
Exploring the Potential of Mediterranean Edible Halophytes as Novel Crops: Ecological and Nutritional Insights from Tuscany’s Salt Marshes




Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Literature Survey
2.3. Field Monitoring
2.4. Data Processing
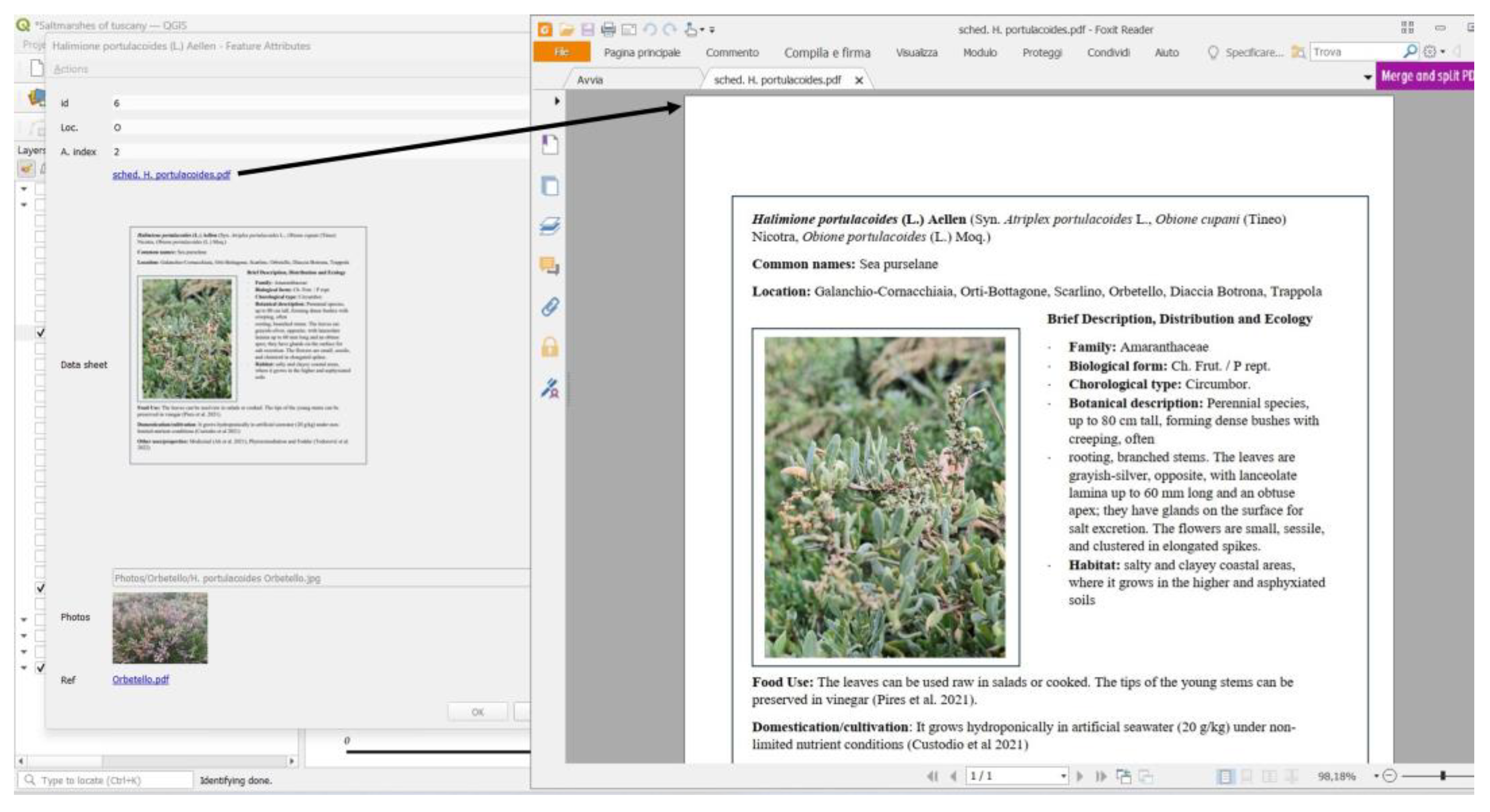
2.5. QGIS Interactive Maps
2.6. Statistical Analysis
3. Results and Discussion
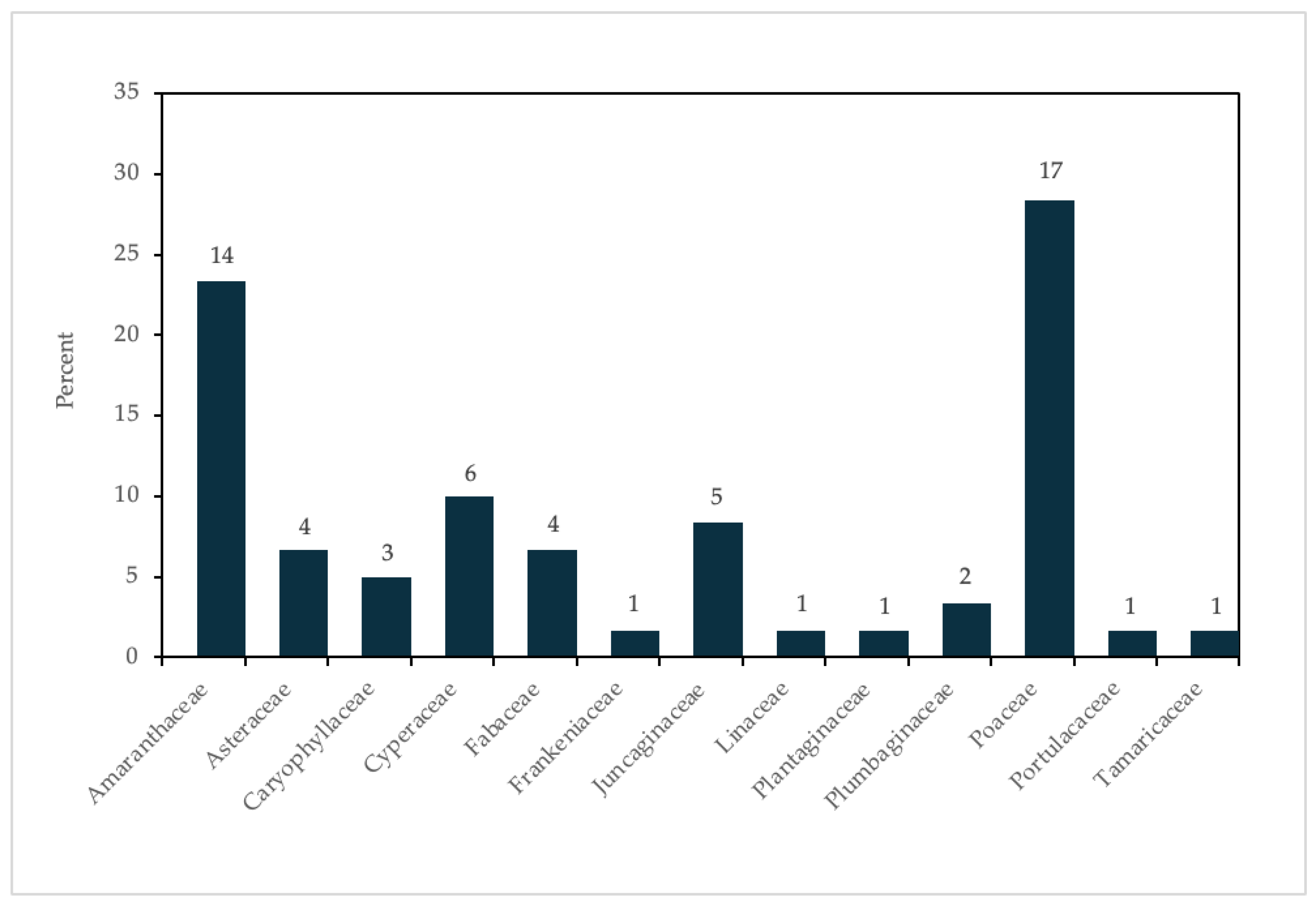
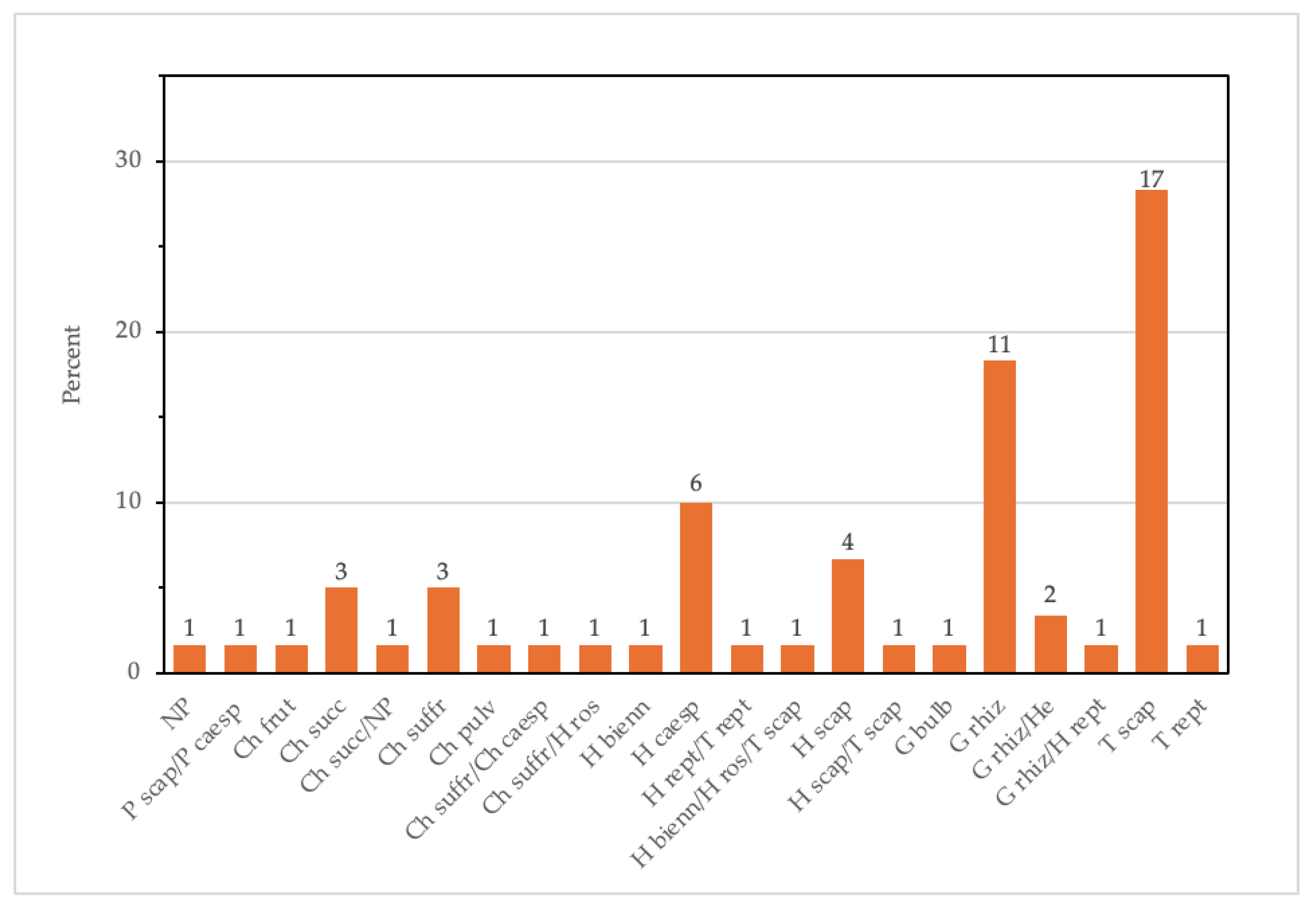
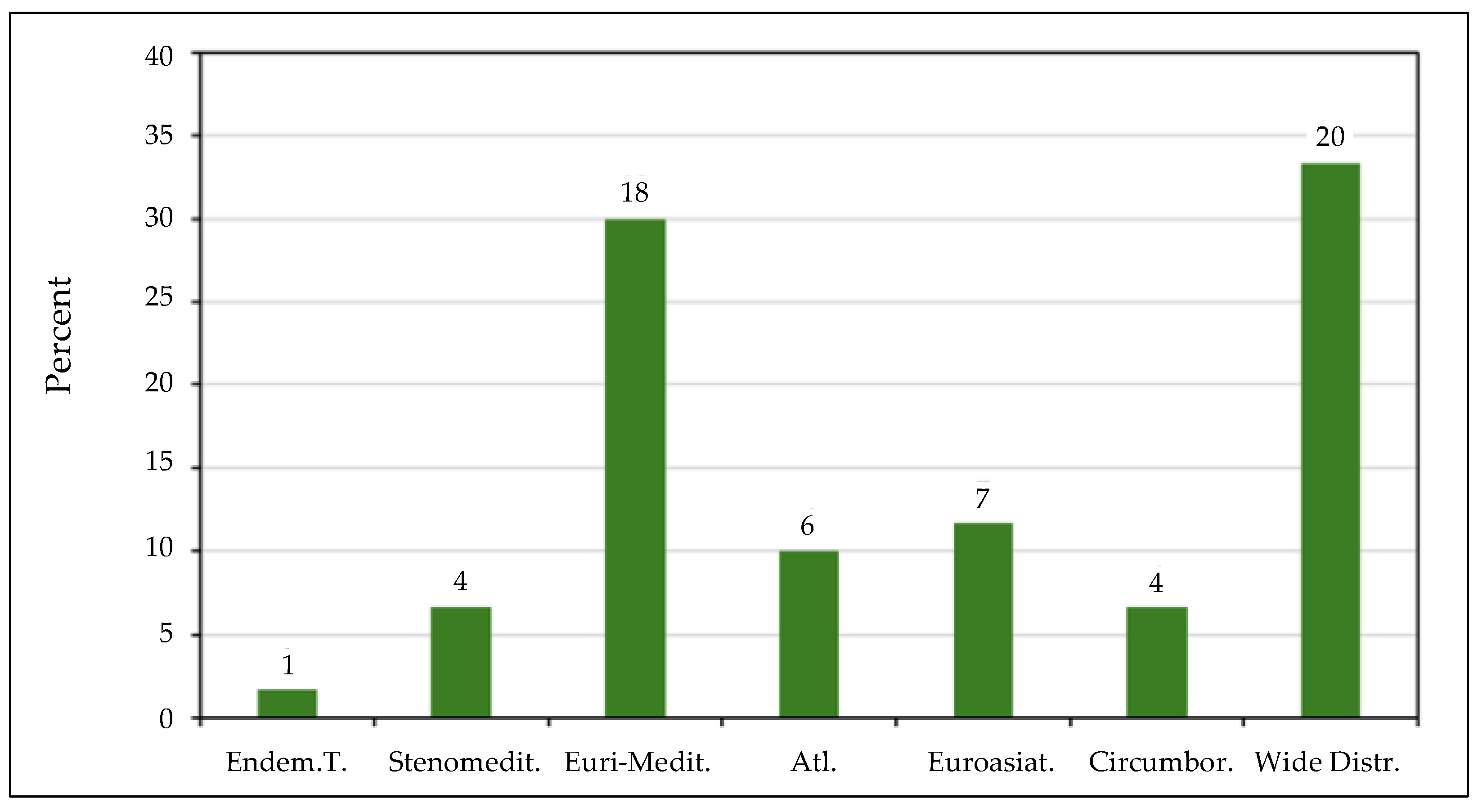
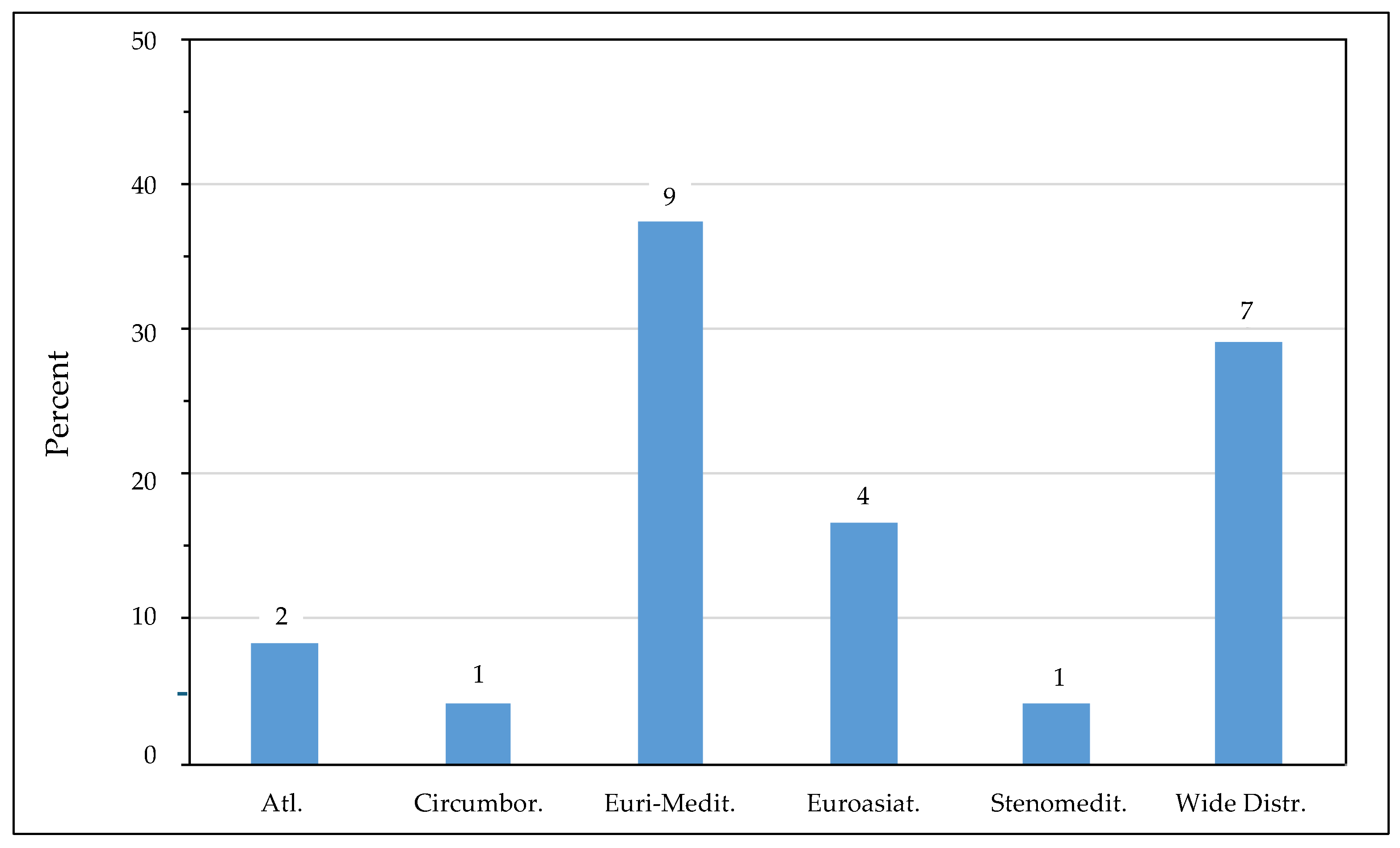
3.1. Literature Survey
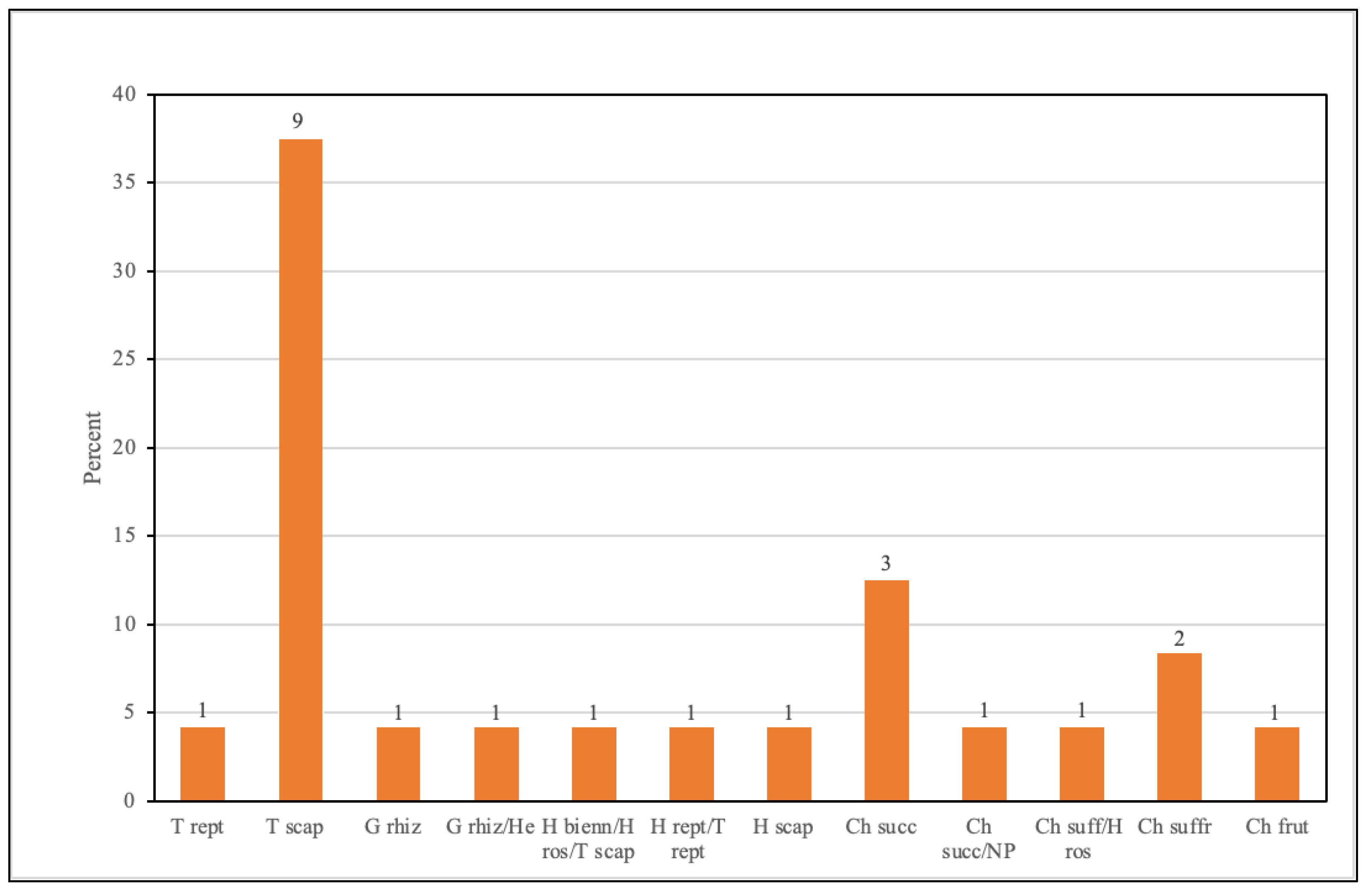
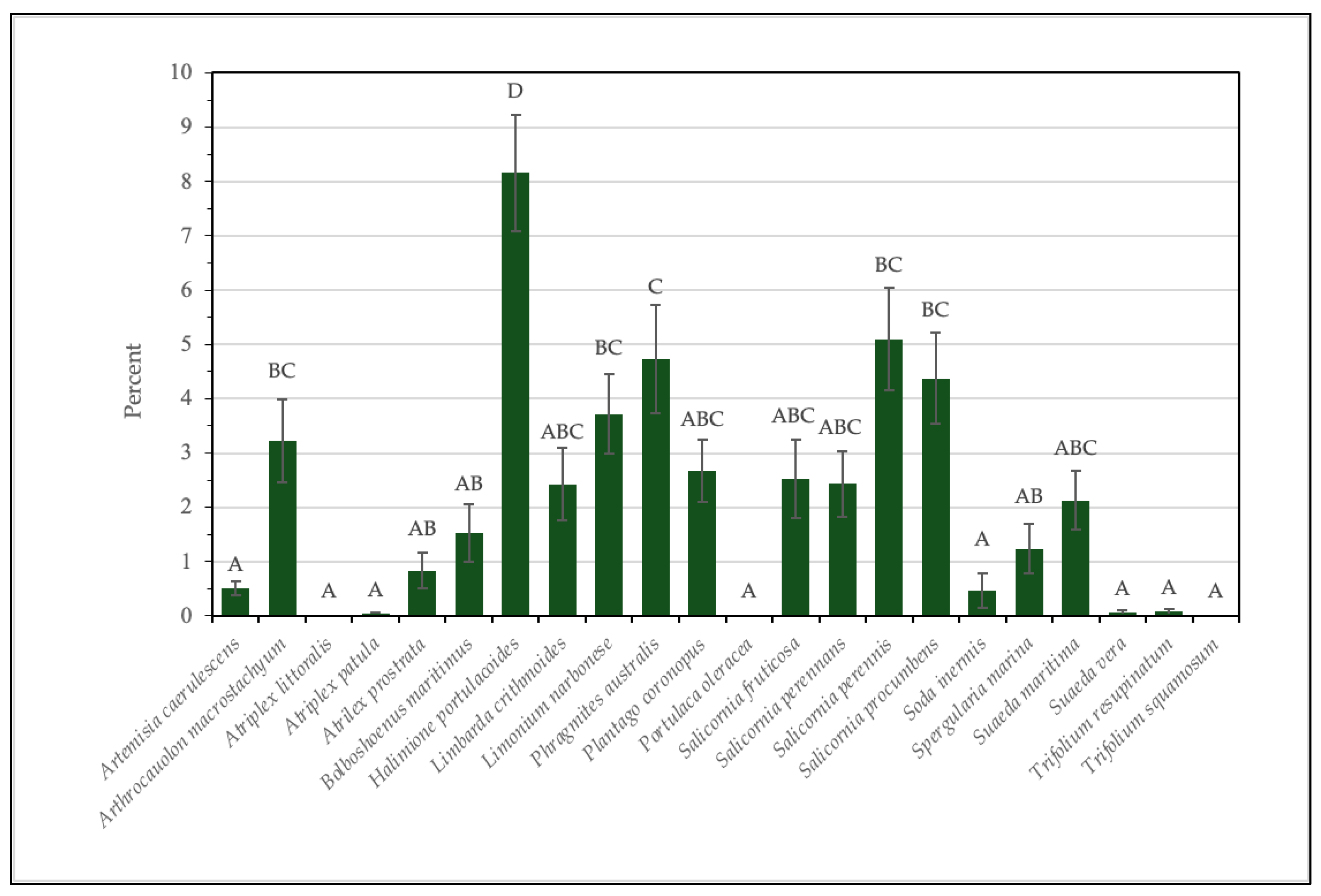
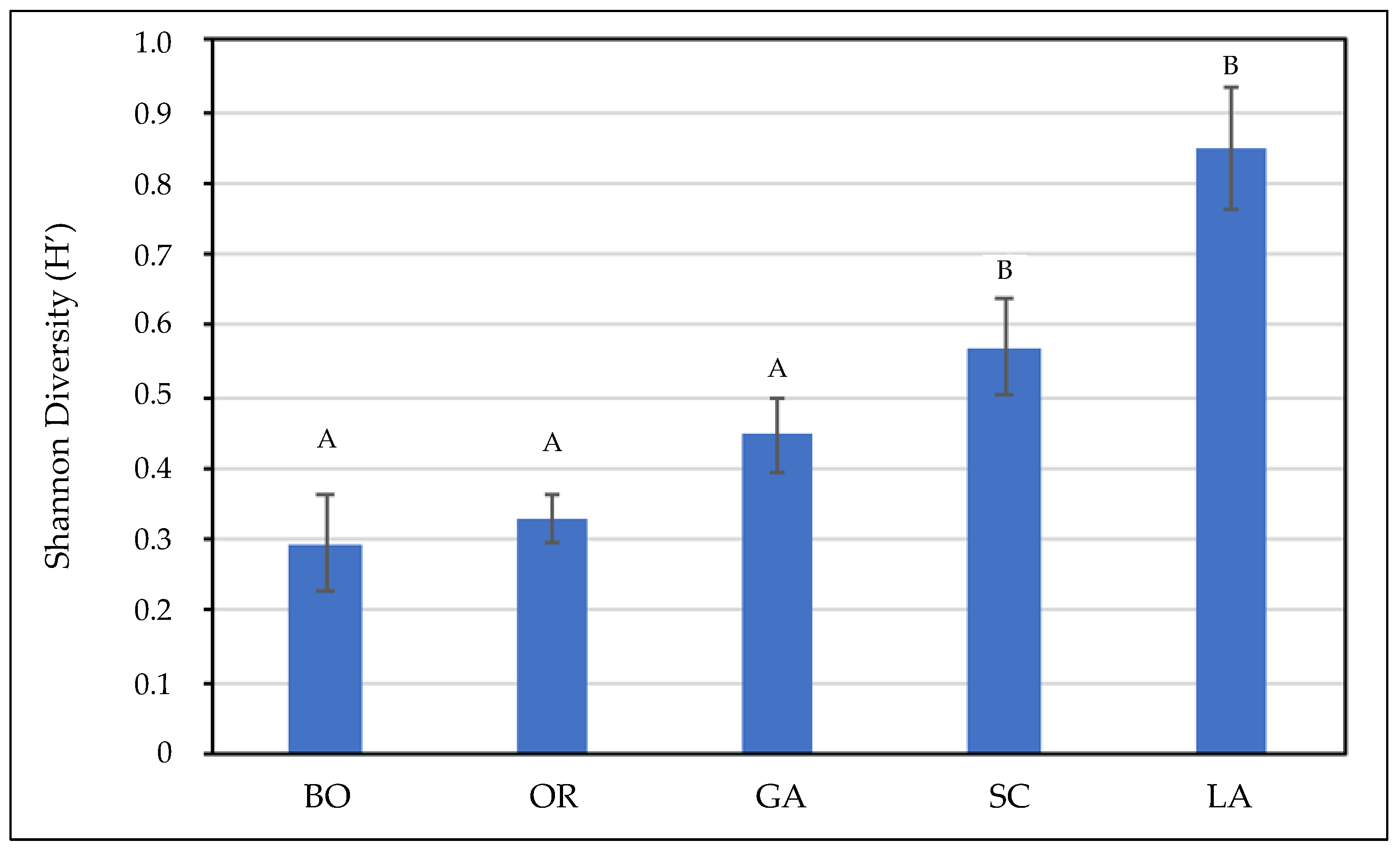
3.2. Field Surveys and Data Processing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family/Plant Name | Cs | Life Form | Chorotype | Presence/Absence | Edible Parts | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| La | Ga | Bo | Sc | Or | Di | Tr | |||||
| Amaranthaceae | |||||||||||
| Arthrocaulon macrostachyum (Moric.) Piirainen and G.Kadereit | Ch succ | Medit.-Macaron. | * | * | * | * | Shoots [76] | ||||
| Atriplex littoralis L. | T scap | Europ. | * | * | * | Leaves, whole plant [77] | |||||
| Atriplex patula L. subsp. patula | T scap | Subcosmop. | * | * | * | Leaves [78] | |||||
| Atriplex prostrata Boucher ex DC. | T scap | Paleotemp. | * | * | * | * | * | * | * | Leaves, whole plant [79] | |
| Beta vulgaris L. subsp. maritima (L.) Arcang.* | LC | H scap | Euri-Medit. | * | * | Aerial parts [80] | |||||
| Halimione portulacoides (L.) Aellen | Ch frut/P rept | Circumbor. | * | * | * | * | * | * | Leaves [81] | ||
| Halocnemum cruciatum (Forssk.) Tod | Ch succ/NP | S-Medit | * | Shoots [82] | |||||||
| Salicornia fruticosa (L.) L. | Ch succ | Euri-Medit./Afric. | * | * | * | Shoots [83] | |||||
| Salicornia perennans Willd. subsp. perennans | T scap | Euri-Medit. | * | * | * | * | * | * | * | Shoots [84] | |
| Salicornia perennis Mill. | LC | Ch succ | Medit.-At. (Euri-) | * | * | * | * | * | * | * | Shoots [85] |
| Salicornia procumbens Sm. Subsp. procumbens | T scap | Steno-Medit | * | * | Shoots [86] | ||||||
| Soda inermis Fourr. | T scap | Paleotemp. | * | * | * | * | * | Leaves [87] | |||
| Suaeda maritima (L.) Dumort. | T scap | Cosmop. | * | * | * | * | * | * | * | Leaves [88] | |
| Suaeda vera J.F. Gmel. | NP | Cosmop. | * | * | Leaves [87] | ||||||
| Asteraceae | |||||||||||
| Artemisia caerulescens L. | Ch suffr | Euri-Medit. | * | * | * | * | * | * | Leaves, flowers [89] | ||
| Limbarda crithmoides (L.) Dumort. subsp. longifolia (Arcang.) Greuter | Ch suffr | Medit.-Atl. (Steno-) | * | * | * | * | * | * | * | Leaves [90] | |
| Caryophyllaceae | |||||||||||
| Spergularia marina (L.) Besser | LC | T rept | Subcosmop. | * | * | * | * | * | * | * | Leaves, aereal part [91] |
| Cyperaceae | |||||||||||
| Bolboschoenus maritimus (L.) Palla | G rhiz | Cosmop. | * | * | * | * | Seeds, root [92] | ||||
| Fabaceae | |||||||||||
