
Changes in Soil Properties, Content of Cd, and Cd-Resistant Bacterial Community with Biochar After One-Time Addition Under Soybean Continuous Cropping and Crop Rotation Patterns

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Description and Experimental Design
2.2. Sample Collection
2.3. Analysis of Soil Physicochemical Properties
2.4. Soil DNA Extraction, Quantitative PCR, and High-Throughput Sequencing
2.5. Statistical Analysis
3. Results
3.1. Soil Properties
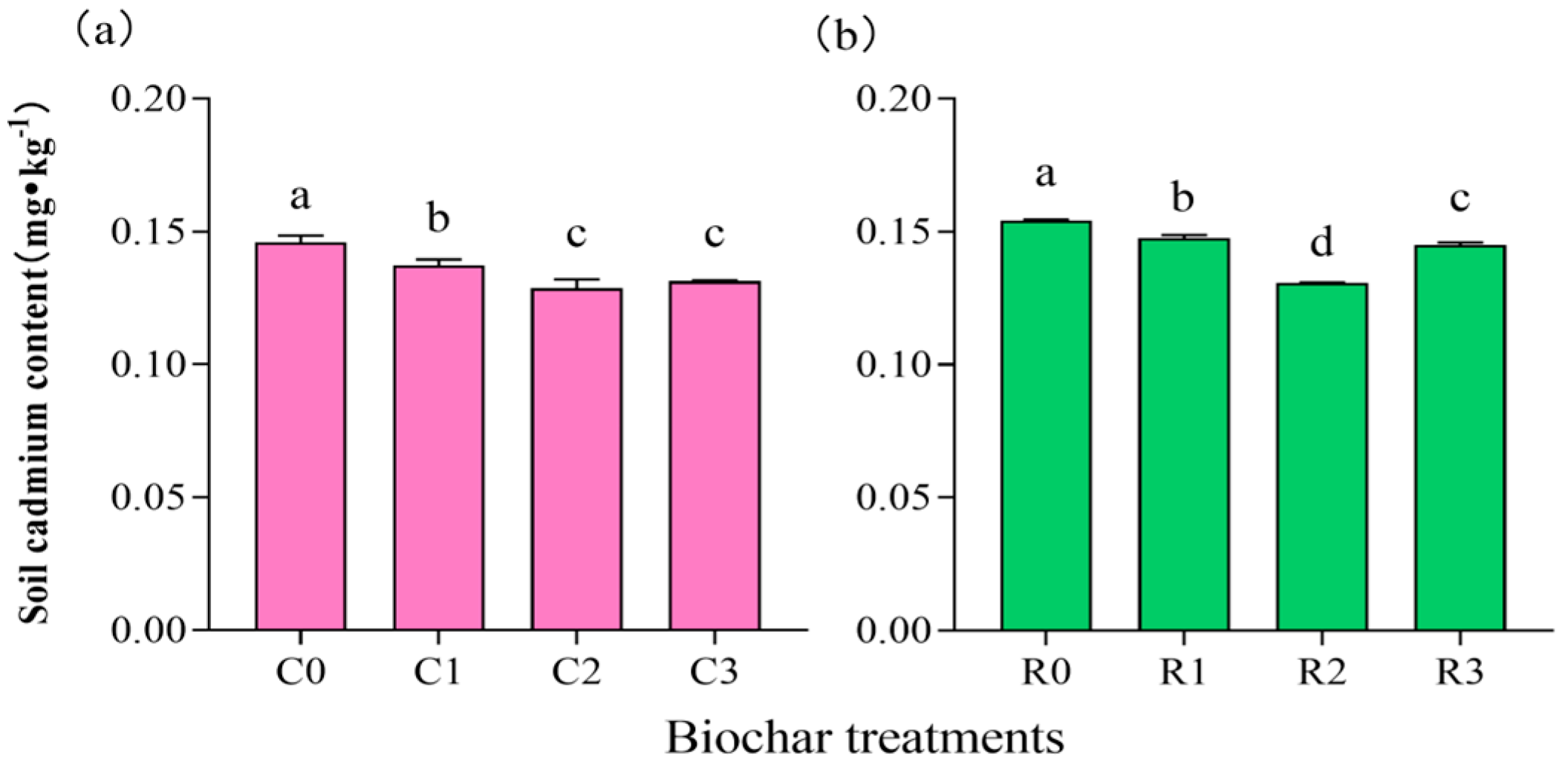
3.2. Cd Content of Soil and Soybean Grain
3.3. Absolute Abundance of the czcA Gene
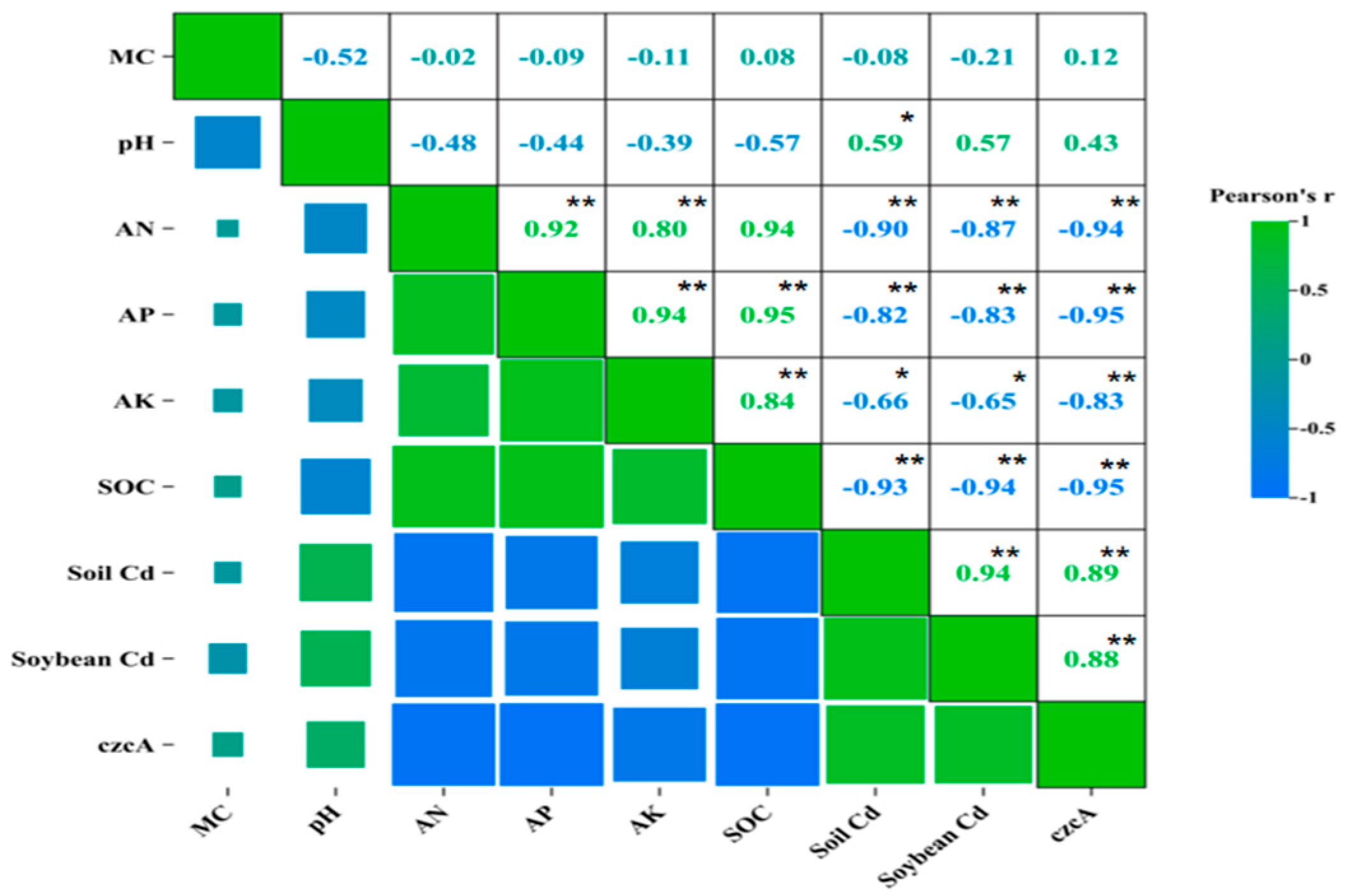
3.4. Correlations Between the Soil Properties, Cd Content, and czcA Abundance
3.5. Analysis of czcA Gene Microbial Community
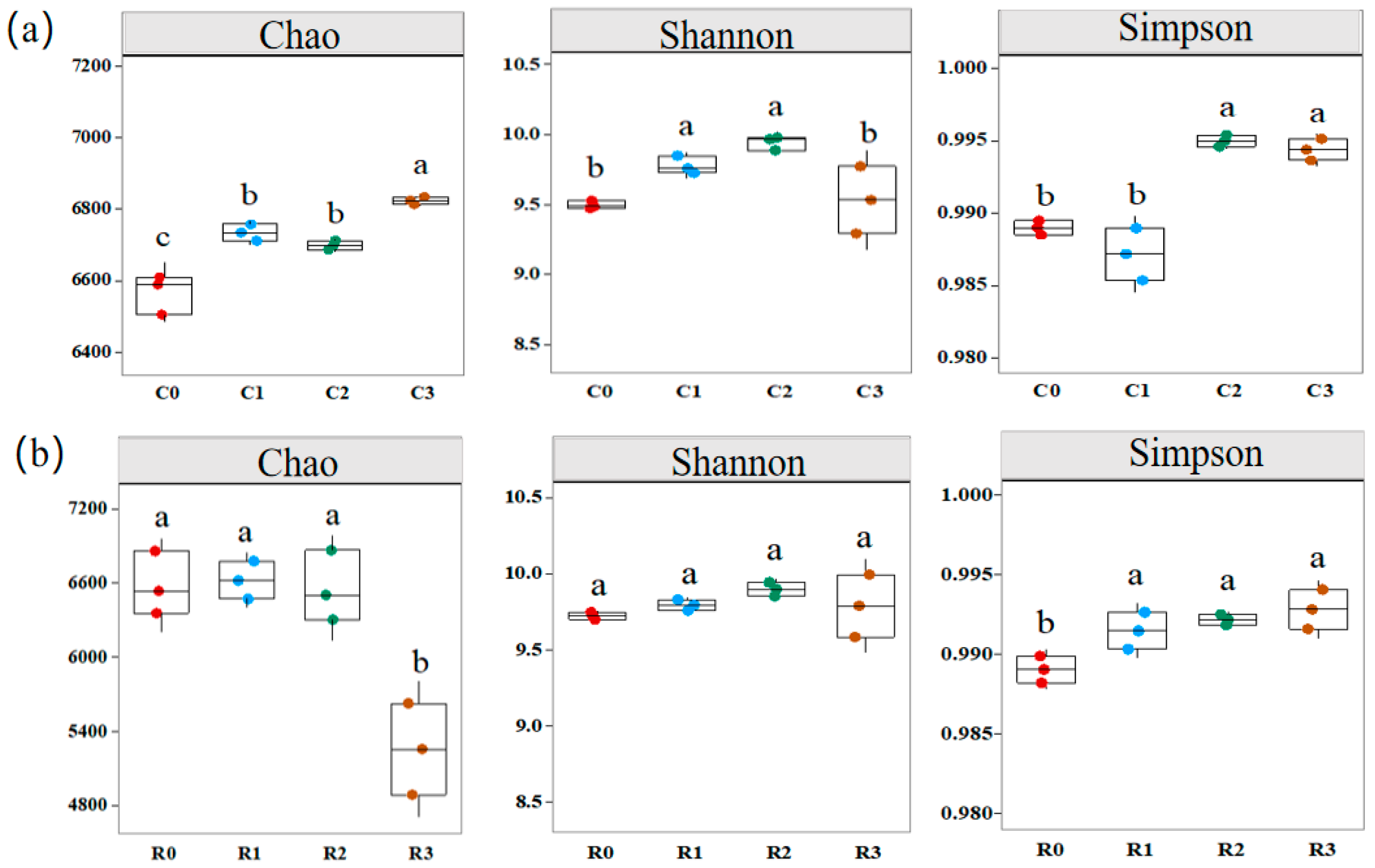
3.5.1. Alpha Diversity
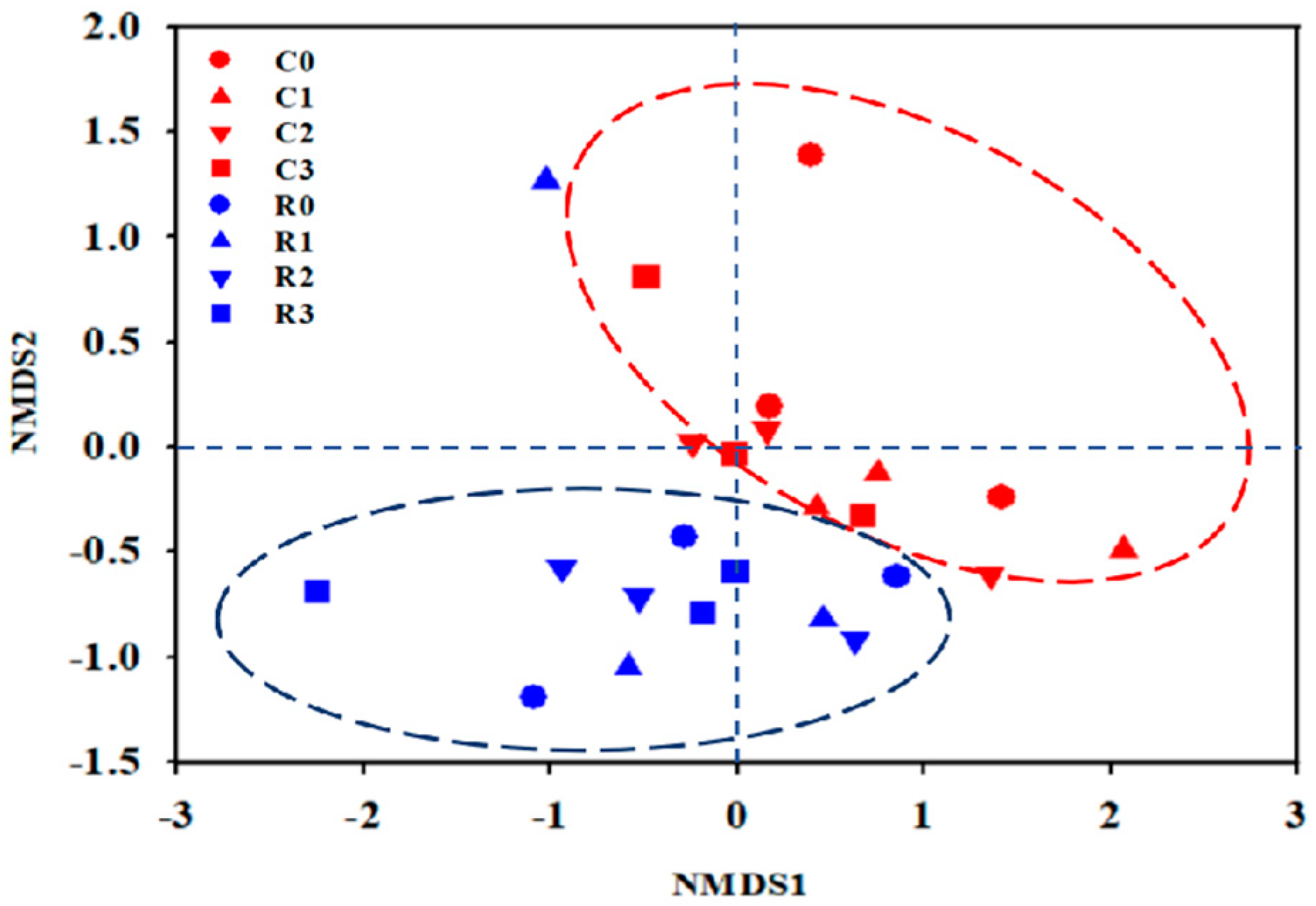
3.5.2. Beta Diversity
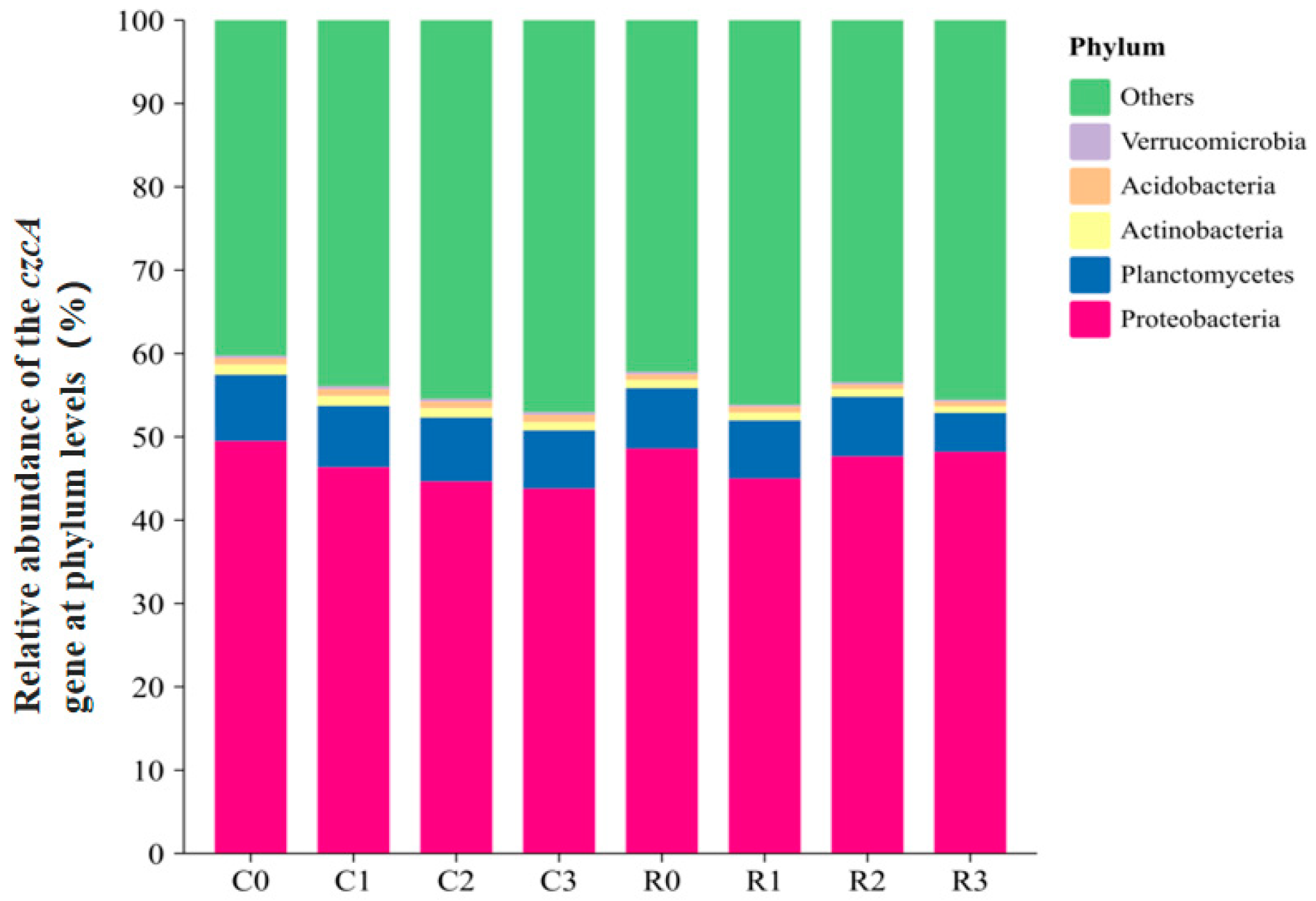
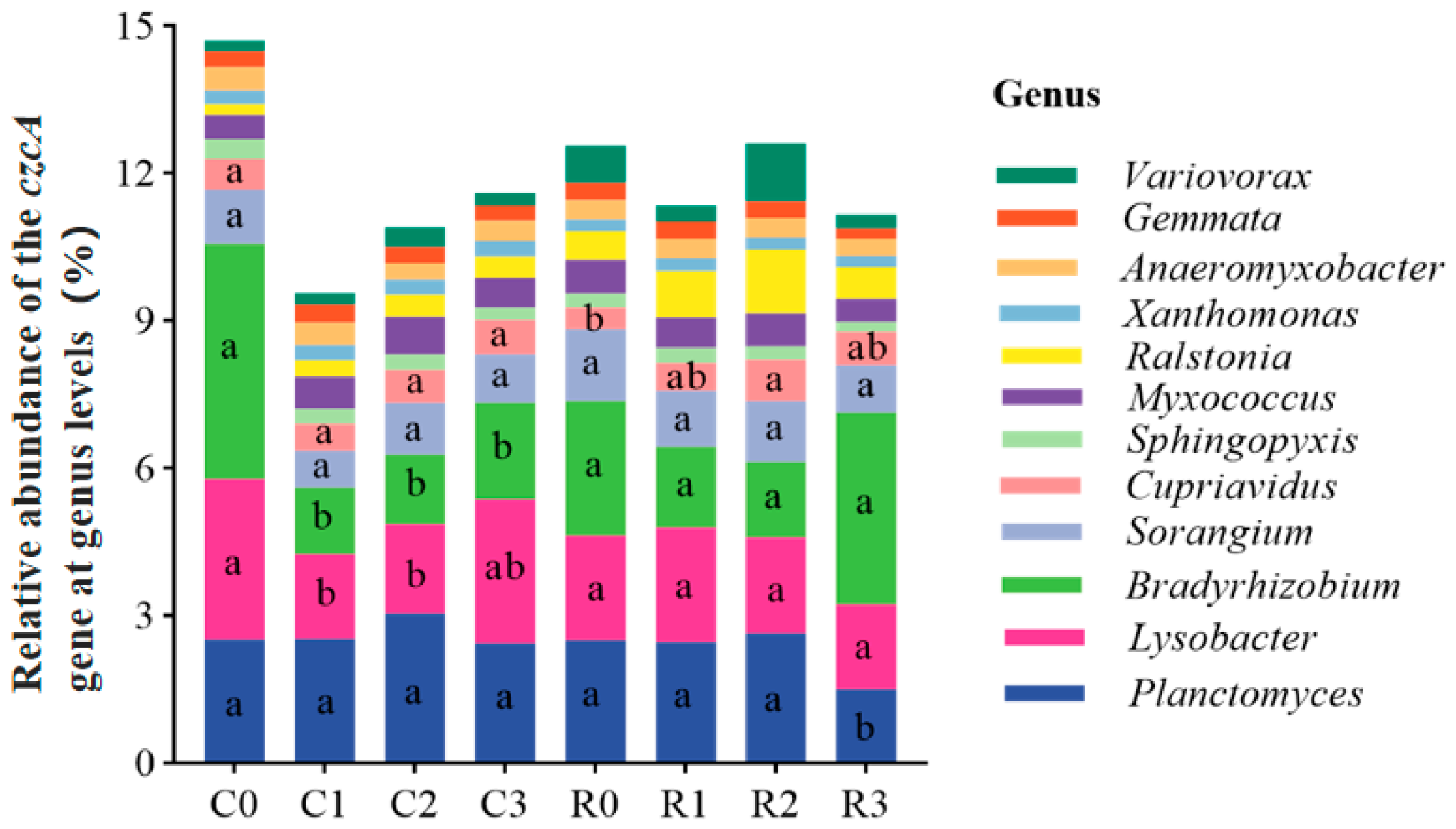
3.5.3. Relative Abundance of the czcA Gene at Phylum and Genus Levels
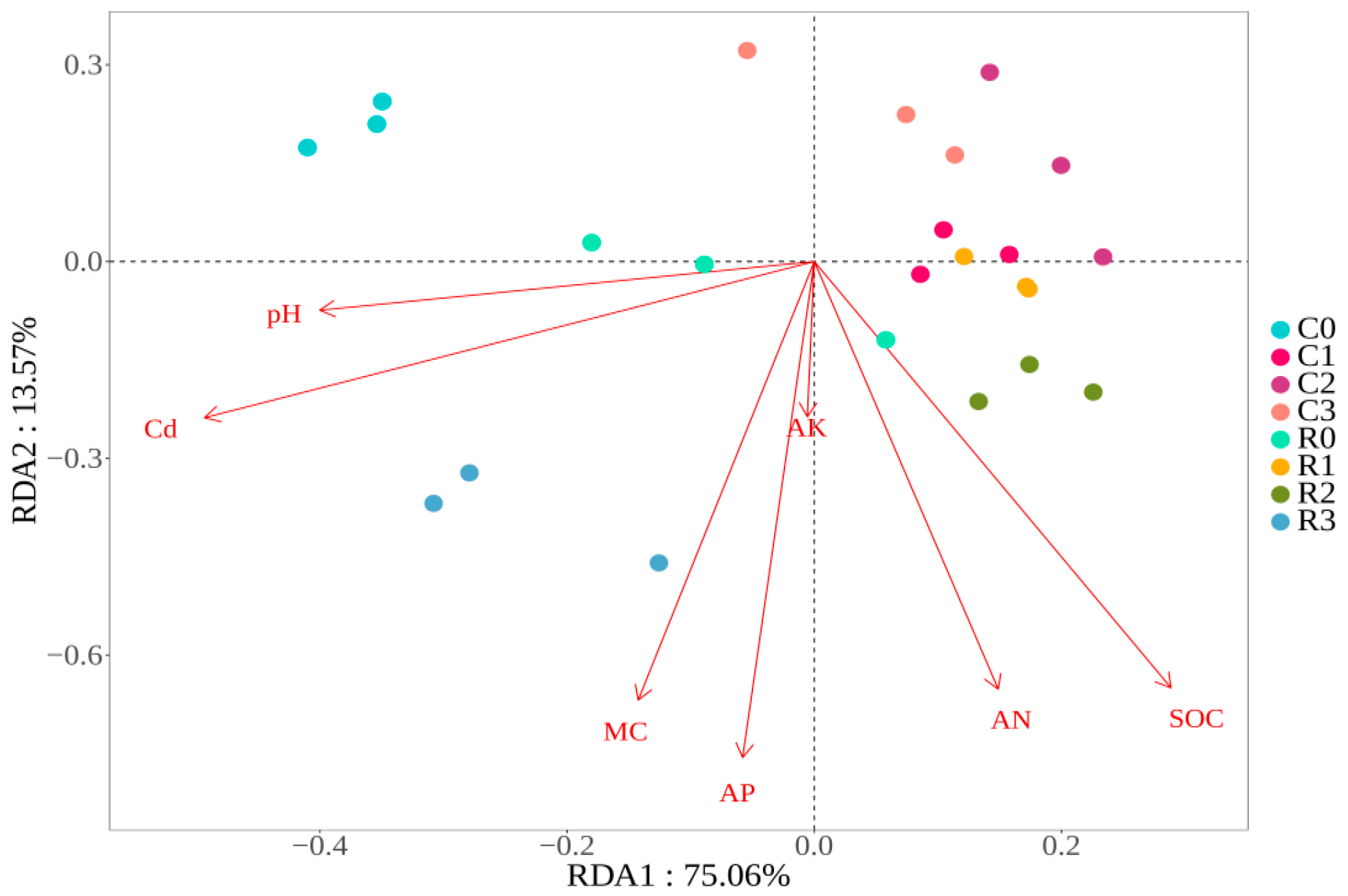
3.6. Redundancy Analysis of czcA Community Structure and Soil Properties
4. Discussion
4.1. Effects of Biochar on Soil Properties
4.2. Effects of Biochar on Cd Contents of Soil and Soybean
4.3. Effects of Biochar on the Expression of the czcA Gene
4.4. Effects of Biochar on Soil Microbial Community Structure of czcA Gene
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Järup, L. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F.; Hannan, F.; Rinklebe, J.; Ok, Y.S. Effect of biochar on cadmium bioavailability and uptake in wheat (Triticum aestivum L.) grown in a soil with aged contamination. Ecotoxicol. Environ. Saf. 2017, 140, 37–47. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Yang, Y.; Li, B.; Wu, Y.; Sun, H.; Yang, Y. Cadmium accumulation characteristics in turnip landraces from China and assessment of their phytoremediation potential for contaminated soils. Front. Plant Sci. 2016, 7, 1862. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ma, Y.; Luo, S. Spatiotemporal evolution and influencing factors of soybean production in Heilongjiang province, China. Land 2023, 12, 2090. [Google Scholar] [CrossRef]
- Wang, X. Characteristic and environmental risk assessment of heavy metals in farmland soil of based on specia-tion analysis. Inform. Manag. Sci. I 2013, 204, 213–220. [Google Scholar] [CrossRef]
