

Evaluation of Commercial Tomato Hybrids for Climate Resilience and Low-Input Farming: Yield and Nutritional Assessment Across Cultivation Systems

 , ,
, ,  , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Methodology
2.2. Traits Evaluated
2.2.1. Descriptive Characteristics
2.2.2. Yield Characteristics
2.2.3. Leaf Nutrient Diagnostics
2.2.4. Fruit Quality and Nutritional Value Characteristics
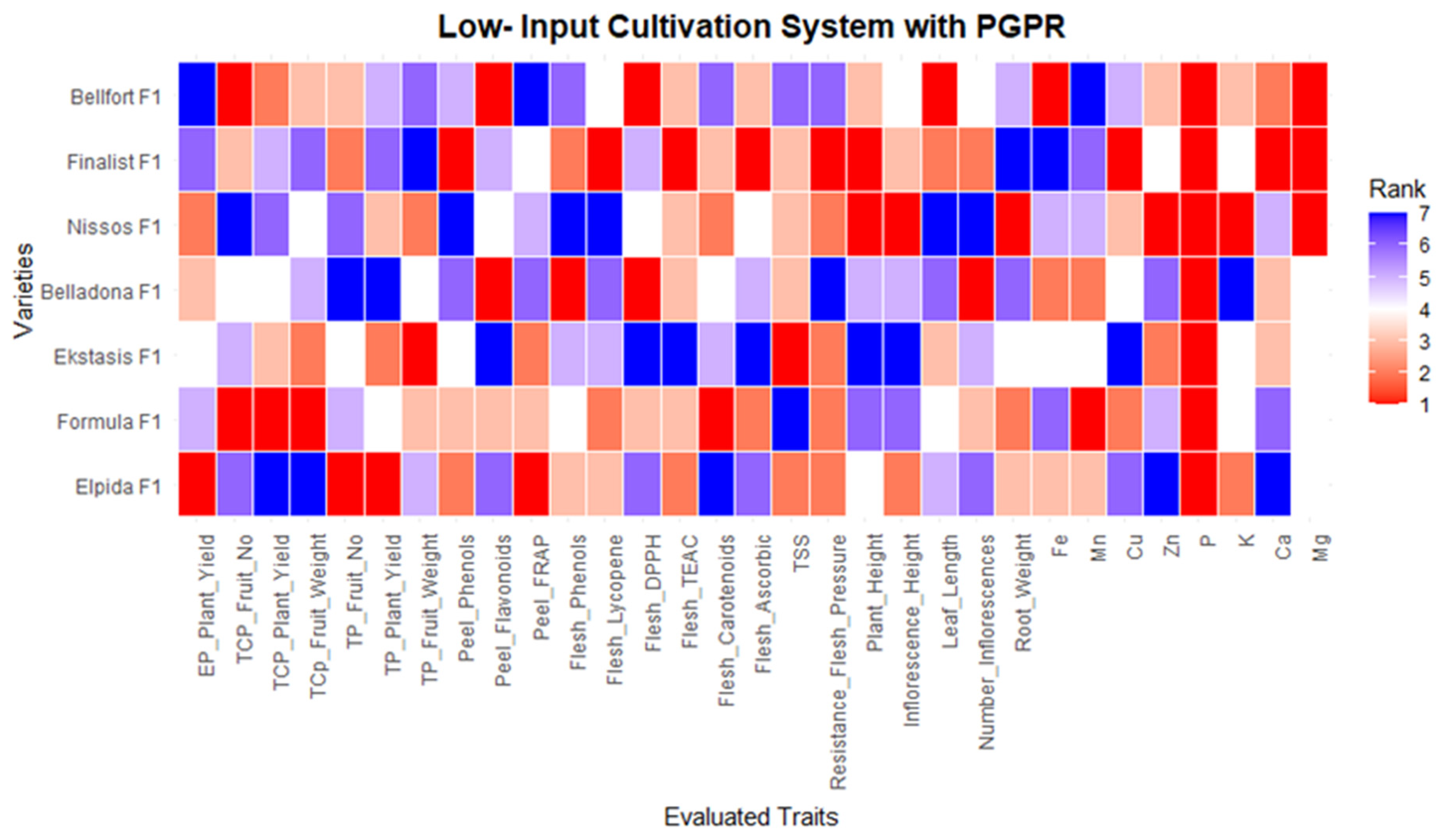
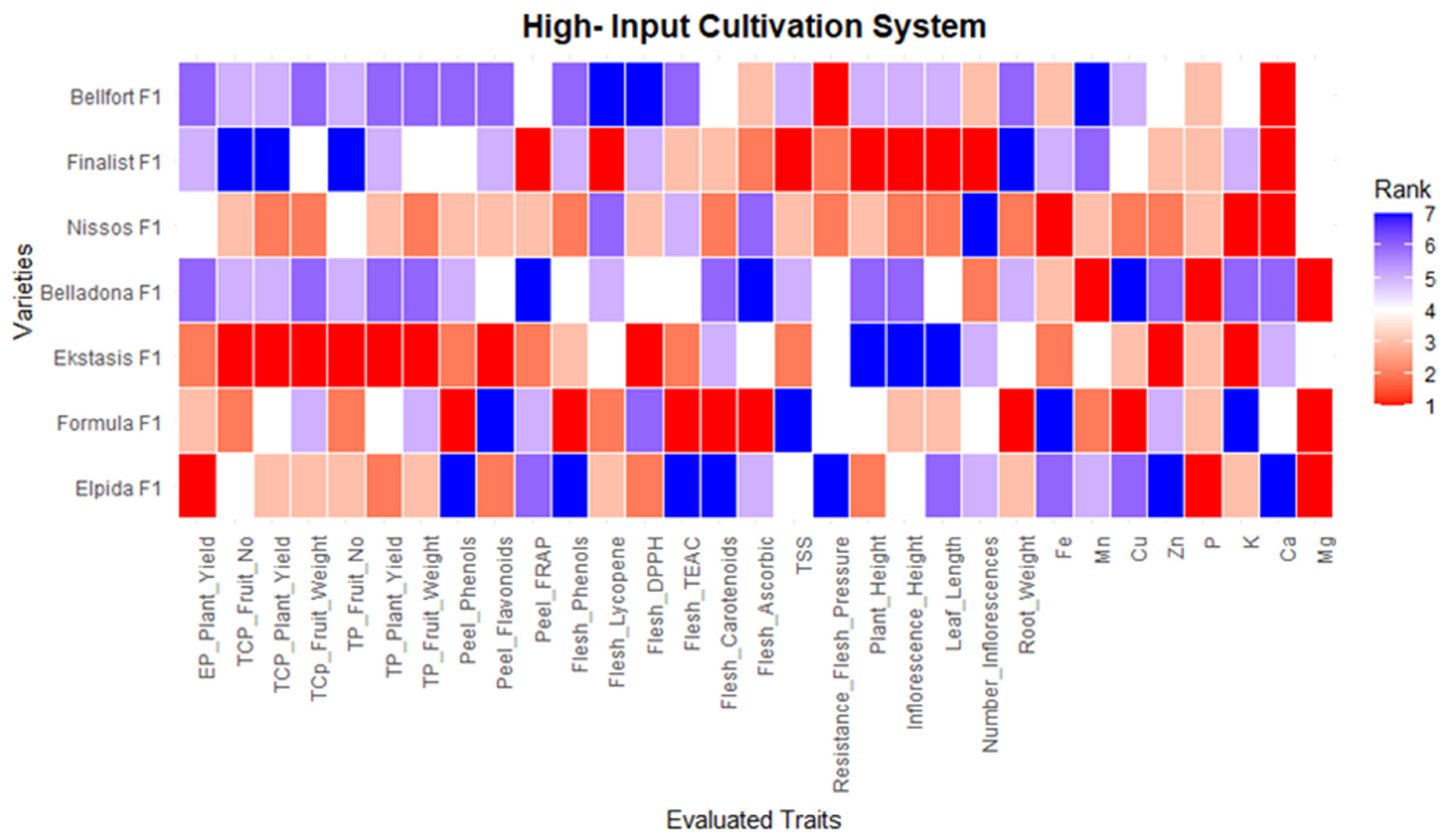
2.3. Statistical Analysis Techno-Economic Analysis and Visualizations
3. Results
3.1. Descriptive Characteristics
3.2. Yield Characteristics
3.3. Leaf Nutrient Diagnostics
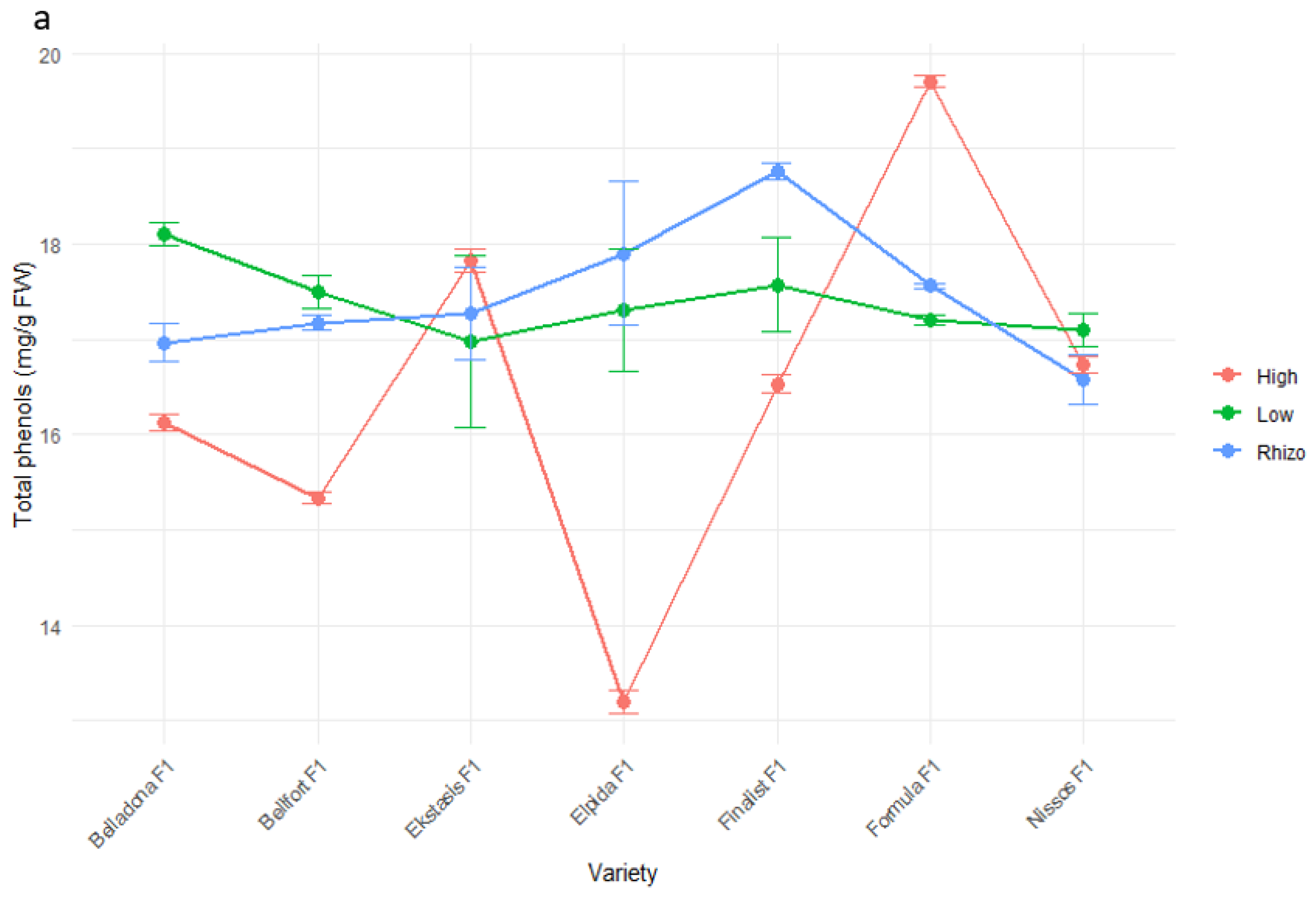
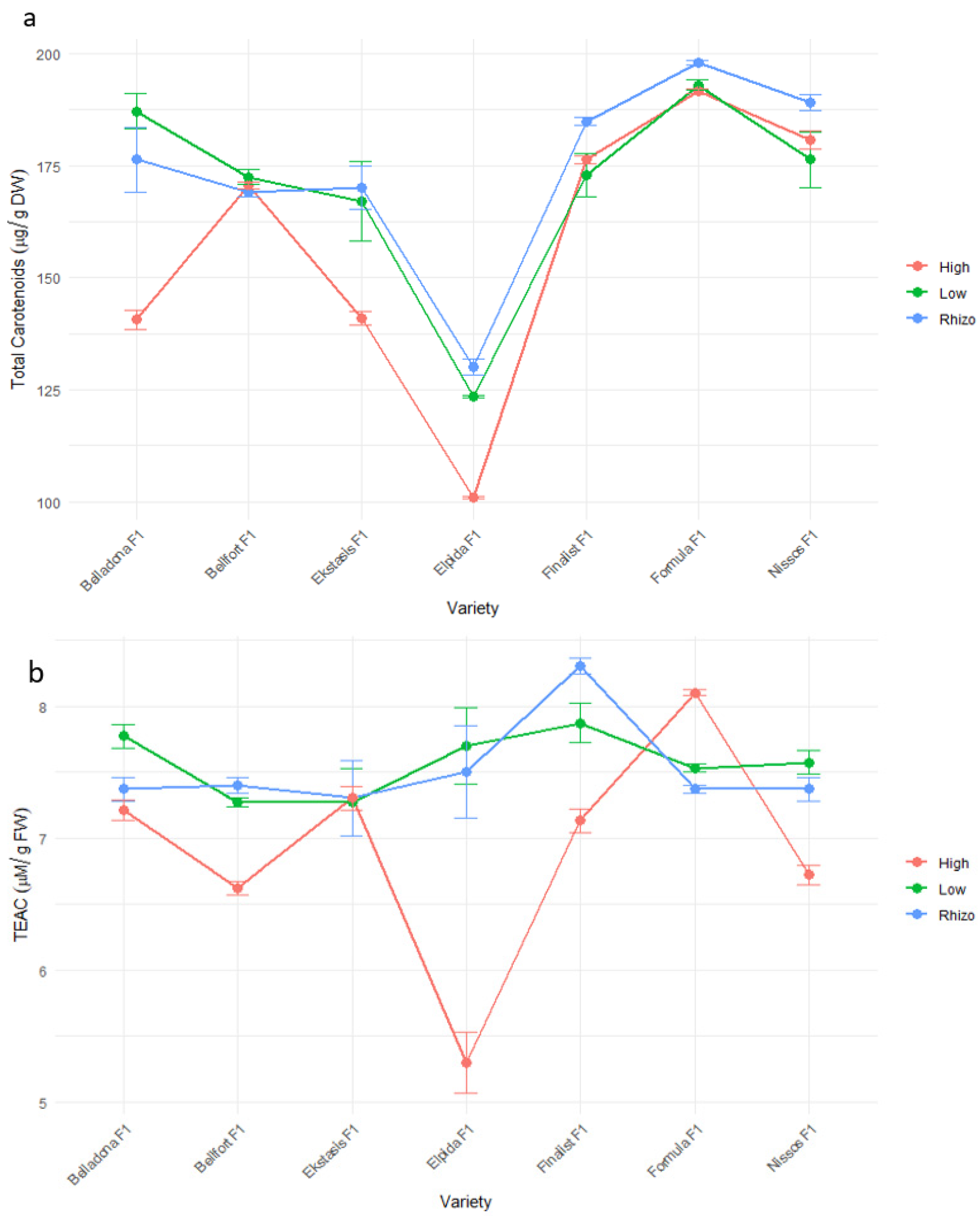
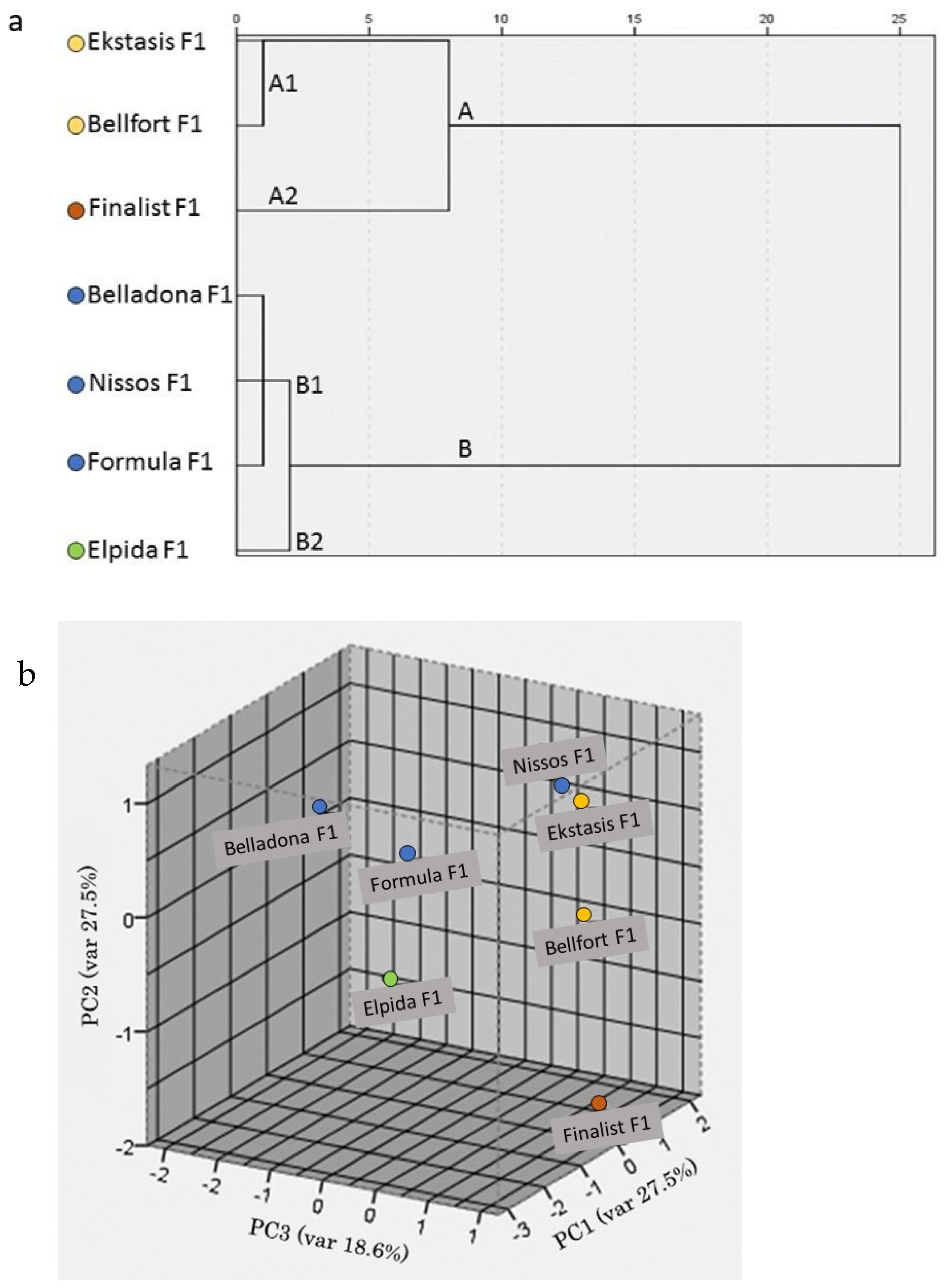
3.4. Fruit Quality and Nutritional Value Characteristics
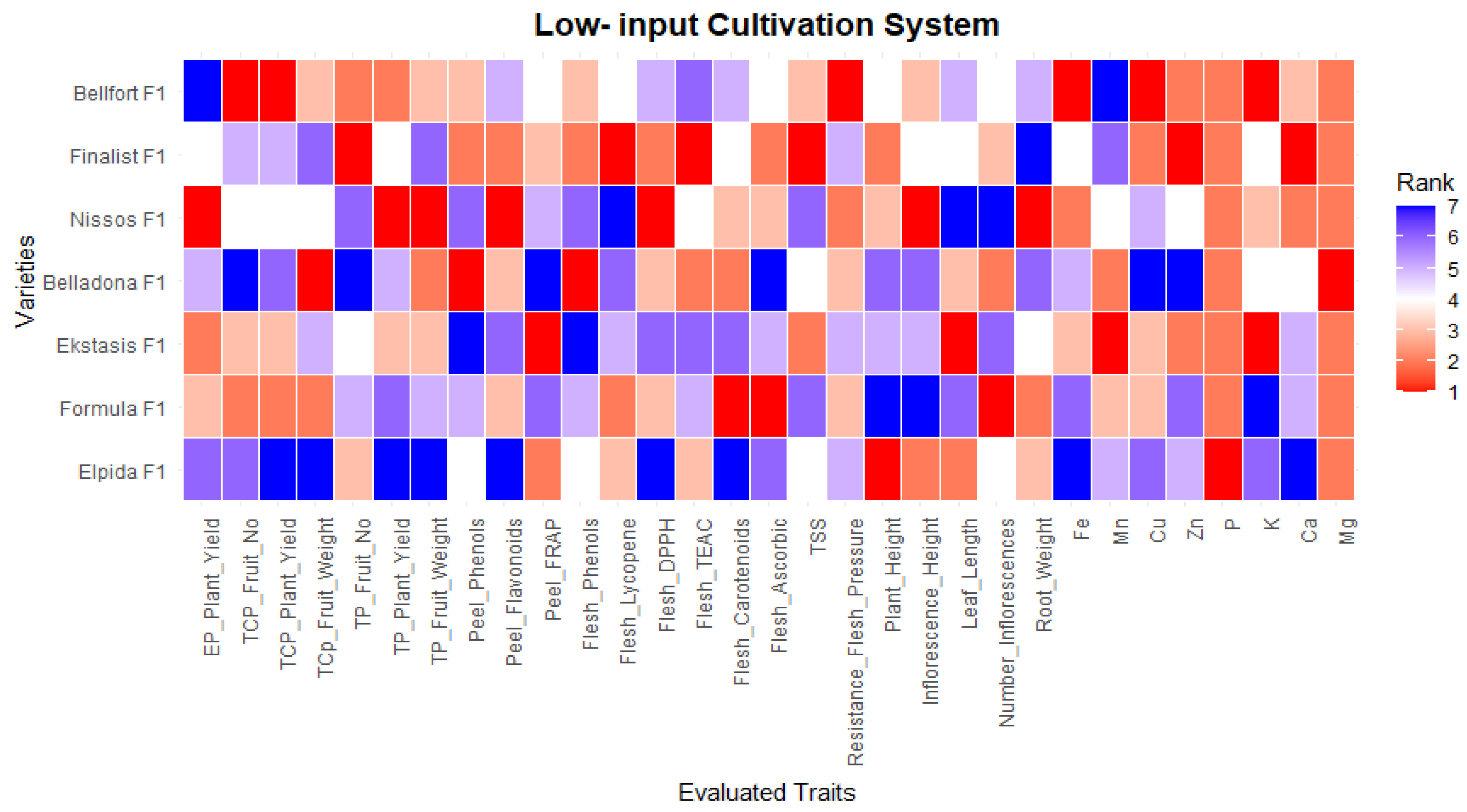
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions. The European Green Deal. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex:52019DC0640 (accessed on 12 February 2025).
- Prada, J.; Dinis, L.-T.; Soriato, E.; Vandelle, E.; Soletkin, O.; Uysal, Ş.; Dihazi, A.; Santos, C.; Santos, J.A. Climate Change Impact on Mediterranean Viticultural Regions and Site-Specific Climate Risk-Reduction Strategies. Mitig. Adapt. Strat. Glob. Change 2024, 29, 52. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The Relation of Climate Extremes with Global Warming in the Mediterranean Region and Its North versus South Contrast. Reg. Environ. Change 2020, 20, 31. [Google Scholar] [CrossRef]
- Zagaria, C.; Schulp, C.J.; Malek, Ž.; Verburg, P.H. Potential for Land and Water Management Adaptations in Mediterranean Croplands under Climate Change. Agric. Syst. 2023, 205, 103586. [Google Scholar] [CrossRef]
- Pomoni, D.I.; Koukou, M.K.; Vrachopoulos, M.G.; Vasiliadis, L. A Review of Hydroponics and Conventional Agriculture Based on Energy and Water Consumption, Environmental Impact, and Land Use. Energies 2023, 16, 1690. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture—Statistical Yearbook 2021; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Tagiakas, R.I.; Avdikos, I.D.; Goula, A.; Koutis, K.; Nianiou-Obeidat, I.; Mavromatis, A.G. Characterization and Evaluation of Greek Tomato Landraces for Productivity and Fruit Quality Traits Related to Sustainable Low-Input Farming Systems. Front. Plant Sci. 2022, 13, 994530. [Google Scholar] [CrossRef]
- Dash, P.K.; Guo, B.; Leskovar, D.I. Optimizing Hydroponic Management Practices for Organically Grown Greenhouse Tomato under Abiotic Stress Conditions. HortScience 2023, 58, 1129–1138. [Google Scholar] [CrossRef]
- Felföldi, Z.; Ranga, F.; Socaci, S.A.; Farcas, A.; Plazas, M.; Sestras, A.F.; Vodnar, D.C.; Prohens, J.; Sestras, R.E. Physico-Chemical, Nutritional, and Sensory Evaluation of Two New Commercial Tomato Hybrids and Their Parental Lines. Plants 2021, 10, 2480. [Google Scholar] [CrossRef]
- Kumar, M.; Ahmad, S.; Singh, R.P. Plant Growth Promoting Microbes: Diverse Roles for Sustainable and Ecofriendly Agriculture. Energy Nexus 2022, 7, 100133. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Chakraborty, I.V.I.; Siddique, W. Characterization of Determinate Tomato Hybrids: Search for Better Processing Qualities. J. Food Process. Technol. 2013, 4, 222. [Google Scholar] [CrossRef]
- Foolad, M.R. Genome Mapping and Molecular Breeding of Tomato. Int. J. Plant Genom. 2007, 2007, 064358. [Google Scholar] [CrossRef]
- Khatri, L.; Kunwar, A.; Bist, D.R. Hydroponics: Advantages and Challenges in Soilless Farming. Big Data Agric. (BDA) 2024, 6, 81–88. [Google Scholar]
