
Enhancing Nutritional and Functional Properties of Broccoli Leaves Through Selenium Biofortification: Potential for Sustainable Agriculture and Bioactive Compound Valorization

 , ,
, ,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Foliar Application of Selenium
2.3. Morphological and Physiological Parameters
2.4. Biochemical Parameters
2.4.1. Soluble Protein Content
2.4.2. GSH-Px Activity
2.4.3. Total Phenolic Compound
2.4.4. Evaluation of Total Antioxidant Capacity
2.4.5. Extraction, Purification, and Quantification of Endogenous Leaves Hormones
2.4.6. Total Se and Mineral Elements Determination
2.5. Preparation of Broccoli Leaf Aqueous Extracts
2.6. In Vitro Antimicrobial Activity of Aqueous Extracts on the Growth Inhibition of Fusarium solani
2.7. Statistical Analysis
3. Results and Discussion
3.1. Evaluation of Changes in Morphological and Physiological Parameters During the Growth Stages of Broccoli Plants
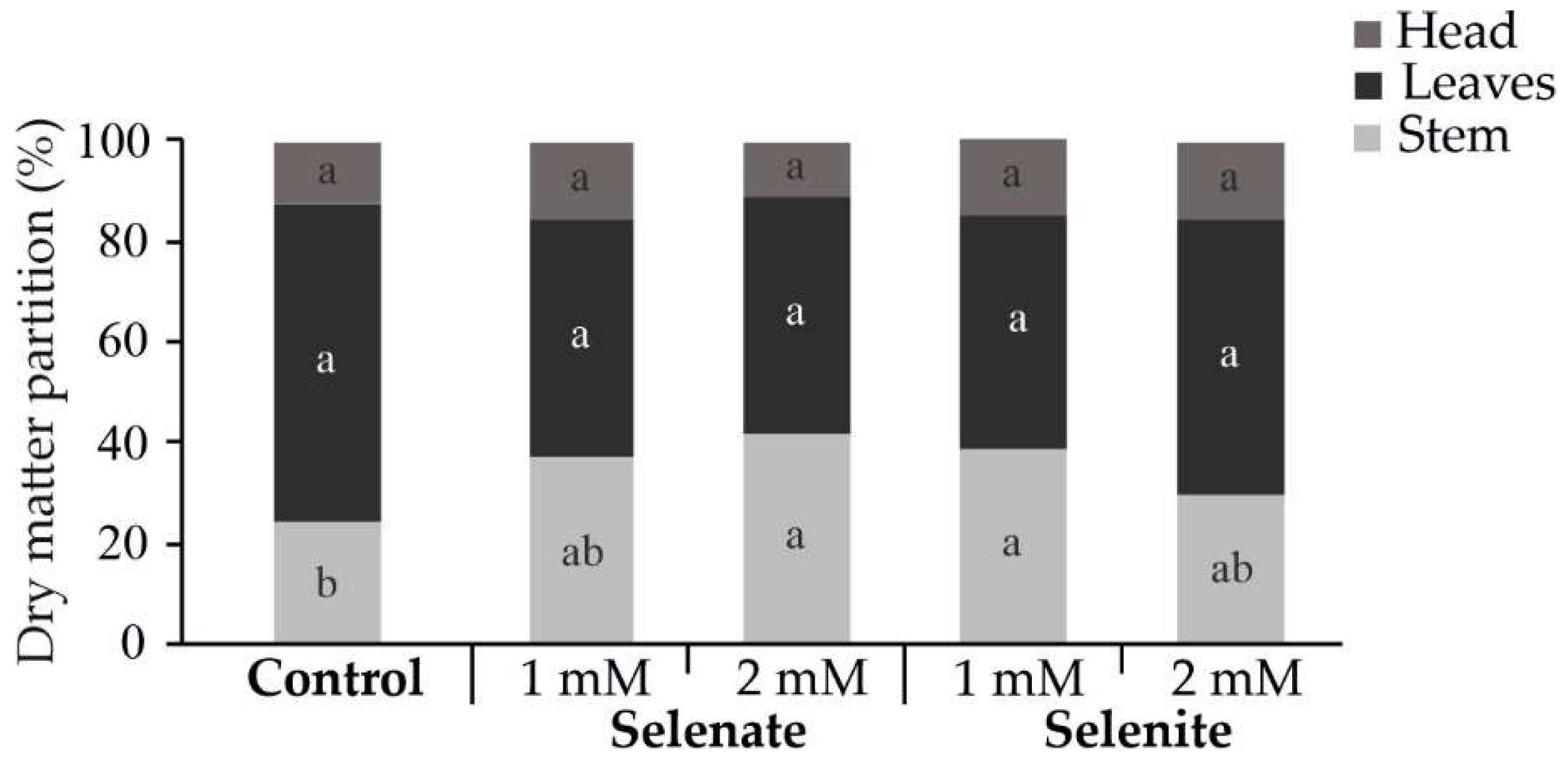
3.2. Effect of Foliar Selenium Application on Biomass Distribution, Fresh Weight, Firmness, and Diameter of Broccoli Heads
3.3. Principal Component Analysis of Selenium Treatments on Growth, Morphological, and Yield Parameters in ‘Belstar’ Broccoli
3.4. Selenium Improves Nutraceutical Parameters in Broccoli By-Products
3.4.1. Biochemical Parameters
3.4.2. Foliar Application of Selenium Increases the Mineral Content of Broccoli By-Products
3.5. Differential Effects of Selenate and Selenite on Phytohormone Regulation in Broccoli Leaves: Balancing Growth and Defense Responses
3.6. Antifungal Potential of Selenium-Biofortified Broccoli Leaf Extracts Against Fusarium solani
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef] [PubMed]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Tinggi, U. Selenium: Its role as antioxidant in human health. Environ. Health Prev. Med. 2008, 13, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Shanaida, M.; Lysiuk, R.; Antonyak, H.; Klishch, I.; Shanaida, V.; Peana, M. Selenium: An Antioxidant with a Critical Role in Anti-Aging. Molecules 2022, 27, 6613. [Google Scholar] [CrossRef]
- Galić, L.; Vinković, T.; Ravnjak, B.; Lončarić, Z. Agronomic Biofortification of Significant Cereal Crops with Selenium—A Review. Agronomy 2021, 11, 1015. [Google Scholar] [CrossRef]
- Wan, H.-S.; Zhang, W.-C.; Wu, W.; Liu, H.-B. The Controlling Factors of Soil Selenium Content in a Selenium-Deficient Area in Southwest China. Agronomy 2023, 13, 1031. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.-Q.; Broadley, M. Selenium Biofortification. In Selenium in Plants: Molecular, Physiological, Ecological and Evolutionary Aspects; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.-Q., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 231–255. [Google Scholar]
- Poblaciones, M.J.; Broadley, M.R. Foliar selenium biofortification of broccolini: Effects on plant growth and mineral accumulation. J. Hortic. Sci. Biotechnol. 2022, 97, 730–738. [Google Scholar] [CrossRef]
- Subirana, M.A.; Boada, R.; Xiao, T.; Llugany, M.; Valiente, M. Direct and indirect selenium speciation in biofortified wheat: A tale of two techniques. Physiol. Plant. 2023, 175, e13843. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Arroyo, I.; Pickering, I.J.; Yang, S.I.; Freeman, J.L. Selenium biofortification of broccoli and carrots grown in soil amended with Se-enriched hyperaccumulator Stanleya pinnata. Food Chem. 2015, 166, 603–608. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Arroyo, I.S.; Dangi, S.R.; Zambrano, M.C. Continued Selenium Biofortification of Carrots and Broccoli Grown in Soils Once Amended with Se-Enriched S. pinnata. Front. Plant Sci. 2016, 7, 1251. [Google Scholar] [CrossRef]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current Knowledge on Selenium Biofortification to Improve the Nutraceutical Profile of Food: A Comprehensive Review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, F.F.; Stoffel, M.M.; Céccoli, G.; Trod, B.S.; Daurelio, L.D.; Bouzo, C.A.; Guevara, M.G. Improving the foliar biofortification of broccoli with selenium without commercial quality losses. Crop Sci. 2021, 61, 4218–4228. [Google Scholar] [CrossRef]
- Golubkina, N.; Antoshkina, M.; Bondareva, L.; Sekara, A.; Campagna, E.; Caruso, G. Effect of Foliar Application of Sodium Selenate on Mineral Relationships in Brassicaceae Crops. Horticulturae 2023, 9, 535. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Ghasemi, K.; Pirdashti, H.; Asgharzadeh, R. Effect of selenium enrichment on the growth, photosynthesis and mineral nutrition of broccoli. Not. Sci. Biol. 2016, 8, 109–203. [Google Scholar] [CrossRef]
- Hsu, F.C.; Wirtz, M.; Heppel, S.C.; Bogs, J.; Krämer, U.; Khan, M.S.; Bub, A.; Hell, R.; Rausch, T. Generation of Se-fortified broccoli as functional food: Impact of Se fertilization on S metabolism. Plant Cell Environ. 2011, 34, 192–207. [Google Scholar] [CrossRef]
- Sindelarova, K.; Szakova, J.; Tremlova, J.; Mestek, O.; Praus, L.; Kana, A.; Najmanova, J.; Tlustos, P. The response of broccoli (Brassica oleracea convar. italica) varieties on foliar application of selenium: Uptake, translocation, and speciation. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 2027–2038. [Google Scholar] [CrossRef]
- Lozano, L.; Tálamo, A.; Artinián, A.L.; Fernández, J.; Arroyo, C. Evaluación de dos híbridos de brócoli (Brassica oleracea var. italica). Horticultura 2017, 36, 37–48. [Google Scholar]
- Dominguez-Perles, R.; Martinez-Ballesta, M.C.; Carvajal, M.; Garcia-Viguera, C.; Moreno, D.A. Broccoli derived by-products a promising source of bioactive ingredients. J. Food Sci. 2010, 75, C383–C392. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, L.; Ser, S.L.; Cumming, J.R.; Ku, K.M. Comparative Phytonutrient Analysis of Broccoli By-Products: The Potentials for Broccoli By-Product Utilization. Molecules 2018, 23, 900. [Google Scholar] [CrossRef]
- Artés-Hernández, F.; Martínez-Zamora, L.; Cano-Lamadrid, M.; Hashemi, S.; Castillejo, N. Genus Brassica By-Products Revalorization with Green Technologies to Fortify Innovative Foods: A Scoping Review. Foods 2023, 12, 561. [Google Scholar] [CrossRef]
- Hwang, J.H.; Lim, S.B. Antioxidant and anticancer activities of broccoli by-products from different cultivars and maturity stages at harvest. Prev. Nutr. Food Sci. 2015, 20, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Francioso, O. Current and future perspectives for biomass waste management and utilization. Sci. Rep. 2024, 14, 9635. [Google Scholar] [CrossRef] [PubMed]
- Gudiño, I.; Martín, A.; Casquete, R.; Prieto, M.H.; Ayuso, M.C.; Córdoba, M.G. Evaluation of broccoli (Brassica oleracea var. italica) crop by-products as sources of bioactive compounds. Sci. Hortic. 2022, 304, 111284. [Google Scholar] [CrossRef]
- Tripathi, N.; Hills, C.D.; Singh, R.S.; Atkinson, C.J. Biomass waste utilisation in low-carbon products: Harnessing a major potential resource. npj Clim. Atmos. Sci. 2019, 2, 35. [Google Scholar] [CrossRef]
