
Effects of Geraniol on Survival, Reproduction, Endophytes, and Transcriptome of Tea Green Leafhoppers (Empoasca onukii)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Selectivity of Tea Green Leafhoppers to Geraniol
2.3. Determination of the Effect of Geraniol Treatment on the Survival Rate of Tea Green Leafhoppers
2.4. Determination of the Total Number of Eggs Laid by Tea Green Leafhoppers
2.5. Determination of the Egg Hatching Rate of Tea Green Leafhoppers
2.6. Determination of Endophytes of Tea Green Leafhoppers
2.7. Determination of the Transcriptome of Tea Green Leafhoppers
3. Results
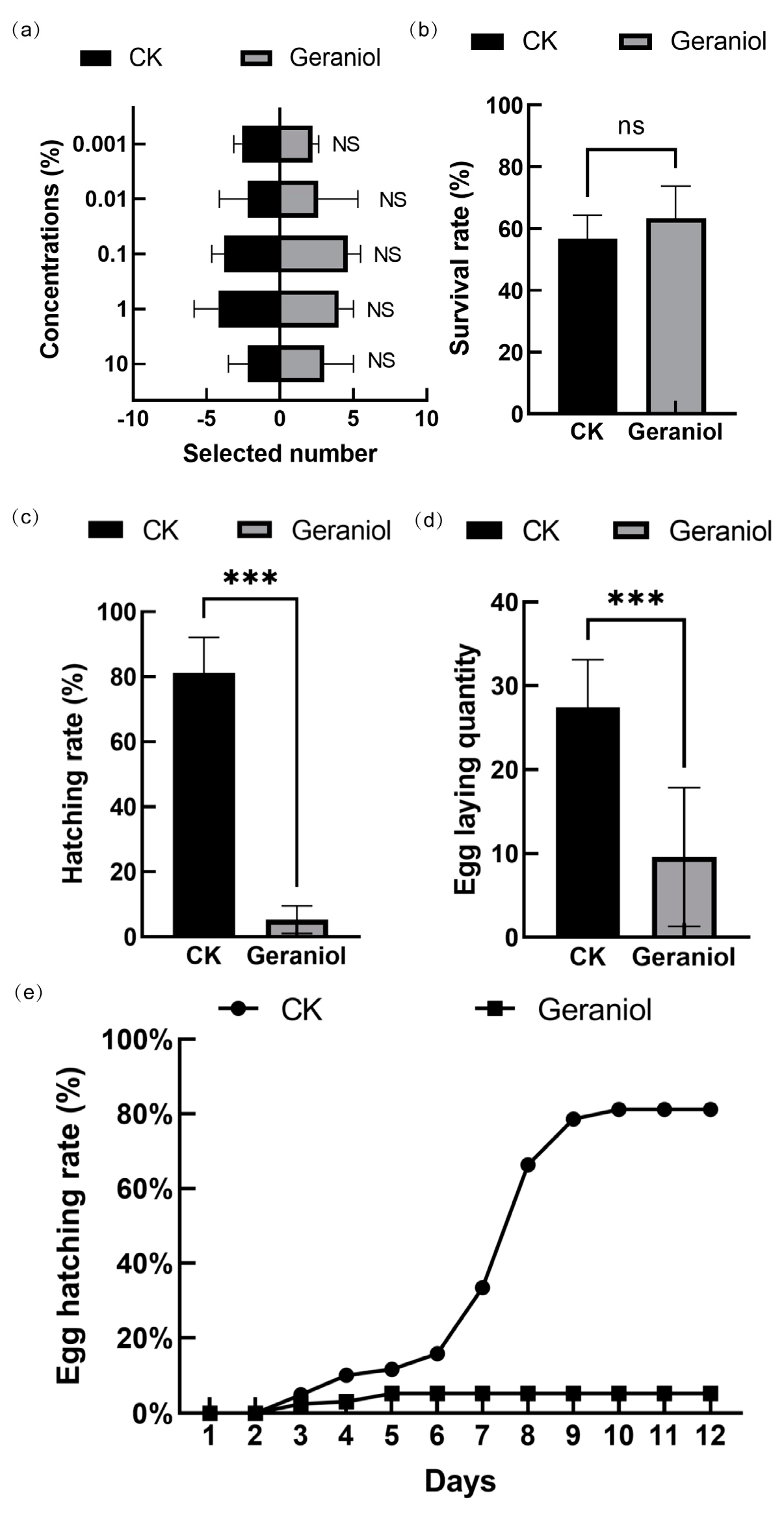
3.1. Geraniol Treatment Reduced the Oviposition and Hatching Rates of Tea Green Leafhoppers Without Affecting Their Selectivity and Survival
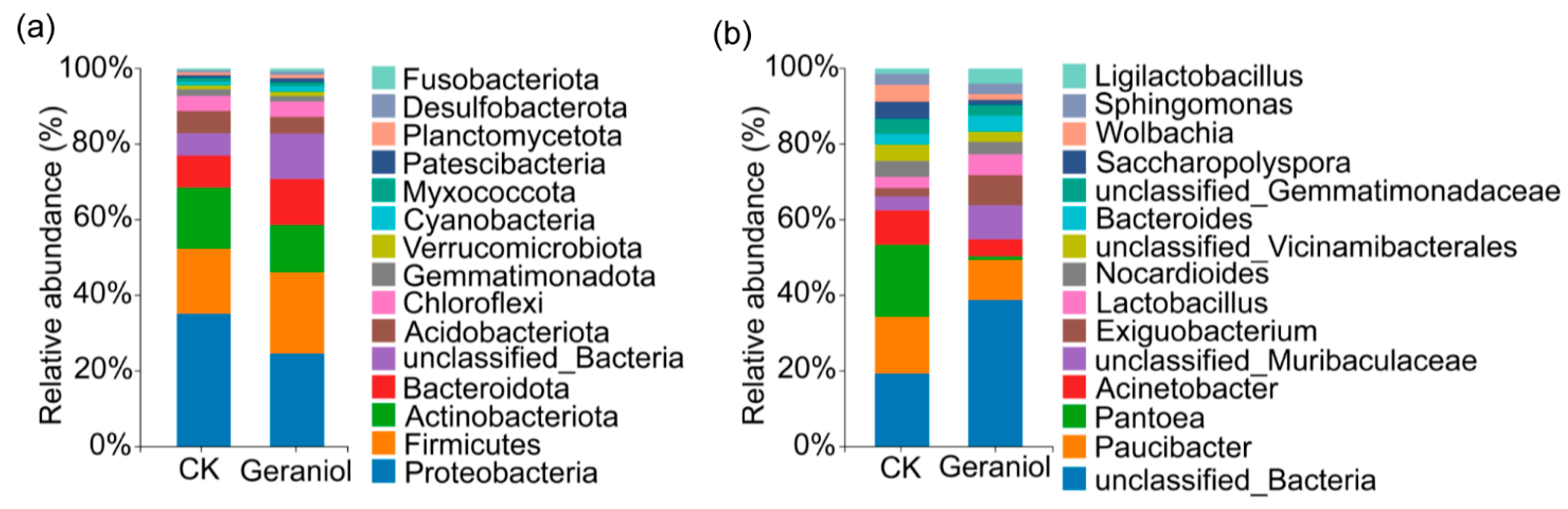
3.2. Effect of Geraniol Treatment on the Endophyte Community of Tea Green Leafhoppers
3.2.1. Geraniol Treatment Altered the Endophyte Community of Tea Green Leafhoppers
3.2.2. Geraniol Treatment Altered the Alpha and Beta Diversity of Endophytes in Tea Green Leafhoppers
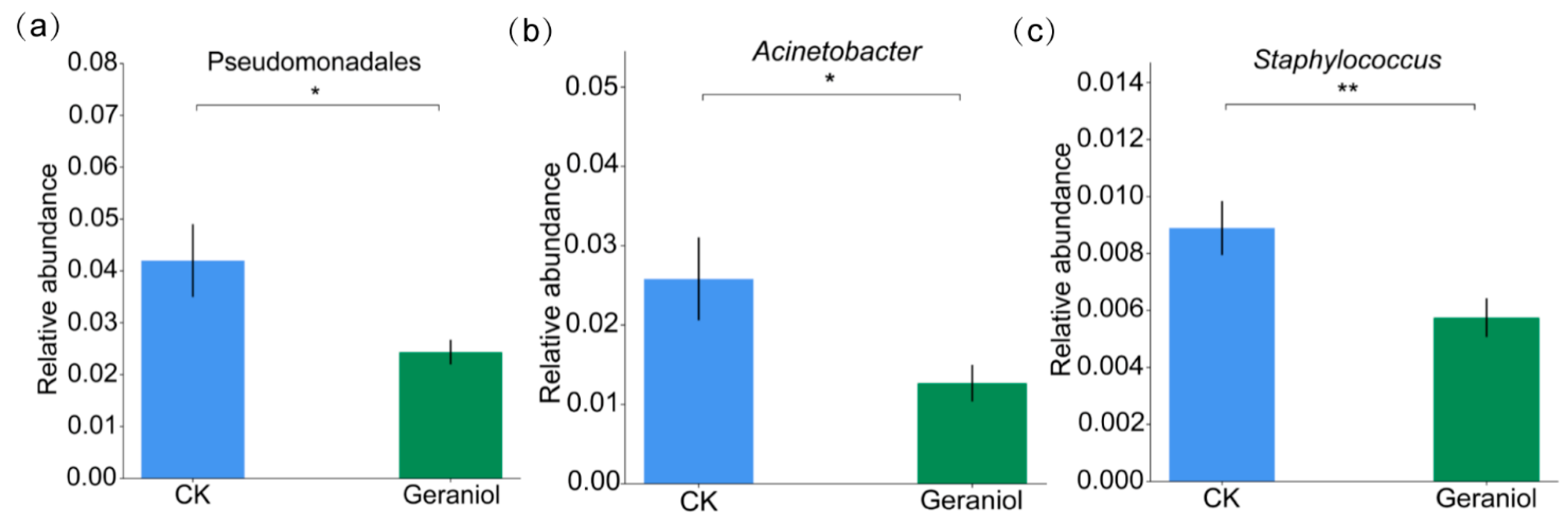
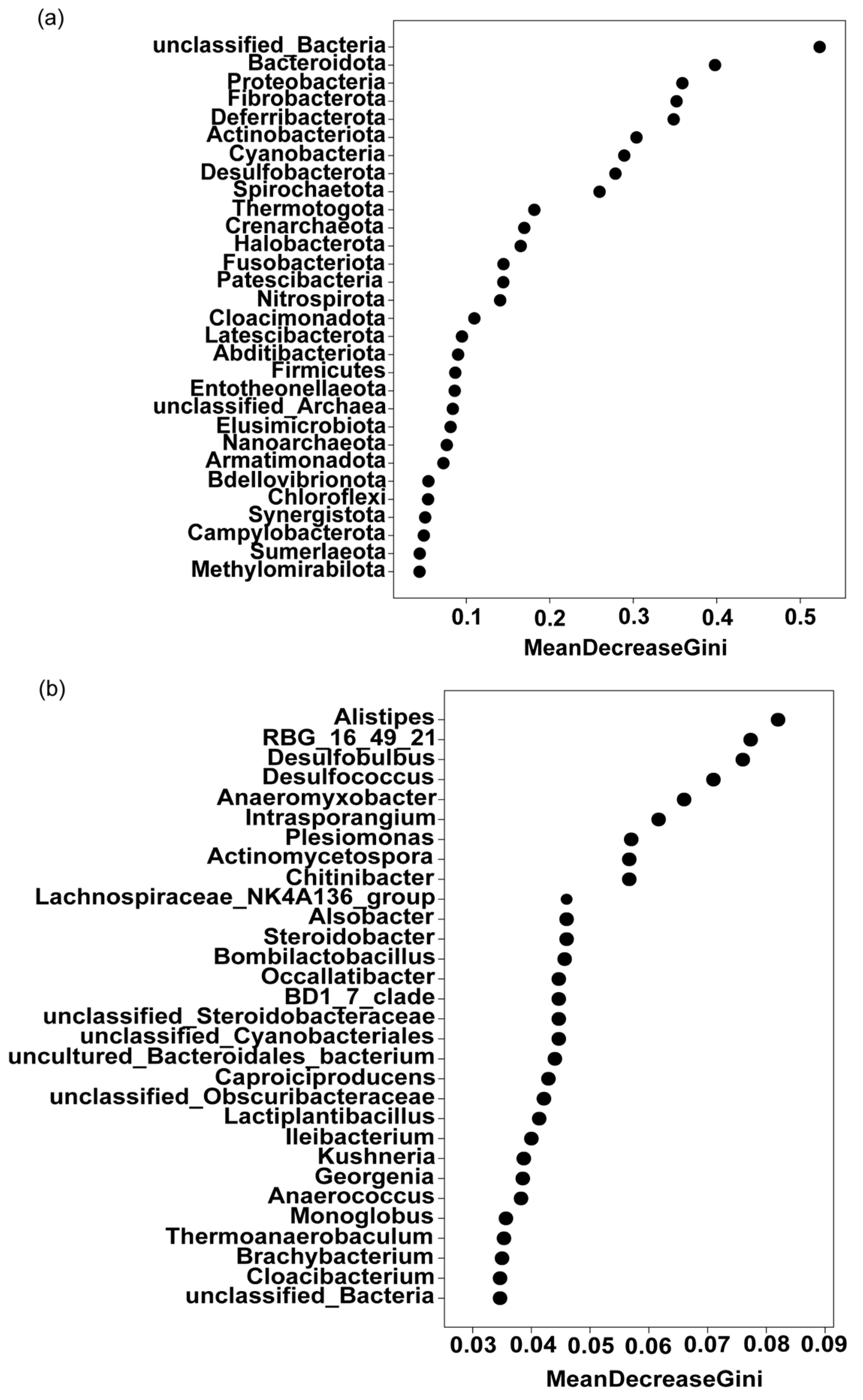
3.2.3. Analysis of Key Differential Endophytic Flora of Tea Green Leafhoppers
3.3. Geraniol Treatment Activated the Expression of Defense-Related Genes in Tea Green Leafhoppers
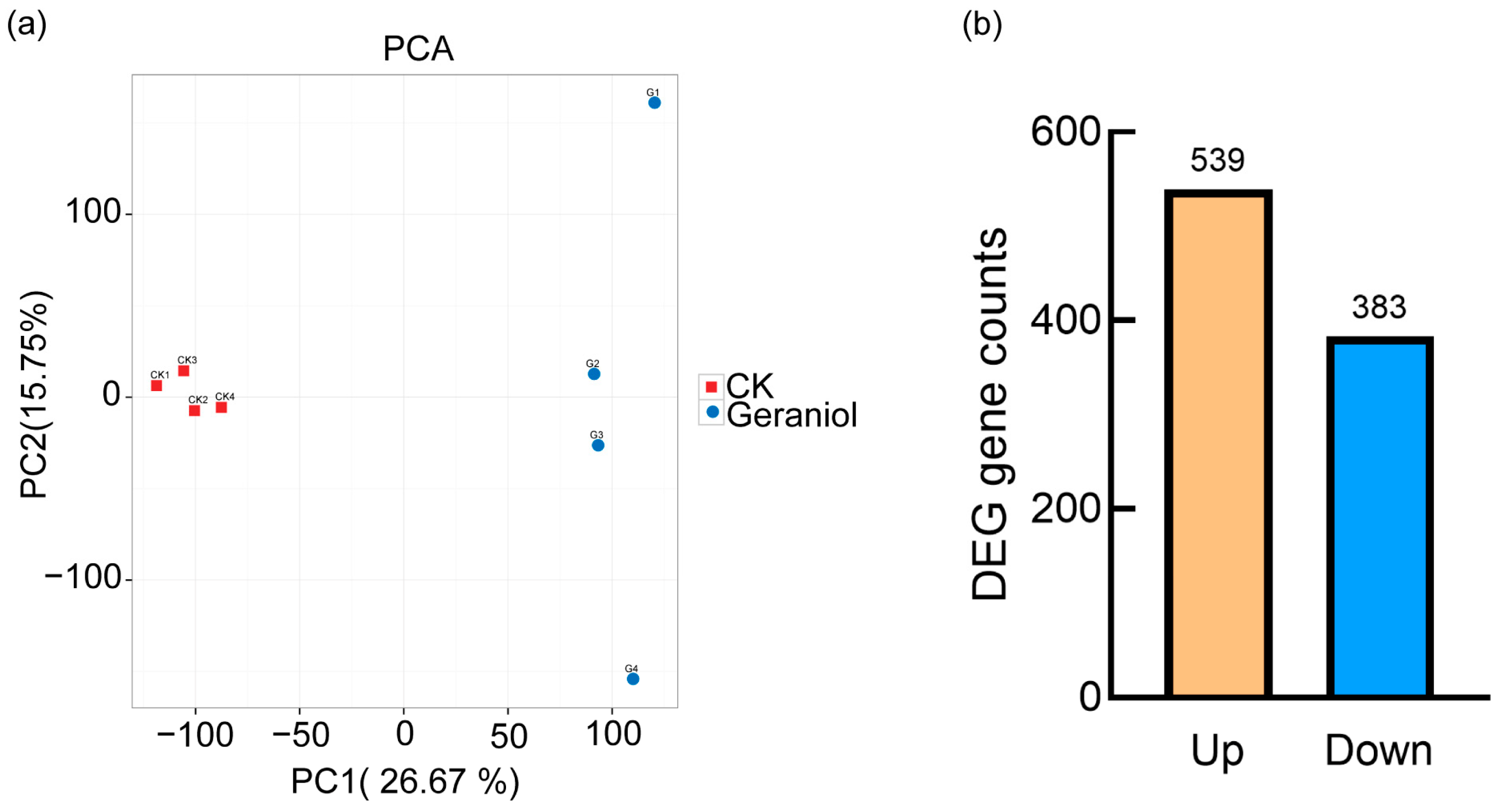
3.3.1. Transcriptome Quality of Tea Green Leafhoppers
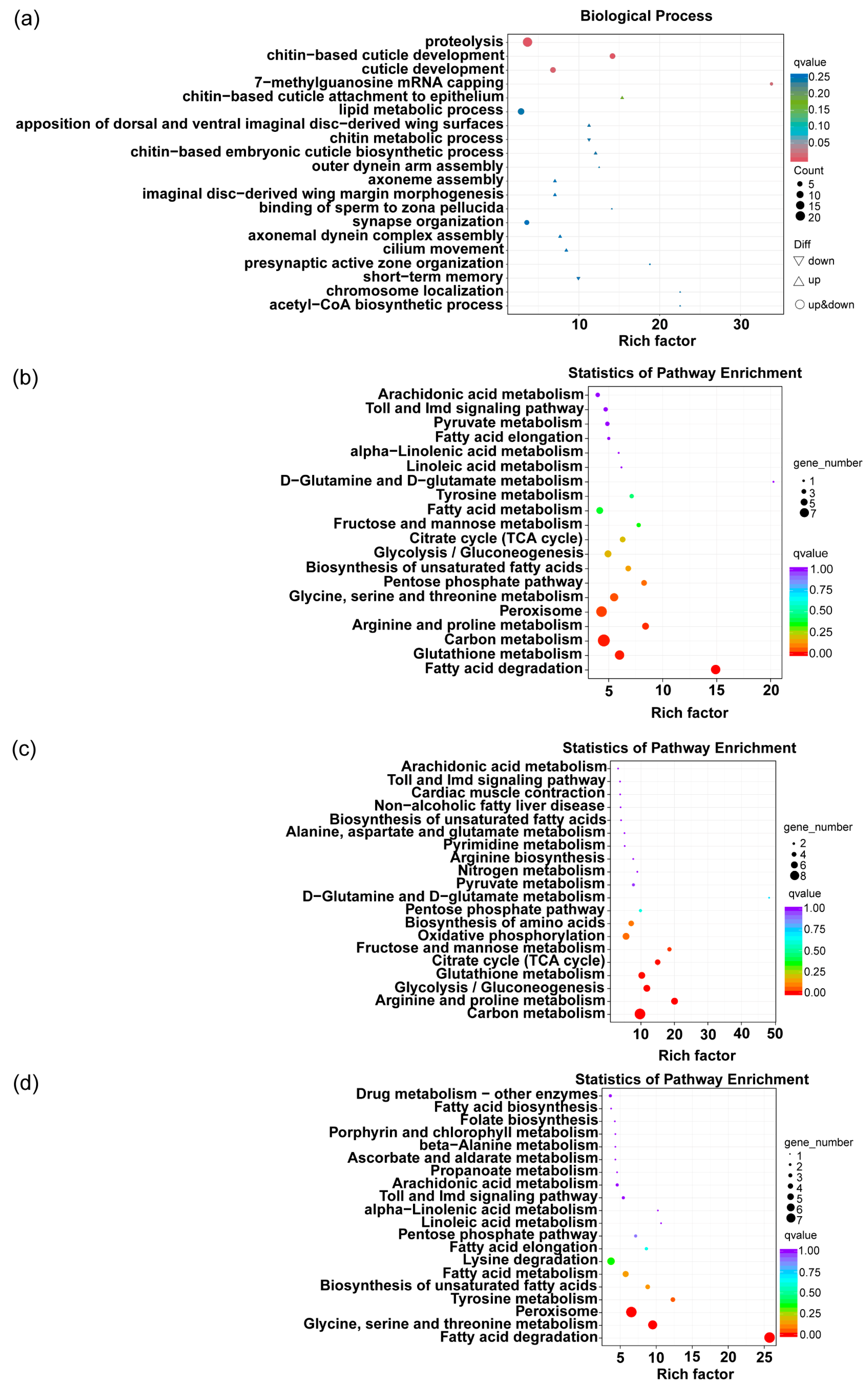
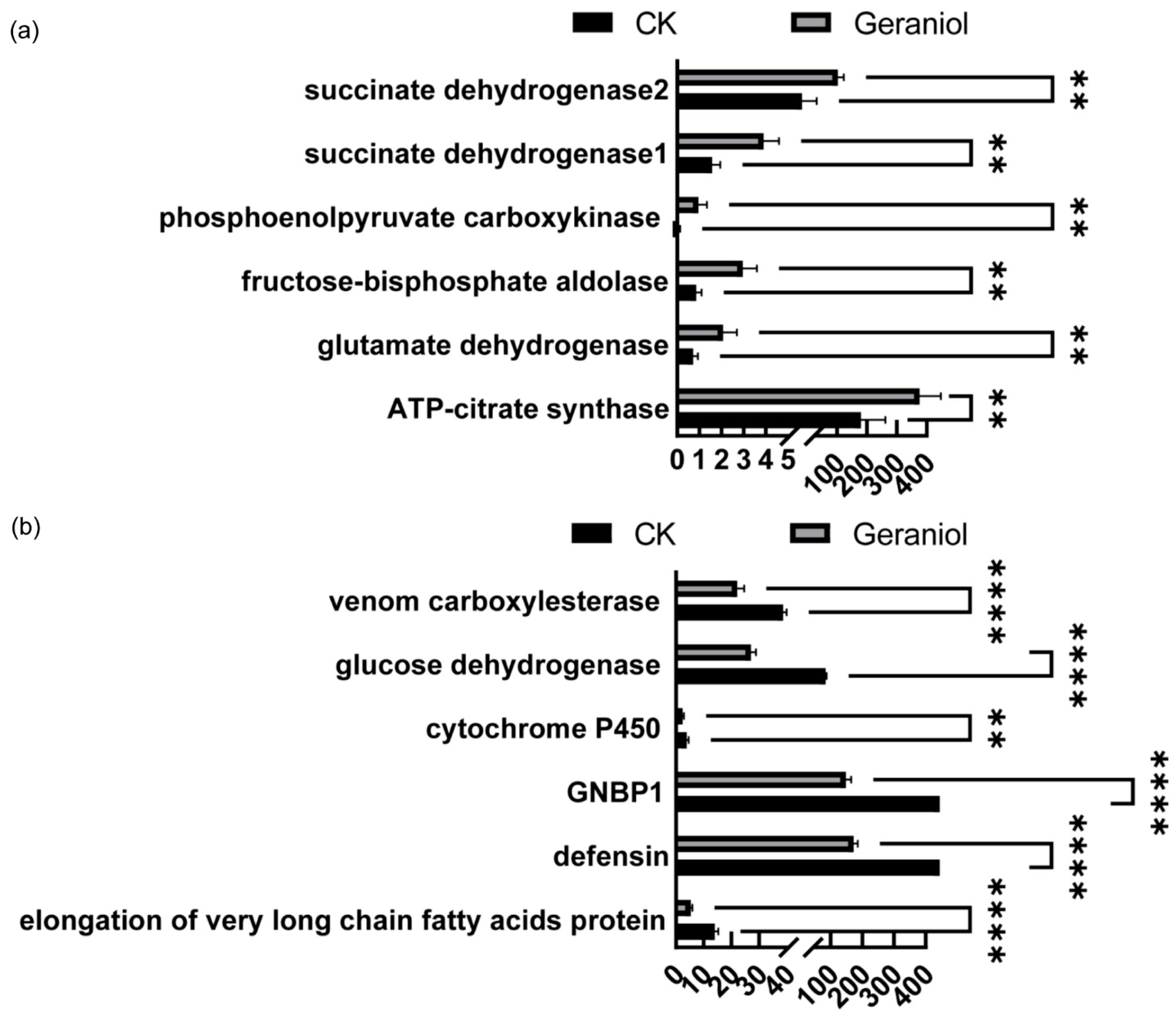
3.3.2. Transcriptome Analysis of Tea Green Leafhoppers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Lin, G.; Batool, K.; Zhang, S.; Chen, M.; Xu, J.; Wu, J.; Jin, L.; Gelbic, I.; Xu, L.; et al. Alimentary Tract Transcriptome Analysis of the Tea Geometrid, Ectropis oblique (Lepidoptera: Geometridae). J. Econ. Entomol. 2018, 111, 1411–1419. [Google Scholar] [CrossRef]
