad Nacional de Córdoba: Córdoba, Argentina, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 3 February 2023).
- Steinauer, K.; Tilman, D.; Wragg, P.D.; Cesarz, S.; Cowles, J.M.; Pritsch, K.; Reich, P.B.; Weisser, W.W.; Eisenhauer, N. Plant diversity effects on soil microbial functions and enzymes are stronger than warming in a grassland experiment. Ecology 2015, 96, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhu, H.; Fu, S.; Yao, Q. Variation in soil microbial community structure associated with different legume species is greater than that associated with different grass species. Front. Microbiol. 2017, 8, 1007. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, Y.; Li, T.; Chen, L.; Chen, Y.; Sui, P. Changes in soil microbial biomass, diversity, and activity with crop rotation in cropping systems: A global synthesis. Appl. Soil Ecol. 2023, 186, 104815. [Google Scholar] [CrossRef]
- Ball, K.R.; Baldock, J.A.; Pendolf, C.; Power, S.A.; Woodin, S.J.; Smith, P.; Pendall, E. Soil organic carbon and nitrogen pools are increased by mixed grass and legume cover crops in vineyard agroecosystems: Detecting short-term management effects using infrared spectroscopy. Geoderma 2020, 379, 114619. [Google Scholar] [CrossRef]
- Wittwer, R.A.; Dorn, B.; Jossi, W.; Heijden, M.G.V.D. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 41911. [Google Scholar] [CrossRef]
- Sievers, T.; Cook, R.L. Aboveground and root decomposition of cereal rye and hairy vetch cover crops. Soil Sci. Soc. Am. J. 2018, 82, 147–155. [Google Scholar] [CrossRef]
- Adhikari, A.D.; Shrestha, P.; Ghimire, R.; Liu, Z.; Pollock, D.A.; Acharya, P.; Aryal, D.R. Cover crop residue quality regulates litter decomposition dynamics and soil carbon mineralization kinetics in semi-arid cropping systems. Appl. Soil. Ecol. 2024, 193, 105160. [Google Scholar] [CrossRef]
- Duval, M.E.; Galantini, J.A.; Capurro, J.E.; Martinez, J.M. Winter cover crops in soybean monoculture: Effects on soil organic carbon and its fractions. Soil Tillage Res. 2016, 161, 95–105. [Google Scholar] [CrossRef]
- Marschner, P.; Hatam, Z.; Cavagnaro, T.R. Soil respiration, microbial biomass and nutrient availability after the second amendment are influenced by legacy effects of prior residue addition. Soil Biol. Biochem. 2015, 88, 169–177. [Google Scholar] [CrossRef]
