
Virus–Host Interactions and Genetic Exchange in Mixed Infections of Tomato Yellow Leaf Curl Virus (TYLCV), Tomato Leaf Curl New Delhi Virus (ToLCNDV), and Tomato Chlorosis Virus (ToCV)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Periods
2.2. Whiteflies, Virus Isolates, and Virus Inoculation
2.3. Symptom Evaluation and Virus Detection by Tissue Blot Hybridization
2.4. Detection of Recombination Events in Mixed Infections by HTS
2.5. Cloning and Sequencing of def-DNA and Recombinant Molecules
2.6. Statistical Analyses
3. Results
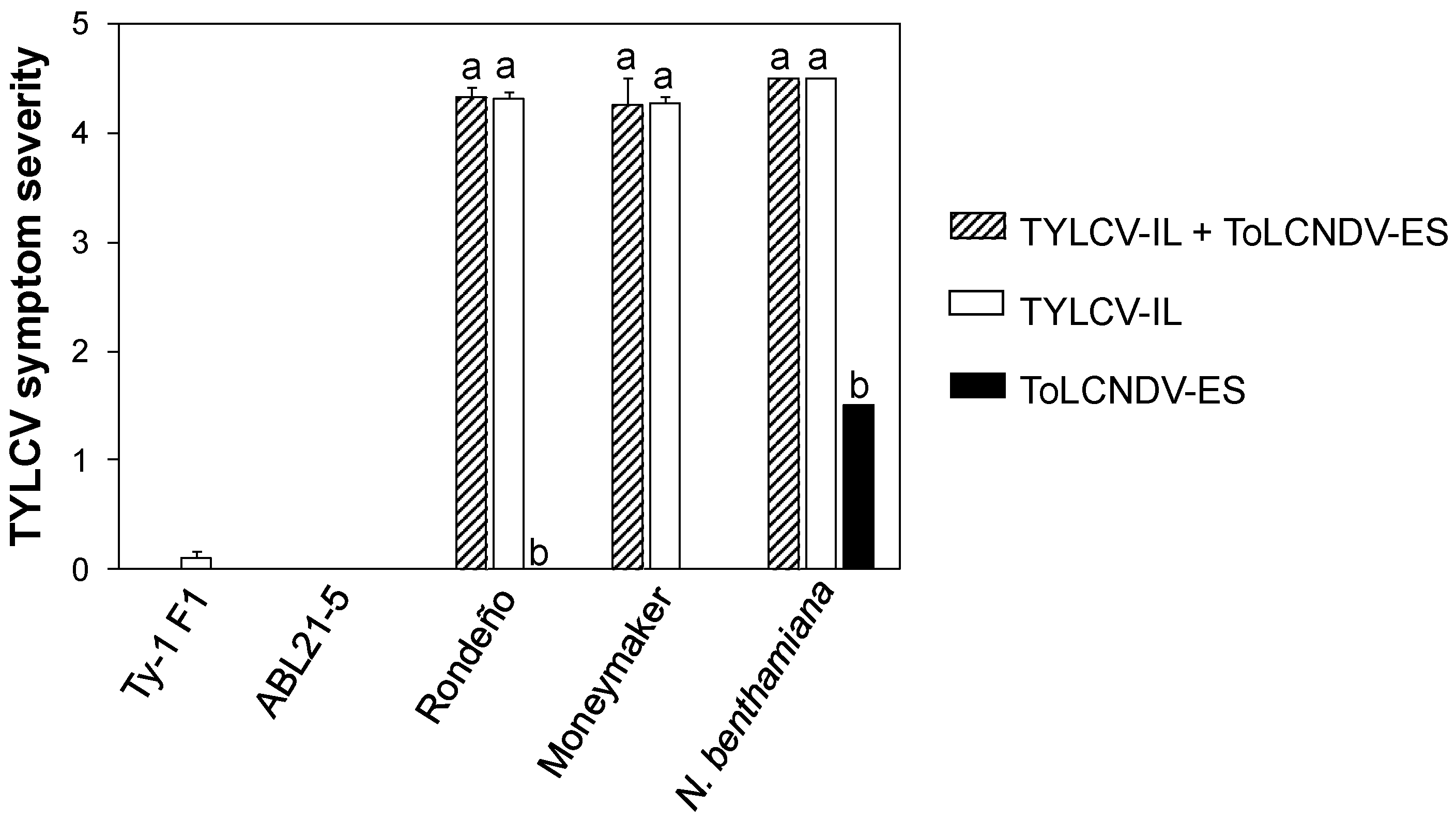
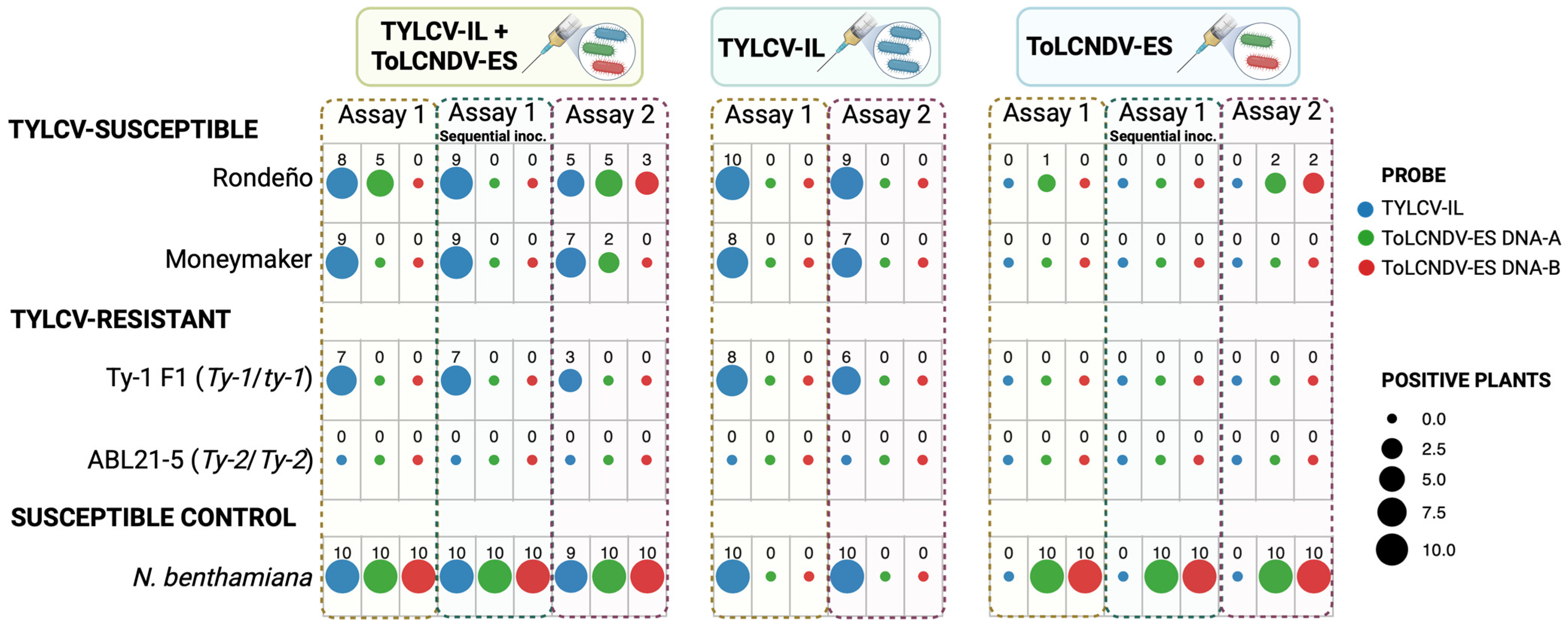
3.1. ToLCNDV-ES and TYLCV-IL Co-Infection in TYLCV-Susceptible, TYLCV-Resistant Tomato Plants and N. benthamiana
3.2. Absence of Heterologous Trans-Replication of ToLCNDV-ES DNA-B by TYLCV-IL
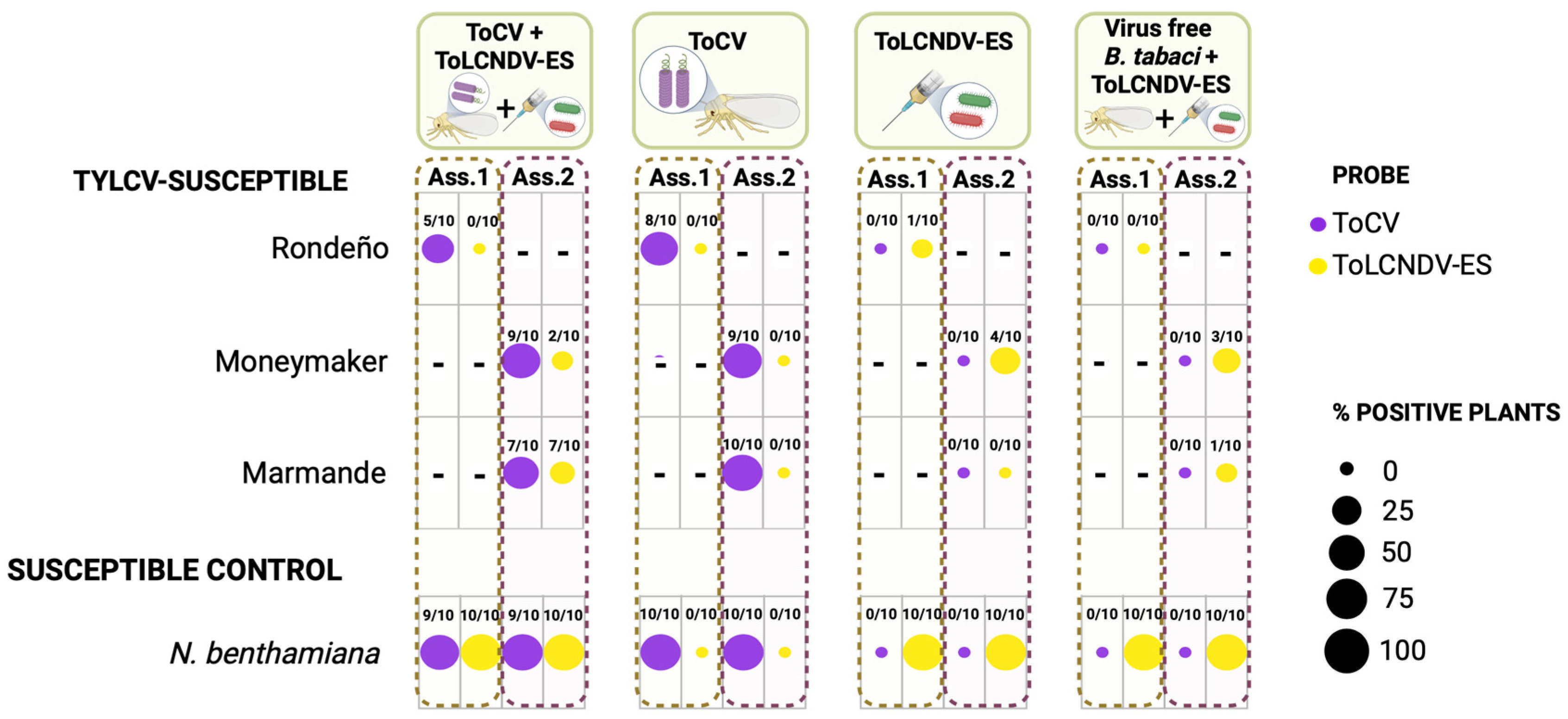
3.3. Lack of Enhancement of ToLCNDV-ES Infection in Tomato Plants by the Crinivirus ToCV
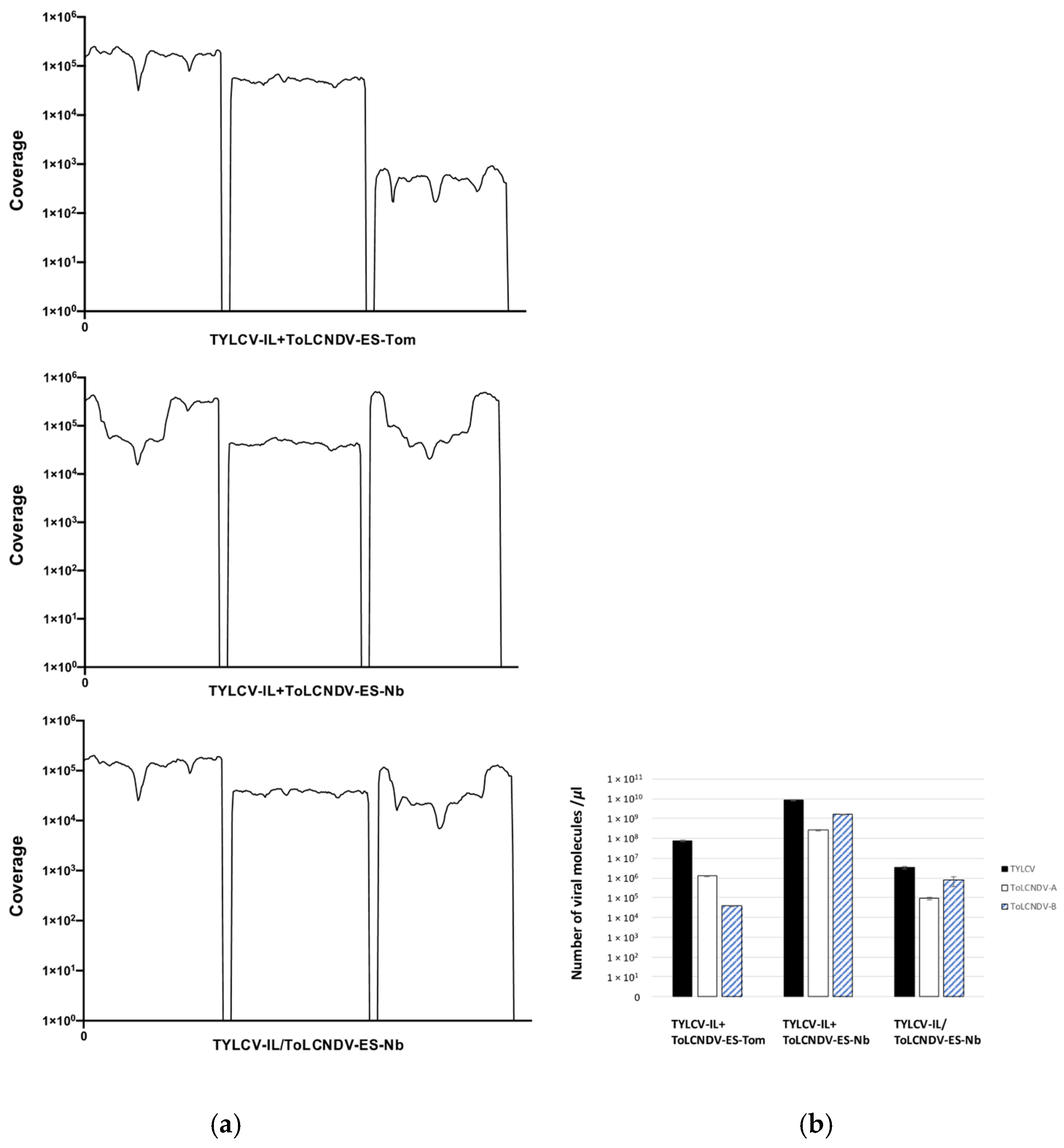
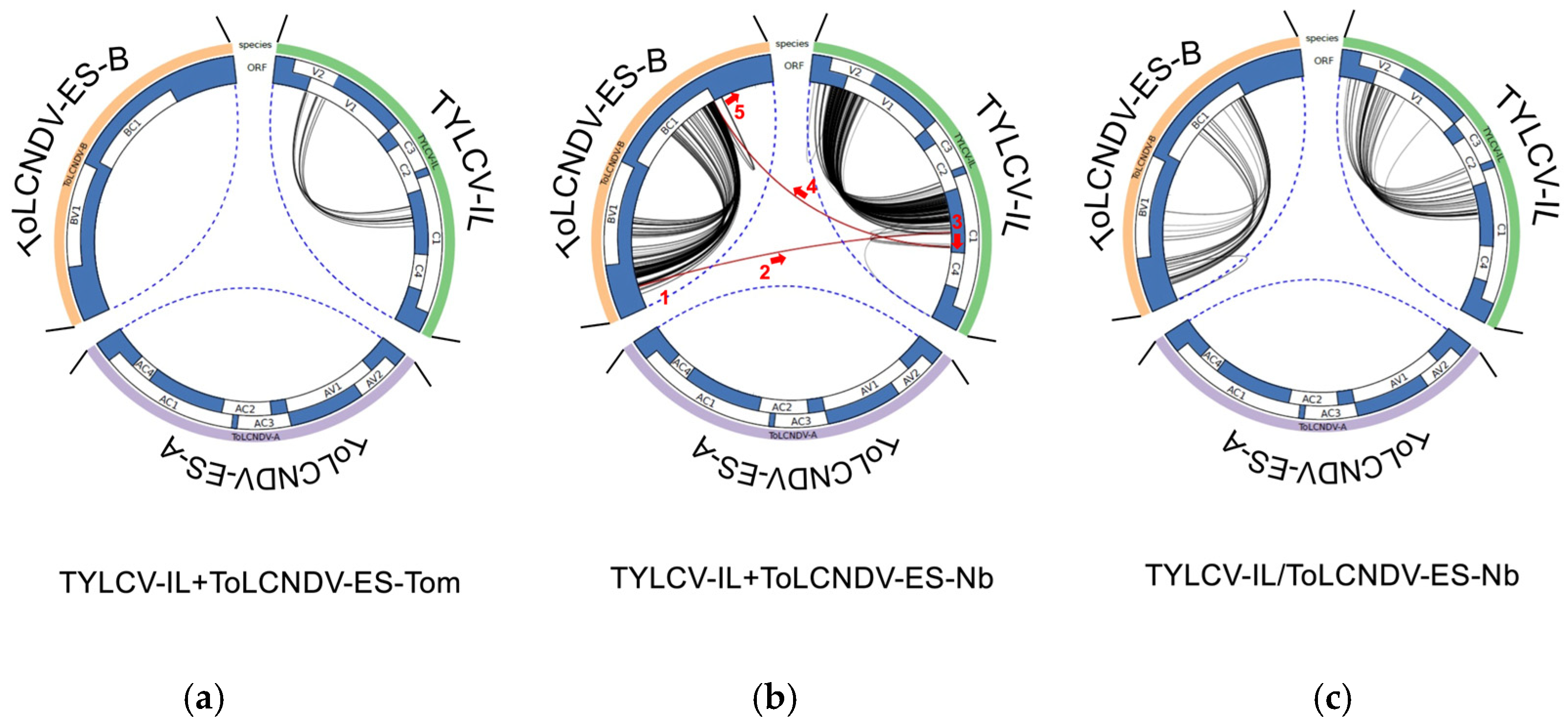
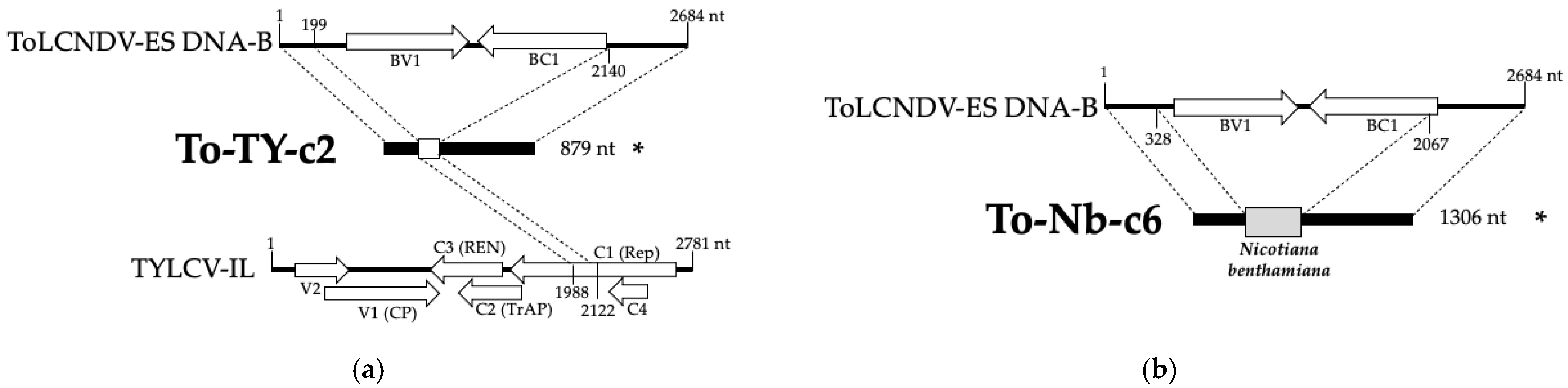
3.4. Recombination Events Between TYLCV-IL and ToLCNDV-ES in Mixed-Infected Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rybicki, E.P.; Pietersen, G. Plant virus disease problems in the developing world. Adv. Virus Res. 1999, 53, 127–175. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Lopez-Moya, J.J.; Aranda, M.A. Whitefly-transmitted RNA viruses that affect intensive vegetable production. Ann. Appl. Biol. 2014, 165, 155–171. [Google Scholar] [CrossRef]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Rivera Bustamante, R.F.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef] [PubMed]
- Dalton, R. Whitefly infestations: The Christmas Invasion. Nature 2006, 443, 898–900. [Google Scholar] [CrossRef]
- De Barro, P.J. Bemisia tabaci, the capacity to invade. In The Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants; Thompson, W.M.O., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 181–204. [Google Scholar]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef]