- Wakamura, S.; Yasuda, T.; Mochizuki, F. Mating behavior of the tea tussock moth, Euproctis pseudoconspersa (strand) (Lepidoptera: Lymantriidae). Appl. Entomol. Zool. 1996, 31, 619–621. [Google Scholar] [CrossRef]
- Zhang, R.; Lun, X.; Zhang, Y.; Zhao, Y.; Xu, X.; Zhang, Z. Characterization of Ionotropic Receptor Gene EonuIR25a in the Tea Green Leafhopper, Empoasca onuki Matsuda. Plants 2023, 12, 2034. [Google Scholar] [CrossRef]
- Lu, C.; Shen, N.; Jiang, W.; Xie, B.; Zhao, R.; Zhou, G.; Zhao, D.; He, Y.; Chen, W. Different Tea Germplasms Distinctly Influence the Adaptability of Toxoptera aurantii (Hemiptera: Aphididae). Insects 2023, 14, 695. [Google Scholar] [CrossRef]
- Hazarika, L.K.; Bhuyan, M.; Hazarika, B.N. Insect pests of tea and their management. Annu. Rev. Entomol. 2009, 54, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Kanu, U.C.; Wang, Z.; Qiu, C.; Wen, Q.; Li, X.; Qiu, D.; Gan, Y.; Mao, R. Redefining the tea green leafhopper: Empoasca onukii Matsuda (Hemiptera: Cicadellidae) as a vital asset in premium tea production. Life 2025, 15, 133. [Google Scholar] [CrossRef]
- Roy, C.; Naskar, S.; Ghosh, S.; Rahaman, P.; Mahanta, S.; Sarkar, N.; Chaudhuri, R.K.; Babu, A.; Roy, S.; Chakraborti, D. Sucking pest management in tea (Camellia sinensis (L.) Kuntze) cultivation: Integrating conventional methods with bio-control strategies. Crop Prot. 2024, 183, 106759. [Google Scholar] [CrossRef]
- Jin, S.; Chen, Z.M.; Backus, E.A.; Sun, X.L.; Xiao, B. Characterization of EPG waveforms for the tea green leafhopper, Empoasca vitis Gothe (Hemiptera: Cicadellidae), on tea plants and their correlation with stylet activities. J. Insect Physiol. 2012, 58, 1235–1244. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H.; Wang, D.; Wu, G.; Wang, Q. Effects of Artemisia argyi volatiles on the behavior of the tea green leafhopper (Empoasca onukii) in tea plantations. Chin. J. Appl. Entomol. 2022, 59, 773–784. [Google Scholar] [CrossRef]