- Mazzuchelli, R.C.L.; Araujo, A.S.F.; Moro, E.; Araujo, F.F. Changes in soil properties and crop yield as a function of early desiccation of pastures. J. Soil Sci. Plant Nutr. 2020, 20, 840–848. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Cavagnaro, T.R.; Ngo, H.T.T.; Marschner, P. Soil respiration, microbial biomass and nutrient availability in soil amended with high and low C/N residue–Influence of interval between residue additions. Soil Biol. Biochem. 2016, 95, 189–197. [Google Scholar] [CrossRef]
- Spohn, M. Microbial respiration per unit microbial biomass depends on litter layer carbon-to-nitrogen ratio. Biogeosciences 2015, 12, 817–823. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Hämmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon: Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef]
- Novak, E.; Carvalho, L.A.; Santiago, E.F.; Portilho, I.I.R. Chemical and microbiological attributes under different soil cover. Cerne 2017, 23, 19–30. [Google Scholar] [CrossRef]
- Balota, E.L.; Calegari, A.; Nakatani, A.S.; Coyne, M.S. Benefits of winter cover crops and no-tillage for microbial parameters in a Brazilian Oxisol: A long-term study. Agric. Ecosyst. Environ. 2014, 197, 31–40. [Google Scholar] [CrossRef]
- Segatelli, C.R.; Câmara, G.M.D.S.; Aguila, L.S.H.D.; Aguila, J.S.D.; Francisco, E.A.B.; Piedade, S.M.S. Soybean yield under no-tillage system with an early Eleusine coracana fertilization. Rev. Caatinga 2022, 35, 308–319. [Google Scholar] [CrossRef]
- Mubvumba, P.; Tyler, H.L. Evaluation of single and mixed cover crops species in a sandy loam soil under corn production. Agron. J. 2024, 116, 1655–1669. [Google Scholar] [CrossRef]
- Baptistella, J.L.C.; Andrade, S.A.L.; Favarin, J.L.; Mazzafera, P. Urochloa in tropical agroecosystems. Front. Sust. Food Syst. 2020, 4, 119. [Google Scholar] [CrossRef]
- Costa, N.R.; Andreotti, M.; Crusciol, C.A.C.; Pariz, C.M.; Bossolani, J.W.; Pascoaloto, I.M.; Calonego, J.C. Soybean yield and nutrition after tropical forage grasses. Nutr. Cycl. Agroecosyst. 2021, 121, 31–49. [Google Scholar] [CrossRef]
- Crusciol, C.A.; Nascente, A.S.; Borghi, E.; Soratto, R.P.; Martins, P.O. Improving soil fertility and crop yield in a tropical region with palisadegrass cover crops. Agron. J. 2015, 107, 2271–2280. [Google Scholar] [CrossRef]
- Romanuik, R.I.; Beltran, M.J.; Brutti, L.; Constantini, A.O.; Bacigaluppo, S.; Sainz Rozas, H.; Salvagiotti, F. Soil organic carbon, macro- and micronutrient changes in soil fractions with different lability in response to crop intensification. Soil Tillage Res. 2018, 181, 136–143. [Google Scholar] [CrossRef]
