

Microbial Community in the Composting Process and Its Positive Impact on the Soil Biota in Sustainable Agriculture

 ,
,  and
and 
Abstract
1. Introduction
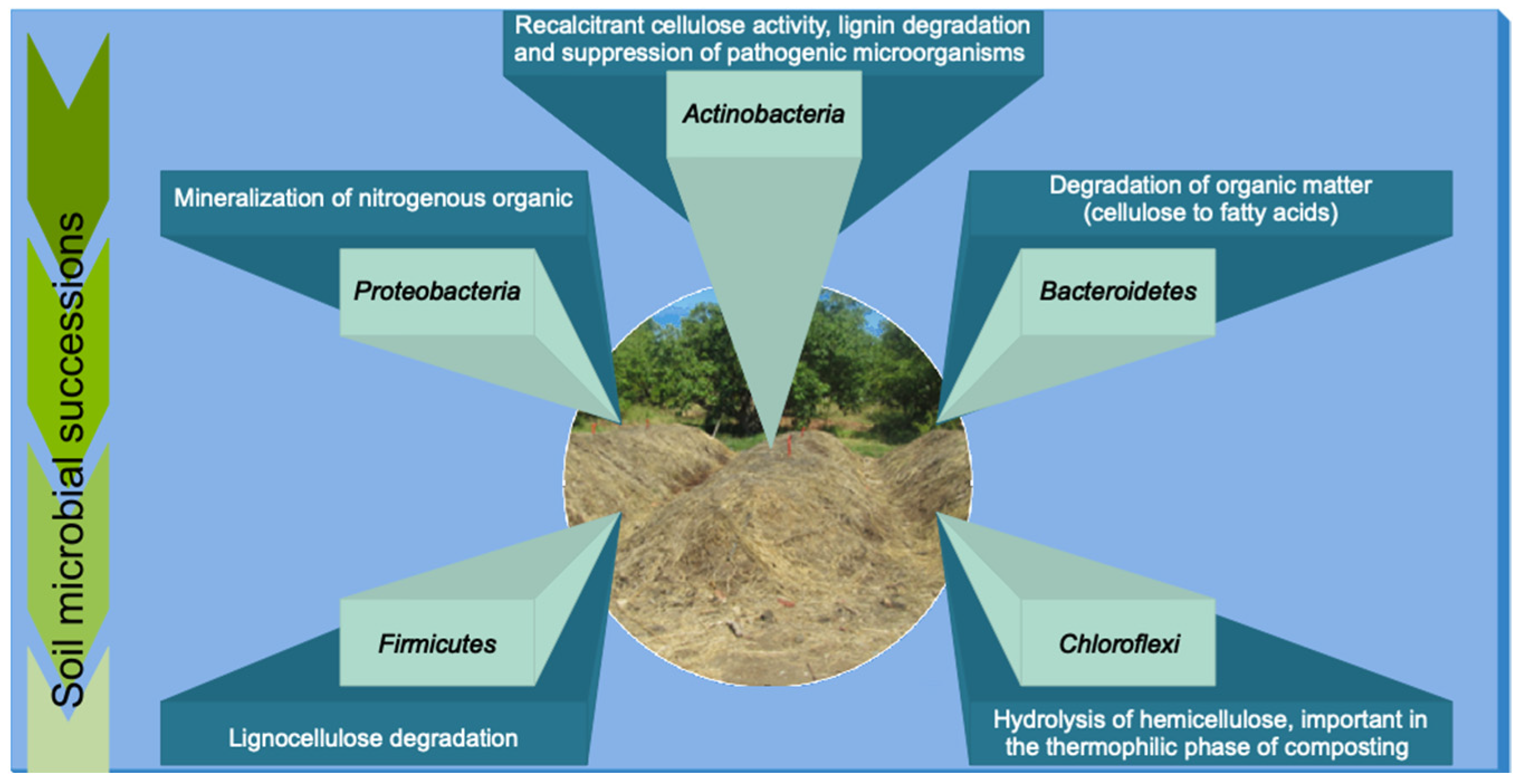
2. The Microbiota in the Composting Process
2.1. Firmicutes
2.2. Actinobacteria
2.3. Proteobacteria
2.4. Bacteroidetes
2.5. Chloroflexi
2.6. Fungi
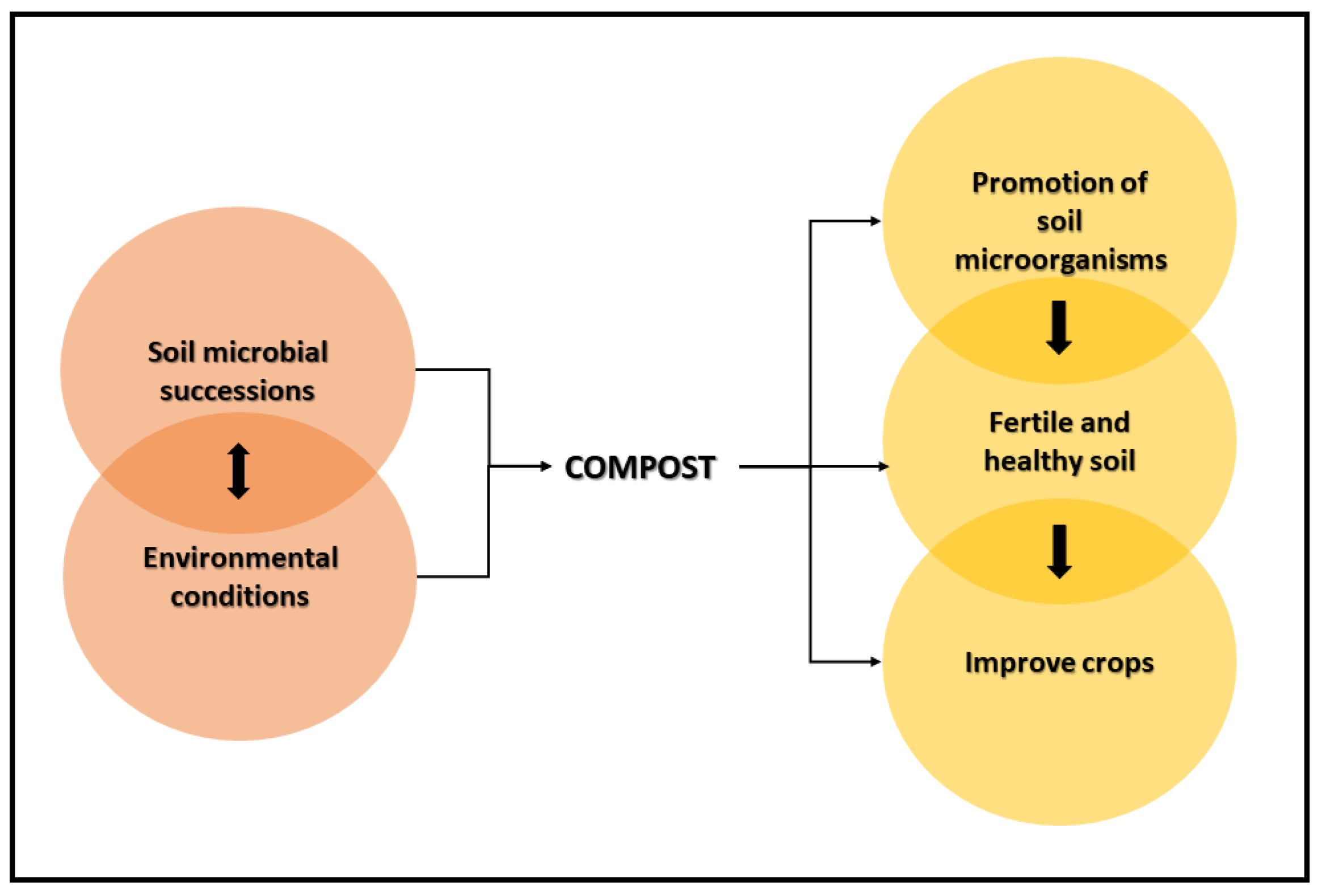
3. Conditions That Affect the Succession of Microbial Communities in the Composting Process
3.1. Temperature