- Wei, Q.; Mu, X.-C.; Yu, H.-Y.; Niu, C.-D.; Wang, L.-X.; Zheng, C.; Chen, Z.; Gao, C.-F. Susceptibility of Empoasca vitis (Hemiptera: Cicadellidae) populations from the main tea-growing regions of China to thirteen insecticides. Crop Prot. 2017, 96, 204–210. [Google Scholar] [CrossRef]
- Pu, X.-Y.; Feng, M.-G.; Shi, C.-H. Impact of three application methods on the field efficacy of a Beauveria bassiana-based mycoinsecticide against the false-eye leafhopper, Empoasca vitis (Homoptera: Cicadellidae) in the tea canopy. Crop Prot. 2005, 24, 167–175. [Google Scholar] [CrossRef]
- Ye, G.-Y.; Xiao, Q.; Chen, M.; Chen, X.-x.; Yuan, Z.-J.; Stanley, D.W.; Hu, C. Tea: Biological control of insect and mite pests in China. Biol. Control. 2014, 68, 73–91. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef]
- Szendrei, Z.; Rodriguez-Saona, C. A meta-analysis of insect pest behavioral manipulation with plant volatiles. Entomol. Exp. Appl. 2010, 134, 201–210. [Google Scholar] [CrossRef]
- Song, H.; Dong, Z.; Li, L.; Lu, Z.; Li, C.; Yu, Y.; Men, X. Relationships among the feeding behaviors of a mirid bug on cotton leaves of different ages and plant biochemical substances. J. Insect Sci. 2021, 21, 15. [Google Scholar] [CrossRef]
- Liu, C.M.; Matsuyama, S.; Kainoh, Y.A.-O. Synergistic effects of volatiles from host-infested plants on host-searching behavior in the parasitoid wasp Lytopylus rufipes (Hymenoptera:Braconidae). J. Chem. Ecol. 2019, 45, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Veyrat, N.; Robert, C.A.M.; Xu, H.; Frey, M.; Ton, J.; Turlings, T.C.J. Indole is an essential herbivore-induced volatile priming signal in maize. Nat. Commun. 2015, 6, 6273. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Huang, S.; Jiang, T.; Wang, C.; Tao, Z.; He, C.; Tang, Q.; Li, P. Volatile DMNT directly protects plants against Plutella xylostella by disrupting the peritrophic matrix barrier in insect midgut. eLife 2021, 10, e63938. [Google Scholar] [CrossRef]
- Gasmi, L.; Martínez-Solís, M.; Frattini, A.; Ye, M.; Collado, M.C.; Turlings, T.C.J.; Erb, M.; Herrero, S. Can Herbivore-Induced Volatiles Protect Plants by Increasing the Herbivores’ Susceptibility to Natural Pathogens? Appl. Environ. Microbiol. 2018, 85, e01468-18. [Google Scholar] [CrossRef]
- von Mérey, G.; Veyrat, N.; Mahuku, G.; Valdez, R.L.; Turlings, T.C.J.; D’Alessandro, M. Dispensing synthetic green leaf volatiles in maize fields increases the release of sesquiterpenes by the plants, but has little effect on the attraction of pest and beneficial insects. Phytochemistry 2011, 72, 1838–1847. [Google Scholar] [CrossRef]
- Ye, M.A.-O.; Glauser, G.A.-O.; Lou, Y.A.-O.; Erb, M.A.-O.; Hu, L.A.-O. Molecular dissection of early defense signaling underlying volatile-mediated defense regulation and herbivore resistance in rice. Plant Cell 2019, 31, 687–698. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.R.; Rodriguez-Saona, L.E.; Frost, C.J. Herbivore-induced volatiles in the perennial shrub, vaccinium corymbosum, and their role in inter-branch signaling. J. Chem. Ecol. 2009, 35, 163–175. [Google Scholar] [CrossRef]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Zakir, A.; Bengtsson, M.; Sadek, M.M.; Hansson, B.S.; Witzgall, P.; Anderson, P. Specific response to herbivore-induced de novo synthesized plant volatiles provides reliable information for host plant selection in a moth. J. Exp. Biol. 2013, 216, 3257–3263. [Google Scholar] [CrossRef] [PubMed]