| Trifolium resupinatum L. | H rept/T rept | Paleotemp. | * | * | Leaves, flowers [93] | ||||||
| Trifolium squamosum L. | T scap | Euri-Medit. | * | * | Leaves, flowers [93] | ||||||
| Plantago coronopus L. | H bienn/H ros/T scap | Euri-Medit. | * | * | * | * | * | * | * | Leaves [94] | |
| Plumbaginaceae | |||||||||||
| Limonium narbonese Mill. | Ch suff/H ros | Euri-Medit. | * | * | * | * | * | * | * | Leaves [95] | |
| Poaceae | |||||||||||
| Phragmites australis L. | LC | G rhiz/He | Subcosmop. | * | * | * | * | * | * | Leaves, young shoots [96] | |
| Portulacaceae | |||||||||||
| Portulaca oleracea L. | LC | T scap | Subcosmop. | * | * | Leaves, stems, flowers, seeds [97] | |||||
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaman, M.; Shahid, S.A.; Heng, L.; Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Cham, Switzerland, 2018; pp. 43–53. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Byerlee, D. A Window of Opportunities for Poor Farmers: Investing for Long-Term Food Supply. Dev. Outreach 2008, 10, 9–12. [Google Scholar] [CrossRef]
- Shrestha, S. Effects of Climate Change in Agricultural Insect Pest. Acta Sci. Agric. 2019, 3, 74–80. [Google Scholar] [CrossRef]
- Byerlee, D. Policies to Promote Cereal Intensification in Ethiopia: A Review of Evidence and Experience. In IFPRI Discussion Paper. 707; International Food Policy Research Institute (IFPRI): Washington, DC, USA, 2007. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2007: Synthesis Report; Working Group Contribution to the Fourth Assessment Report of Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2007. [Google Scholar]
- Stavi, I. Food Security among Dryland Pastoralists and Agropastoralists: The Climate, Land-Use Change, and Population Dynamics Nexus. Anthr. Rev. 2022, 9, 299–323. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt Tolerance and Crop Potential of Halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Duarte, B. Nutritional Valuation and Food Safety of Endemic Mediterranean Halophytes Species Cultivated in Abandoned Salt Pans under a Natural Irrigation Scheme. Estuar. Coast. Shelf Sci. 2022, 265, 107733. [Google Scholar] [CrossRef]
- Negacz, K.; Vellinga, P. The Emergence of a Governance Landscape for Saline Agriculture. In Halt Soil Salinization, Boost Soil Productivity. In Proceedings of the Global Symposium on Salt-Affected Soils, Rome, Italy, 20 October 2022; pp. 284–285. [Google Scholar]
- Corrêa, R.C.G. Halophytes for Future Horticulture: The Case of Small-Scale Farming in the Mediterranean Basin. In Handbook of Halophytes: From Molecules to Ecosystems Towards Biosaline Agriculture; Springer Nature: Berlin, Germany, 2021; pp. 2367–2393. [Google Scholar]
- Anzidei, M. Coastal Structure, Sea-Level Changes and Vertical Motion of the Land in the Mediterranean. Geol. Soc. Lond. Spec. Publ. 2014, 388, 453–479. [Google Scholar] [CrossRef]
- Meli, M.; Camargo, C.M.; Olivieri, M.; Slangen, A.B.; Romagnoli, C. Sea-Level Trend Variability in the Mediterranean during the 1993–2019 Period. Front. Mar. Sci. 2023, 10, 1150488. [Google Scholar] [CrossRef]