- Hanssen, I.M.; Lapidot, M.; Thomma, B.P.H.J. Emerging Viral Diseases of Tomato Crops. Mol. Plant-Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef]
- Moriones, E.; Praveen, S.; Chakraborty, S. Tomato leaf curl New Delhi virus: An emerging virus complex threatening vegetable and fiber crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Tomato chlorosis virus, an emergent plant virus still expanding its geographical and host ranges. Mol. Plant Pathol. 2019, 20, 1307–1320. [Google Scholar] [CrossRef]
- Tatineni, S.; Hein, G.L. Plant viruses of agricultural importance: Current and future perspectives of virus disease management strategies. Phytopathology 2023, 113, 117–141. [Google Scholar] [CrossRef]
- Lefeuvre, P.; Martin, D.P.; Harkins, G.; Lemey, P.; Gray, A.J.A.; Meredith, S.; Lakay, F.; Monjane, A.; Lett, J.-M.; Varsani, A.; et al. The spread of tomato yellow leaf curl virus from the Middle East to the world. PLoS Pathog. 2010, 6, e1001164. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Lett, J.-M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV Virus Taxonomy Profile: Geminiviridae 2021. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 105–140. [Google Scholar] [CrossRef]
- Arguello-Astorga, G.R.; Ruiz-Medrano, R. An iteron-related domain is associated to Motif 1 in the replication proteins of geminiviruses: Identification of potential interacting amino acid-base pairs by a comparative approach. Arch. Virol. 2001, 146, 1465–1485. [Google Scholar] [CrossRef]
- Gong, P.; Tan, H.; Zhao, S.; Li, H.; Liu, H.; Ma, Y.; Zhang, X.; Rong, J.; Fu, X.; Lozano-Durán, R.; et al. Geminiviruses encode additional small proteins with specific subcellular localizations and virulence function. Nat. Commun. 2021, 12, 4278. [Google Scholar] [CrossRef]