- Dwivedi, S.; Goldman, I.; Ortiz, R. Pursuing the Potential of Heirloom Cultivars to Improve Adaptation, Nutritional, and Culinary Features of Food Crops. Agronomy 2019, 9, 441. [Google Scholar] [CrossRef]
- Francis, P.B.; Stark, C.R. Heirloom Tomato Production in Conventional and Transitional-Organic Managed Systems. HortScience 2012, 47, 1034–1037. [Google Scholar] [CrossRef]
- Avdikos, I.D.; Tagiakas, R.; Mylonas, I.; Xynias, I.N.; Mavromatis, A.G. Assessment of Tomato Recombinant Lines in Conventional and Organic Farming Systems for Productivity and Fruit Quality Traits. Agronomy 2021, 11, 129. [Google Scholar] [CrossRef]
- Sharma, M.; Adarsh, M.N.; Kumari, P.; Thakur, M.; Kumar, R.; Sharma, R.; Gautam, N. Hybrid Breeding in Tomato. Int. J. Farm Sci. 2015, 5, 233–250. [Google Scholar]
- Cheema, D.S.; Dhaliwal, M.S. Hybrid Tomato Breeding. J. New Seeds 2004, 6, 1–14. [Google Scholar] [CrossRef]
- Reganold, J.P.; Wachter, J.M. Organic Agriculture in the Twenty-First Century. Nat. Plants 2016, 2, 1–8. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Horváth, K.Z.; Andryei, B.; Ilahy, R.; Takács, S.; Neményi, A.; Pék, Z.; Helyes, L. Impact of Plant Growth-Promoting Rhizobacteria Inoculation on the Physiological Response and Productivity Traits of Field-Grown Tomatoes in Hungary. Horticulturae 2022, 8, 641. [Google Scholar] [CrossRef]
- Barriuso, J.; Solano, B.R.; Gutiérrez Mañero, F.J. Protection Against Pathogen and Salt Stress by Four Plant Growth-Promoting Rhizobacteria Isolated from Pinus Sp. on Arabidopsis thaliana. Phytopathology 2008, 98, 666–672. [Google Scholar] [CrossRef]
- Liu, T.; Yang, X.; Batchelor, W.D.; Liu, Z.; Zhang, Z.; Wan, N.; Sun, S.; He, B.; Gao, J.; Bai, F. A Case Study of Climate-Smart Management in Foxtail Millet (Setaria Italica) Production under Future Climate Change in Lishu County of Jilin, China. Agric. For. Meteorol. 2020, 292, 108131. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Razdan, N.; Saharan, K. Rhizospheric Microbiome: Bio-Based Emerging Strategies for Sustainable Agriculture Development and Future Perspectives. Microbiol. Res. 2022, 254, 126901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cui, G.; Zhang, W.; Lang, D.; Li, Z.; Zhang, X. Bacillus Sp. G2 Improved the Growth of Glycyrrhiza Uralensis Fisch. Related to Antioxidant Metabolism and Osmotic Adjustment. Acta Physiol. Plant. 2021, 43, 152. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Ryu, C.-M. Chatting with a Tiny Belowground Member of the Holobiome: Communication between Plants and Growth-Promoting Rhizobacteria. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 82, pp. 135–160. [Google Scholar]
- Pereira, S.I.A.; Abreu, D.; Moreira, H.; Vega, A.; Castro, P.M.L. Plant Growth-Promoting Rhizobacteria (PGPR) Improve the Growth and Nutrient Use Efficiency in Maize (Zea mays L.) under Water Deficit Conditions. Heliyon 2020, 6, e05106. [Google Scholar] [CrossRef]
- Andryei, B.; Horváth, K.Z.; Agyemang Duah, S.; Takács, S.; Égei, M.; Szuvandzsiev, P.; Neményi, A. Use of Plant Growth Promoting Rhizobacteria (PGPRs) in the Mitigation of Water Deficiency of Tomato Plants (Solanum lycopersicum L.). J. Cent. Eur. Agric. 2021, 22, 167–177. [Google Scholar]
- Singh, V.K.; Singh, A.K.; Singh, P.P.; Kumar, A. Interaction of Plant Growth Promoting Bacteria with Tomato under Abiotic Stress: A Review. Agric. Ecosyst. Environ. 2018, 267, 129–140. [Google Scholar] [CrossRef]
- Adedayo, A.A.; Babalola, O.O.; Prigent-Combaret, C.; Cruz, C.; Stefan, M.; Kutu, F.; Glick, B.R. The Application of Plant Growth-Promoting Rhizobacteria in Solanum Lycopersicum Production in the Agricultural System: A Review. PeerJ 2022, 10, e13405. [Google Scholar] [CrossRef]
