
Differences in Fatty Acid and Central Carbon Metabolite Distribution among Different Tissues of Alfalfa–Rhizobia Symbiotic System

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Rhizobia Strains, and Treatments
2.2. Indicators Measurement
2.2.1. Plant Phenotype
2.2.2. Nodule Formation
2.2.3. Nitrogen Fixation Capacity
2.2.4. Activity of Key Enzymes Involved in Nitrogen Metabolism
2.3. Sample Collection for Targeted Metabolomics
Quantification of Central Carbon and F.A. Metabolites
2.4. Data Analysis
3. Results
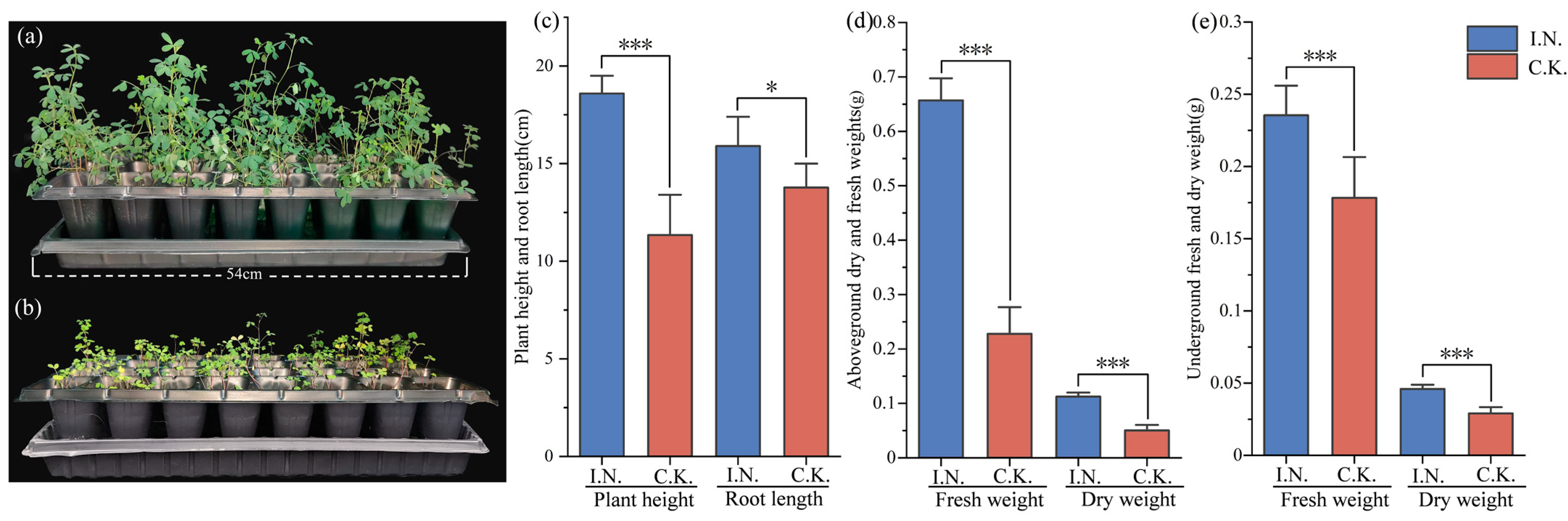
3.1. Plant Phenotype and Biomass
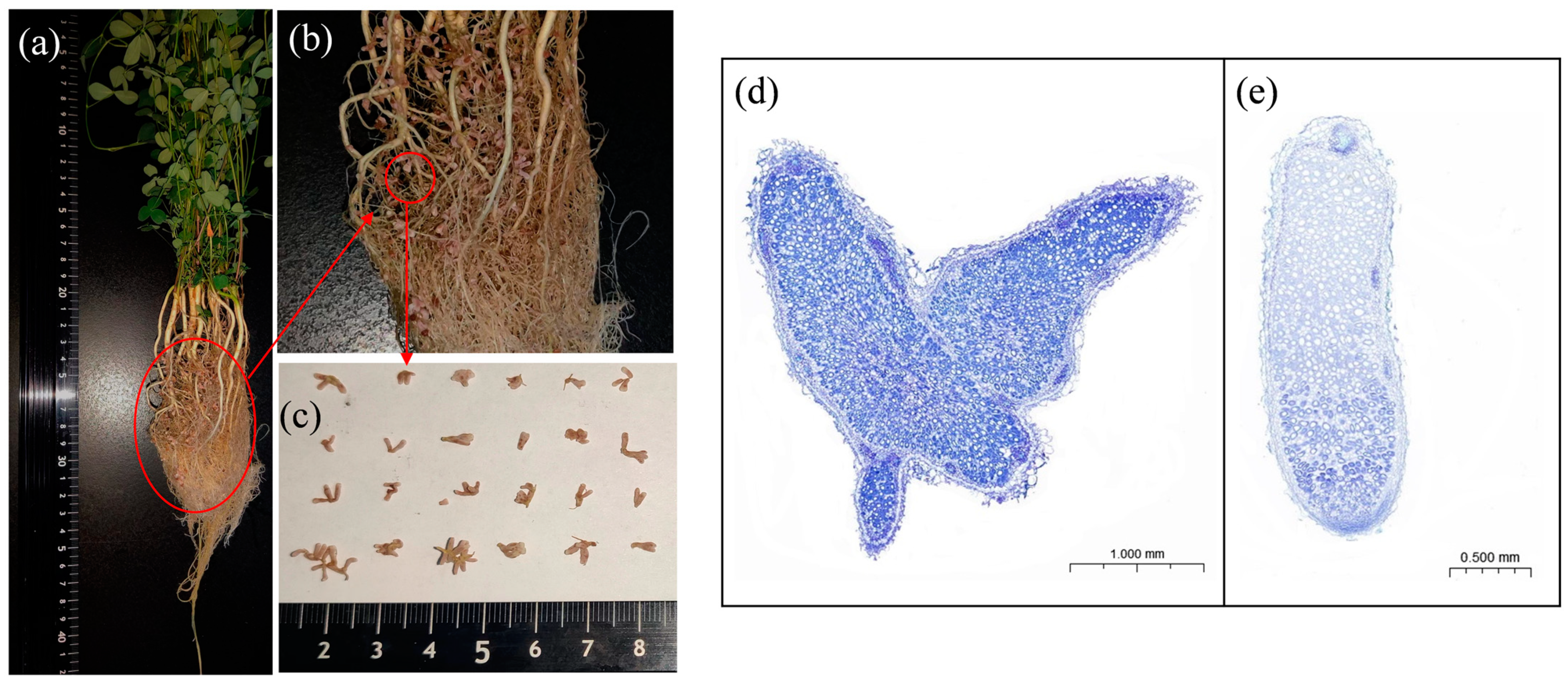
3.2. Nodulation and Nitrogen Fixation Capacity
3.3. Activity of Key Enzymes Involved in Nitrogen Metabolism
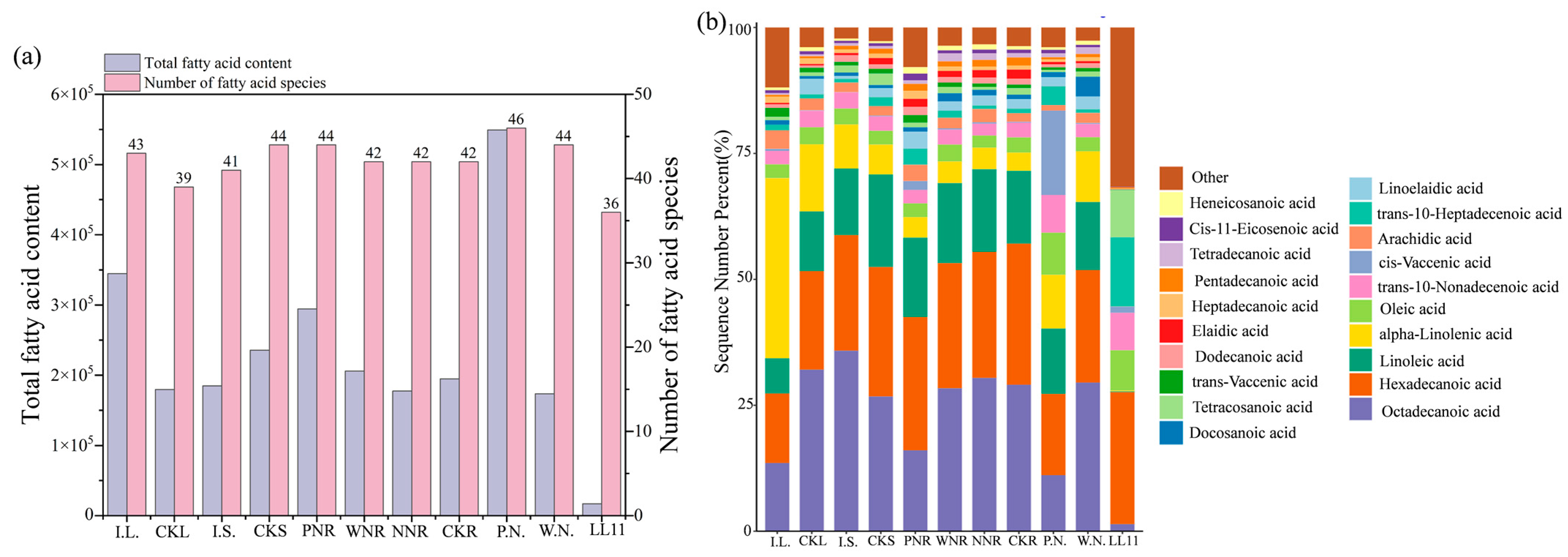
3.4. F.A. and Central Carbon-Targeted Metabolomic Analysis of Rhizobia
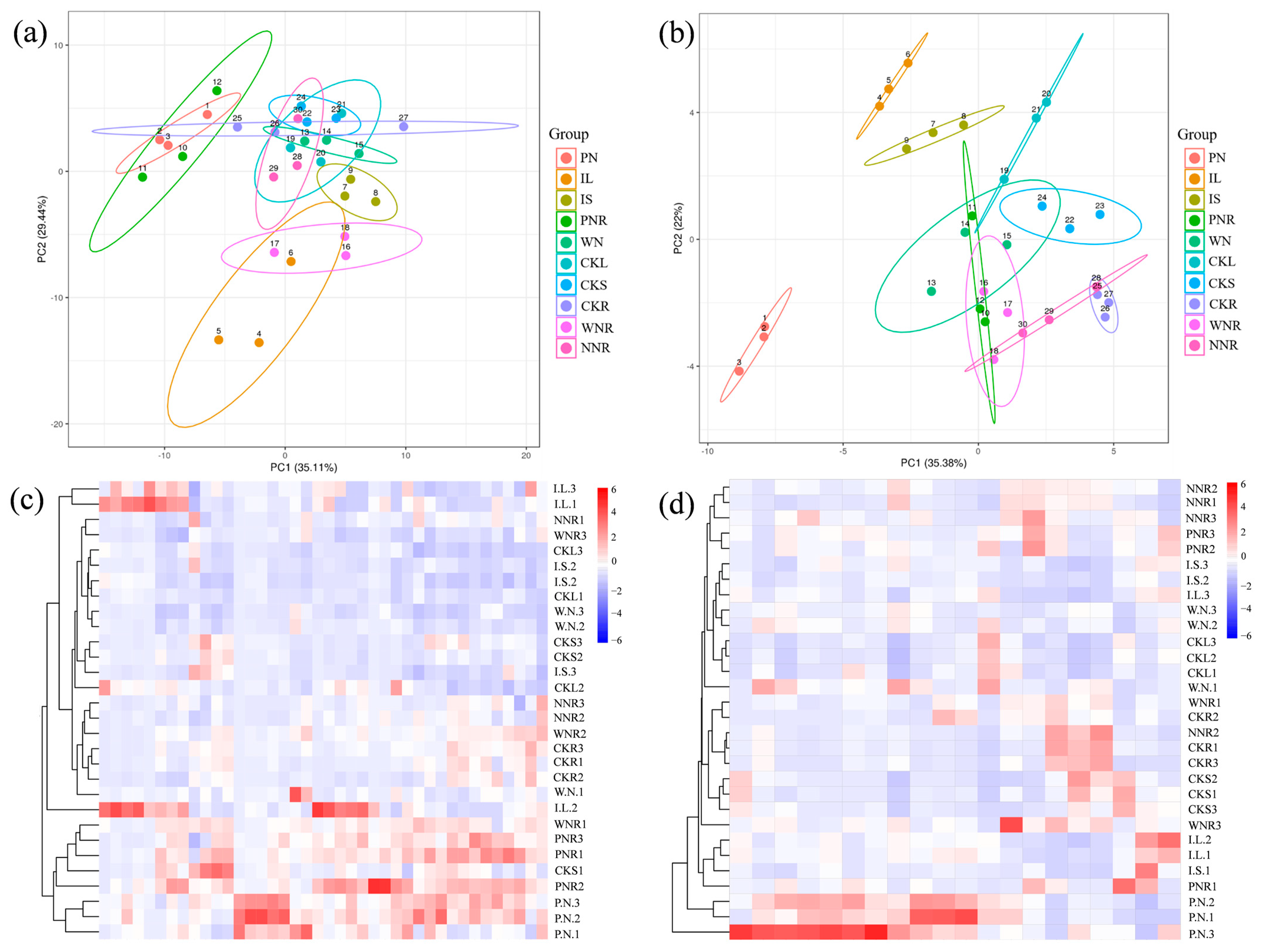
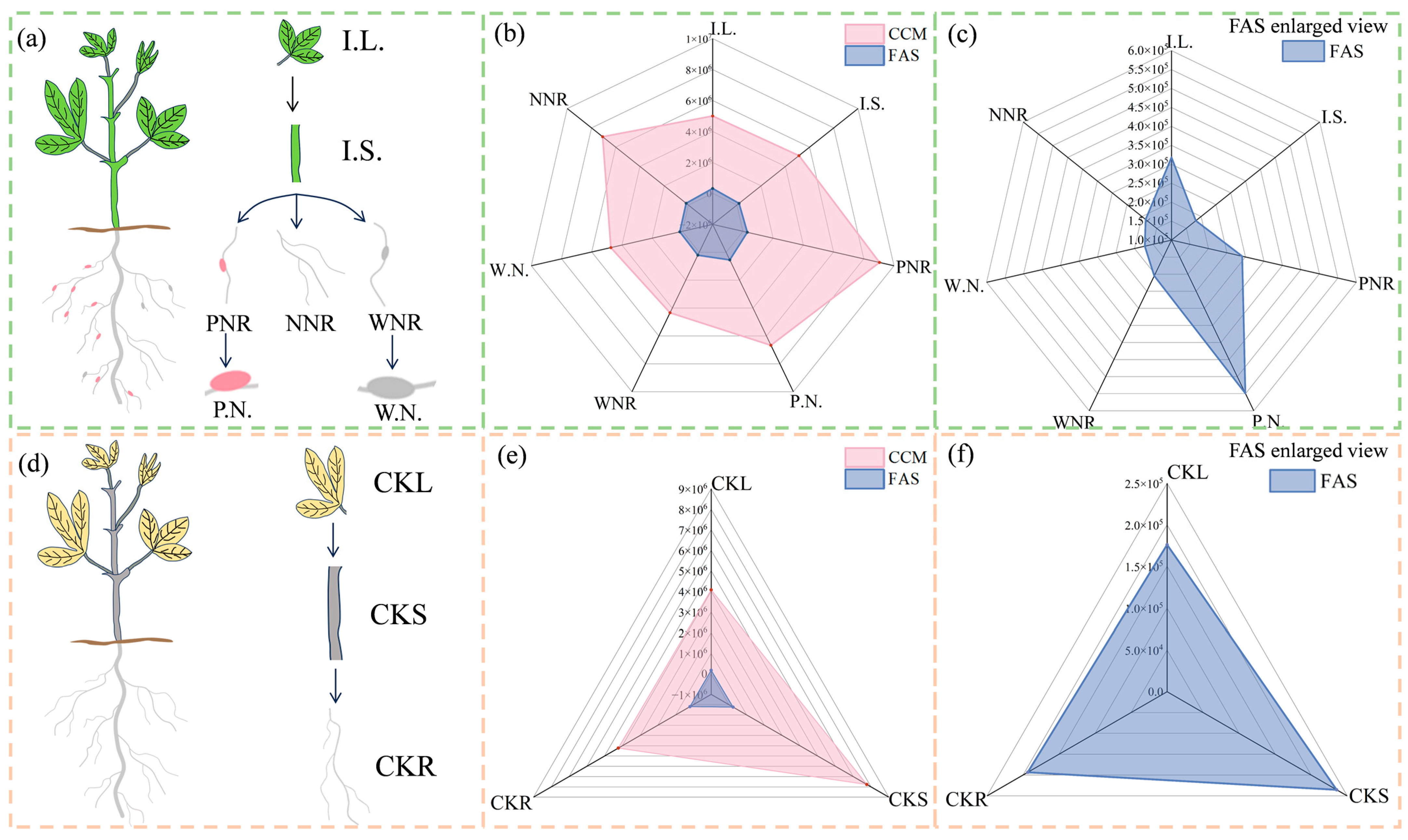
3.5. F.A. and Central Carbon-Targeted Metabolic Analyses of Plant Tissues
3.5.1. Differences in Leaf and Stem Metabolism
3.5.2. Differences in Root Metabolism
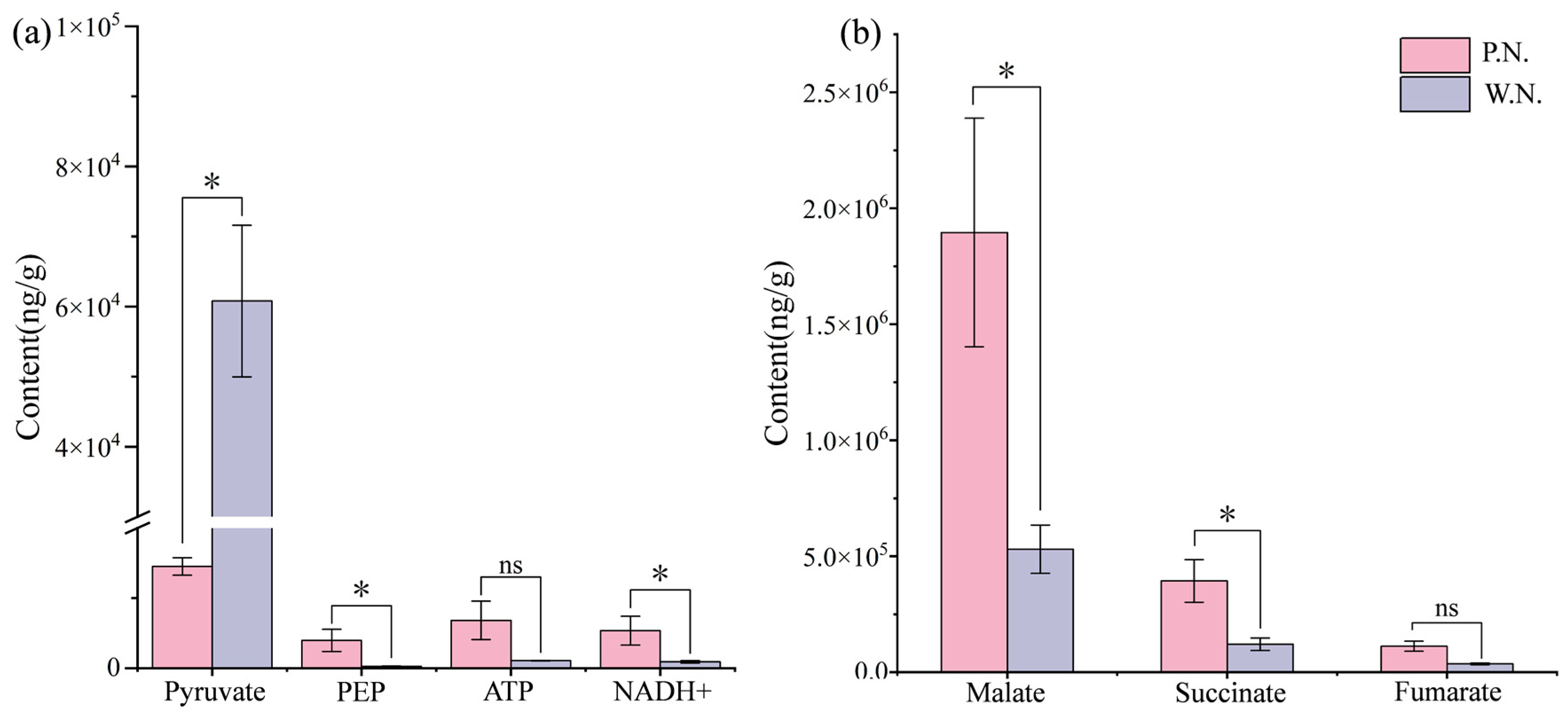
3.5.3. Differences in Nodule Metabolism
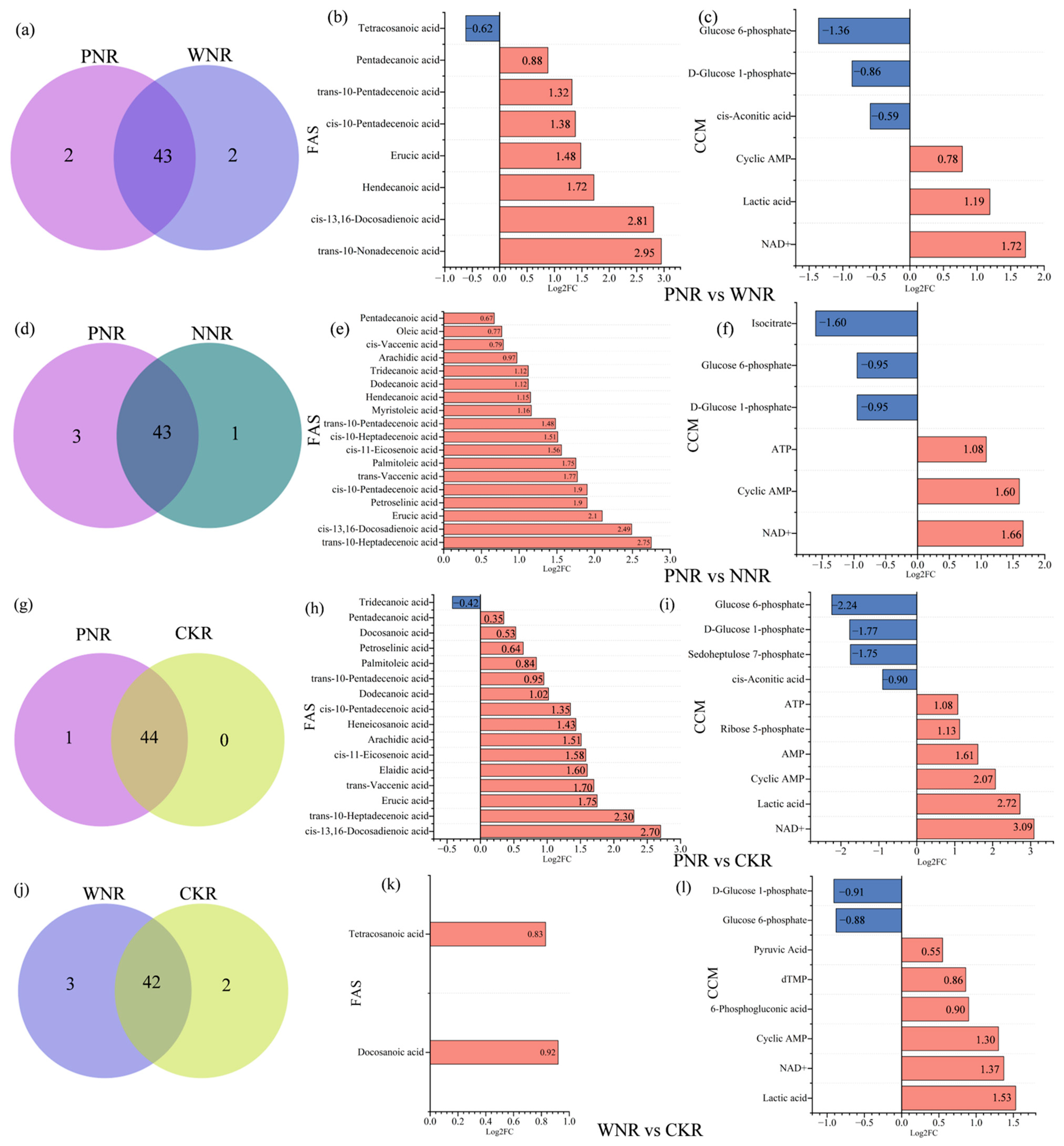
3.5.4. Differences in Root and Nodule Metabolism
3.6. Differences in F.A. Metabolism between Rhizobia and Symbiotic Plants
3.7. Differences in F.A. and Central Carbon Changes between Symbiotic and C.K. Plants
4. Discussion
4.1. Effects of Rhizobia Inoculation on Plant Biomass, Nodules, Nitrogen Fixation, and Key Enzymes Related to Nitrogen Metabolism
4.2. Differences in Energy Metabolism among Different Tissues
4.2.1. Differences in Central Carbon Metabolites
4.2.2. Differences in F.A. Metabolite Abundance