- Sadh, P.K.; Chawla, P.; Kumar, S.; Das, A.; Kumar, R.; Bains, A.; Sridhar, K.; Duhan, J.S.; Sharma, M. Recovery of agricultural waste biomass: A path for circular bioeconomy. Sci. Total Environ. 2023, 870, 161904. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, Y. Recent progress in the conversion of biomass wastes into functional materials for value-added applications. Sci. Technol. Adv. Mater. 2020, 21, 787–804. [Google Scholar] [CrossRef]
- Shinali, T.S.; Zhang, Y.; Altaf, M.; Nsabiyeze, A.; Han, Z.; Shi, S.; Shang, N. The Valorization of Wastes and Byproducts from Cruciferous Vegetables: A Review on the Potential Utilization of Cabbage, Cauliflower, and Broccoli Byproducts. Foods 2024, 13, 1163. [Google Scholar] [CrossRef]
- Damas-Job, M.C.; Soriano-Melgar, L.A.A.; Rodríguez-Herrera, R.; Peralta-Rodríguez, R.D.; Rivera-Cabrera, F.; Martínez-Vazquez, D.G. Effect of broccoli fresh residues-based extracts on the postharvest quality of cherry tomato (Solanum lycopersicum L.) fruits. Sci. Hortic. 2023, 317, 112076. [Google Scholar] [CrossRef]
- Eugui, D.; Velasco, P.; Abril-Urías, P.; Escobar, C.; Gómez-Torres, Ó.; Caballero, S.; Poveda, J. Glucosinolate-extracts from residues of conventional and organic cultivated broccoli leaves (Brassica oleracea var. italica) as potential industrially-scalable efficient biopesticides against fungi, oomycetes and plant parasitic nematodes. Ind. Crops Prod. 2023, 200, 116841. [Google Scholar] [CrossRef]
- Trod, B.S.; Buttarelli, M.S.; Stoffel, M.M.; Céccoli, G.; Olivella, L.; Barengo, P.B.; Llugany, M.; Guevara, M.G.; Muñoz, F.F.; Daurelio, L.D. Postharvest commercial quality improvement of broccoli (Brassica oleracea L.) after foliar biofortification with selenium. Crop Sci. 2023, 63, 784–800. [Google Scholar] [CrossRef]
- Meier, U. (Ed.) Growth stages of mono-and dicotyledonous plants. In BBCH Monograph, 4th ed.; Federal Biological Research Centre for Agriculture and Forestry: Quedlinburg, Germany; Julius Kuhn-Institut: Quedlinburg, Germany, 2018; p. 204. [Google Scholar]
- Céccoli, G.; Seen, M.; Bustos, D.; Ortega, L.I.; Cordoba, A.; Vegetti, A.; Taleisnik, E. Genetic variability for responses to short- and long-term salt stress in vegetative sunflower plants. J. Plant Nutr. Soil Sci. 2012, 175, 882–890. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. Available online: https://pubmed.ncbi.nlm.nih.gov/6066618/ (accessed on 1 January 2025). [PubMed]
- Lemoine, M.; Chaves, A.R.; Martínez, G. Influence of combined hot air and UV-C treatment on the antioxidant system of minimally processed broccoli (Brassica oleracea L. var. italica). LWT—Food Sci. Technol. 2010, 43, 1313–1319. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Bartoszek, A.; Wolska, L.; Drzewiecki, J.; Gorinstein, S.; Namieśnik, J. Partial characterization of white cabbages (Brassica oleracea var. capitata f. alba) from different regions by glucosinolates, bioactive compounds, total antioxidant activities and proteins. LWT—Food Sci. Technol. 2008, 41, 1–9. [Google Scholar] [CrossRef]
- Llugany, M.; Martin, S.R.; Barceló, J.; Poschenrieder, C. Endogenous jasmonic and salicylic acids levels in the Cd-hyperaccumulator Noccaea (Thlaspi) praecox exposed to fungal infection and/or mechanical stress. Plant Cell Rep. 2013, 32, 1243–1249. [Google Scholar] [CrossRef]
- Segarra, G.; Jáuregui, O.; Casanova, E.; Trillas, I. Simultaneous quantitative LC–ESI-MS/MS analyses of salicylic acid and jasmonic acid in crude extracts of Cucumis sativus under biotic stress. Phytochemistry 2006, 67, 395–401. [Google Scholar] [CrossRef]