- Miller, R.O. High-Temperature Oxidation: Dry Ashing. In Proceedings of the Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 2019; pp. 53–56. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef]
- Fish, W.W.; Perkins-Veazie, P.; Collins, J.K. A Quantitative Assay for Lycopene That Utilizes Reduced Volumes of Organic Solvents. J. Food Compos. Anal. 2002, 15, 309–317. [Google Scholar] [CrossRef]
- Su, C.; Li, T.; Wang, Y.; Ge, Z.; Xiao, J.; Shi, X.; Wang, B. Comparison of Phenolic Composition, Vitamin C, Antioxidant Activity, and Aromatic Components in Apricots from Xinjiang. J. Food Sci. 2022, 87, 231–250. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. [34] Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Deepa, N.; Kaur, C.; Singh, B.; Kapoor, H.C. Antioxidant Activity in Some Red Sweet Pepper Cultivars. J. Food Compos. Anal. 2006, 19, 572–578. [Google Scholar] [CrossRef]
- Giannakoula, A.E.; Ilias, I.F. The Effect of Water Stress and Salinity on Growth and Physiology of Tomato (Lycopersicon esculentum Mil.). Arch. Biol. Sci. 2013, 65, 611–620. [Google Scholar] [CrossRef]
- Allaire, J. RStudio: Integrated Development Environment for R, Version 4.3.2; Computer Software; RStudio: Boston, USA, 2023. [Google Scholar]
- de Mendiburu, F. agricolae: Statistical Procedures for Agricultural Research, Version 1.3-7; R package; CRAN: Vienna, Austria, 2019; Available online: https://CRAN.R-project.org/package=agricolae (accessed on 14 January 2025).
- Wickham, H. Data Analysis. In ggplot2; Use R! Springer International Publishing: Cham, Switzerland, 2016; pp. 189–201. ISBN 978-3-319-24275-0. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps; CRAN: Vienna, Austria, 2019; Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 14 January 2025).
- Filipovic, J.; Stankovic, S.; Rahovic, D.; Tomic, V.; Ljiljanic, N.; Radisic, R.; Maslovaric, M. Gross Margin and Economic Parameters of Grape Production in Pomoravlje Region, Serbia. In Proceedings of the VIII International Scientific Agriculture Symposium, “Agrosym 2017”, Jahorina, Bosnia and Herzegovina, 5–8 October 2017; Book of Proceedings; Agricultural Advisory Service of Jagodina: Jagodina, Serbia, 2017. [Google Scholar]
- Avdikos, I.D.; Tagiakas, R.; Tsouvaltzis, P.; Mylonas, I.; Xynias, I.N.; Mavromatis, A.G. Comparative Evaluation of Tomato Hybrids and Inbred Lines for Fruit Quality Traits. Agronomy 2021, 11, 609. [Google Scholar] [CrossRef]
- Labroo, M.R.; Studer, A.J.; Rutkoski, J.E. Heterosis and Hybrid Crop Breeding: A Multidisciplinary Review. Front. Genet. 2021, 12, 643761. [Google Scholar] [CrossRef]
- Carbonell, P.; Salinas, J.F.; Alonso, A.; Grau, A.; Cabrera, J.A.; García-Martínez, S.; Ruiz, J.J. Effect of Low Inputs and Salinity on Yield and Quality–A 3 Year Study in Virus-Resistant Tomato (Solanum lycopersicum L.) Breeding Lines and Hybrids. Sci. Hortic. 2020, 260, 108889. [Google Scholar] [CrossRef]
- Collinson, S.; Hamdziripi, E.; De Groote, H.; Ndegwa, M.; Cairns, J.E.; Albertsen, M.; Ligeyo, D.; Mashingaidze, K.; Olsen, M.S. Incorporating Male Sterility Increases Hybrid Maize Yield in Low Input African Farming Systems. Commun. Biol. 2022, 5, 729. [Google Scholar] [CrossRef]
- Ter Steeg, E.M.; Struik, P.C.; Visser, R.G.; Lindhout, P. Crucial Factors for the Feasibility of Commercial Hybrid Breeding in Food Crops. Nat. Plants 2022, 8, 463–473. [Google Scholar] [CrossRef]
- Shahzad, R.; Tayade, R.; Shahid, M.; Hussain, A.; Ali, M.W.; Yun, B.-W. Evaluation Potential of PGPR to Protect Tomato against Fusarium Wilt and Promote Plant Growth. PeerJ 2021, 9, e11194. [Google Scholar]