4.2.3. Differences in F.A. and Central Carbon Content between Symbiotic and C.K. Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medina, C.A.; Kaur, H.; Ray, I.; Yu, L.X. Strategies to Increase Prediction Accuracy in Genomic Selection of Complex Traits in Alfalfa (Medicago sativa L.). Cells 2021, 10, 3372. [Google Scholar] [CrossRef] [PubMed]
- Haag, A.F.; Arnold, M.F.; Myka, K.K.; Kerscher, B.; Dall’Angelo, S.; Zanda, M.; Mergaert, P.; Ferguson, G.P. Molecular insights into bacteroid development during Rhizobia—Legume symbiosis. FEMS Microbiol. Rev. 2013, 37, 364–383. [Google Scholar] [CrossRef] [PubMed]
- Booth, N.J.; Smith, P.M.C.; Ramesh, S.A.; Day, D.A. Malate Transport and Metabolism in Nitrogen-Fixing Legume Nodules. Molecules 2021, 26, 6876. [Google Scholar] [CrossRef] [PubMed]
- diCenzo, G.C.; Tesi, M.; Pfau, T.; Mengoni, A.; Fondi, M. Genome-scale metabolic reconstruction of the symbiosis between a leguminous plant and a nitrogen-fixing bacterium. Nat. Commun. 2020, 11, 2574. [Google Scholar] [CrossRef]
- Zhang, G.; Ahmad, M.Z.; Chen, B.; Manan, S.; Zhang, Y.; Jin, H.; Wang, X.; Zhao, J. Lipidomic and transcriptomic profiling of developing nodules reveals the essential roles of active glycolysis and fatty acid and membrane lipid biosynthesis in soybean nodulation. Plant J. 2020, 103, 1351–1371. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef]
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by rhizobia-the roots of a success story. Curr. Opin. Plant Biol. 2018, 44, 7–15. [Google Scholar] [CrossRef]
- Sulieman, S.; Tran, L.S. Symbiotic nitrogen fixation in legume nodules: Metabolism and regulatory mechanisms. Int. J. Mol. Sci. 2014, 15, 19389–19393. [Google Scholar] [CrossRef]
- Yang, J.; Liu, C.W.; Li, X. Advances in the legume–rhizobia symbiosis. Plant Physiol. J. 2023, 59, 1407–1435. [Google Scholar] [CrossRef]