- Vecchio, A.; Anzidei, M.; Serpelloni, E. Sea Level Rise Projections up to 2150 in the Northern Mediterranean Coasts. Environ. Res. Lett. 2024, 19, 014050. [Google Scholar] [CrossRef]
- Canfora, L.; Salvati, L.; Benedetti, A.; Dazzi, C.; Lo Papa, G. Saline soils in Italy: Distribution, ecological processes and socioeconomic issues. Riv. Econ. Agrar. 2017, 72, 63–77. [Google Scholar]
- Salvati, L. Towards a Process-Based Evaluation of Land Vulnerability to Soil Degradation in Italy. Ecol. Indic. 2011, 11, 1216–1227. [Google Scholar] [CrossRef]
- Costantini, E.A.; Dazzi, C. The Soils of Italy; Springer: Berlin, Germany, 2013; ISBN 94-007-5642-9. [Google Scholar]
- Dazzi, C. Saline Waters and Soil Quality [Italy]. Ital. J. Agron. 2006, 1, 467–474. [Google Scholar] [CrossRef]
- Costantini, E.; Urbano, F.; Aramini, G.; Barbetti, R.; Bellino, F.; Bocci, M.; Bonati, G.; Fais, A.; L’Abate, G.; Loj, G. Rationale and Methods for Compiling an Atlas of Desertification in Italy. Land Degrad. Dev. 2009, 20, 261–276. [Google Scholar] [CrossRef]
- Raspini, F.; Bianchini, S.; Ciampalini, A.; Del Soldato, M.; Montalti, R.; Solari, L.; Tofani, V.; Casagli, N. Persistent Scatterers Continuous Streaming for Landslide Monitoring and Mapping: The Case of the Tuscany Region (Italy). Landslides 2019, 16, 2033–2044. [Google Scholar] [CrossRef]
- Stragapede, F. LE COSTE: Caratteristiche, tendenze evolutive, erosione e interventi di difesa. In Monografie di geologia ambientale; Edizione SIGEA: Milano, Italy, 2023. [Google Scholar]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Sposimo, P.; Castelli, C. La Biodiversità in Toscana. Specifiee Habitat in Pericolo; Archivio Del Repertorio Naturalistico Toscano (RENATO): Firenze, Italy, 2005. [Google Scholar]
- Selvi, F. Diversity, Geographic Variation and Conservationof the Serpentine Flora of Tuscany (Italy). Biodivers. Conserv. 2007, 16, 1423–1439. [Google Scholar] [CrossRef]
- Foggi, B.; Viciani, D.; Baldini, R.M.; Carta, A.; Guidi, T. Conservation Assessment of the Endemic Plants of the Tuscan Archipelago, Italy. Oryx 2015, 49, 118–126. [Google Scholar] [CrossRef]
- Bedini, G.; Garbari, F.; Peruzzi, L. Chrobase. It–Chromosome Numbers for the Italian Flora. 2016. Available online: https://italianbotanist.pensoft.net/article/8818/ (accessed on 1 January 2025).
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E. An Updated Checklist of the Vascular Flora Native to Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S. An Updated Checklist of the Vascular Flora Alien to Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M. Red List of Threatened Vascular Plants in Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- Fiaschi, T.; Bonari, G.; Frignani, F.; Gizzi, G.; Landi, M.; Magrini, S.; Quilghini, G.; Pafumi, E.; Scoppola, A.; Angiolini, C. Vascular Flora of the Isthmus of Feniglia (Southern Tuscany, Italy). Ital. Bot. 2024, 17, 77–101. [Google Scholar] [CrossRef]