- Gashash, E.A.; Osman, N.A.; Alsahli, A.A.; Hewait, H.M.; Ashmawi, A.E.; Alshallash, K.S.; El-Taher, A.M.; Azab, E.S.; Abd El-Raouf, H.S.; Ibrahim, M.F. Effects of Plant-Growth-Promoting Rhizobacteria (PGPR) and Cyanobacteria on Botanical Characteristics of Tomato (Solanum lycopersicon L.) Plants. Plants 2022, 11, 2732. [Google Scholar] [CrossRef]
- Cochard, B.; Giroud, B.; Crovadore, J.; Chablais, R.; Arminjon, L.; Lefort, F. Endophytic PGPR from Tomato Roots: Isolation, in Vitro Characterization and in Vivo Evaluation of Treated Tomatoes (Solanum lycopersicum L.). Microorganisms 2022, 10, 765. [Google Scholar] [CrossRef] [PubMed]
- Paull, J. Organic Agriculture in Europe: EU Sets Goal of Growing Organic Farmland from 10% to 25% by 2030. Eur. J. Agric. Food Sci. 2024, 6, 26–31. [Google Scholar] [CrossRef]
- Lyu, D.; Zajonc, J.; Pagé, A.; Tanney, C.A.; Shah, A.; Monjezi, N.; Msimbira, L.A.; Antar, M.; Nazari, M.; Backer, R. Plant Holobiont Theory: The Phytomicrobiome Plays a Central Role in Evolution and Success. Microorganisms 2021, 9, 675. [Google Scholar] [CrossRef] [PubMed]
- Antar, M.; Lyu, D.; Nazari, M.; Shah, A.; Zhou, X.; Smith, D.L. Biomass for a Sustainable Bioeconomy: An Overview of World Biomass Production and Utilization. Renew. Sustain. Energy Rev. 2021, 139, 110691. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst Phytohormones from Planta and PGPR under Biotic and Abiotic Stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Naveed, M.; Ahmad, M.; Zahir, Z.A. Rhizosphere Bacteria for Crop Production and Improvement of Stress Tolerance: Mechanisms of Action, Applications, and Future Prospects. In Plant Microbes Symbiosis: Applied Facets; Arora, N.K., Ed.; Springer: New Delhi, India, 2015; pp. 1–36. ISBN 978-81-322-2067-1. [Google Scholar]
- Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal Plants; Egamberdieva, D., Shrivastava, S., Varma, A., Eds.; Soil Biology; Springer International Publishing: Cham, Switzerland, 2015; Volume 42, ISBN 978-3-319-13400-0. [Google Scholar]
- Mena-Violante, H.G.; Olalde-Portugal, V. Alteration of Tomato Fruit Quality by Root Inoculation with Plant Growth-Promoting Rhizobacteria (PGPR): Bacillus Subtilis BEB-13bs. Sci. Hortic. 2007, 113, 103–106. [Google Scholar] [CrossRef]
- Zameer, M.; Zahid, H.; Tabassum, B.; Ali, Q.; Nasir, I.A.; Saleem, M.; Butt, S.J. PGPR Potentially Improve Growth of Tomato Plants in Salt-Stressed Environment. Turk. J. Agric.-Food Sci. Technol. 2016, 4, 455–463. [Google Scholar] [CrossRef]
- Almaghrabi, O.A.; Massoud, S.I.; Abdelmoneim, T.S. Influence of Inoculation with Plant Growth Promoting Rhizobacteria (PGPR) on Tomato Plant Growth and Nematode Reproduction under Greenhouse Conditions. Saudi J. Biol. Sci. 2013, 20, 57–61. [Google Scholar] [CrossRef]
- Hasan, A.; Tabassum, B.; Hashim, M.; Khan, N. Role of Plant Growth Promoting Rhizobacteria (PGPR) as a Plant Growth Enhancer for Sustainable Agriculture: A Review. Bacteria 2024, 3, 59–75. [Google Scholar] [CrossRef]
- Ghosh, A.; Acharya, R.; Shaw, S.; Gangopadhyay, D. Plant Growth-Promoting Rhizobacteria (PGPR): A Potential Alternative Tool for Sustainable Agriculture. In Updates on Rhizobacteria; IntechOpen: London, UK, 2024. [Google Scholar]
- Gagné, S.; Dehbi, L.; Le Quéré, D.; Cayer, F.; Morin, J.-L.; Lemay, R.; Fournier, N. Increase of Greenhouse Tomato Fruit Yields by Plant Growth-Promoting Rhizobacteria (PGPR) Inoculated into the Peat-Based Growing Media. Soil Biol. Biochem. 1993, 25, 269–272. [Google Scholar] [CrossRef]
- Yagmur, B.; Gunes, A. Evaluation of the Effects of Plant Growth Promoting Rhizobacteria (PGPR) on Yield and Quality Parameters of Tomato Plants in Organic Agriculture by Principal Component Analysis (PCA). Gesunde Pflanz. 2021, 73, 219–228. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Swapnil, P. Plant Growth-Promoting Rhizobacteria as a Green Alternative for Sustainable Agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Sharafzadeh, S. Effects of PGPR on Growth and Nutrients Uptake of Tomato. Int. J. Adv. Eng. Technol. 2012, 2, 27. [Google Scholar]