- Zhao, S.; Gong, P.; Ren, Y.; Liu, H.; Li, H.; Li, F.; Zhou, X. The novel C5 protein from tomato yellow leaf curl virus is a virulence factor and suppressor of gene silencing. Stress Biol. 2022, 2, 19. [Google Scholar] [CrossRef]
- Liu, H.; Chang, Z.; Zhao, S.; Gong, P.; Zhang, M.; Lozano-Durán, R.; Yan, H.; Zhou, X.; Li, F. Functional identification of a novel C7 protein of tomato yellow leaf curl virus. Virology 2023, 585, 117–126. [Google Scholar] [CrossRef]
- Patil, B.; Dasgupta, I. Defective interfering DNAs of plant viruses. Crit. Rev. Plant Sci. 2006, 25, 47–64. [Google Scholar] [CrossRef]
- Shahid, M.S.; Ito, T.; Kimbara, J.; Onozato, A.; Natsuaki, K.T.; Ikegami, M. Evaluation of tomato hybrids carrying Ty-1 and Ty-2 loci to Japanese monopartite begomovirus species. J. Phytopathol. 2013, 161, 205–209. [Google Scholar] [CrossRef]
- Prasanna, H.C.; Sinha, D.P.; Rai, G.K.; Krishna, R.; Kashyap, S.P.; Singh, N.K.; Singh, M.; Malathi, V.G. Pyramiding Ty-2 and Ty-3 genes for resistance to monopartite and bipartite tomato leaf curl viruses of India. Plant Pathol. 2015, 64, 256–264. [Google Scholar] [CrossRef]
- Albuquerque, L.; Varsani, A.; Fernandes, F.; Pinheiro, B.; Martin, D.; de Tarso Oliveira Ferreira, P.; Lemos, T.O.; Inoue-Nagata, A.K. Further characterization of tomato-infecting begomoviruses in Brazil. Arch. Virol. 2012, 157, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Tiendrebeogo, F.; Lefeuvre, P.; Hoareau, M.; Harimalala, M.; De Bruyn, A.; Villemot, J.V.; Traoré, V.S.E.; Konaté, G.; Traoré, A.S.; Barro, N.; et al. Evolution of African cassava mosaic virus by recombination between bipartite and monopartite begomoviruses. Virol. J. 2012, 9, 67. [Google Scholar] [CrossRef]
- Vanitharani, R.; Chellappan, P.; Pita, J.S.; Fauquet, C.M. Differential roles of AC2 and AC4 of cassava geminiviruses in mediating synergism and suppression of posttranscriptional gene silencing. J. Virol. 2004, 78, 9487–9498. [Google Scholar] [CrossRef]