- Xu, L.; Jin, S.; Su, Y.; Lyu, X.; Yan, S.; Wang, C.; Cao, L.; Yan, C.; Ma, C. Combined metagenomics and metabolomic analysis of microbial community structure and metabolic function in continuous soybean crop soils of Songnen Plain, China. Chem. Biol. Technol. Agric. 2024, 11, 46. [Google Scholar] [CrossRef]
- Aschi, A.; Aubert, M.; Riah-Anglet, W.; Nélieu, S.; Dubois, C.; Akpa-Vincentas, M.; Trinsoutrot-Gattin, I. Introduction of Faba bean in crop rotation: Impacts on soil chemical and biological characteristics. Appl. Soil Ecol. 2017, 120, 219–228. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Zhang, H.; Shi, W.; Liu, Y. Bacterial heavy-metal and antibiotic resistance genes in a copper tailing dam area in northern China. Front. Microbiol. 2019, 10, 1916. [Google Scholar] [CrossRef]
- Geisseler, D.; Linquist, B.A.; Lazicki, P.A. Effect of fertilization on soil microorganisms in paddy rice systems–a meta-analysis. Soil Biol. Biochem. 2017, 115, 452–460. [Google Scholar] [CrossRef]
- Xin, Y.; Liu, M.; Wei, L.; Gao, Y.; Ruan, Y.; Wang, Q.; Zhang, Z. Changes in soil chemical properties and rhizosphere bacterial community induced by soil amendments associated with reduction in cadmium accumulation by rice. Agronomy 2023, 13, 3051. [Google Scholar] [CrossRef]
- Guo, J.; Yan, C.; Luo, Z.; Fang, H.; Hu, S.; Cao, Y. Synthesis of a novel ternary HA/Fe-Mn oxides-loaded biochar composite and its application in cadmium(II) and arsenic(V) adsorption. J. Environ. Sci. 2019, 85, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xu, R.; Sun, X.; Han, F.; Xiao, E.; Chen, L.; Qiu, L.; Sun, W. Microbiome–environment interactions in antimony-contaminated rice paddies and the correlation of core microbiome with arsenic and antimony contamination. Chemosphere 2021, 263, 128227. [Google Scholar] [CrossRef] [PubMed]
- Gillard, B.; Chatzievangelou, D.; Thomsen, L.; Ullrich, M.S. Heavy-metal-resistant microorganisms in deep-sea sediments disturbed by mining activity: An application toward the development of experimental in vitro systems. Front. Mar. Sci. 2019, 6, 462. [Google Scholar] [CrossRef]
- Xie, X.; Yuan, K.; Chen, X.; Zhao, Z.; Huang, Y.; Hu, L.; Liu, H.; Luan, T.; Chen, B. Characterization of metal resistance genes carried by waterborne free-living and particle-attached bacteria in the Pearl River estuary. Environ. Pollut. 2023, 327, 121547. [Google Scholar] [CrossRef]