- Wang, D.; Dong, W.; Murray, J.; Wang, E. Innovation and appropriation in mycorrhizal and rhizobial Symbioses. Plant Cell 2022, 34, 1573–1599. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef]
- Keymer, A.; Gutjahr, C. Cross-kingdom lipid transfer in arbuscular mycorrhiza symbiosis and beyond. Curr. Opin. Plant Biol. 2018, 44, 137–144. [Google Scholar] [CrossRef]
- Luginbuehl, L.; HMenard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.C.C.R.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.L. Study Mesorhizobia Huakuii 7653R Bacteroid Differentiation and Nitrogen Fixation Mechanism Based Global Transcriptomr Analysis. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2014. [Google Scholar]
- Dai, J. Involvement of Utilization of the Plant-Derived Sugars and Lipids and the Protease CSE1 in the Symbiotic Interactions between Metarhizium Robertsii and Plants. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2022. [Google Scholar] [CrossRef]
- Xu, H.; Li, Z.; Tong, Z.; He, F.; Li, X. Metabolomic analyses reveal substances that contribute to the increased freezing tolerance of alfalfa (Medicago sativa L.) after continuous water deficit. BMC Plant Biol. 2020, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Frank, M.; Reid, D. No Home without Hormones: How Plant Hormones Control Legume Nodule Organogenesis. Plant Commun. 2020, 1, 100104. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Chen, W.; Wang, W.; Zhang, H.; Liu, X.; Luo, J. Comprehensive profiling and natural variation of flavonoids in rice. J. Integr. Plant Biol. 2014, 56, 876–886. [Google Scholar] [CrossRef]