- Guevara, M.G.; Oliva, C.R.; Huarte, M.; Daleo, G.R. An aspartic protease with antimicrobial activity is induced after infection and wounding in intercellular fluids of potato tubers. Eur. J. Plant Pathol. 2002, 108, 131–137. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M.; Robledo, C. InfoStat. Statistical Software; Grupo Infostat FCA UNC: Córdoba, Argentina, 2017. [Google Scholar]
- Thuc, L.V.; Sakagami, J.I.; Hung, L.T.; Huu, T.N.; Khuong, N.Q.; Vi, L.L.V. Foliar selenium application for improving drought tolerance of sesame (Sesamum indicum L.). Open Agric. 2021, 6, 93–101. [Google Scholar] [CrossRef]
- Zhou, W.; Duan, Y.; Zhang, Y.; Wang, H.; Huang, D.; Zhang, M. Effects of foliar selenium application on growth and rhizospheric soil micro-ecological environment of Atractylodes macrocephala Koidz. S. Afr. J. Bot. 2021, 137, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Skrypnik, L.; Feduraev, P.; Golubkina, N.; Maslennikov, P.; Antipina, M.; Katserov, D.; Murariu, O.C.; Tallarita, A.V.; Caruso, G. Foliar spraying of selenium in inorganic and organic forms stimulates plant growth and secondary metabolism of sage (Salvia officinalis L.) through alterations in photosynthesis and primary metabolism. Sci. Hortic. 2024, 338, 113633. [Google Scholar] [CrossRef]
- Lin, Y.; Cao, S.; Wang, X.; Liu, Y.; Sun, Z.; Zhang, Y.; Li, M.; Wang, Y.; He, W.; Zhang, Y.; et al. Foliar application of sodium selenite affects the growth, antioxidant system, and fruit quality of strawberry. Front. Plant Sci. 2024, 15, 1449157. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Su, Y.; Yang, R.; Xie, Z.; Gong, H. Effect of Foliar Application of Silicon and Selenium on the Growth, Yield and Fruit Quality of Tomato in the Field. Horticulturae 2023, 9, 1126. [Google Scholar] [CrossRef]
- Cheng, H.; Shi, X.; Li, L. The Effect of Exogenous Selenium Supplementation on the Nutritional Value and Shelf Life of Lettuce. Agronomy 2024, 14, 1380. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Lu, Y.; Liao, Y.; Nie, J.; Yuan, X.; Chen, F. Use of a leaf chlorophyll content index to improve the prediction of above-ground biomass and productivity. PeerJ 2019, 2019, e6240. [Google Scholar] [CrossRef]
- Francescangeli, N.; Sangiacomo, M.; Marti, H. Effects of plant density in broccoli on yield and radiation use efficiency. Sci. Hortic. 2006, 110, 135–143. [Google Scholar] [CrossRef]
- Huang, J.; Sedano, F.; Huang, Y.; Ma, H.; Li, X.; Liang, S.; Tian, L.; Zhang, X.; Fan, J.; Wu, W. Assimilating a synthetic Kalman filter leaf area index series into the WOFOST model to improve regional winter wheat yield estimation. Agric. For. Meteorol. 2016, 216, 188–202. [Google Scholar] [CrossRef]
- Lindemann-Zutz, K.; Fricke, A.; Stützel, H. Prediction of time to harvest and its variability of broccoli (Brassica oleracea var. italica) part II. Growth model description, parameterisation and field evaluation. Sci. Hortic. 2016, 200, 151–160. [Google Scholar] [CrossRef]
- Casierra-Posada, F.; Peña-Olmos, J.E. Prolonged waterlogging reduces growth and yield in Broccoli Plants (Brassica oleracea var. italica). Gesunde Pflanz. 2022, 74, 249–257. [Google Scholar] [CrossRef]
- El-Mogy, M.M.; Mahmoud, A.W.M.; El-Sawy, M.B.I.; Parmar, A. Pre-Harvest Foliar Application of Mineral Nutrients to Retard Chlorophyll Degradation and Preserve Bio-Active Compounds in Broccoli. Agronomy 2019, 9, 711. [Google Scholar] [CrossRef]
- Palencia, P.; Martinez, F.; Burducea, M.; Oliveira, J.A.; Giralde, I. Efectos del enriquecimiento con Selenio en spad, calidad de la fruta y parámetros de crecimiento de plantas de fresa en un sistema de cultivo sin suelo. Rev. Bras. Frutic. 2016, 38, 202–212. [Google Scholar] [CrossRef]