- Hu, H. Faculty opinions recommendation of Antibiotic resistomes in plant microbiomes. Fac. Opin.—Post-Publ. Peer Rev. Biomed. Lit. 2019, 24, 530–554. [Google Scholar] [CrossRef]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qiu, J.; Wang, X.; Zhong, Y.; Lan, C.; Shu, W. Cadmium contamination in orchard soils and fruit trees and its potential health risk in Guangzhou, China. Environ. Sci. Pollut. R. 2006, 143, 159–165. [Google Scholar] [CrossRef]
- Visconti, D.; Álvarez-Robles, M.J.; Fiorentino, N.; Fagnano, M.; Clemente, R. Use of Brassica juncea and Dactylis glomerata for the phytostabilization of mine soils amended with compost or biochar. Chemosphere 2020, 260, 127661. [Google Scholar] [CrossRef]
- Singh, R.; Babu, J.N.; Kumar, R.; Srivastava, P.; Singh, P.; Raghubanshi, A.S. Multifaceted application of crop residue biochar as a tool for sustainable agriculture: An ecological perspective. Ecol. Eng. 2015, 77, 324–347. [Google Scholar] [CrossRef]
- Mustafa, A.; Brtnicky, M.; Hammerschmiedt, T.; Kucerik, J.; Kintl, A.; Chorazy, T.; Naveed, M.; Skarpa, P.; Baltazar, T.; Malicek, O.; et al. Food and agricultural wastes-derived biochars in combination with mineral fertilizer as sustainable soil amendments to enhance soil microbiological activity, nutrient cycling and crop production. Front. Plant Sci. 2022, 13, 1028101. [Google Scholar] [CrossRef] [PubMed]
- Amalina, F.; Abd Razak, A.S.; Zularisam, A.W.; Aziz, M.A.A.; Krishnan, S.; Nasrullah, M. Comprehensive assessment of biochar integration in agricultural soil conditioning: Advantages, drawbacks, and future prospectives. Phys. Chem. Earth Parts A/B/C 2023, 132, 103508. [Google Scholar] [CrossRef]
- Liang, Y.; Cao, X.; Zhao, L.; Arellano, E. Biochar- and phosphate-induced immobilization of heavy metals in contaminated soil and water: Implication on simultaneous remediation of contaminated soil and groundwater. Environ. Sci. Pollut. Res. 2014, 21, 4665–4674. [Google Scholar] [CrossRef] [PubMed]
- Abou Jaoude, L.; Castaldi, P.; Nassif, N.; Pinna, M.V.; Garau, G. Biochar and compost as gentle remediation options for the recovery of trace elements-contaminated soils. Sci. Total. Environ. 2020, 711, 134511. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ni, X.; Huang, Q.; Liu, D.; Ye, Z. Effect of bamboo biochar on reducing grain cadmium content in two contrasting wheat genotypes. Environ. Sci. Pollut. Res. 2021, 28, 17405–17416. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, H.; Yang, K.; Er Ze, L.; Lu, Z.; Li, Y.; Mu, L.; Zhang, N. Reducing Cd and Pb accumulation in potatoes: The role of soil passivators in contaminated mining soils. Life 2024, 14, 1615. [Google Scholar] [CrossRef]