- Singh, A.K.; Kushwaha, N.; Chakraborty, S. Synergistic interaction among begomoviruses leads to the suppression of host defense-related gene expression and breakdown of resistance in chilli. Appl. Microbiol. Biotechnol. 2016, 100, 4035–4049. [Google Scholar] [CrossRef] [PubMed]
- Voorburg, C.M.; Yan, Z.; Bergua-Vidal, M.; Wolters, A.-M.A.; Bai, Y.; Kormelink, R. Ty-1, a universal resistance gene against geminiviruses that is compromised by co-replication of a betasatellite. Mol. Plant Pathol. 2020, 21, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Monci, F.; Sanchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef]
- Davino, S.; Miozzi, L.; Panno, S.; Rubio, L.; Davino, M.; Accotto, G.P. Recombination profiles between Tomato yellow leaf curl virus and Tomato yellow leaf curl Sardinia virus in laboratory and field conditions: Evolutionary and taxonomic implications. J. Gen. Virol. 2012, 93, 2712–2717. [Google Scholar] [CrossRef]
- Díaz-Pendón, J.; Sánchez-Campos, S.; Fortes, I.; Moriones, E. Tomato yellow leaf curl Sardinia virus, a begomovirus species evolving by mutation and recombination: A challenge for virus control. Viruses 2019, 11, 45. [Google Scholar] [CrossRef]
- Chakraborty, S.; Vanitharani, R.; Chattopadhyay, B.; Fauquet, C.M. Supervirulent pseudorecombination and asymmetric synergism between genomic components of two distinct species of begomovirus associated with severe tomato leaf curl disease in India. J. Gen. Virol. 2008, 89, 818–828. [Google Scholar] [CrossRef]
- Vuillaume, F.; Thebaud, G.; Urbino, C.; Forfert, N.; Granier, M.; Froissart, R.; Blanc, S.; Peterschmitt, M. Distribution of the phenotypic effects of random homologous recombination between two virus species. PLoS Pathog. 2011, 7, e1002028. [Google Scholar] [CrossRef]