- Vélez-Terreros, P.Y.; Romero-Estévez, D.; Navarrete, H.; Yánez-Jácome, G.S. Nutritional Quality of Conventional, Organic, and Hydroponic Tomatoes Commercialized in Quito, Ecuador. Foods 2024, 13, 1348. [Google Scholar] [CrossRef]
- Fernandes, I.; Leça, J.M.; Aguiar, R.; Fernandes, T.; Marques, J.C.; Cordeiro, N. Influence of Crop System Fruit Quality, Carotenoids, Fatty Acids and Phenolic Compounds in Cherry Tomatoes. Agric Res 2021, 10, 56–65. [Google Scholar] [CrossRef]
- Tahiri, A.; Meddich, A.; Raklami, A.; Alahmad, A.; Bechtaoui, N.; Anli, M.; Göttfert, M.; Heulin, T.; Achouak, W.; Oufdou, K. Assessing the Potential Role of Compost, PGPR, and AMF in Improving Tomato Plant Growth, Yield, Fruit Quality, and Water Stress Tolerance. J. Soil Sci. Plant Nutr. 2022, 22, 743–764. [Google Scholar] [CrossRef]
- Shilpa; Sharma, A.K.; Chauhan, M.; Bijalwan, P. Plant Growth Promoting Rhizobacteria, Organic Manures, and Chemical Fertilizers: Impact on Crop Productivity and Soil Health of Capsicum (Capsicum annuum L.) in North Western Himalayan Region. J. Plant Nutr. 2024, 47, 448–467. [Google Scholar] [CrossRef]
- Flaviano, T.; Bernardo, M.H.J.; Angelica, G.; Gabriele, C.; Fabrizio, L.; Walter, P.; Cristiano, P.; Clemencia, C.-L.; Fabio, S. Impact of Organic and Conventional Cropping Systems on Plant Growth-Promoting Rhizobacteria (PGPR). Biocatal. Agric. Biotechnol. 2025, 64, 103527. [Google Scholar] [CrossRef]
- Avdikos, I.D.; Nteve, G.-M.; Apostolopoulou, A.; Tagiakas, R.; Mylonas, I.; Xynias, I.N.; Papathanasiou, F.; Kalaitzis, P.; Mavromatis, A.G. Analysis of Re-Heterosis for Yield and Fruit Quality in Restructured Hybrids, Generated from Crossings among Tomato Recombinant Lines. Agronomy 2021, 11, 822. [Google Scholar] [CrossRef]
- Gerakari, M.; Kyriakoudi, A.; Nokas, D.; Mourtzinos, I.; Chronopoulou, E.G.; Tani, E.; Avdikos, I. Evaluation of the Potential Use of Wild Relatives of Tomato (Solanum pennellii) to Improve Yield and Fruit Quality Under Low-Input and High-Salinity Cultivation Conditions. Agronomy 2024, 14, 3042. [Google Scholar] [CrossRef]
- Miller, J.C.; Tanksley, S.D. RFLP Analysis of Phylogenetic Relationships and Genetic Variation in the Genus Lycopersicon. Theoret. Appl. Genet. 1990, 80, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Lindhout, P. Domestication and Breeding of Tomatoes: What Have We Gained and What Can We Gain in the Future? Ann. Bot. 2007, 100, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Tsiouni, M.; Aggelopoulos, S.; Pavloudi, A.; Siggia, D. Economic and Financial Sustainability Dependency on Subsidies: The Case of Goat Farms in Greece. Sustainability 2021, 13, 7441. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Scoring System |
|---|---|
| Plant growth type | 1: Determinate → 2: Indeterminate |
| Number of inflorescences on the main stem | Count |
| Plant height | cm |
| Height of the 4th inflorescence | cm |
| Length of internodes | cm |
| Anthocyanin coloration of the stem | 1: Absent → 5: Very strong |
| Leaf attitude | 1: Erect → 3: Horizontal → 5: Drooping |
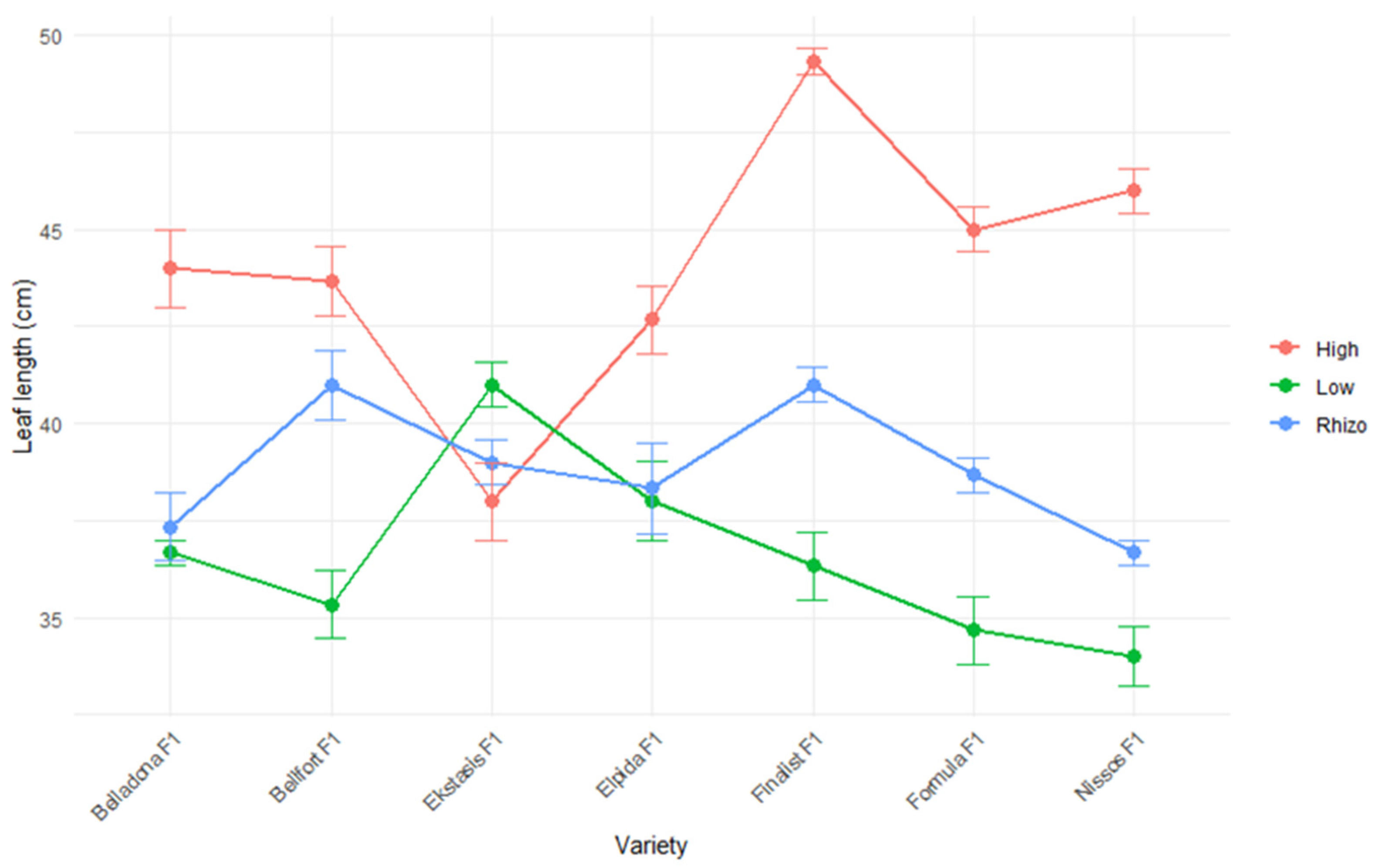
| Leaf length | cm |
| Leaf width | cm |
| Leaf’s intensity of green color | 1: Very light → 9: Very dark |
| Attitude of the petiolule of leaflets in relation to the petiole | 1: Erect → 2: Horizontal → 3: Drooping |
| Type of leaf | 1: Pinnate → 2: Bipinnate |
| Type of inflorescence | 1: Mainly uniparous → 2: Equally uniparous and multiparous → 3: Mainly multiparous |