- Xing, D.; Cheng, H.; Ning, Z.; Liu, Y.; Lin, S.; Li, Y.; Wang, X.; Hill, P.; Chadwick, D.; Jones, D.L. Field aging declines the regulatory effects of biochar on cadmium uptake by pepper in the soil. J. Environ. Manag. 2022, 321, 115832. [Google Scholar] [CrossRef]
- Yang, X.; Liu, J.; McGrouther, K.; Huang, H.; Lu, K.; Guo, X.; He, L.; Lin, X.; Che, L.; Ye, Z.; et al. Effect of biochar on the extractability of heavy metals (Cd, Cu, Pb, and Zn) and enzyme activity in soil. Environ. Sci. Pollut. Res. 2016, 23, 974–984. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Yuan, H.M.; Xue, W.; Roiloa, S.; Yao, J.; Yu, F.H. Increasing biochar diversity promotes the impacts of plant diversity on remediating cadmium in soil. J. Plant Ecol. 2024, 17, rtae068. [Google Scholar] [CrossRef]
- Walker, N.J. Real-time and quantitative PCR: Applications to mechanism-based toxicology. J. biochem. Mol. Toxicol. 2001, 15, 121–127. [Google Scholar] [CrossRef]
- Hou, X.; He, W.; Zhang, Y.; Zhang, N.; Yan, J.; Chen, Y. Maize-straw biochar enhances soil properties and grain yield of foxtail millet in a newly reclaimed land. Agronomy 2024, 14, 2465. [Google Scholar] [CrossRef]
- Malghani, S.; Kim, J.; Lee, S.-H.; Yoo, G.-y.; Kang, H. Application of two contrasting rice-residue-based biochars triggered gaseous loss of nitrogen under denitrification-favoring conditions: A short-term study based on acetylene inhibition technique. Appl. Soil Ecol. 2018, 127, 112–119. [Google Scholar] [CrossRef]
- Fan, S.; Zuo, J.; Dong, H. Changes in soil properties and bacterial community composition with biochar amendment after six years. Agronomy 2020, 10, 746. [Google Scholar] [CrossRef]
- Houssou, A.A.; Jeyakumar, P.; Niazi, N.K.; Van Zwieten, L.; Li, X.; Huang, L.; Wei, L.; Zheng, X.; Huang, Q.; Huang, Y.; et al. Biochar and soil properties limit the phytoavailability of lead and cadmium by Brassica chinensis L. in contaminated soils. Biochar 2022, 4, 5. [Google Scholar] [CrossRef]
- Oladele, S.O. Changes in physicochemical properties and quality index of an Alfisol after three years of rice husk biochar amendment in rainfed rice—Maize cropping sequence. Geoderma 2019, 353, 359–371. [Google Scholar] [CrossRef]
- Futa, B.; Oleszczuk, P.; Andruszczak, S.; Kwiecińska-Poppe, E.; Kraska, P. Effect of natural aging of biochar on soil enzymatic activity and physicochemical properties in long-term field experiment. Agronomy 2020, 10, 449. [Google Scholar] [CrossRef]