- Bertacchi, A.; Lombardi, T.; Saggese, A.; Lazzeri, V. The Vegetation of a Relict Salt Marsh Area in the Pisan Coast in the Context of Brackish Wetlands of Tuscany. Plant Sociol. 2021, 58, 41–53. [Google Scholar] [CrossRef]
- Gennai, M.; Angiolini, C.; Bertacchi, A.; Gabellini, A.; Sarmati, S.; Viciani, D.; Foggi, B. Studying Local Species Assemblages of Salt-Affected Vegetation for Monitoring Natura 2000 Habitats. Plant Sociol. 2022, 59, 1–10. [Google Scholar] [CrossRef]
- Ungaro, F.; Calzolari, C.; Fantappiè, M.; Napoli, R.; Barbetti, R.; Tarocco, P.; Staffilani, F.; Puddu, R.; Fanni, S.; Ragazzi, F. National Mapping and Reporting of Salt-Affected Soils of Italy; Food and Agriculture Organization: FAO, Rome, 2024. [Google Scholar] [CrossRef]
- Singh, A. A Review of Wastewater Irrigation: Environmental Implications. Resour. Conserv. Recycl. 2021, 168, 105454. [Google Scholar]
- Bedini, G.; Peruzzi, L. Wikiplantbase# Toscana-Verso Un Catalogo Collaborativo, Online e Gratuito Delle Piante Vascolari Di Toscana. In Riunione scientifica del Gruppo per la Floristica, Società Botanica Italiana, 18-19/10/2013, Roma; ITA: Rome, Italy, 2013. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982; Volume 13, ISBN 88-506-2449-2. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Milano, Italy, 2017; Volume 2019, pp. 1–4. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Alessandrini, A.; Ardenghi, N.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D. A Second Update to the Checklist of the Vascular Flora Native to Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2024, 158, 219–296. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Alessandrini, A.; Ardenghi, N.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D. A Second Update to the Checklist of the Vascular Flora Alien to Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2024, 158, 297–340. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a Database of Salt-Tolerant Plants: Helping Put Halophytes to Work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef]
- Renna, M.; Gonnella, M. Ethnobotany, Nutritional Traits, and Healthy Properties of Some Halophytes Used as Greens in the Mediterranean Basin. In Handbook of Halophytes: From Molecules to Ecosystems Towards Biosaline Agriculture; Springer Nature: Berlin/Heidelberg, Germany, 2020; pp. 1–19. [Google Scholar]
- Ortuño, J.A.; Verde, A.; Fajardo, J.; Rivera, D.; Obón, C.; Alcaraz, F. Halophytes in Arts and Crafts: Ethnobotany of Glassmaking. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture; Springer: Berlin/Heidelberg, Germany, 2021; pp. 2675–2706. [Google Scholar]
- Lombardi, T.; Ventura, I.; Bertacchi, A. Floristic Inventory of Ethnobotanically Important Halophytes of North-Western Mediterranean Coastal Brackish Areas, Tuscany, Italy. Agronomy 2023, 13, 615. [Google Scholar] [CrossRef]
- Farzana, T.; Guo, Q.; Rahman, M.S.; Rose, T.J.; Barkla, B.J. Salinity and Nitrogen Source Affect Productivity and Nutritional Value of Edible Halophytes. PLoS ONE 2023, 18, e0288547. [Google Scholar] [CrossRef]