- Conversa, G.; Lazzizera, C.; Bonasia, A.; Elia, A. Harvest Season and Genotype Affect Head Quality and Shelf-Life of Ready-to-Use Broccoli. Agronomy 2020, 10, 527. [Google Scholar] [CrossRef]
- Dixon, G.; Wells, R. Vegetable Brassicas and Related Crucifers, 2nd ed.; CABI Digital Library: Wallingford, UK, 2024; p. 544. [Google Scholar] [CrossRef]
- Bouranis, D.L.; Stylianidis, G.P.; Manta, V.; Karousis, E.N.; Tzanaki, A.; Dimitriadi, D.; Bouzas, E.A.; Siyiannis, V.F.; Constantinou-Kokotou, V.; Chorianopoulou, S.N.; et al. Floret Biofortification of Broccoli Using Amino Acids Coupled with Selenium under Different Surfactants: A Case Study of Cultivating Functional Foods. Plants 2023, 12, 1272. [Google Scholar] [CrossRef]
- Martirosyan, V.V.; Kostyuchenko, M.N.; Kryachko, T.I.; Malkina, V.D.; Zhirkova, E.V.; Golubkina, N.A. The Beneficial Effect of Selenium-Enriched Broccoli on the Quality Characteristics of Bread. Processes 2023, 11, 3037. [Google Scholar] [CrossRef]
- Liu, L.; Wang, C.; Yin, K.; Ni, M.; Ding, Y.; Li, C.; Zheng, S.-J. The Dual Effect of Selenium Application in Reducing Fusarium Wilt Disease Incidence in Banana and Producing Se-Enriched Fruits. Plants 2024, 13, 3435. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Garousi, F.; Domokos-Szabolcsy, E.; Jánószky, M.; Balláné Kovács, A.; Veres, S.; Soós, A.; Kovács, B. Selenoamino acid-enriched Green Pea as a value-added plant protein source for humans and livestock. Plant Foods Hum. Nutr. 2017, 72, 168–175. [Google Scholar] [CrossRef]
- Newman, R.; Waterland, N.; Moon, Y.; Tou, J.C. Selenium biofortification of agricultural crops and effects on plant nutrients and bioactive compounds important for human health and disease prevention—A Review. Plant Foods Hum. Nutr. 2019, 74, 449–460. [Google Scholar] [CrossRef]
- Li, H.; Xia, Y.; Liu, H.Y.; Guo, H.; He, X.Q.; Liu, Y.; Wu, D.T.; Mai, Y.H.; Li, H.B.; Zou, L.; et al. Nutritional values, beneficial effects, and food applications of broccoli (Brassica oleracea var. italica Plenck). Trends Food Sci. Technol. 2022, 119, 288–308. [Google Scholar] [CrossRef]
- Pazurkiewicz-Kocot, K.; Kita, A.; Pietruszka, M. Effect of Selenium on Magnesium, Iron, Manganese, Copper, and Zinc Accumulation in Corn Treated by Indole-3-acetic Acid. Commun. Soil Sci. Plant Anal. 2008, 39, 2303–2318. [Google Scholar] [CrossRef]
- Saffaryazdi, A.; Lahouti, M.; Ganjeali, A.; Bayat, H. Impact of selenium supplementation on growth and selenium accumulation on spinach (Spinacia oleracea L.) plants. Not. Sci. Biol. 2012, 4, 95–100. [Google Scholar] [CrossRef]
- He, P.P.; Lv, X.Z.; Wang, G.Y. Effects of Se and Zn supplementation on the antagonism against Pb and Cd in vegetables. Environ. Int. 2004, 30, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.J.; Blasco, B.; Leyva, R.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M. Nutritional balance changes in lettuce plant grown under different doses and forms of selenium. J. Plant Nutr. 2014, 36, 1344–1354. [Google Scholar] [CrossRef]
- Narváez-Ortiz, W.A.; Martínez-Hernández, M.; Fuentes-Lara, L.O.; Benavides-Mendoza, A.; Valenzuela-García, J.R.; González-Fuentes, J.A. Effect of selenium application on mineral marco- and micronutrients and antioxidant status in strawberries. J. Appl. Bot. Food Qual. 2018, 91, 321–331. [Google Scholar] [CrossRef]