| Flower color | 1: Yellow → 2: Orange |
| Pedicel abscission layer | 1: Absent → 9: Present |
| Pubescence of pedicel | 1: Absent → 9: Very strong |
| Presence of green shoulder in fruits | 1: Absent → 9: Present |
| Extent of green shoulder | 1: Very small → 9: Very large |
| Intensity of green color of shoulder | 1: Very light → 9: Very dark |
| Intensity of green color excluding the shoulder | 1: Very light → 9: Very dark |
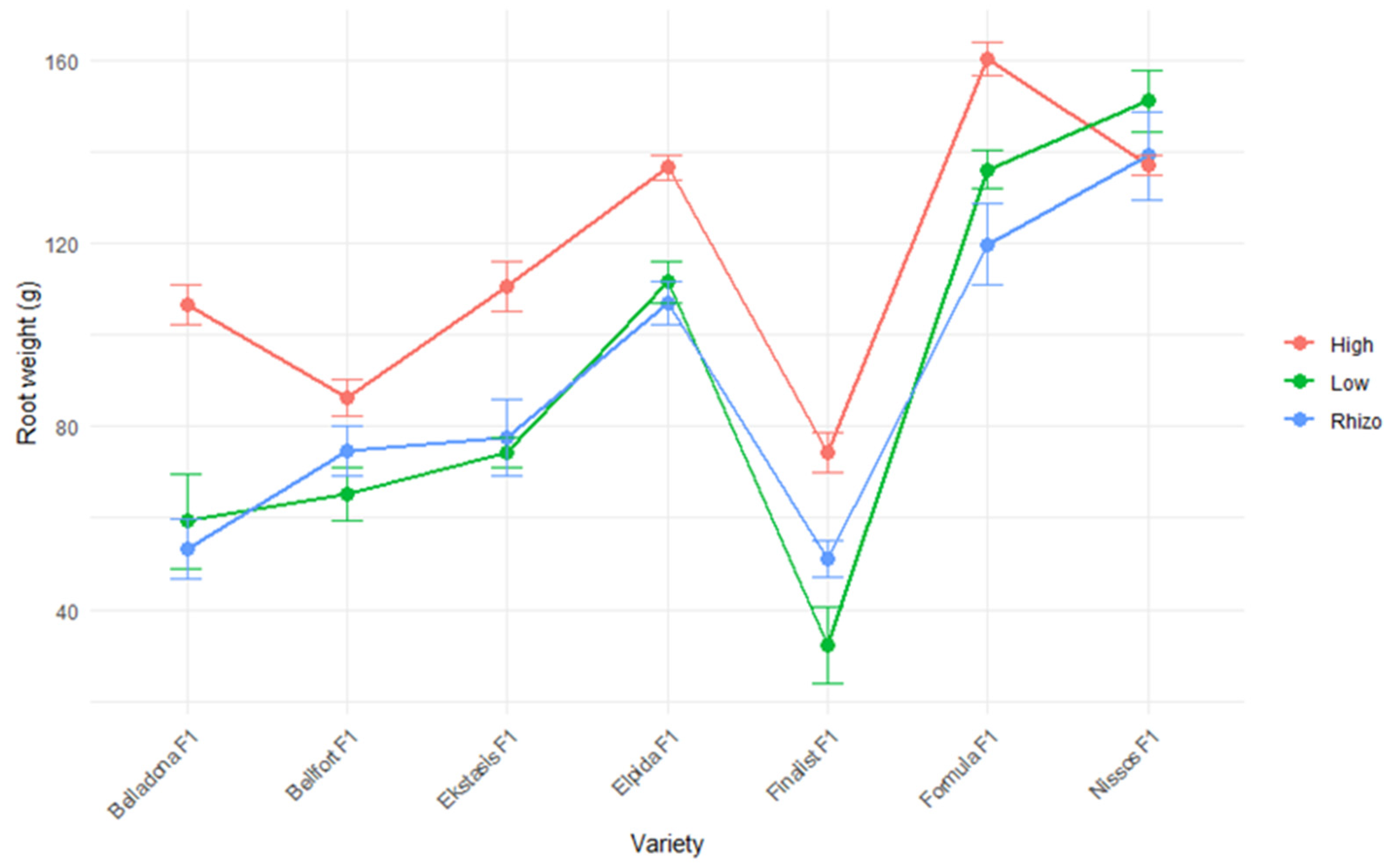
| Root weight | g |
| Variety | Plant Height (cm) (65 DAT) | 4th Inflorescence’s Height (cm) | Leaf Length (cm) | Number of Inflorescences | Root Weight (g) |
|---|---|---|---|---|---|
| Elpida F1 | 147.78 ± 27.73 ab | 94.11 ± 14.19 b | 39.67 ± 2.40 a | 6.67 ± 0.50 c | 119.76 ± 8.59 b |
| Formula F1 | 121.67 ± 17.01 c | 82.89 ± 8.53 c | 39.44 ± 2.76 a | 7.22 ± 0.25 ab | 138.64 ± 15.68 a |
| Ekstasis F1 | 124.00 ± 22.89 c | 79.00 ± 10.73 c | 39.33 ± 1.15 a | 6.78 ± 0.38 bc | 87.38 ± 13.40 c |
| Belladona F1 | 125.11 ± 19.81 c | 82.44 ± 10.16 c | 39.33 ± 2.14 a | 7.67 ± 0.41 a | 73.08 ± 15.98 c |
| Nissos F1 | 146.67 ± 27.47 ab | 108.67 ± 17.94 a | 38.89 ± 3.22 a | 6.11 ± 0.35 d | 142.33 ± 11.43 a |
| Finalist F1 | 154.44 ± 25.87 a | 94.56 ± 9.42 b | 42.22 ± 3.33 a | 7.67 ± 0.58 a | 52.53 ± 11.69 d |
| Bellfort F1 | 139.78 ± 26.96 b | 90.78 ± 12.72 b | 40.00 ± 2.65 a | 7.33 ± 0.41 a | 75.35 ± 6.88 c |
| Low input | 163.43 ± 10.62 a | 102.57 ± 8.41 a | 36.57 ± 1.62 c | 6.62 ± 0.34 b | 89.93 ± 25.31 b |
| Low input and PGPR | 164.19 ± 13.40 a | 104.90 ± 7.69 a | 38.86 ± 1.73 b | 6.86 ± 0.49 b | 87.97 ± 21.26 b |
| High input | 83.57 ± 3.94 b | 63.57 ± 3.31 b | 44.10 ± 2.03 a | 7.71 ± 0.45 a | 115.9 ± 16.99 a |
| Variety | Early Yield | ||
|---|---|---|---|
| Fruit Number | Plant Yield (g) | Fruit Weight (g) | |
| Elpida F1 | 13.45 ± 0.55 a | 2379.77 ± 262.2 a | 181.09 ± 21.59 a |
| Formula F1 | 12.19 ± 0.35 bc | 2149.69 ± 137.08 a | 175.16 ± 9.55 a |
| Ekstasis F1 | 12.90 ± 0.39 ab | 2429.10 ± 171.79 a | 187.05 ± 11.46 a |
| Belladona F1 | 12.21 ± 0.38 abc | 2163.61 ± 141 a | 176.90 ± 10.44 a |
| Nissos F1 | 11.88 ± 0.48 bc | 2295.06 ± 305.92 a | 184.46 ± 19.92 a |
| Finalist F1 | 13.13 ± 0.52 ab | 2051.91 ± 149.84 a | 161.64 ± 12.25 a |
| Bellfort F1 | 11.56 ± 0.74 c | 2060.75 ± 204.75 a | 172.66 ± 13.11 a |
| Low input | 12.33 ± 0.88 a | 1629.43 ± 263.26 a | 128.90 ± 19.86 b |
| Low input and PGPR | 12.62 ± 1.19 a | 1566.00 ± 185.57 a | 128.43 ± 10.30 b |
| High input | 12.72 ± 0.46 a | 3159.05 ± 259.26 a | 250.01 ± 17.72 a |
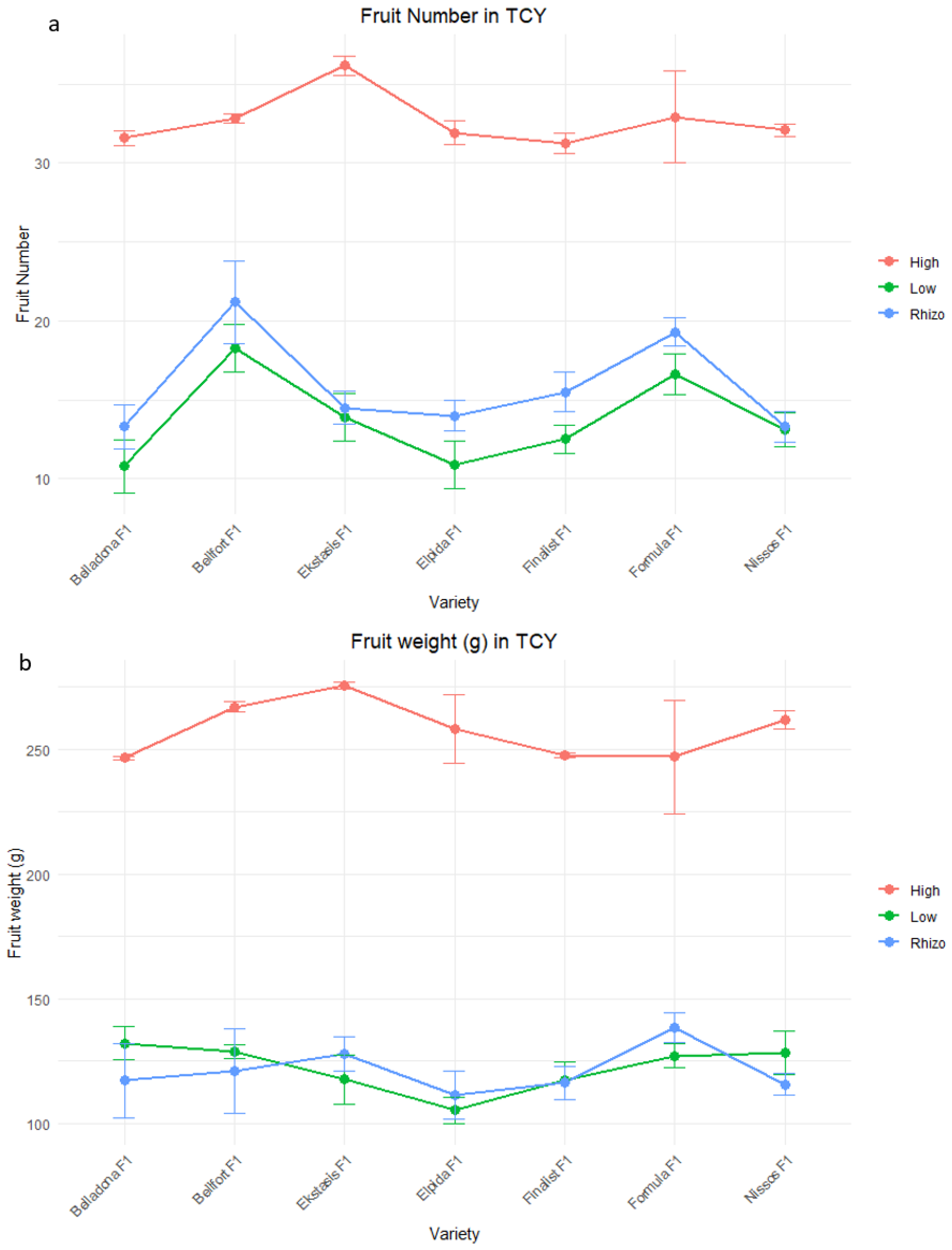
| Variety | Total Commercial Yield | ||
|---|---|---|---|
| Fruit Number | Plant Yield (g) | Fruit Weight (g) | |
| Elpida F1 | 20.03 ± 1.82 c | 4022.29 ± 650.99 c | 166.36 ± 14.51 a |
| Formula F1 | 23.56 ± 1.39 a | 4522.63 ± 500.28 ab | 175.57 ± 10.13 a |
| Ekstasis F1 | 22.71 ± 2.02 b | 4924.65 ± 725.05 a | 181.98 ± 13.89 a |
| Belladona F1 | 20.32 ± 1.96 c | 4178.57 ± 599.90 c | 177.43 ± 12.45 a |
| Nissos F1 | 20.28 ± 1.69 c | 4162.50 ± 588.93 bc | 174.41 ± 12.47 a |
| Finalist F1 | 20.47 ± 1.59 c | 3920.53 ± 531.53 c | 165.82 ± 11.65 a |
| Bellfort F1 | 24.66 ± 1.48 a | 4843.38 ± 572.54 ab | 178.16 ± 13.32 a |
| Low input | 14.14 ± 1.99 c | 1675.81 ± 349.85 c | 120.33 ± 12.83 b |
| Low input and PGPR | 15.76 ± 1.75 b | 1966.05 ± 289.64 b | 123.52 ± 6.47 b |
| High input | 32.6 ± 1.02 a | 8298.68 ± 492.76 a | 254.80 ± 8.25 a |
| Variety | Total Yield | ||
|---|---|---|---|
| Fruit Number | Plant Yield (g) | Fruit Weight (g) | |
| Elpida F1 | 28.03 ± 1.03 a | 4945.48 ± 570.87 ab | 165.69 ± 14.83 a |
| Formula F1 | 26.25 ± 1.09 ab | 4662.44 ± 486.19 b | 165.83 ± 11.33 a |
| Ekstasis F1 | 27.77 ± 1.37 ab | 5554.23 ± 643.14 a | 182.52 ± 13.85 a |