- Paz-Carrasco, L.C.; Castillo-Urquiza, G.P.; Lima, A.T.; Xavier, C.A.; Vivas-Vivas, L.M.; Mizubuti, E.S.; Zerbini, F.M. Begomovirus diversity in tomato crops and weeds in Ecuador and the detection of a recombinant isolate of rhynchosia golden mosaic Yucatan virus infecting tomato. Arch. Virol. 2014, 159, 2127–2132. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, J.-c.; Ding, T.-b.; Chu, D. Synergistic effects of a Tomato chlorosis virus and Tomato yellow leaf curl virus mixed infection on host tomato plants and the whitefly vector. Front. Plant Sci. 2021, 12, 672400. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros, I.; López-Moya, J.J.; Díaz-Pendón, J.A. Coinfection of tomato plants with tomato yellow leaf curl virus and tomato chlorosis virus affects the interaction with host and whiteflies. Phytopathology 2022, 112, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Pita, J.S.; Fondong, V.N.; Sangare, A.; Otim-Nape, G.W.; Ogwal, S.; Fauquet, C.M. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef]
- Kumari, P.; Singh, A.K.; Sharma, V.K.; Chattopadhyay, B.; Chakraborty, S. A novel recombinant tomato-infecting begomovirus capable of transcomplementing heterologous DNA-B components. Arch. Virol. 2011, 156, 769–783. [Google Scholar] [CrossRef]
- Fortes, M.I.; Sanchez-Campos, S.; Fiallo-Olive, E.; Diaz-Pendón, J.A.; Navas-Castillo, J.; Moriones, E. A novel strain of tomato leaf curl New Delhi virus has spread to the Mediterranean basin. Viruses 2016, 8, 307. [Google Scholar] [CrossRef]
- Ruiz, L.; Simon, A.; Velasco, L.; Janssen, D. Biological characterization of Tomato leaf curl New Delhi virus from Spain. Plant Pathol. 2017, 66, 376–382. [Google Scholar] [CrossRef]
- Vo, T.T.B.; Troiano, E.; Lal, A.; Hoang, P.T.; Kil, E.-J.; Lee, S.; Parrella, G. ToLCNDV-ES infection in tomato is enhanced by TYLCV: Evidence from field survey and agroinoculation. Front. Microbiol. 2022, 13, 954460. [Google Scholar] [CrossRef]
- Morilla, G.; Janssen, D.; Garcia-Andres, S.; Moriones, E.; Cuadrado, I.M.; Bejarano, E.R. Pepper (Capsicum annuum) is a dead-end host for Tomato yellow leaf curl virus. Phytopathology 2005, 95, 1089–1097. [Google Scholar] [CrossRef]