- Hussain, W.; Abbas, Q.; Saleem, S.; Khan, S.W.; Shah, M.A. Assessment of Floristic Diversity and Traditional Knowledge from the Selected Mountainous Valleys of District Gilgit, Gilgit-Baltistan, Pakistan. Ethnobot. Res. Appl. 2024, 27, 1–22. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge Der Vegetationskunde; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 3-7091-4078-1. [Google Scholar]
- Van der Maarel, E. Transformation of Cover-Abundance Values in Phytosociology and Its Effects on Community Similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Costanza, R.; Cumberland, J.H.; Daly, H.; Goodland, R.; Norgaard, R.B. An Introduction to Ecological Economics; CRC Press: Boca Raton, FL, USA, 1997; ISBN 1-00-304084-5. [Google Scholar]
- Assessment, M.E. Ecosystems and Human Well-Being: Wetlands and Water; World Resources Institute: Washington, DC, USA, 2005; ISBN 1-56973-597-2. [Google Scholar]
- Sforzi, S.; Selvi, F. Flora Vascolare Della Palude “Diaccia Botrona”(Castiglione Della Pescaia, Grosseto). Atti Soc. Toscana Sci. Nat. Resid. Pisa Mem. Ser. B 1999, 106, 99–114. [Google Scholar]
- Cutini, M.; Agostinelli, E.; Acosta, T.; Molina, J. Coastal Salt-marsh Zonation in Tyrrhenian Central Italy and Its Relationship with Other Mediterranean Wetlands. Plant Biosyst. 2010, 144, 1–11. [Google Scholar] [CrossRef]
- Viciani, D.; Lombardi, L. La Vegetazione Del Padule Di Orti-Bottagone (Piombino, Toscana Meridionale) e La Sua Importanza Botanica Ai Fini Conservazionistici. Parlatorea 2001, 5, 101–118. [Google Scholar]
- Viciani, D.; Gabellini, A.; Biagini, P. La Vegetazione Del Padule Di Scarlino (Con Note Illustrative Della Carta Della Vegetazione, Scala 1: 12.000); S.EL.CA.: Firenze, Italy, 2001; pp. 1–48. [Google Scholar]
- Arrigoni, P.V. La Flora Vascolare Del Parco Della Maremma (Toscana, Italia Centrale). Webbia 2003, 58, 151–240. [Google Scholar] [CrossRef]
- Andreucci, F. La Vegetazione Alofila Della Laguna Di Orbetello (Toscana, Grosseto). Fitosociologia 2004, 41, 31–49. [Google Scholar]
- Sani, A.; Tomei, P.E. La Vegetazione Psammofila Del Litorale Di San Rossore (Toscana Settentrionale) e La Sua Importanza Conservazionistica. Parlatorea 2006, 8, 99–119. [Google Scholar]
- Bertacchi, A.; Lombardi, T.; Tomei, P. Le Aree Umide Salmastre Della Tenuta Di San Rossore (PI): Zonazione e Successione Delle Specie Vegetali in Relazione Alla Salinità Del Suolo; INTER NOS, Volume 1, Pisa, Italia: ETS; 2007; pp. 1–14. Available online: https://www.researchgate.net/publication/283301704_Le_aree_umide_salmastre_della_Tenuta_di_San_Rossore_PI_zonazione_e_successione_delle_specie_vegetali_in_relazione_alla_salinita_del_suolo (accessed on 1 February 2025).
- Londi, G.; Biagini, P.; Campedelli, T.; Mini, L.; Tellini Florenzano, G. Storia Ed Ecologia Del Padule Di Scarlino; Comune di Scarlino: Scarlino, Italy, 2007. [Google Scholar]