| Belladona F1 | 24.43 ± 1.42 c | 4740.68 ± 521.97 b | 178.57 ± 11.25 a |
| Nissos F1 | 25.50 ± 1.20 bc | 5051.19 ± 570.10 ab | 183.14 ± 16.11 a |
| Finalist F1 | 27.16 ± 0.83 ab | 4579.13 ± 464.59 b | 159.27 ± 12.28 a |
| Bellfort F1 | 27.50 ± 1.38 ab | 5029.75 ± 557.32 b | 171.83 ± 13.80 a |
| Low input | 22.33 ± 1.46 b | 2555.19 ± 429.23 b | 115.90 ± 17.77 b |
| Low input and PGPR | 22.86 ± 1.54 b | 2801.86 ± 246.88 b | 123.33 ± 8.49 b |
| High input | 33.17 ± 1.04 a | 8402.26 ± 491.43 a | 253.32 ± 8.45 a |
| Variety | Fe (ppm) | Mn (ppm) | Cu (ppm) | Zn (ppm) |
|---|---|---|---|---|
| Elpida F1 | 61.36 ± 7.57 bc | 79.43 ± 7.48 bc | 10.40 ± 1.29 bc | 20.20 ± 1.49 c |
| Formula F1 | 58.65 ± 3.94 c | 87.74 ± 8.36 ab | 12.00 ± 0.97 a | 21.05 ± 2.06 c |
| Ekstasis F1 | 67.72 ± 5.08 ab | 85.32 ± 6.28 ab | 11.36 ± 0.97 abc | 25.55 ± 2.20 a |
| Belladona F1 | 64.42 ± 5.02 bc | 91.02 ± 5.07 a | 10.02 ± 1.02 c | 20.06 ± 1.95 c |
| Nissos F1 | 72.96 ± 7.44 a | 84.21 ± 7.82 abc | 11.82 ± 1.25 ab | 24.78 ± 1.96 ab |
| Finalist F1 | 58.72 ± 4.31 c | 74.69 ± 6.40 cd | 11.70 ± 0.73 ab | 23.44 ± 1.77 b |
| Bellfort F1 | 74.05 ± 7.60 a | 69.33 ± 6.67 d | 11.25 ± 0.64 abc | 23.23 ± 1.50 b |
| Low input | 61.13 ± 8.01 b | 78.81 ± 8.93 b | 10.45 ± 0.96 b | 20.29 ± 1.72 b |
| Low input and PGPR | 61.69 ± 4.78 b | 75.50 ± 6.77 b | 11.02 ± 0.75 b | 21.38 ± 1.08 b |
| High input | 73.42 ± 4.26 a | 90.73 ± 4.13 a | 12.2 ± 1.14 a | 25.97 ± 1.83 a |
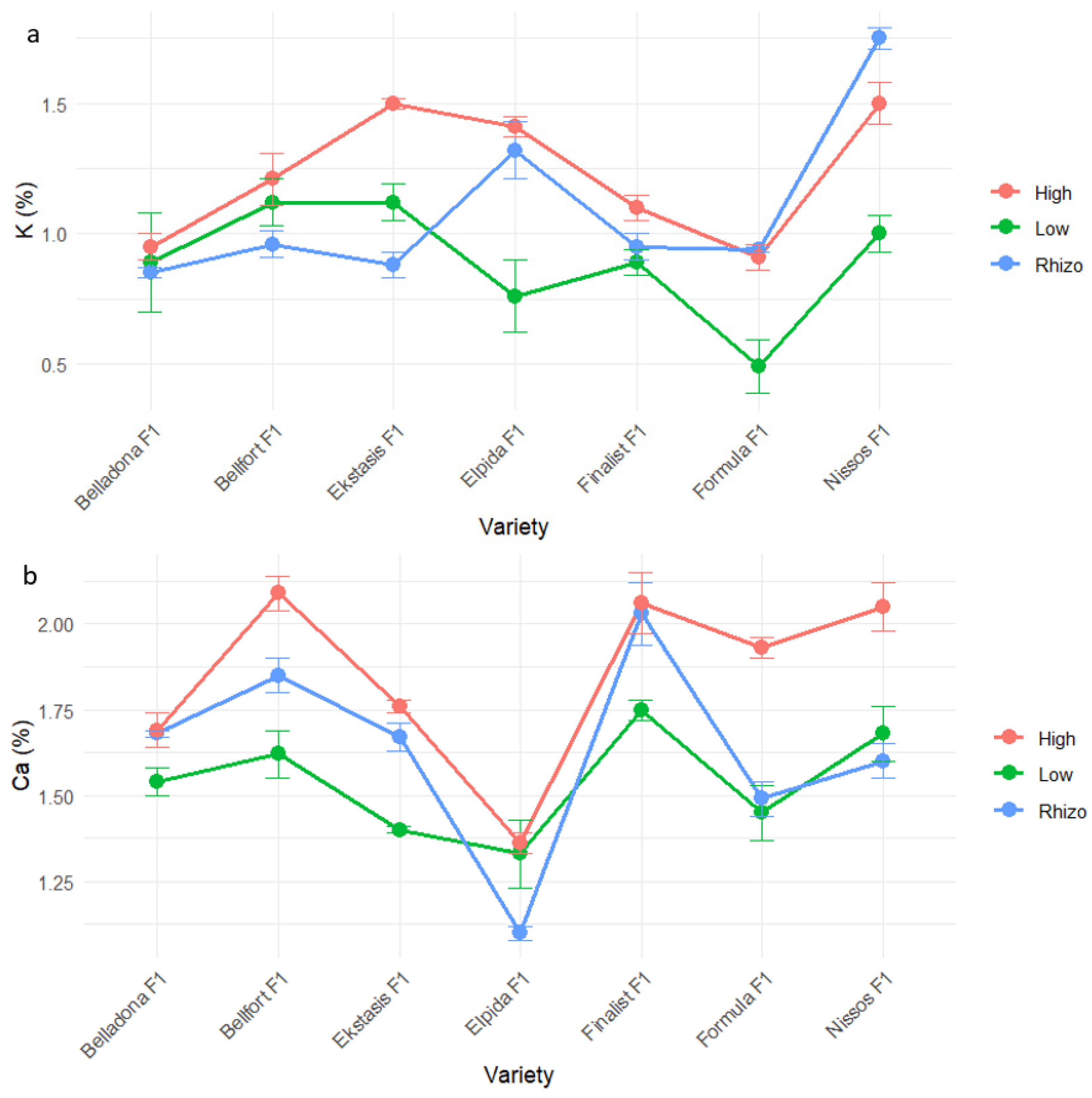
| P (%) | K (%) | Ca (%) | Mg (%) | |
| Elpida F1 | 0.18 ± 0.12 a | 1.17 ± 0.28 ab | 1.27 ± 0.23 b | 0.15 ± 0.02 a |
| Formula F1 | 0.10 ± 0.04 a | 0.78 ± 0.18 c | 1.62 ± 0.25 a | 0.16 ± 0.03 a |
| Ekstasis F1 | 0.11 ± 0.02 a | 1.17 ± 0.36 ab | 1.61 ± 0.13 a | 0.09 ± 0.01 b |
| Belladona F1 | 0.11 ± 0.04 a | 0.90 ± 0.10 bc | 1.64 ± 0.13 a | 0.16 ± 0.02 a |
| Nissos F1 | 0.09 ± 0.02 a | 1.41 ± 0.22 a | 1.78 ± 0.22 a | 0.14 ± 0.03 a |
| Finalist F1 | 0.11 ± 0.01 a | 0.98 ± 0.10 bc | 1.95 ± 0.21 a | 0.13 ± 0.02 a |
| Bellfort F1 | 0.07 ± 0.01 a | 1.10 ± 0.10 abc | 1.85 ± 0.18 a | 0.14 ± 0.02 a |
| Low input | 0.10 ± 0.05 a | 0.90 ± 0.19 b | 1.54 ± 0.16 b | 0.13 ± 0.03 a |
| Low input and PGPR | 0.08 ± 0.01 a | 1.09 ± 0.19 ab | 1.63 ± 0.21 b | 0.15 ± 0.02 a |
| High input | 0.14 ± 0.07 a | 1.23 ± 0.27 a | 1.85 ± 0.26 a | 0.14 ± 0.03 a |
| Variety | Total Soluble Solids (Brixo) | Resistance of Flesh to Pressure |
|---|---|---|
| Elpida F1 | 5.76 ± 0.29 b | 0.90 ± 0.09 d |
| Formula F1 | 4.80 ± 0.14 c | 0.95 ± 0.05 bcd |
| Ekstasis F1 | 6.07 ± 0.28 a | 0.94 ± 0.04 cd |
| Belladona F1 | 5.51 ± 0.12 bc | 0.88 ± 0.06 d |
| Nissos F1 | 5.27 ± 0.29 bc | 1.06 ± 0.07 ab |
| Finalist F1 | 5.20 ± 0.38 a | 1.03 ± 0.09 abc |
| Bellfort F1 | 5.52 ± 0.16 bc | 1.12 ± 0.12 a |
| Low input | 5.46 ± 0.25 b | 1.00 ± 0.06 a |
| Low input and PGPR | 5.57 ± 0.26 b | 0.97 ± 0.07 a |
| High input | 6.05 ± 0.29 a | 0.98 ± 0.13 a |
| Variety | Fruit Peel | ||
|---|---|---|---|
| Phenols (mg/100 g FW) | Flavonoids (mg/g FW) | FRAP (mmol/Fe2+ FW) | |
| Elpida F1 | 16.13 ± 1.37 e | 57.61 ± 4.68 a | 54.89 ± 5.22 b |
| Formula F1 | 18.16 ± 0.68 a | 57.04 ± 1.90 a | 53.01 ± 4.14 cd |
| Ekstasis F1 | 17.36 ± 0.56 bc | 58.73 ± 5.85 a | 57.81 ± 2.55 a |
| Belladona F1 | 17.07 ± 0.51 bcd | 60.27 ± 2.06 a | 51.21 ± 3.83 d |
| Nissos F1 | 16.80 ± 0.21 cd | 61.07 ± 2.31 a | 54.57 ± 1.78 bc |
| Finalist F1 | 17.62 ± 0.61 ab | 58.70 ± 1.89 a | 58.06 ± 1.30 a |
| Bellfort F1 | 16.67 ± 0.59 de | 58.36 ± 1.43 a | 52.73 ± 2.52 cd |
| Low input | 17.39 ± 0.44 a | 58.21 ± 2.13 b | 57.94 ± 1.47 a |
| Low input and PGPR | 17.45 ± 0.50 a | 55.43 ± 3.03 c | 57.65 ± 2.20 a |
| High input | 16.50 ± 1.11 b | 62.84 ± 2.88 a | 48.25 ± 2.90 b |