- Monci, F.; Garcia-Andres, S.; Maldonado, J.A.; Moriones, E. Resistance to monopartite begomoviruses associated with the bean leaf crumple disease in Phaseolus vulgaris controlled by a single dominant gene. Phytopathology 2005, 95, 819–826. [Google Scholar] [CrossRef]
- Garcia-Cano, E.; Navas-Castillo, J.; Moriones, E.; Fernandez-Munoz, R. Resistance to Tomato chlorosis virus in Wild Tomato Species that Impair Virus Accumulation and Disease Symptom Expression. Phytopathology 2010, 100, 582–592. [Google Scholar] [CrossRef]
- Friedmann, M.; Lapidot, M.; Cohen, S.; Pilowsky, M. A novel source of resistance to tomato yellow leaf curl virus exhibiting a symptomless reaction to viral infection. J. Am. Soc. Hortic. Sci. 1998, 123, 1004–1007. [Google Scholar] [CrossRef]
- Garcia-Cano, E.; Resende, R.O.; Fernandez-Munoz, R.; Moriones, E. Synergistic interaction between Tomato chlorosis virus and Tomato spotted wilt virus results in breakdown of resistance in tomato. Phytopathology 2006, 96, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef]
- Yamamoto, H.; Wakita, Y.; Kitaoka, T.; Fujishiro, T.; Kesumawati, E.; Koeda, S. Southeast Asian isolate of the Tomato leaf curl New Delhi virus shows higher pathogenicity against tomato and cucurbit crops compared to that of the Mediterranean isolate. Hortic. J. 2021, 90, 314–325. [Google Scholar] [CrossRef]
- Sivalingam, P.N.; Varma, A. Role of betasatellite in the pathogenesis of a bipartite begomovirus affecting tomato in India. Arch. Virol. 2012, 157, 1081–1092. [Google Scholar] [CrossRef]
- Seoane, P.; Díaz-Martínez, L.; Viguera, E.; Claros, M.G.; Grande-Pérez, A. QuasiFlow: A bioinformatic tool for genetic variability analysis from next generation sequencing data. bioRxiv 2022. [Google Scholar] [CrossRef]
- Inkscape Project. Inkscape [Computer Software]. 2020. Available online: https://inkscape.org (accessed on 3 March 2023).