- Ahmed, N.; Zhang, B.; Bozdar, B.; Chachar, S.; Rai, M.; Li, J.; Li, Y.; Hayat, F.; Chachar, Z.; Tu, P. The power of magnesium: Unlocking the potential for increased yield, quality, and stress tolerance of horticultural crops. Front. Plant Sci. 2023, 14, 1285512. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S.; Wu, F. Effects of Se on the uptake of essential elements in Pteris vittata L. Plant Soil 2009, 325, 123–132. [Google Scholar] [CrossRef]
- Boldrin, P.F.; Faquin, V.; Ramos, S.J.; Furtini Boldrin, K.V.; Ávila, F.W.; Guimarães Guilherme, L.R. Soil and foliar application of selenium in rice biofortification. J. Food Compos. Anal. 2013, 31, 238–244. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Devi, D.D.; Shanker, A.K.; Sheeba, J.A.; Bangarusamy, U. Selenium—An antioxidative protectant in soybean during senescence. Plant Soil 2005, 272, 77–86. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef]
- Freeman, J.L.; Tamaoki, M.; Stushnoff, C.; Quinn, C.F.; Cappa, J.J.; Devonshire, J.; Fakra, S.C.; Marcus, M.A.; McGrath, S.P.; Van Hoewyk, D.; et al. Molecular mechanisms of selenium tolerance and hyperaccumulation in Stanleya pinnata. Plant Physiol. 2010, 153, 1630–1652. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, M.; Schiavon, M.; dall’Acqua, S.; Pilon-Smits, E.A.H. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.A.; Elyamine, A.M.; Zhao, Y.Y.; Moussa, M.G.; Rana, M.S.; Afzal, J.; Imran, M.; Zhao, X.H.; Hu, C.X. Can Selenium and Molybdenum Restrain Cadmium Toxicity to Pollen Grains in Brassica napus? Int. J. Mol. Sci. 2018, 19, 2163. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stress. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Schiavon, M.; Pilon, M.; Malagoli, M.; Pilon-Smits, E.A.H. Exploring the importance of sulfate transporters and ATP sulphurylases for selenium hyperaccumulation-a comparison of Stanleya pinnata and Brassica juncea (Brassicaceae). Front. Plant. Sci. 2015, 6, 2. [Google Scholar] [CrossRef]
- Feng, R.W.; Wei, C.Y. Antioxidative mechanisms on selenium accumulation in Pteris vittata L., a potential selenium phytoremediation plant. Plant Soil Environ. 2012, 58, 105–110. [Google Scholar] [CrossRef]
- Chao, W.; Rao, S.; Chen, Q.; Zhang, W.; Liao, Y.; Ye, J.; Cheng, S.; Yang, X.; Xu, F. Advances in Research on the Involvement of Selenium in Regulating Plant Ecosystems. Plants 2022, 11, 2712. [Google Scholar] [CrossRef]
- Fan, S.; Wu, H.; Gong, H.; Guo, J. The salicylic acid mediates selenium-induced tolerance to drought stress in tomato plants. Sci. Hortic. 2022, 300, 111092. [Google Scholar] [CrossRef]
- Tamaoki, M.; Freeman, J.L.; Marquès, L.; Pilon-Smits, E.A.H. New insights into the roles of ethylene and jasmonic acid in the acquisition of selenium resistance in plants. Plant Signal. Behav. 2008, 3, 865–867. [Google Scholar] [CrossRef]
- Mao, X.; Hua, C.; Yang, L.; Zhang, Y.; Sun, Z.; Li, L.; Li, T. The effects of selenium on wheat fusarium head blight and DON accumulation were selenium compound-dependent. Toxins 2020, 12, 573. [Google Scholar] [CrossRef] [PubMed]
- Troni, E.; Beccari, G.; D’Amato, R.; Tini, F.; Baldo, D.; Senatore, M.T.; Beone, G.M.; Fontanella, M.C.; Prodi, A.; Businelli, D.; et al. In Vitro Evaluation of the Inhibitory Activity of Different Selenium Chemical Forms on the Growth of a Fusarium proliferatum Strain Isolated from Rice Seedlings. Plants 2021, 10, 1725. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xian, L.; Yuan, L.; Lin, Z.; Chen, X.; Wang, J.; Li, T. The use of selenium for controlling plant fungal diseases and insect pests. Front. Plant Sci. 2023, 14, 1102594. [Google Scholar] [CrossRef]