- Lv, H.; Guo, S. Comparative analysis of flavonoid metabolites from different parts of Hemerocallis citrina. BMC Plant Biol. 2023, 23, 491. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Cao, X.; Wei, S.; Huang, X.; Ouyang, H.; Chang, Y.; Shi, R.; He, J. Quantitative Comparison and Chemical Profile of Different Botanical Parts of Panax notoginseng from Different Regions. Front. Nutr. 2022, 9, 841541. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, W.; Liu, C.; Chen, P.; Zhou, L. Study on the accumulation pattern of anthocyanins, sugars and organic acids in medicinal Vitis vinifera ‘SuoSuo’ during ripening. Food Chem. 2024, 433, 137294. [Google Scholar] [CrossRef]
- He, L.; Shi, S.L.; Kang, W.J.; Liu, C.C.; Wang, W.J.; Wu, B. Location and Sterilization of Endogenous Rhizobia in Alfalfa Seeds. Acta Sgrestia Sin. 2022, 30, 2892–2898. [Google Scholar]
- Kang, W.J.; Lu, B.F.; Guan, J.; He, L.; Liu, C.C.; Nan, P.; Ma, R.H. A Hydroponic Device That Is Easy to Move and Observe the Nodulation Status of Alfalfa Roots. Patent CN202222289363.4, 14 March 2023. [Google Scholar]
- Wang, S.Q.; Han, X.Z.; Qiao, Y.F.; Yan, J.; Li, X.H. Effects of low molecular organic acids on nitrogen accumulation, nodulation, and nitrogen fixation of soybean (Glycine max L.) under phosphorus deficiency stress. Chin. J. Appl. Ecol. 2009, 20, 1079–1084. [Google Scholar] [CrossRef]
- Zuo, Y.M.; Liu, Y.X.; Zhang, F.S. Effects of Improvement of Iron Nutrition by Mixed Cropping with Maize on Nodule Microstructure and Leghaemoglobin Content of Peanut. J. Plant Physiol. Mol. Biol. 2003, 29, 33–38. [Google Scholar]
- Basile, L.A.; Lepek, V.C. Legume-rhizobia dance: An agricultural tool that could be improved? Microb. Biotechnol. 2021, 14, 1897–1917. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, Y.; Yang, X.; Deng, L.; Lu, X. Enhancing Soybean Yield: The Synergy of Sulfur and Rhizobia Inoculation. Plants 2023, 12, 3911. [Google Scholar] [CrossRef] [PubMed]