- Almeida, D.S.; Delai, L.B.; Sawaya, A.C.H.F.; Rosolem, C.A. Exudation of organic acid anions by tropical grasses in response to low phosphorus availability. Sci. Rep. 2020, 10, 16955. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Hao, M.; Shao, M.; Gale, W.J. Changes in soil properties and the availability of soil micronutrients after 18 years of cropping and fertilization. Soil Tillage Res. 2006, 91, 120–130. [Google Scholar] [CrossRef]
- Cordeiro, C.F.S.; Echer, F.R.; Araujo, F.F. Cover crops impact crops yields by improving microbiological activity and fertility in sandy soil. J. Soil Sci. Plant Nutr. 2021, 21, 1968–1977. [Google Scholar] [CrossRef]
- Colombo, C.; Iorio, E.; Liu, Q.; Jiang, Z.; Barrón, B. Iron oxide nonanoparticles in soils: Environmental and agronomical importance. J. Nanisci. Nanotechnol. 2017, 17, 4449–4460. [Google Scholar] [CrossRef]
- Beltrán, M.; Galantini, J.A.; Salvagiotti, F.; Tognetti, P.; Bacigaluppo, S.; Sainz Rozas, H.R.; Barraco, M.; Barbieri, P.A. Do soil carbon sequestration and soil fertility increase by including a gramineous cover crop in continuous soybean? Soil Sci. Soc. Am. J. 2021, 85, 1380–1394. [Google Scholar] [CrossRef]
- Lopes, A.S.; Cox, F.R. A Survey of the Fertility Status of Surface Soils Under “Cerrado” Vegetation in Brazil. Soil Sci. Soc. Am. J. 1977, 41, 742–747. [Google Scholar] [CrossRef]
- Pires, M.F.M.; Souza, H.A.; Medeiros, J.C.; Dalla Rosa, J.; Martins, R.V.S.; Sobral, A.H.S.; Carvalho, S.P.; Vera, G.S.; Vieira, P.F.M.J.; Sagrilo, E. Nutrient uptake by soybean plants in succession of cover crops in northeast of Brazil. Commun. Soil Sci. Plant Anal. 2023, 54, 945–963. [Google Scholar] [CrossRef]
- Rodrigues, L.U.; Silva, R.R. Boron availability in building up fertility in Cerrado soil of Tocantins. Commun. Soil Sci. Plant Anal. 2020, 51, 595–603. [Google Scholar] [CrossRef]
- Silva, L.S.; Laroca, J.V.S.; Coelho, A.P.; Gonçalves, E.C.; Gomes, R.P.; Pacheco, L.P.; Carvalho, P.C.F.; Pires, G.C.; Oliveira, R.L.; Souza, J.M.A.; et al. Does grass-legume intercropping change soil quality and grain yield in integrated crop-livestock systems? Appl. Soil Ecol. 2022, 170, 104257. [Google Scholar] [CrossRef]
- Finney, D.M.; White, C.M.; Kaye, J.P. Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agron. J. 2016, 108, 39–52. [Google Scholar] [CrossRef]