- Xiu, L.; Zhang, W.; Sun, Y.; Wu, D.; Meng, J.; Chen, W. Effects of biochar and straw returning on the key cultivation limitations of Albic soil and soybean growth over 2 years. Catena 2019, 173, 481–493. [Google Scholar] [CrossRef]
- Ma, X.; Liu, Z.; Liu, Z.; Xie, G.; Rookes, J.; An, F. Root secretion of oxalic and malic acids mitigates the rubber tree aluminum toxicity. J. Rubber Res. 2021, 24, 381–390. [Google Scholar] [CrossRef]
- Yu, F.; Zhao, S.; Zhao, Y.; Wang, Y.; Zhai, C.; Zhong, R.; Zhang, J.; Meng, Q. Long-term cattle manure application to saline-sodic soil increases maize yield by decreasing key obstacle factors in the black soil region of Northeastern China. Int. J. Agric. Biol. Eng. 2023, 16, 176–183. [Google Scholar] [CrossRef]
- Huang, X.F.; Li, S.Q.; Li, S.Y.; Ye, G.Y.; Lu, L.J.; Zhang, L.; Yang, L.Y.; Qian, X.; Liu, J. The effects of biochar and dredged sediments on soil structure and fertility promote the growth, photosynthetic and rhizosphere microbial diversity of Phragmites communis (Cav.) Trin. ex Steud. Sci. Total. Environ. 2019, 697, 134073. [Google Scholar] [CrossRef]
- Naeem, M.B.; Jahan, S.; Rashid, A.; Shah, A.A.; Raja, V.; El-Sheikh, M.A. Improving maize yield and drought tolerance in field conditions through activated biochar application. Sci. Rep. 2024, 14, 25000. [Google Scholar] [CrossRef] [PubMed]
- Lemanowicz, J.; Bartkowiak, A.; Lamparski, R.; Wojewódzki, P.; Pobereżny, J.; Wszelaczyńska, E.; Szczepanek, M. Physicochemical and enzymatic soil properties influenced by cropping of primary wheat under organic and conventional farming systems. Agronomy 2020, 10, 1652. [Google Scholar] [CrossRef]
- Haddad, S.A.; Lemanowicz, J. Benefits of corn-cob biochar to the microbial and enzymatic activity of soybean plants grown in soils contaminated with heavy metals. Energies 2021, 14, 5763. [Google Scholar] [CrossRef]
- Han, J.; Wu, D.; Yang, J.; Shi, Y.; Abid, G.; Wang, L.; Li, Z. A biochar-based amendment improved cadmium (Cd) immobilization, reduced its bioaccumulation, and increased rice yield. Front. Env. Sci. 2024, 12, 1487190. [Google Scholar] [CrossRef]
- Lang, Q.; Xia, Y.; Li, Y.; Wang, C.; Liu, Z.; Zou, G.; Sun, Q. CaO-modified hydrochar reduces soil cadmium bioavailability by altering soil properties, shifting bacterial community, and promoting microbial metabolisms. Environ. Technol. Innov. 2024, 35, 103698. [Google Scholar] [CrossRef]