- Allito, B.B.; Ewusi-Mensah, N.; Logah, V.; Hunegnaw, D.K. Legume-rhizobia specificity effect on nodulation, biomass production and partitioning of faba bean (Vicia faba L.). Sci. Rep. 2021, 11, 3678. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.H.; Sui, X.H.; Hu, Y.G.; Chen, D.M.; Chen, W.X.; Gao, R.l. 3Screening of highly-effective Sinorhizobia meliloti strains for Medicago sativa cultivars and their field inoculation. Acta Prataculturae Sin. 2004, 13, 95–100. [Google Scholar]
- Li, S.S.; Yang, Z.; Li, H.; Yang, W.G.; Wang, X.L.; Chai, H.; Wu, Y. Effect of rhizobia inoculation on yield and quality of alfalfa. Heilongjiang Anim. Sci. Vet. Med. 2021, 20, 98–101+107. [Google Scholar] [CrossRef]
- Li, X.L.; Gao, D.S.; Mi, R.F. Study on the niteate reductase activity of apple trees in orchard. J. Shandong Agric. Univ. 1997, 3–6. [Google Scholar]
- Li, Y.; Ma, C.M.; Gong, Z.P. Study on glutamine synthetase activity in spring soybean. J. Northeast. Agric. Univ. 2006, 729–732. [Google Scholar] [CrossRef]
- Sun, F. Studies on Enzyme Activities of Nitrogen Metabolism and the Content of Nitrogen-Containing Compounds in Roots of Soybean Varieties Released at Different Age. Ph.D. Thesis, Jilin Agricultural University, Changchun, China, 2020. [Google Scholar] [CrossRef]
- Liu, S.X. Study on the Mechanism of Sinorhizobia Meliloti SD101 Improving Salt Tolerance of Alfalfa (Medicago sativa L.). Ph.D. Thesis, Heilongjiang Bayi Agricultural University, Heilongjiang, China, 2022. [Google Scholar] [CrossRef]
- Yoneyama, T.; Fujimori, T.; Yanagisawa, S.; Hase, T.; Suzuki, A. 15N Tracing Studies on In Vitro Reactions of Ferredoxin-Dependent Nitrite Reductase and Glutamate Synthase Using Reconstituted Electron Donation Systems. Plant Cell Physiol. 2015, 56, 1154–1161. [Google Scholar] [CrossRef] [PubMed]