| Variety | Fruit Flesh | |||||
|---|---|---|---|---|---|---|
| Phenols (mg/g FW) | Lycopene (mg/g FW) | DPPH (%) | TEAC (μM/g FW) | Carotenoids (μg/100 g DW) | Ascorbic Acid (mg/g FW) | |
| Elpida F1 | 501.44 ± 75.77 c | 531.99 ± 16.97 c | 50.14 ± 4.16 bc | 6.83 ± 0.70 d | 118.17 ± 7.66 e | 2789.88 ± 145.63 c |
| Formula F1 | 588.21 ± 8.43 a | 593.70 ± 18.73 b | 49.73 ± 1.58 bc | 7.67 ± 0.19 a | 194.16 ± 1.76 a | 3493.68 ± 222.69 a |
| Ekstasis F1 | 564.33 ± 17.88 b | 440.30 ± 8.79 d | 50.88 ± 4.98 ab | 7.29 ± 0.19 bc | 159.34 ± 9.43 d | 2855.52 ± 159.07 c |
| Belladona F1 | 584.37 ± 44.01 a | 385.49 ± 21.93 f | 52.20 ± 1.70 ab | 7.38 ± 0.20 b | 168.00 ± 12.86 c | 2558.74 ± 165.31 d |
| Nissos F1 | 569.30 ± 10.08 ab | 374.74 ± 18.81 f | 52.97 ± 1.95 a | 7.21 ± 0.24 bc | 182.00 ± 4.55 b | 2916.36 ± 83.38 c |
| Finalist F1 | 555.33 ± 44.93 b | 651.06 ± 13.49 a | 50.43 ± 1.49 ab | 7.76 ± 0.31 a | 178.01 ± 3.92 b | 3416.60 ± 127.53 a |
| Bellfort F1 | 513.33 ± 55.74 c | 402.28 ± 36.26 e | 47.90 ± 2.44 c | 7.09 ± 0.22 c | 170.66 ± 1.29 c | 3148.34 ± 111.04 b |
| Low input | 581.90 ± 16.45 a | 489.91 ± 51.26 a | 50.54 ± 1.85 b | 7.57 ± 0.19 a | 170.33 ± 12.97 a | 3028.29 ± 113.07 ab |
| Low input and PGPR | 593.40 ± 20.84 a | 483.95 ± 51.02 ab | 48.14 ± 2.63 c | 7.51 ± 0.24 a | 173.85 ± 12.38 a | 2945.58 ± 198.05 b |
| High input | 485.99 ± 56.63 b | 474.51 ± 82.56 b | 53.14 ± 3.40 a | 6.87 ± 0.47 b | 157.39 ± 17.28 b | 3102.90 ± 335.35 a |
| Low-Input System | Low-Input and RGPR System | High-Input System | |
|---|---|---|---|
| Production (kg/acre) | 5.110 | 5.603 | 16.804 |
| Price (€) | 0.70 | 0.70 | 0.50 |
| Gross Revenue | 3.570 | 3.922 | 8.402 |
| Variable costs | |||
| Fertilizers (€/acre) | 80 | 80 | 740 |
| Plant Protection (€/acre) | 60 | 60 | 340 |
| Plant Capital (€/acre) | 500 | 500 | 500 |
| Irrigation (€/acre) | 70 | 70 | 130 |
| Hired labour (€/acre) | 900 | 900 | 450 |
| Other (€/acre) | 180 | 200 | 250 |
| Interest in operating capital (€/acre) | 95 | 95 | 130 |
| Total variable cost | 1.885 | 1.905 | 2.540 |
| Gross Margin | 1.685 | 2.017 | 5.862 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerakari, M.; Mitkou, D.; Antoniadis, C.; Giannakoula, A.; Stefanou, S.; Hilioti, Z.; Chatzidimopoulos, M.; Tsiouni, M.; Pavloudi, A.; Xynias, I.N.; et al. Evaluation of Commercial Tomato Hybrids for Climate Resilience and Low-Input Farming: Yield and Nutritional Assessment Across Cultivation Systems. Agronomy 2025, 15, 929. https://doi.org/10.3390/agronomy15040929
Gerakari M, Mitkou D, Antoniadis C, Giannakoula A, Stefanou S, Hilioti Z, Chatzidimopoulos M, Tsiouni M, Pavloudi A, Xynias IN, et al. Evaluation of Commercial Tomato Hybrids for Climate Resilience and Low-Input Farming: Yield and Nutritional Assessment Across Cultivation Systems. Agronomy. 2025; 15(4):929. https://doi.org/10.3390/agronomy15040929
Chicago/Turabian StyleGerakari, Maria, Diamantia Mitkou, Christos Antoniadis, Anastasia Giannakoula, Stefanos Stefanou, Zoe Hilioti, Michael Chatzidimopoulos, Maria Tsiouni, Alexandra Pavloudi, Ioannis N. Xynias, and et al. 2025. "Evaluation of Commercial Tomato Hybrids for Climate Resilience and Low-Input Farming: Yield and Nutritional Assessment Across Cultivation Systems" Agronomy 15, no. 4: 929. https://doi.org/10.3390/agronomy15040929
APA StyleGerakari, M., Mitkou, D., Antoniadis, C., Giannakoula, A., Stefanou, S., Hilioti, Z., Chatzidimopoulos, M., Tsiouni, M., Pavloudi, A., Xynias, I. N., & Avdikos, I. D. (2025). Evaluation of Commercial Tomato Hybrids for Climate Resilience and Low-Input Farming: Yield and Nutritional Assessment Across Cultivation Systems. Agronomy, 15(4), 929. https://doi.org/10.3390/agronomy15040929