- Lewis, K.L.; Burke, J.A.; Keeling, W.S.; McCallister, D.M.; DeLaune, P.B.; Keeling, J.W. Soil benefits and yield limitations of cover crop use in Texas High Plains cotton. Agron. J. 2018, 110, 1616–1623. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Farias, J.R.; Neumaier, N.; Nepomuceno, A.L. Modeling nitrogen accumulation and use by soybean. Field Crop Res. 2003, 81, 149–158. [Google Scholar] [CrossRef]
- Cafaro La Menza, N.; Monzón, J.P.; Specht, J.E.; Lindquist, J.L.; Arkebauer, T.J.; Gref, G.; Grassini, P. Nitrogen limitation in high-yield soybean: Seed yield, N accumulation, and N-use efficiency. Field Crop Res. 2019, 237, 74–81. [Google Scholar] [CrossRef]
- Kirkpatrick, D.; Roberts, T.L.; Brye, K.; Ross, J. Influence of cover crops on soybean yield and partial returns as an alternative to double-crop soybean in Arkansas. Agron. J. 2023, 115, 1373–1383. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH (H2O) | TOC | P | K+ | Ca2+ | Mg2+ | S-SO42− | Al3+ | CEC |
| g kg−1 | mg dm−3 | cmolc dm−3 | mg dm−3 | cmolc dm−3 | ||||
| 5.35 | 15 | 85 | 0.04 | 0.97 | 0.44 | 5.73 | 0.10 | 4.66 |
| BS | Cu | Fe | Mn | Zn | B | sand | silt | clay |
| % | mg dm−3 | g kg−1 | ||||||
| 31 | 0.09 | 64 | 0.33 | 2.9 | 0.20 | 774 | 92 | 134 |
| Cover Crop | Biomass | N | P | K | Ca | Mg | S |
| kg ha−1 | g kg−1 | ||||||
| Fallow | 1515 ± 247 c | 41 ± 2 a | 3.8 ± 0.2 a | 15 ± 0 b | 7.1 ± 0.7 b | 5.1 ± 0.5 b | 1.7 ± 0.2 b |
| Millet | 2027 ± 456 b | 12 ± 1 d | 2.4 ± 0.4 b | 13 ± 1 b | 5.0 ± 1.1 c | 3.6 ± 0.4 c | 1.1 ± 0.4 b |
| Marandu | 2100 ± 361 b | 15 ± 3 d | 2.7 ± 0.4 b | 26 ± 2 a | 3.6 ± 0.4 c | 4.5 ± 1.4 b | 1.3 ± 0.3 b |
| Ruziziensis | 1813 ± 280 b | 14 ± 3 d | 2.6 ± 0.6 b | 28 ± 0 a | 4.6 ± 1.4 c | 4.0 ± 0.8 c | 2.4 ± 0.8 a |
| Tanzania | 1980 ± 275 b | 13 ± 1 d | 1.1 ± 0.5 c | 25 ± 2 a | 5.0 ± 0 c | 2.8 ± 0.9 c | 1.0 ± 0.2 b |
| Massai | 3093 ± 193 a | 10 ± 1 d | 1.4 ± 0.7 c | 25 ± 4 a | 3.7 ± 0.5 c | 2.6 ± 0.3 c | 1.3 ± 0.2 b |
| Cowpea | 728 ± 271 c | 32 ± 4 b | 4.3 ± 0.5 a | 27 ± 3 a | 12.1 ± 1.4 a | 6.9 ± 0.3 a | 2.3 ± 0.2 a |
| Pigeon pea | 1480 ± 175 c | 20 ± 3 c | 4.3 ± 0.1 a | 17 ± 1 b | 5.3 ± 0.3 c | 2.8 ± 0.3 c | 2.2 ± 0.4 a |
| C. juncea | 1068 ± 412 c | 20 ± 1 c | 3.6 ± 0.1 a | 16 ± 4 b | 4.6 ± 0.7 c | 3.4 ± 0.7 c | 1.3 ± 0.3 b |
| F test | ** | ** | ** | ** | ** | ** | ** |
| CV (%) | 16.8 | 13.2 | 15.7 | 11.1 | 13.4 | 17.7 | 23.7 |
| Cover Crop | Cu | Fe | Zn | Mn | B | Lignin/N | C/N |
| mg kg−1 | |||||||
| Fallow | 9 ± 3 b | 326 ± 24 a | 42 ± 8 a | 14 ± 1 b | 18 ± 2 b | 0.8 ± 0.1 c | 10 ± 1 d |
| Millet | 12 ± 2 b | 213 ± 25 b | 46 ± 1 a | 25 ± 1 b | 13 ± 4 b | 5.6 ± 1.2 a | 35 ± 3 a |
| Marandu | 14 ± 4 b | 178 ± 4 c | 22 ± 3 c | 31 ± 12 a | 11 ± 1 b | 3.2 ± 1.2 b | 27 ± 6 b |