- Brechenmacher, L.; Lei, Z.; Libault, M.; Findley, S.; Sugawara, M.; Sadowsky, M.J.; Sumner, L.W.; Stacey, G. Soybean metabolites regulated in root hairs in response to the symbiotic bacterium Bradyrhizobia japonicum. Plant Physiol. 2010, 153, 1808–1822. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Teng, C.C.; Liu, Y.J.; Zhang, J.F.; Hou, W.W.; He, T.; Zhang, X.L.; Wang, J.Z. Identification of Vicia faba Rhizobia and drought resistance verification in arid region of Qinghai. Acta Microbiol. Sin. 2022, 62, 4030–4046. [Google Scholar] [CrossRef]
- Orgambide, G.G.; Huang, Z.H.; Gage, D.A.; Dazzo, F.B. Phospholipid and fatty acid compositions of Rhizobia leguminosarum biovar trifolii ANU843 in relation to flavone-activated pSym nod gene expression. Lipids 1993, 28, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.M. Cloning and Functional Analysis of FAD2 Gene Family of Artemisia sphaerocephala and Evaluation of Transgenic Alfalfa. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2021. [Google Scholar] [CrossRef]
- Fan, Q.; Creamer, R.; Li, Y. Time-course metabolic profiling in alfalfa leaves under Phoma medicaginis infection. PLoS ONE. 2018, 13, e0206641. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of Omega-3 Fatty Acids on Immune Cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef]
- Ogden, A.J. Metabolic Progression during Development of Symbiotic Nitrogen Fixation in the Medicago-Sinorhizobia Symbiosis and the Role of HslUV and ClpXP Protease Machinery; Washington State University: Washington, DC, USA, 2018. [Google Scholar]
- Colebatch, G.; Desbrosses, G.; Ott, T.; Krusell, L.; Montanari, O.; Kloska, S.; Kopka, J.; Udvardi, M.K. Global changes in transcription orchestrate metabolic differentiation during symbiotic nitrogen fixation in Lotus japonicus. Plant J. 2004, 39, 487–512. [Google Scholar] [CrossRef]