- Garcia-Andres, S.; Tomas, D.M.; Sanchez-Campos, S.; Navas-Castillo, J.; Moriones, E. Frequent occurrence of recombinants in mixed infections of tomato yellow leaf curl disease-associated begomoviruses. Virology 2007, 365, 210–219. [Google Scholar] [CrossRef]
- Briddon, R.W.; Prescott, A.G.; Lunness, P.; Chamberlin, L.C.L.; Markham, P.G. Rapid production of full-length, infectious geminivirus clones by abutting primer PCR (AbP-PCR). J. Virol. Methods 1993, 43, 7–20. [Google Scholar] [CrossRef]
- Blawid, R.; Van, D.T.; Maiss, E. Transreplication of a Tomato yellow leaf curl Thailand virus DNA-B and replication of a DNA− component by Tomato leaf curl Vietnam virus and Tomato yellow leaf curl Vietnam virus. Virus Res. 2008, 136, 107–117. [Google Scholar] [CrossRef]
- Ouattara, A.; Tiendrébéogo, F.; Becker, N.; Urbino, C.; Thébaud, G.; Hoareau, M.; Allibert, A.; Chiroleu, F.; Vernerey, M.S.; Traoré, E.V.; et al. Synergy between an emerging monopartite begomovirus and a DNA-B component. Sci. Rep. 2022, 12, 695. [Google Scholar] [CrossRef] [PubMed]
- Maliano, M.R.; Rojas, M.R.; Macedo, M.A.; Barboza, N.; Gilbertson, R.L. The invasion biology of tomato begomoviruses in Costa Rica reveals neutral synergism that may lead to increased disease pressure and economic loss. Virus Res. 2022, 317, 198793. [Google Scholar] [CrossRef]
- Nogueira, A.M.; Barbosa, T.M.C.; Quadros, A.F.F.; Orílio, A.F.; Bigão, M.C.J.; Xavier, C.A.D.; Ferro, C.G.; Zerbini, F.M. Specific nucleotides in the common region of the begomovirus tomato rugose mosaic virus (ToRMV) are responsible for the negative interference over tomato severe rugose virus (ToSRV) in mixed infection. Viruses 2023, 15, 2074. [Google Scholar] [CrossRef] [PubMed]
- Mascia, T.; Cillo, F.; Fanelli, V.; Finetti-Sialer, M.M.; De Stradis, A.; Palukaitis, P.; Gallitelli, D. Characterization of the interactions between Cucumber mosaic virus and Potato virus Y in Mixed Infections in Tomato. Mol. Plant-Microbe Interact. 2010, 23, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Luria, N.; Smith, E.; Hadad, L.; Bakelman, E.; Sela, N.; Belausov, E.; Lachman, O.; Leibman, D.; Dombrovsky, A. Tomato brown rugose fruit virus contributes to enhanced Pepino mosaic virus titers in tomato Plants. Viruses 2020, 12, 879. [Google Scholar] [CrossRef]
- Ohnishi, J.; Yamaguchi, H.; Saito, A. Analysis of the Mild strain of tomato yellow leaf curl virus, which overcomes Ty-2 gene-mediated resistance in tomato line H24. Arch. Virol. 2016, 161, 2207–2217. [Google Scholar] [CrossRef]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef]
- Saeed, M.; Zafar, Y.; Randles, J.W.; Rezaian, M.A. A monopartite begomovirus-associated DNA beta satellite substitutes for the DNA B of a bipartite begomovirus to permit systemic infection. J. Gen. Virol. 2007, 88, 2881–2889. [Google Scholar] [CrossRef]
- Simon, A.; Ruiz, L.; Velasco, L.; Janssen, D. Absolute quantification of Tomato leaf curl New Delhi virus Spain strain, ToLCNDV-ES: Virus accumulation in a host-specific manner. Plant Dis. 2017, 102, 165. [Google Scholar] [CrossRef]
- Luna, A.P.; Morilla, G.; Voinnet, O.; Bejarano, E.R. Functional analysis of gene-silencing suppressors from tomato yellow leaf curl disease viruses. Mol. Plant-Microbe Interact. 2012, 25, 1294–1306. [Google Scholar] [CrossRef]
- Gover, O.; Peretz, Y.; Mozes-Koch, R.; Maori, E.; Rabinowitch, H.D.; Sela, I. Only minimal regions of tomato yellow leaf curl virus (TYLCV) are required for replication, expression and movement. Arch. Virol. 2014, 159, 2263–2274. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.T.B.; Lal, A.; Nattanong, B.; Tabassum, M.; Qureshi, M.A.; Troiano, E. Coat protein is responsible for tomato leaf curl New Delhi virus pathogenicity in tomato. Front. Plant Sci. 2023, 14, 1206255. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Navas-Castillo, J. Tomato chlorosis virus, a promiscuous virus with multiple host plants and whitefly vectors. Ann. Appl. Biol. 2022, 182, 29–36. [Google Scholar] [CrossRef]
- Martinez-Zubiaur, Y.; Fiallo-Olive, E.; Carrillo-Tripp, J.; Rivera-Bustamante, R. First report of tomato chlorosis virus Infecting tomato in single and mixed infections with tomato yellow leaf curl virus in Cuba. Plant Dis. 2008, 92, 836. [Google Scholar] [CrossRef]
- Macedo, M.A.; Inoue-Nagata, A.K.; Silva, T.N.Z.; Freitas, D.M.S.; Rezende, J.A.M.; Barbosa, J.C.; Michereff-Filho, M.; Nascimento, A.R.; Lourenção, A.L.; Bergamin Filho, A. Temporal and spatial progress of the diseases caused by the crinivirus Tomato chlorosis virus and the begomovirus Tomato severe rugose virus in tomatoes in Brazil. Plant Pathol. 2019, 68, 72–84. [Google Scholar] [CrossRef]
- Wege, C.; Siegmund, D. Synergism of a DNA and an RNA virus: Enhanced tissue infiltration of the begomovirus Abutilon mosaic virus (AbMV) mediated by Cucumber mosaic virus (CMV). Virology 2007, 357, 10–28. [Google Scholar] [CrossRef]