| Ruziziensis | 17 ± 5 a | 209 ± 36 b | 25 ± 1 b | 36 ± 13 a | 14 ± 3 b | 2.8 ± 0.1 b | 31 ± 6 b |
| Tanzania | 14 ± 1 b | 193 ± 1 c | 18 ± 3 c | 21 ± 5 b | 7 ± 1 c | 3.1 ± 0.4 b | 30 ± 4 b |
| Massai | 14 ± 1 b | 81 ± 4 d | 16 ± 1 c | 48 ± 16 a | 5 ± 3 c | 6.6 ± 1.4 a | 40 ± 2 a |
| Cowpea | 19 ± 1 a | 218 ± 20 b | 47 ± 8 a | 34 ± 7 a | 27 ± 4 a | 1.5 ± 0.2 c | 11 ± 2 d |
| Pigeon pea | 20 ± 3 a | 72 ± 17 d | 31 ± 6 b | 14 ± 1 b | 14 ± 4 b | 7.6 ± 1.3 a | 23 ± 4 c |
| C. juncea | 19 ± 3 a | 54 ± 6 d | 28 ± 8 b | 20 ± 6 b | 11 ± 2 b | 4.3 ± 1.2 b | 21 ± 1 c |
| F test | ** | ** | ** | ** | ** | ** | ** |
| CV (%) | 18.3 | 9.7 | 17.9 | 25.1 | 20.6 | 22.1 | 14.4 |
| Cover Crop | TOC | MBC | MBN | BSR | qCO2 | qMic | DHA | FDA |
| g kg−1 | mg kg−1 | mg C-CO2 kg−1 d−1 | mg C-CO2 kg−1 MBC d−1 | % | µl de H g−1 | µg FDA g−1 | ||
| Fallow | 18 ± 0.7 b | 105 + 7 b | 0.23 + 0 c | 25.7 + 6.0 b | 0.21 + 0.02 b | 0.57 + 0.03 b | 4.3 + 0.1 b | 20.1 + 2.3 |
| Millet | 16 ± 2.4 b | 84 + 6 b | 0.98 + 0.1 b | 23.6 + 3.8 b | 0.32 + 0.02 a | 0.52 + 0.06 b | 4.5 + 1.3 b | 32.7 + 0.9 |
| Marandu | 20 ± 1.1 a | 123 + 12 a | 0.80 + 0.2 b | 28.4 + 5.0 a | 0.25 + 0.05 b | 0.63 + 0.10 a | 5.7 + 0.6 a | 26.6 + 0.8 |
| Ruziziensis | 19 ± 0.4 a | 139 + 19 a | 0.72 + 0.2 b | 29.6 + 1.0 a | 0.22 + 0.02 b | 0.72 + 0.06 a | 3.9 + 0.7 b | 26.4 + 4.0 |
| Tanzania | 21 ± 1.0 a | 140 + 9 a | 0.73 + 0.1 b | 24.1 + 1.5 b | 0.20 + 0.03 b | 0.68 + 0.02 a | 4.3 + 1.2 b | 24.9 + 1.1 |
| Massai | 17 ± 2.9 b | 100 + 14 b | 0.59 + 0.1 b | 33.1 + 2.0 a | 0.29 + 0.07 a | 0.69 + 0.04 a | 3.7 + 0.9 b | 26.0 + 3.2 |
| Cowpea | 21 ± 0.6 a | 115 + 20 a | 1.34 + 0.2 a | 23.5 + 0.4 b | 0.21 + 0.03 b | 0.66 + 0.04 a | 6.4 + 0.2 a | 26.9 + 4.0 |
| Pigeon pea | 22 ± 1.3 a | 98 + 6 b | 2.13 + 0.3 a | 25.9 + 3.1 b | 0.24 + 0.01 b | 0.45 + 0.03 b | 4.0 + 0.1 b | 29.8 + 1.4 |
| Crotalaria juncea | 20 ± 0.1 a | 99 + 7 b | 1.76 + 0.3 a | 25.3 + 0.1 b | 0.25 + 0.01 b | 0.48 + 0.03 b | 2.5 + 0.2 b | 28.1 + 1.5 |
| F test | ** | ** | ** | ** | ** | ** | ** | ns |
| CV (%) | 7.4 | 13.3 | 19.4 | 10.6 | 13.5 | 10.1 | 17.7 | 8.9 |
| Cover Crop | pH (H2O) | P | K+ | Ca2+ | Mg2+ | S-SO42− |
| mg dm−3 | ------------ cmolc dm−3 ------------ | mg dm−3 | ||||
| Fallow | 5.35 ± 0.3 | 54 ± 13 a | 0.10 ± 0.00 b | 0.86 ± 0.01 d | 0.49 ± 0.05 b | 4.88 ± 1.3 a |
| Millet | 5.17 ± 0.2 | 47 ± 1 b | 0.09 ± 0.00 b | 0.84 ± 0.12 d | 0.44 ± 0.09 b | 3.15 ± 0.7 b |