- Wang, X.; Ding, S.; Cheng, T.; Wang, S.; Qing, Z.; Hu, J.; Wang, X. Mechanism of Cd adsorption by sulphur modified biochar and its application in Cd-contaminated soil. Pol. J. Environ. Stud. 2024, 33, 5901–5912. [Google Scholar] [CrossRef]
- Tian, Y.; Cui, L.; Lin, Q.; Li, G.; Zhao, X. The sewage sludge biochar at low pyrolysis temperature had better improvement in urban soil and turf grass. Agronomy 2019, 9, 156. [Google Scholar] [CrossRef]
- Jin, J.; Li, Y.; Zhang, J.; Wu, S.; Cao, Y.; Liang, P.; Zhang, J.; Wong, M.H.; Wang, M.; Shan, S.; et al. Influence of pyrolysis temperature on properties and environmental safety of heavy metals in biochars derived from municipal sewage sludge. J. Hazard. Mater. 2016, 320, 417–426. [Google Scholar] [CrossRef]
- Mohamed, I.; Zhang, G.S.; Li, Z.G.; Liu, Y.; Chen, F.; Dai, K. Ecological restoration of an acidic Cd contaminated soil using bamboo biochar application. Ecol. Eng. 2015, 84, 67–76. [Google Scholar] [CrossRef]
- Irfan, M.; Mudassir, M.; Khan, M.J.; Dawar, K.M.; Muhammad, D.; Mian, I.A.; Ali, W.; Fahad, S.; Saud, S.; Hayat, Z.; et al. Heavy metals immobilization and improvement in maize (Zea mays L.) growth amended with biochar and compost. Sci. Rep. 2021, 11, 18416. [Google Scholar] [CrossRef]
- Haider, F.U.; Coulter, J.A.; Cheema, S.A.; Farooq, M.; Wu, J.; Zhang, R.; Guo, S.J.; Cai, L.Q. Co-application of biochar and microorganisms improves soybean performance and remediate cadmium-contaminated soil. Ecotoxicol. Environ. Saf. 2021, 214, 112112. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhou, M.; Zhang, H.; Yuan, L.; Lv, P.; Wang, L.; Zhang, J. Changes in Cd forms and Cd resistance genes in municipal sludge during coupled earthworm and biochar composting. Ecotoxicol. Environ. Saf. 2024, 286, 117–179. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Xue, X.M.; Kappler, A.; Rosen, B.P.; Meharg, A.A. Linking genes to microbial biogeochemical cycling: Lessons from arsenic. Environ. Sci. Technol. 2017, 51, 7326–7339. [Google Scholar] [CrossRef]
- Khan, Z.; Nisar, M.A.; Hussain, S.Z.; Arshad, M.N.; Rehman, A. Cadmium resistance mechanism in Escherichia coli P4 and its potential use to bioremediate environmental cadmium. Appl. Microbiol. Biotechnol. 2015, 99, 10745–10757. [Google Scholar] [CrossRef]