- Selvi, F. A Critical Checklist of the Vascular Flora of Tuscan Maremma (Grosseto Province, Italy). Flora Mediterr. 2010, 20, 47–139. [Google Scholar]
- Viciani, D.; Foggi, B.; Ferretti, G. The Mediterranean Salt Steppes (Order Limonietalia Br.-Bl. & O. Bolòs 1958) in Tuscany (Central Italy). Acta Bot. Gall. 2012, 159, 85–96. [Google Scholar]
- Angiolini, C.; Landi, M.; Pieroni, G.; Frignani, F.; Finoia, M.G.; Gaggi, C. Soil Chemical Features as Key Predictors of Plant Community Occurrence in a Mediterranean Coastal Ecosystem. Estuar. Coast. Shelf Sci. 2013, 119, 91–100. [Google Scholar] [CrossRef]
- Biondi, E.; Casavecchia, S.; Estrelles, E.; Soriano, P. Halocnemum M. Bieb. Vegetation in the Mediterranean Basin. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2013, 147, 536–547. [Google Scholar]
- Taramelli, A.; Valentini, E.; Piedelobo, L.; Righini, M.; Cappucci, S. Assessment of State Transition Dynamics of Coastal Wetlands in Northern Venice Lagoon, Italy. Sustainability 2021, 13, 4102. [Google Scholar] [CrossRef]
- Rozema, J.; Schat, H. Salt Tolerance of Halophytes, Research Questions Reviewed in the Perspective of Saline Agriculture. Environ. Exp. Bot. 2013, 92, 83–95. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of Halophytes: Multiple Origins of Salt Tolerance in Land Plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Soussi, M.; Khadri, M.; Pliego, L.; Ocaña, A.; Lluch, C. Effects of Salt Stress on the Symbiosis Cicer arietinum L.-Mesorhizobium cicero; European Association for Grain Legume Research: Villadolid, Spain, 1998. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Plant Salt Tolerance: Adaptations in Halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Grigore, M.-N.; Toma, C. Anatomical Adaptations of Halophytes. A Rev. Class. Lit. Recent 2017, 338, 9–36. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt Tolerance and Salinity Effects on Plants: A Review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Adamson, R.S. The Classification of Life-Forms of Plants. Bot. Rev. 1939, 5, 546–561. [Google Scholar] [CrossRef]
- Cain, S.A. Life-Forms and Phytoclimate. Bot. Rev. 1950, 16, 1–32. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Hot-Spots Analysis for Conservation of Plant Biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Zohary, D. The Origin of Cultivated Cereals and Pulses in the Near East. Chromosomes Today 1973, 4, 307–320. [Google Scholar]
- Weber, M.; Knoy, C.; Kindscher, K.; Brown, R.C.; Niemann, S.; Chapman, J. Identification of Medicinally Active Compounds in Prairie Plants by HPLC Coupled to Electron Impact-Mass Spectrometry. Am. Lab. 2007, 39, 9. [Google Scholar]
- Luczaj, L.; Pieroni, A.; Tardío, J.; Pardo-de-Santayana, M.; Sõukand, R.; Svanberg, I.; Kalle, R. Wild Food Plant Use in 21st Century Europe: The Disappearance of Old Traditions and the Search for New Cuisines Involving Wild Edibles. Acta Soc. Bot. Pol. 2012, 81, 359–370. [Google Scholar] [CrossRef]
- Pericin, C. Piante Selvatiche Commestibili Dell’Istria; Centro di Ricerche Storiche Rovigno: Rovinj, Croatia, 2020. [Google Scholar]