- Butterbach, P.; Verlaan, M.G.; Dullemans, A.; Lohuis, D.; Visser, R.G.F.; Bai, Y.; Kormelink, R. Tomato yellow leaf curl virus resistance by Ty-1 involves increased cytosine methylation of viral genomes and is compromised by cucumber mosaic virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 12942–12947. [Google Scholar] [CrossRef]
- Landeo-Ríos, Y.; Navas-Castillo, J.; Moriones, E.; Cañizares, M. The heterologous expression of the p22 RNA silencing suppressor of the crinivirus Tomato chlorosis virus from Tobacco rattle virus and potato virus X enhances disease severity but does not complement suppressor-defective mutant viruses. Viruses 2017, 9, 358. [Google Scholar] [CrossRef]
- Saunders, K.; Salim, N.; Mali, V.R.; Malathi, V.G.; Briddon, R.; Markham, P.G.; Stanley, J. Characterisation of Sri Lankan cassava mosaic virus and Indian cassava mosaic virus: Evidence for acquisition of a DNA B component by a monopartite Begomovirus. Virology 2002, 293, 63–74. [Google Scholar] [CrossRef]
- Shafiq, M.; Asad, S.; Zafar, Y.; Briddon, R.W.; Mansoor, S. Pepper leaf curl Lahore virus requires the DNA B component of Tomato leaf curl New Delhi virus to cause leaf curl symptoms. Virol. J. 2010, 7, 367. [Google Scholar] [CrossRef]
- Chakraborty, S.; Pandey, P.K.; Banerjee, M.K.; Kalloo, G.; Fauquet, C.M. Tomato leaf curl Gujarat virus, a new begomovirus species causing a severe leaf curl disease of tomato in Varanasi, India. Phytopathology 2003, 93, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Carter, S.A.; Cole, A.B.; Cheng, N.H.; Nelson, R.S. A natural variant of a host RNA-dependent RNA polymerase is associated with increased susceptibility to viruses by Nicotiana benthamiana. Proc. Natl. Acad. Sci. USA 2004, 101, 6297–6302. [Google Scholar] [CrossRef]
- Hančinský, R.; Mihálik, D.; Mrkvová, M.; Candresse, T.; Glasa, M. Plant viruses infecting solanaceae family members in the cultivated and wild environments: A review. Plants 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Bellido, A.; Hoyer, J.S.; Dubey, D.; Jeannot, R.B.; Duffy, S. Interspecies recombination has driven the macroevolution of cassava mosaic begomoviruses. J. Virol. 2021, 95, e00541-21. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.B. Defective viral genomes: Critical danger signals of viral infections. J. Virol. 2014, 88, 8720–8723. [Google Scholar] [CrossRef]
- Manzoni, T.B.; Lopez, C.B. Defective (interfering) viral genomes re-explored: Impact on antiviral immunity and virus persistence. Future Virol. 2018, 13, 493–503. [Google Scholar] [CrossRef]
- Catoni, M.; Noris, E.; Vaira, A.M.; Jonesman, T.; Matić, S.; Soleimani, R.; Behjatnia, S.A.A.; Vinals, N.; Paszkowski, J.; Accotto, G.P. Virus-mediated export of chromosomal DNA in plants. Nat. Commun. 2018, 9, 5308. [Google Scholar] [CrossRef]
- Patil, B.L.; Dutt, N.; Briddon, R.W.; Bull, S.E.; Rothenstein, D.; Borah, B.K.; Dasgupta, I.; Stanley, J.; Jeske, H. Deletion and recombination events between the DNA-A and DNA-B components of Indian cassava-infecting geminiviruses generate defective molecules in Nicotiana benthamiana. Virus Res. 2007, 124, 59–67. [Google Scholar] [CrossRef]
- Stanley, J.; Saunders, K.; Pinner, M.S.; Wong, S.M. Novel defective interfering DNAs associated with ageratum yellow vein geminivirus infection of Ageratum conyzoides. Virology 1997, 239, 87–96. [Google Scholar] [CrossRef]
- Hillman, B.I.; Carrington, J.C.; Morris, T.J. A defective interfering RNA that contains a mosaic of a plant virus genome. Cell 1987, 51, 427–433. [Google Scholar] [CrossRef]
- Stanley, J.; Frischmuth, T.; Ellwood, S. Defective viral DNA ameliorates symptoms of geminivirus infection in transgenic plants. Proc. Natl. Acad. Sci. USA 1990, 87, 6291–6295. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fortes, I.M.; Díaz-Martínez, L.; Moriones, E.; Grande-Pérez, A. Virus–Host Interactions and Genetic Exchange in Mixed Infections of Tomato Yellow Leaf Curl Virus (TYLCV), Tomato Leaf Curl New Delhi Virus (ToLCNDV), and Tomato Chlorosis Virus (ToCV). Agronomy 2025, 15, 1006. https://doi.org/10.3390/agronomy15051006
Fortes IM, Díaz-Martínez L, Moriones E, Grande-Pérez A. Virus–Host Interactions and Genetic Exchange in Mixed Infections of Tomato Yellow Leaf Curl Virus (TYLCV), Tomato Leaf Curl New Delhi Virus (ToLCNDV), and Tomato Chlorosis Virus (ToCV). Agronomy. 2025; 15(5):1006. https://doi.org/10.3390/agronomy15051006
Chicago/Turabian StyleFortes, Isabel M., Luis Díaz-Martínez, Enrique Moriones, and Ana Grande-Pérez. 2025. "Virus–Host Interactions and Genetic Exchange in Mixed Infections of Tomato Yellow Leaf Curl Virus (TYLCV), Tomato Leaf Curl New Delhi Virus (ToLCNDV), and Tomato Chlorosis Virus (ToCV)" Agronomy 15, no. 5: 1006. https://doi.org/10.3390/agronomy15051006
APA StyleFortes, I. M., Díaz-Martínez, L., Moriones, E., & Grande-Pérez, A. (2025). Virus–Host Interactions and Genetic Exchange in Mixed Infections of Tomato Yellow Leaf Curl Virus (TYLCV), Tomato Leaf Curl New Delhi Virus (ToLCNDV), and Tomato Chlorosis Virus (ToCV). Agronomy, 15(5), 1006. https://doi.org/10.3390/agronomy15051006