- Liu, K.; Sun, M.; Ye, M.; Chao, H.; Zhao, Y.; Xia, B.; Jiao, W.; Feng, Y.; Zheng, X.; Liu, M.; et al. Coexistence and association between heavy metals, tetracycline and corresponding resistance genes in vermicomposts originating from different substrates. Environ. Pollut. 2019, 244, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; He, L.; Chen, J.; Lyu, H.; Wang, Y.; Yang, L.; Yang, S.; Liu, Y. Long-term successive biochar amendments alter the composition and α-diversity of bacterial community of paddy soil in rice-wheat rotation. Front. Environ. Sci. 2022, 10, 921766. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Marris, E. Putting the carbon back: Black is the new green. Nature 2006, 442, 7103. [Google Scholar] [CrossRef]
- Yan, T.; Xue, J.; Zhou, Z.; Wu, Y. Biochar-based fertilizer amendments improve the soil microbial community structure in a karst mountainous area. Sci. Total Environ. 2021, 794, 148757. [Google Scholar] [CrossRef]
- Wang, P.; Yan, S.; Zhang, W.; Xie, X.; Li, M.; Ren, T.; Gu, L.; Zhang, Z. Effect of soil management systems on the rhizosphere bacterial community structure of tobacco: Continuous cropping vs. paddy-upland rotation. Ecol. Eng. 2022, 13, 996858. [Google Scholar] [CrossRef]
- Semenov, M.V.; Zhelezova, A.D.; Ksenofontova, N.A.; Ivanova, E.A.; Nikitin, D.A.; Semenov, V.M. Microbiological indicators for assessing the effects of agricultural practices on soil health: A review. Agronom. 2025, 15, 335. [Google Scholar] [CrossRef]
- Gao, W.; Gao, K.; Guo, Z.; Liu, Y.; Jiang, L.; Liu, C.; Liu, X.; Wang, G. Different responses of soil bacterial and fungal communities to 3 years of biochar amendment in an alkaline soybean soil. Front. Microbiol. 2021, 12, 630418. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Yuan, P.; Ullah, S.; Iqbal, A.; Zhao, Q.; Liang, H.; Khan, A.; Imran; Zhang, H.; Wu, X.; et al. Biochar amendment and nitrogen fertilizer contribute to the changes in soil properties and microbial communities in a paddy field. Front. Microbiol. 2022, 13, 834751. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Xu, H.; Huang, Y.; Xiong, X.; Zhu, H.; Lin, J.; Shi, J.; Tang, C.; Xu, J. Changes in soil Cd contents and microbial communities following Cd-containing straw return. Environ. Pollut. 2023, 330, 121753. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | MC (%) | pH | AN (mg·kg−1) | AP (mg·kg−1) | AK (mg·kg−1) | SOC (mg·kg−1) |
|---|---|---|---|---|---|---|
| C0 | 16.50 ± 0.05 a | 8.44 ± 0.01 a | 85.58 ± 0.52 c | 7.48 ± 0.30 d | 303.00 ± 2.00 c | 26.51 ± 0.06 d |