| Marandu | 5.20 ± 0.2 | 61 ± 9 a | 0.11 ± 0.02 a | 1.00 ± 0.00 c | 0.49 ± 0.02 b | 5.79 ± 0.3 a |
| Ruziziensis | 5.35 ± 0.1 | 65 ± 7 a | 0.09 ± 0.01 b | 1.08 ± 0.02 c | 0.50 ± 0.05 b | 4.09 ± 0.9 b |
| Tanzania | 5.31 ± 0.1 | 39 ± 5 b | 0.09 ± 0.00 b | 1.10 ± 0.02 c | 0.50 ± 0.04 b | 3.77 ± 0.7 b |
| Massai | 5.27 ± 0.3 | 40 ± 5 b | 0.11 ± 0.00 a | 0.83 ± 0.01 d | 0.54 ± 0.15 b | 2.65 ± 0.9 b |
| Cowpea | 5.33 ± 0.1 | 54 ± 3 a | 0.11 ± 0.01 a | 1.42 ± 0.28 b | 0.68 ± 0.13 a | 3.87 ± 0.3 b |
| Pigeon pea | 5.28 ± 0.3 | 64 ± 8 a | 0.09 ± 0.01 b | 1.80 ± 0.02 a | 0.71 ± 0.06 a | 5.06 ± 0.6 a |
| Crotalaria juncea | 5.26 ± 0.1 | 48 ± 2 b | 0.11 ± 0.01 a | 1.04 ± 0.05 c | 0.57 ± 0.08 b | 4.07 ± 0.2 b |
| F test | ns | ** | ** | ** | ** | ** |
| CV (%) | 3.5 | 13.1 | 8.0 | 9.0 | 14.5 | 16.3 |
| Cover Crop | Cu | Fe | Mn | Zn | B | CEC |
| mg dm−3 | cmolc dm−3 | |||||
| Fallow | 0.11 ± 0.02 b | 36 ± 3 | 0.52 ± 0.03 | 0.70 ± 0.29 b | 0.19 ± 0.01 | 5.38 ± 0.4 |
| Millet | 0.12 ± 0.01 b | 46 ± 1 | 0.51 ± 0.10 | 1.61 ± 0.42 a | 0.18 ± 0.02 | 5.06 ± 0.5 |
| Marandu | 0.12 ± 0.03 b | 47 ± 14 | 0.62 ± 0.21 | 1.39 ± 0.31 a | 0.19 ± 0.02 | 4.99 ± 0.4 |
| Ruziziensis | 0.11 ± 0.00 b | 45 ± 7 | 0.66 ± 0.04 | 1.21 ± 0.51 a | 0.20 ± 0.01 | 5.13 ± 0.0 |
| Tanzania | 0.11 ± 0.02 b | 35 ± 11 | 0.49 ± 0.09 | 1.18 ± 0.36 a | 0.17 ± 0.03 | 5.20 ± 0.6 |
| Massai | 0.10 ± 0.03 b | 46 ± 3 | 0.56 ± 0.02 | 0.85 ± 0.48 b | 0.18 ± 0.03 | 4.99 ± 0.7 |
| Cowpea | 0.10 ± 0.01 b | 38 ± 1 | 0.68 ± 0.03 | 1.46 ± 0.12 a | 0.18 ± 0.01 | 5.81 ± 0.5 |
| Pigeon pea | 0.17 ± 0.05 a | 42 ± 9 | 0.62 ± 0.21 | 0.92 ± 0.21 b | 0.18 ± 0.03 | 6.11 ± 0.3 |
| Crotalaria juncea | 0.11 ± 0.02 b | 47 ± 13 | 0.46 ± 0.17 | 0.57 ± 0.20 b | 0.14 ± 0.02 | 5.26 ± 0.4 |
| F test | * | ns | ns | ** | ns | ns |
| CV (%) | 18.2 | 19.6 | 16.7 | 23.8 | 11.8 | 8.0 |
| Cover Crop | N | P | K | Ca | Mg | S | Cu | Fe | Mn | Zn | B |
| -------------------g kg−1 ------------------- | --------------mg kg−1 -------------- | ||||||||||
| Fallow | 49.8 ± 0.0 | 4.2 ± 0.3 | 22.7 ± 0.8 b | 6.2 ± 0.3 | 4.3 ± 0.2 | 3.3 ± 0.1 | 9 ± 0.6 | 84 ± 5 b | 86 ± 19 a | 121 ± 33 | 75 ± 0 b |
| Millet | 49.7 ± 1.4 | 4.0 ± 0.3 | 20.7 ± 0.3 b | 5.7 ± 0.7 | 4.3 ± 0.1 | 3.2 ± 0.0 | 10 ± 1.7 | 82 ± 7 b | 69 ± 1 b | 84 ± 2 | 74 ± 6 b |
| Marandu | 50.3 ± 1.7 | 4.4 ± 0.2 | 24.9 ± 1.8 a | 6.1 ± 0.0 | 4.2 ± 0.2 | 3.9 ± 0.9 | 10 ± 2.6 | 90 ± 6 a | 109 ± 1 a | 111 ± 38 | 65 ± 1 b |
| Ruziziensis | 50.6 ± 1.4 | 4.7 ± 0.4 | 24.5 ± 1.0 a | 5.8 ± 0.9 | 4.3 ± 0.1 | 3.6 ± 0.7 | 9 ± 1.6 | 90 ± 6 a | 88 ± 38 a | 131 ± 23 | 76 ± 9 b |