- Théberge, M.C.; Prévost, D.; Chalifour, F.P. The effect of different temperatures on the fatty acid composition of Rhizobia leguminosarum bv. viciae in the faba bean symbiosis. New Phytol. 1996, 134, 657–664. [Google Scholar] [CrossRef]
- Ledermann, R.; Schulte, C.C.M.; Poole, P.S. How Rhizobia Adapt to the Nodule Environment. J. Bacteriol. 2021, 203, e0053920. [Google Scholar] [CrossRef] [PubMed]
- Westhoek, A.; Clark, L.J.; Culbert, M.; Dalchau, N.; Griffiths, M.; Jorrin, B.; Karunakaran, R.; Ledermann, R.; Tkacz, A.; Webb, I.; et al. Conditional sanctioning in a legume—Rhizobia mutualism. Proc. Natl. Acad. Sci. USA 2021, 118, e2025760118. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Site | Abbreviation |
|---|---|---|
| Inoculation of rhizobia LL11 | Leaf | I.L. |
| Stem | I.S. | |
| Pink nodule roots | PNR | |
| White nodule roots | WNR | |
| No nodule roots | NNR | |
| Pink root nodules | P.N. | |
| White root nodules | W.N. | |
| Inoculation of the same amount of sterile water C.K. | C.K. Leaves | CKL |
| C.K. stems | CKS | |
| C.K. roots | CKR | |
| Sinorhizobia meliloti LL11 | Thallus | LL11 |
| Items | I.N. | C.K. |
|---|---|---|
| Number of effective nodules | 7.100 ± 0.8307 | 0. |
| Number of invalid nodules | 1.7143 ± 0.6999 | 0 |
| Nodule weight per plant(g) | 0.0295 ± 0.0009 | 0 |
| Nodule diameter(mm) | 0.8731 ± 0.3057 | 0 |
| Nitrogenase activity(μmol·g−1·h−1) | 0.962 ± 0.0462 | 0 |
| Leghemoglobin content(mg·g−1) | 2.5636 ± 0.0548 | 0 |
| Mean nitrogen fixation potential (μmol·h−1) | 0.0283 ± 0.0011 | 0 |
| Group | Saturated Fatty Acid (SFA) | Unsaturated Fatty Acid (UFA) | ||||
|---|---|---|---|---|---|---|