- Nedelcheva, A. An Ethnobotanical Study of Wild Edible Plants in Bulgaria. EurAsian J. BioSciences 2013, 7, 77–94. [Google Scholar] [CrossRef]
- Tan, A.; Adanacıoglu, N.; Karabak, S.; Aysar, N.; Tan, A.S.; Aykas, L. Sea Beets [Beta vulgaris Subsp. maritima (L.) Arcang.] Wild Edible Beets and Home Garden Beets of Turkey. ANADOLU Ege Tarımsal Araştırma Enstitüsü Derg. 2017, 27, 54–61. [Google Scholar]
- Pires, E.d.O., Jr.; Di Gioia, F.; Rouphael, Y.; Ferreira, I.C.; Caleja, C.; Barros, L.; Petropoulos, S.A. The Compositional Aspects of Edible Flowers as an Emerging Horticultural Product. Molecules 2021, 26, 6940. [Google Scholar] [CrossRef]
- Nasernakhaei, F.; Zahraei, M. Halocnemum strobilaceum (Pall.) M. Bieb.: A Review of Its Botany, Phytochemistry, Pharmacology and Ethnobotany. J. Med. Plants 2021, 20, 1–12. [Google Scholar] [CrossRef]
- Farhat, M.B.; Beji-Serairi, R.; Selmi, S.; Saidani-Tounsi, M.; Abdelly, C. Salicornia fruticosa L. and Portulaca oleracea L. Antioxidants as Affected by Domestic Cooking Processes. Int. J. Gastron. Food Sci. 2022, 27, 100462. [Google Scholar] [CrossRef]
- Loconsole, D.; Cristiano, G.; De Lucia, B. Glassworts: From Wild Salt Marsh Species to Sustainable Edible Crops. Agriculture 2019, 9, 14. [Google Scholar] [CrossRef]
- Antunes, M.D.; Gago, C.; Guerreiro, A.; Sousa, A.R.; Julião, M.; Miguel, M.G.; Faleiro, M.L.; Panagopoulos, T. Nutritional Characterization and Storage Ability of Salicornia ramosissima and Sarcocornia perennis for Fresh Vegetable Salads. Horticulturae 2021, 7, 6. [Google Scholar] [CrossRef]
- Lopes, M.; Silva, A.S.; Séndon, R.; Barbosa-Pereira, L.; Cavaleiro, C.; Ramos, F. Towards the Sustainable Exploitation of Salt-Tolerant Plants: Nutritional Characterisation, Phenolics Composition, and Potential Contaminants Analysis of Salicornia ramosissima and Sarcocornia perennis alpini. Molecules 2023, 28, 2726. [Google Scholar] [CrossRef] [PubMed]
- Ríos, S.; Obon, C.; Martínez-Francés, V.; Verde, A.; Ariza, D.; Laguna, E. Halophytes as Food: Gastroethnobotany of Halophytes. In Handbook of Halophytes: From Molecules to Ecosystems Towards Biosaline Agriculture; Springer Nature: Berlin, Germany, 2020; pp. 1–36. [Google Scholar]
- Pornpitakdamrong, A.; Sudjaroen, Y. Seablite (Suaeda maritima) Product for Cooking, Samut Songkram Province, Thailand. Food Nutr. Sci. 2014, 5, 850–856. [Google Scholar]
- Łuczaj, Ł.; Jug-Dujaković, M.; Dolina, K.; Vitasović-Kosić, I. Plants in Alcoholic Beverages on the Croatian Islands, with Special Reference to Rakija Travarica. J. Ethnobiol. Ethnomed. 2019, 15, 51. [Google Scholar] [CrossRef]
- Iltaf, M.; Niaz, S.I.; Majeed, M.K.; Saleem, M.; Shah, M.; Ali, M.; Shakeel Abbas, S.; Amin, A. DFT, GC-MS Analysis and Biological Evaluation of Limbarda crithmoides L. Dumort Essential Oil; an Important Edible Halophyte Grown in Pakistan. Nat. Prod. Res. 2024, 1–8. [Google Scholar] [CrossRef]