- Hanson, B.; Garifullina, G.F.; Lindblom, S.D.; Wangeline, A.; Ackley, A.; Kramer, K.; Norton, A.P.; Lawrence, C.B.; Pilon-Smits, E.A.H. Selenium accumulation protects Brassica juncea from invertebrate herbivory and fungal infection. New Phytol. 2003, 159, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Jia, W.; Hu, C.; Shi, G.; Yang, D.; Cai, M.; Zhan, T.; Tang, Y.; Zhou, Y.; Sun, X.; et al. Enhancement and improvement of selenium in soil to the resistance of rape stem against Sclerotinia sclerotiorum and the inhibition of dissolved organic matter derived from rape straw on mycelium. Environ. Pollut. 2020, 265, 114827. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, H.; Hu, T.; Wang, Z.; Wu, W.; Liang, Y.; Guo, Y. Sunflower resistance against Sclerotinia sclerotiorum is potentiated by selenium through regulation of redox homeostasis and hormones signaling pathways. Environ. Sci. Pollut. Res. 2022, 29, 38097–38109. [Google Scholar] [CrossRef]
- Companioni, B.; Medrano, J.; Torres, J.A.; Flores, A.; Rodríguez, E.; Benavides, A. Protective action of sodium selenite against fusarium wilt in tomato: Total protein contents, levels of phenolic compounds and changes in antioxidant potential. ISHS Acta Hortic. 2012, 947, 321–327. [Google Scholar] [CrossRef]
- Somalraju, A.; Mccallum, J.L.; Main, D.; Peters, R.D.; Fofana, B. Foliar selenium application reduces late blight severity and incidence in potato and acts as a pathogen growth inhibitor and elicitor of induced plant defence. Can. J. Plant Pathol. 2021, 44, 39–55. [Google Scholar] [CrossRef]
- Wu, Z.-L.; Yin, X.-B.; Lin, Z.-Q.; Bañuelos, G.S.; Yuan, L.-X.; Liu, Y.; Li, M. Ihibitory effect of selenium against Penicillium expansum and its possible mechanisms of action. Curr. Microbiol. 2014, 69, 192–201. [Google Scholar] [CrossRef]
- Zohri, A.; Saber, S.M.; Mostafa, M.E. Effect of selenite and tellurite on the morphological growth and toxin production of Aspergillus parasiticus var. globosus IMI 120920. Mycopathologia 1997, 139, 51–57. [Google Scholar] [CrossRef]
- Nowruzi, B.; Jalil, B.S.; Metcalf, J.S. Antifungal screening of selenium nanoparticles biosynthesized by microcystin-producing Desmonostoc alborizicum. BMC Biotechnol. 2023, 23, 41. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Mukherjee, A.; Das, A.; Roy, A.; Majumdar, A.; Dhar, A.; Pattanaik, B.K.; Chowardhara, B.; Ghosh, D.; Upadhyay, M.K.; et al. Selenium—An environmentally friendly micronutrient in agroecosystem in the modern era: An overview of 50-year findings. Ecotoxicol. Environ. Saf. 2024, 270, 115832. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| PH (cm) | LA (cm2) | |||||
|---|---|---|---|---|---|---|
| Treatment | Days after Transplanting (DAT) | |||||
| 78 | 85 | 90 | 78 | 85 | 90 | |
| Control | 15.0 ± 1.5 fg | 19.3 ± 2.8 cdef | 24.2 ± 3.1 a | 99.5 ± 3.9 abcd | 103.0 ± 15.8 abc | 107.5 ± 11.7 abc |
| Selenate 1 mM | 13.7 ± 0.9 g | 19.5 ± 0.9 bcdef | 24.0 ± 1.2 ab | 95.9 ± 1.3 bcd | 107.17 ± 5.6 abc | 107.0 ± 6.2 abcd |
| Selenate 2 mM | 15.5 ± 0.3 efg | 18.5 ± 1.5 def | 23.3 ± 2.5 abc | 102.2 ± 5.1 abcd | 121.5 ± 16.8 ab | 110.6 ± 22.8 a |
| Selenite 1 mM | 12.2 ± 0.2 g | 16.7 ± 0.3 efg | 22.3 ± 0.3 abcd | 76.9 ± 5.5 d | 102.1 ± 10.7 abcd | 124.6 ± 16.1 abc |
| Selenite 2 mM | 15.3 ± 0.9 efg | 19.7 ± 1.5 abcde | 22.2 ± 1.6 abcd | 88.9 ± 7.2 cd | 103.2 ± 14.6 abcd | 91.7 ± 11.8 cd |
| Treatment × DAT (p-value) | 0.9542 | 0.9093 | ||||
| Treatment | PRER (cm cm−1 d−1) |
|---|---|
| Control | 0.91 ± 0.09 ab |
| Selenate 1 mM | 0.86 ± 0.04 ab |
| Selenate 2 mM | 0.79 ± 0.15 ab |
| Selenite 1 mM | 0.83 ± 0.01 ab |