| C1 | 16.52 ± 0.05 a | 8.43 ± 0.03 a | 92.05 ± 1.93 b | 9.43 ± 0.09 c | 284.50 ± 1.50 d | 27.59 ± 0.07 c |
| C2 | 16.59 ± 0.10 a | 8.37 ± 0.01 b | 98.79 ± 3.41 a | 14.76 ± 0.13 b | 356.00 ± 3.00 b | 29.00 ± 0.03 b |
| C3 | 16.46 ± 0.11 a | 8.42 ± 0.02 a | 101.50 ± 4.03 a | 18.15 ± 1.00 a | 401.67 ± 4.73 a | 29.24 ± 0.07 a |
| R0 | 16.32 ± 0.20 b | 8.38 ± 0.01 b | 117.54 ± 3.14 b | 15.33 ± 0.09 d | 344.67 ± 12.66 b | 29.46 ± 0.02 b |
| R1 | 17.00 ± 0.01 ab | 8.38 ± 0.04 b | 124.78 ± 2.45 a | 22.83 ± 0.13 c | 327.00 ± 3.46 c | 29.58 ± 0.04 b |
| R2 | 16.79 ± 0.15 ab | 8.41 ± 0.02 ab | 122.27 ± 1.24 ab | 35.36 ± 0.95 b | 357.67 ± 4.04 b | 30.87 ± 0.09 a |
| R3 | 17.47 ± 0.11 a | 8.43 ± 0.01 a | 126.00 ± 3.21 a | 41.56 ± 1.52 a | 397.67 ± 3.51 a | 30.93 ± 0.15 a |
| Soil Properties | Mantel R Statistic | p-Value |
|---|---|---|
| MC | 0.329 * | 0.014 |
| pH | 0.129 | 0.218 |
| AN | 0.298 * | 0.017 |
| AP | 0.396 * | 0.006 |
| AK | 0.038 | 0.653 |
| SOC | 0.334 * | 0.016 |
| Cd | 0.235 * | 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Q.; He, G.; Yan, S.; Song, J.; Zhang, Y.; Zhang, X.; Sun, Y.; Wang, Y.; Li, Y.; Tian, Y.; et al. Changes in Soil Properties, Content of Cd, and Cd-Resistant Bacterial Community with Biochar After One-Time Addition Under Soybean Continuous Cropping and Crop Rotation Patterns. Agronomy 2025, 15, 1084. https://doi.org/10.3390/agronomy15051084
Yao Q, He G, Yan S, Song J, Zhang Y, Zhang X, Sun Y, Wang Y, Li Y, Tian Y, et al. Changes in Soil Properties, Content of Cd, and Cd-Resistant Bacterial Community with Biochar After One-Time Addition Under Soybean Continuous Cropping and Crop Rotation Patterns. Agronomy. 2025; 15(5):1084. https://doi.org/10.3390/agronomy15051084
Chicago/Turabian StyleYao, Qin, Gege He, Shubo Yan, Jie Song, Yuetong Zhang, Xiaoyu Zhang, Yan Sun, Yibo Wang, Yongjuan Li, Yu Tian, and et al. 2025. "Changes in Soil Properties, Content of Cd, and Cd-Resistant Bacterial Community with Biochar After One-Time Addition Under Soybean Continuous Cropping and Crop Rotation Patterns" Agronomy 15, no. 5: 1084. https://doi.org/10.3390/agronomy15051084
APA StyleYao, Q., He, G., Yan, S., Song, J., Zhang, Y., Zhang, X., Sun, Y., Wang, Y., Li, Y., Tian, Y., Zhao, C., & Guo, Y. (2025). Changes in Soil Properties, Content of Cd, and Cd-Resistant Bacterial Community with Biochar After One-Time Addition Under Soybean Continuous Cropping and Crop Rotation Patterns. Agronomy, 15(5), 1084. https://doi.org/10.3390/agronomy15051084