- Ismail, C.; Baraka, A.; Abdallah, R.; El-Dien, O.; Mostafa, D. Spergularia marina: A Potential Source of a Novel Calcium Channel Blocker with Antihypertensive and Diuretic Activities. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 506–517. [Google Scholar]
- Wollstonecroft, M.M.; Ellis, P.R.; Hillman, G.C.; Fuller, D.Q. Advances in Plant Food Processing in the Near Eastern Epipalaeolithic and Implications for Improved Edibility and Nutrient Bioaccessibility: An Experimental Assessment of Bolboschoenus maritimus (L.) Palla (Sea Club-Rush). Veg. Hist. Archaeobotany 2008, 17, 19–27. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J. Trifolium Species–the Latest Findings on Chemical Profile, Ethnomedicinal Use and Pharmacological Properties. J. Pharm. Pharmacol. 2016, 68, 845–861. [Google Scholar] [CrossRef]
- Gonçalves, S.; Romano, A. The Medicinal Potential of Plants from the Genus Plantago (Plantaginaceae). Ind. Crop. Prod. 2016, 83, 213–226. [Google Scholar] [CrossRef]
- González-Orenga, S.; Grigore, M.-N.; Boscaiu, M.; Vicente, O. Constitutive and Induced Salt Tolerance Mechanisms and Potential Uses of Limonium Mill. Species. Agronomy 2021, 11, 413. [Google Scholar] [CrossRef]
- Velez-Gavilan, J. Phragmites australis (Common Reed). CABI Compend. 2024. [Google Scholar] [CrossRef]
- Mattera, M.; Pilla, N.; Aguzzi, A.; Gabrielli, P.; Di Lena, G.; Durazzo, A.; Lucarini, M. Portulaca oleracea L.: Literature Quantitative Research Analysis. Nat. Prod. Res. 2024, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ventura, Y.; Sagi, M. Halophyte Crop Cultivation: The Case for Salicornia and Sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar] [CrossRef]
- Ali, B.; Musaddiq, S.; Iqbal, S.; Rehman, T.; Shafiq, N.; Hussain, A. The Therapeutic Properties, Ethno Pharmacology and Phytochemistry of Atriplex Species: A Review. Pak. J. Biochem. Biotechnol. 2021, 2, 49–64. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombardi, T.; Bedini, S.; Bertacchi, A.; Ventura, I. Exploring the Potential of Mediterranean Edible Halophytes as Novel Crops: Ecological and Nutritional Insights from Tuscany’s Salt Marshes. Agronomy 2025, 15, 634. https://doi.org/10.3390/agronomy15030634
Lombardi T, Bedini S, Bertacchi A, Ventura I. Exploring the Potential of Mediterranean Edible Halophytes as Novel Crops: Ecological and Nutritional Insights from Tuscany’s Salt Marshes. Agronomy. 2025; 15(3):634. https://doi.org/10.3390/agronomy15030634
Chicago/Turabian StyleLombardi, Tiziana, Stefano Bedini, Andrea Bertacchi, and Irene Ventura. 2025. "Exploring the Potential of Mediterranean Edible Halophytes as Novel Crops: Ecological and Nutritional Insights from Tuscany’s Salt Marshes" Agronomy 15, no. 3: 634. https://doi.org/10.3390/agronomy15030634
APA StyleLombardi, T., Bedini, S., Bertacchi, A., & Ventura, I. (2025). Exploring the Potential of Mediterranean Edible Halophytes as Novel Crops: Ecological and Nutritional Insights from Tuscany’s Salt Marshes. Agronomy, 15(3), 634. https://doi.org/10.3390/agronomy15030634