| Species Number | Total Content (ng) | Percentage (%) | Species Number | Total Content (ng) | Percentage (%) | |
| I.L. | 15 | 114,204.01 | 33.03 | 28 | 231,581.54 | 66.97 |
| I.S. | 13 | 121,995.24 | 65.95 | 28 | 62,978.56 | 34.05 |
| PNR | 14 | 148,878.02 | 51.84 | 30 | 138,322.19 | 48.16 |
| WNR | 14 | 131,443.54 | 63.85 | 31 | 74,425.38 | 36.15 |
| NNR | 14 | 118,166.67 | 66.59 | 28 | 59,299.41 | 33.41 |
| P.N. | 14 | 175,968.99 | 32.04 | 32 | 373,254.57 | 67.96 |
| W.N. | 14 | 108,971.11 | 62.81 | 29 | 64,513.52 | 37.19 |
| CKL | 12 | 100,687.87 | 55.92 | 30 | 79,378.65 | 44.08 |
| CKS | 13 | 144,945.31 | 61.50 | 28 | 90,805.00 | 38.50 |
| CKR | 14 | 131,650.49 | 67.57 | 30 | 63,196.08 | 32.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, B.-F.; Kang, W.-J.; Shi, S.-L.; Guan, J.; Jing, F.; Wu, B. Differences in Fatty Acid and Central Carbon Metabolite Distribution among Different Tissues of Alfalfa–Rhizobia Symbiotic System. Agronomy 2024, 14, 511. https://doi.org/10.3390/agronomy14030511
Lu B-F, Kang W-J, Shi S-L, Guan J, Jing F, Wu B. Differences in Fatty Acid and Central Carbon Metabolite Distribution among Different Tissues of Alfalfa–Rhizobia Symbiotic System. Agronomy. 2024; 14(3):511. https://doi.org/10.3390/agronomy14030511
Chicago/Turabian StyleLu, Bao-Fu, Wen-Juan Kang, Shang-Li Shi, Jian Guan, Fang Jing, and Bei Wu. 2024. "Differences in Fatty Acid and Central Carbon Metabolite Distribution among Different Tissues of Alfalfa–Rhizobia Symbiotic System" Agronomy 14, no. 3: 511. https://doi.org/10.3390/agronomy14030511
APA StyleLu, B.-F., Kang, W.-J., Shi, S.-L., Guan, J., Jing, F., & Wu, B. (2024). Differences in Fatty Acid and Central Carbon Metabolite Distribution among Different Tissues of Alfalfa–Rhizobia Symbiotic System. Agronomy, 14(3), 511. https://doi.org/10.3390/agronomy14030511