- Naskar, S.; Roy, C.; Ghosh, S.; Mukhopadhyay, A.; Hazarika, L.K.; Chaudhuri, R.K.; Roy, S.; Chakraborti, D. Elicitation of biomolecules as host defense arsenals during insect attacks on tea plants (Camellia sinensis (L.) Kuntze). Appl. Microbiol. Biotechnol. 2021, 105, 7187–7199. [Google Scholar] [CrossRef]
- Wang, M.; Han, S.; Wu, Y.; Lin, J.; Zhou, J.; Han, B. Tea green leafhopper-induced synomone attracts the egg parasitoids, mymarids to suppress the leafhopper. Pest Manag. Sci. 2023, 79, 3785–3795. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Naseem, T.; Holopainen, J.K.; Liu, T.; Zhang, J.; Zhang, F. Tritrophic interactions among arthropod natural enemies, herbivores and plants considering volatile blends at different scale levels. Cells. 2023, 12, 251. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, X.; Yang, Z. Characterization of terpene synthase from tea green leafhopper being involved in formation of geraniol in tea (Camellia sinensis) leaves and potential effect of geraniol on insect-derived endobacteria. Biomolecules 2019, 9, 808. [Google Scholar] [CrossRef] [PubMed]
- de Lira, M.H.P.; de Andrade Júnior, F.P.; Moraes, G.F.Q.; da Silva Macena, G.; de Oliveira Pereira, F.; Lima, I.O. Antimicrobial activity of geraniol: An integrative review. J. Essent. Oil Res. 2020, 32, 187–197. [Google Scholar] [CrossRef]
- Ramadan, G.R.M.; Abdelgaleil, S.A.M.; Shawir, M.S.; El-bakary, A.S.; Zhu, K.Y.; Phillips, T.W. Terpenoids, DEET and short chain fatty acids as toxicants and repellents for Rhyzopertha dominica (Coleoptera: Bostrichidae) and Lasioderma serricorne (Coleoptera: Ptinidae). J. Stored Prod. Res. 2020, 87, 101610. [Google Scholar] [CrossRef]
- Mu, D.; Cui, L.; Ge, J.; Wang, M.X.; Liu, L.F.; Yu, X.P.; Zhang, Q.H.; Han, B.Y. Behavioral responses for evaluating the attractiveness of specific tea shoot volatiles to the tea green leafhopper, Empoaca vitis. Insect Sci. 2012, 19, 229–238. [Google Scholar] [CrossRef]