| Selenite 2 mM | 0.76 ± 0.02 a |
| Control | Selenate 2 mM | Selenite 2 mM | |
|---|---|---|---|
| Antioxidant capacity (mg ascorbic acid equiv. g−1 DW) | 2.76 ± 0.11 a | 4.38 ± 0.37 b | 3.44 ± 0.14 a |
| GSH-Px activity (U g−1 DW) | 0.70 ± 0.03 a | 1.05 ± 0.13 a | 1.20 ± 0.28 a |
| Phenolic compounds (mg gallic acid equiv. g−1 DW) | 4.27 ± 0.34 a | 6.93 ± 0.84 b | 6.53 ± 0.54 b |
| Soluble proteins content (mg g−1 DW) | 0.15 ± 0.01 a | 0.43 ± 0.02 c | 0.24 ± 0.02 b |
| µg g−1 | Control | Selenate 2 mM | Selenite 2 mM. |
|---|---|---|---|
| Se | 0.27 ± 0.00 a | 27.22 ± 0.26 b | 40.44 ± 1.37 c |
| K | 17,258 ± 1872.09 a | 14,904 ± 393.08 a | 18,808 ± 1932.60 a |
| P | 2910 ± 91.53 a | 2687 ± 146.30 a | 2917 ± 195.65 a |
| Mg | 2811 ± 83.89 ab | 2518 ± 60.11 a | 3200 ± 143.72 b |
| Ca | 12,809 ± 789.73 a | 11,987 ± 519.65 a | 19,826 ± 1183.18 b |
| S | 8257 ± 63.98 a | 9871 ± 446.20 b | 12,971 ± 43.52 c |
| Fe | 27.69 ± 2.27 a | 42.66 ± 3.14 b | 46.48 ± 2.35 b |
| Mn | 36.61 ± 2.67 a | 29.14 ± 3.20 a | 52.34 ± 1.62 b |
| B | 36.53 ± 1.79 a | 37.47 ± 1.88 a | 37.39 ± 3.05 a |
| Zn | 25.66 ± 0.13 a | 26.70 ± 0.95 a | 25.16 ± 1.87 a |
| Mo | 11.53 ± 0.01 a | 9.69 ± 0.69 a | 15.30 ± 0.22 b |
| Cu | 2.32 ± 0.36 a | 2.41 ± 0.25 a | 3.00 ± 0.16 a |
| Ni | 0.30 ± 0.05 a | 0.24 ± 0.01 a | 0.23 ± 0.01 a |
| ng g−1 | Control | Selenate 2 mM | Selenite 2 mM |
|---|---|---|---|
| [MJA] | 15.05 ± 0.92 a | 19.86 ± 3.03 ab | 28.99 ± 2.43 b |
| [JA] | 61.52 ± 5.74 b | 40.54 ± 2.89 a | 54.04 ± 6.32 ab |
| [SA] | 3216.33 ± 204.84 a | 3306.33 ± 112.72 a | 9793.00 ± 633.33 b |
| [IAA] | 12.27 ± 2.54 a | 27.38 ± 3.47 b | 10.87 ± 0.91 a |
| [ABA] | 51.08 ± 3.54 b | 97.44 ± 6.38 c | 28.84 ± 6.51 a |
| [ACC] | 5.85 ± 0.29 b | 6.75 ± 0.68 b | 3.55 ± 0.60 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buttarelli, M.S.; Céccoli, G.; Trod, B.S.; Stoffel, M.M.; Simonutti, M.; Bouzo, C.A.; Turowski, V.R.; Perez, A.A.; Llugany, M.; Sánchez-Martín, M.-J.; et al. Enhancing Nutritional and Functional Properties of Broccoli Leaves Through Selenium Biofortification: Potential for Sustainable Agriculture and Bioactive Compound Valorization. Agronomy 2025, 15, 389. https://doi.org/10.3390/agronomy15020389
Buttarelli MS, Céccoli G, Trod BS, Stoffel MM, Simonutti M, Bouzo CA, Turowski VR, Perez AA, Llugany M, Sánchez-Martín M-J, et al. Enhancing Nutritional and Functional Properties of Broccoli Leaves Through Selenium Biofortification: Potential for Sustainable Agriculture and Bioactive Compound Valorization. Agronomy. 2025; 15(2):389. https://doi.org/10.3390/agronomy15020389
Chicago/Turabian StyleButtarelli, Melina Soledad, Gabriel Céccoli, Betiana Soledad Trod, María Micaela Stoffel, Marianela Simonutti, Carlos Alberto Bouzo, Valeria Rosana Turowski, Adrián Alejandro Perez, Mercè Llugany, María-Jesús Sánchez-Martín, and et al. 2025. "Enhancing Nutritional and Functional Properties of Broccoli Leaves Through Selenium Biofortification: Potential for Sustainable Agriculture and Bioactive Compound Valorization" Agronomy 15, no. 2: 389. https://doi.org/10.3390/agronomy15020389
APA StyleButtarelli, M. S., Céccoli, G., Trod, B. S., Stoffel, M. M., Simonutti, M., Bouzo, C. A., Turowski, V. R., Perez, A. A., Llugany, M., Sánchez-Martín, M.-J., Daurelio, L. D., Guevara, M. G., & Muñoz, F. F. (2025). Enhancing Nutritional and Functional Properties of Broccoli Leaves Through Selenium Biofortification: Potential for Sustainable Agriculture and Bioactive Compound Valorization. Agronomy, 15(2), 389. https://doi.org/10.3390/agronomy15020389