| Tanzania | 49.8 ± 0.7 | 4.7 ± 0.1 | 24.3 ± 1.7 a | 5.7 ± 0.9 | 4.3 ± 0.2 | 3.1 ± 0.1 | 8 ± 1.2 | 92 ± 6 a | 52 ± 3 b | 74 ± 4 | 70 ± 9 b |
| Massai | 50.4 ± 2.4 | 4.8 ± 0.6 | 23.6 ± 0.1 a | 5.2 ± 0.8 | 4.2 ± 0.2 | 3.6 ± 0.1 | 10 ± 0.9 | 82 ± 5 b | 56 ± 1 b | 89 ± 23 | 73 ± 8 b |
| Cowpea | 51.3 ± 0.9 | 4.8 ± 0.8 | 24.0 ± 0.5 a | 5.5 ± 0.0 | 4.1 ± 0.6 | 3.2 ± 0.3 | 12 ± 0.2 | 87 ± 5 a | 101 ± 4 a | 114 ± 28 | 90 ± 0 a |
| Pigeon pea | 48.7 ± 1.3 | 4.5 ± 0.5 | 25.4 ± 3.0 a | 5.7 ± 0.3 | 4.6 ± 0.4 | 3.2 ± 0.6 | 11 ± 1.4 | 78 ± 1 b | 90 ± 32 a | 109 ± 12 | 80 ± 1 a |
| Crotalaria juncea | 50.4 ± 0.1 | 4.8 ± 0.3 | 24.3 ± 1.5 a | 5.5 ± 0.7 | 4.3 ± 0.2 | 3.6 ± 0.2 | 10 ± 1.6 | 87 ± 8 a | 75 ± 25 b | 98 ± 25 | 69 ± 0 b |
| F test | ns | ns | ** | ns | ns | ns | ns | * | ** | ns | ** |
| CV (%) | 2.5 | 7.6 | 5.4 | 7.9 | 6.2 | 13.4 | 13.6 | 5.6 | 19.6 | 20.7 | 7.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, H.A.F.d.; Sagrilo, E.; Oliveira Júnior, J.O.L.d.; Sousa, D.C.d.; Costa, C.P.d.M.; Costa, P.M.; Araújo Neto, R.B.d.; Alcantara, R.M.C.M.d.; Leite, L.F.C.; Furtado, M.B.; et al. Cover Crops Optimize Soil Fertility and Soybean Productivity in the Cerrado of MATOPIBA, Brazil. Agronomy 2025, 15, 1083. https://doi.org/10.3390/agronomy15051083
Andrade HAFd, Sagrilo E, Oliveira Júnior JOLd, Sousa DCd, Costa CPdM, Costa PM, Araújo Neto RBd, Alcantara RMCMd, Leite LFC, Furtado MB, et al. Cover Crops Optimize Soil Fertility and Soybean Productivity in the Cerrado of MATOPIBA, Brazil. Agronomy. 2025; 15(5):1083. https://doi.org/10.3390/agronomy15051083
Chicago/Turabian StyleAndrade, Hosana Aguiar Freitas de, Edvaldo Sagrilo, José Oscar Lustosa de Oliveira Júnior, Daiane Conceição de Sousa, Carlos Pedro de Menezes Costa, Paula Muniz Costa, Raimundo Bezerra de Araújo Neto, Rosa Maria Cardoso Mota de Alcantara, Luiz Fernando Carvalho Leite, Mariléia Barros Furtado, and et al. 2025. "Cover Crops Optimize Soil Fertility and Soybean Productivity in the Cerrado of MATOPIBA, Brazil" Agronomy 15, no. 5: 1083. https://doi.org/10.3390/agronomy15051083
APA StyleAndrade, H. A. F. d., Sagrilo, E., Oliveira Júnior, J. O. L. d., Sousa, D. C. d., Costa, C. P. d. M., Costa, P. M., Araújo Neto, R. B. d., Alcantara, R. M. C. M. d., Leite, L. F. C., Furtado, M. B., Beltrán, M. J., Cafaro La Menza, N., & Souza, H. A. d. (2025). Cover Crops Optimize Soil Fertility and Soybean Productivity in the Cerrado of MATOPIBA, Brazil. Agronomy, 15(5), 1083. https://doi.org/10.3390/agronomy15051083