- Yao, Q.; Wang, M.; Chen, Z. The relative preference of Empoasca onukii (Hemiptera: Cicadellidae) for oviposition on twenty-four tea cultivars. J. Econ. Entomol. 2022, 115, 1521–1530. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. Public Libr. Sci. Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Wu, S.; Zhang, Z.; Li, Y. Global Geographic Diversity and Distribution of the Myxobacteria. Microbiol. Spectr. 2021, 9, e0001221. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal properties of several essential oils on the house fly (Musca domestica L.). Phytother. Res. 2008, 22, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Yarou, B.B.; Bawin, T.; Boullis, A.; Heukin, S.; Lognay, G.; Verheggen, F.J.; Francis, F. Oviposition deterrent activity of basil plants and their essentials oils against Tuta absoluta (Lepidoptera: Gelechiidae). Environ. Sci. Pollut. Res. Int. 2018, 25, 29880–29888. [Google Scholar] [CrossRef] [PubMed]
- Wagan, T.; He, Y.; Long, M.; Chakira, H.; Zhao, J.; Hua, H. Effectiveness of aromatic plant species for repelling and preventing oviposition of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). J. Appl. Entomol. 2018, 142, 287–295. [Google Scholar] [CrossRef]
- Erdemir, T.; Erler, F. Repellent, oviposition-deterrent and egg-hatching inhibitory effects of some plant essential oils against citrus mealybug, Planococcus citri Risso (Hemiptera: Pseudococcidae). J. Plant Dis. Prot. 2017, 124, 473–479. [Google Scholar] [CrossRef]
- Douglas, A.E. Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic bacteria buchnera. Annu. Rev. Entomology. 1998, 43, 17–37. [Google Scholar] [CrossRef]
- Boone, C.K.; Keefover-Ring, K.; Mapes, A.C.; Adams, A.S.; Bohlmann, J.; Raffa, K.F. Bacteria associated with a tree-killing insect reduce concentrations of plant defense compounds. J. Chem. Ecol. 2013, 39, 1003–1006. [Google Scholar] [CrossRef]
- Li, T.; Xiao, J.H.; Wu, Y.Q.; Huang, D.W. Diversity of bacterial symbionts in populations of Sitobion miscanthi (Hemiptera: Aphididae) in China. Environ. Entomol. 2014, 43, 605–611. [Google Scholar] [CrossRef]
- Xu, T.T.; Chen, J.; Jiang, L.Y.; Qiao, G.A.-O. Diversity of bacteria associated with Hormaphidinae aphids (Hemiptera: Aphididae). Insect Sci. 2021, 28, 165–179. [Google Scholar] [CrossRef]
- Duan, X.Z.; Sun, J.T.; Wang, L.T.; Shu, X.H.; Guo, Y.; Keiichiro, M.; Zhu, Y.X.; Bing, X.L.; Hoffmann, A.A.; Hong, X.A.-O. Recent infection by Wolbachia alters microbial communities in wild Laodelphax striatellus populations. Microbiome 2020, 8, 104. [Google Scholar] [CrossRef]
- Xia, X.; Wang, Q.; Gurr, G.M.; Vasseur, L.; Han, S.; You, M. Gut bacteria mediated adaptation of diamondback moth, Plutella xylostella, to secondary metabolites of host plants. mSystems 2023, 8, e00826-23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Huang, X.-Y.; Zi, H.-B.; Gao, T.; Ji, R.-J.; Sheng, J.; Zhi, D.; Zhang, Y.-L.; Gong, C.-M.; et al. Altitude as a key environmental factor shaping microbial communities of tea green leafhoppers (Matsumurasca onukii). Microbiol. Spectr. 2023, 11, e0100923. [Google Scholar] [CrossRef]
- Tang, T.; Zhang, Y.; Cai, T.; Deng, X.; Liu, C.; Li, J.; He, S.; Li, J.; Wan, H. Antibiotics increased host insecticide susceptibility via collapsed bacterial symbionts reducing detoxification metabolism in the brown planthopper, Nilaparvata lugens. J. Pest Sci. 2021, 94, 757–767. [Google Scholar] [CrossRef]
- Wu, C.; Ding, C.; Chen, S.; Wu, X.; Zhang, L.; Song, Y.; Li, W.; Zeng, R.A.-O. Exposure of Helicoverpa armigera larvae to plant volatile organic compounds induces cytochrome P450 monooxygenases and enhances larval tolerance to the insecticide methomyl. Insects 2021, 12, 238. [Google Scholar] [CrossRef]
- Hederstedt, L.; Rutberg, L. Succinate dehydrogenase–A comparative review. Microbiol. Rev. 1981, 45, 542–555. [Google Scholar] [CrossRef]
- Bond, P.A.; Sang, J.H. Glutamate dehydrogenase of Drosophila larvae. J. Insect Physiol. 1968, 14, 341–359. [Google Scholar] [CrossRef]
- Pannebakker, B.A.; Trivedi, U.; Blaxter, M.A.; Watt, R.; Shuker, D.M. The transcriptomic basis of oviposition behaviour in the parasitoid Wasp Nasonia vitripennis. PLoS ONE 2013, 8, e68608. [Google Scholar] [CrossRef]
- Tanji, T.; Hu, X.; Weber, A.N.R.; Ip, Y.T. Toll and IMD pathways synergistically activate an innate immune response in Drosophila melanogaster. Mol. Cell. Biol. 2007, 27, 4578–4588. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, J.; Hong, H.; Mo, S.; Mo, S.; Shi, H.; Shi, J.; Zhou, Y. Effects of Geraniol on Survival, Reproduction, Endophytes, and Transcriptome of Tea Green Leafhoppers (Empoasca onukii). Agronomy 2025, 15, 782. https://doi.org/10.3390/agronomy15040782
Tian J, Hong H, Mo S, Mo S, Shi H, Shi J, Zhou Y. Effects of Geraniol on Survival, Reproduction, Endophytes, and Transcriptome of Tea Green Leafhoppers (Empoasca onukii). Agronomy. 2025; 15(4):782. https://doi.org/10.3390/agronomy15040782
Chicago/Turabian StyleTian, Junjie, Hainuo Hong, Shiliang Mo, Shiqin Mo, Hongliang Shi, Juan Shi, and Ying Zhou. 2025. "Effects of Geraniol on Survival, Reproduction, Endophytes, and Transcriptome of Tea Green Leafhoppers (Empoasca onukii)" Agronomy 15, no. 4: 782. https://doi.org/10.3390/agronomy15040782
APA StyleTian, J., Hong, H., Mo, S., Mo, S., Shi, H., Shi, J., & Zhou, Y. (2025). Effects of Geraniol on Survival, Reproduction, Endophytes, and Transcriptome of Tea Green Leafhoppers (Empoasca onukii). Agronomy, 15(4), 782. https://doi.org/10.3390/agronomy15040782