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composting Stages | Dominant Phyla | Maximum Temperatures (°C) | References |
|---|---|---|---|
| Mesophilic | Actinobacterias | >25 °C | [78] |
| Firmicutes | |||
| Proteobacterias | |||
| Thermophilic | Actinobacterias Firmicutes | 50–65 °C | [86] |
| Cooling | Bacteroidetes | 50 to 40 °C | [86] |
| Maturationn | Chloroflexi | 40 °C to room temperature | [86] |
3.2. Aerobiosis
3.3. pH
3.4. Humidity
3.5. Raw Materials/Composting Methods
3.6. C/N
3.7. Pathogens in the Composting Process
3.8. Enrichment of the Composting Process with External Inoculums
4. Promotion of Soil Microorganisms by Compost Application
4.1. Fungal Community
4.2. Arbuscular Mycorrhizal Fungi (AMF)
4.3. Bacterial Community in Agricultural Soil
4.4. Plant Growth Promoting Bacteria (PGPR)
4.5. Other Microorganisms
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.E.; Gulledge, J.; Engelhaupt, E.; Burow, M.E.; McLachlan, J.A. Pesticides reduce symbiotic efficiency of nitrogen-fixing rhizobia and host plants. Proc. Natl. Acad. Sci. USA 2007, 104, 10282–10287. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Paredes, A.; Valdés, G.; Nuti, M. Ecosystem Functions of Microbial Consortia in Sustainable Agriculture. Agronomy 2020, 10, 1902. [Google Scholar] [CrossRef]
- Yang, W.; Guo, Y.; Wang, X.; Chen, C.; Hu, Y.; Cheng, L.; Gu, S.; Xu, X. Temporal variations of soil microbial community under compost addition in black soil of Northeast China. Appl. Soil Ecol. 2017, 121, 214–222. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and Soil Microbial Community: A Review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- Ma, S.; Xiong, J.; Cui, R.; Sun, X.; Han, L.; Xu, Y.; Kan, Z.; Gong, X.; Huang, G. Effects of intermittent aeration on greenhouse gas emissions and bacterial community succession during large-scale membrane-covered aerobic composting. J. Clean. Prod. 2020, 266, 121551. [Google Scholar] [CrossRef]
- Scotti, R.; Pane, C.; Spaccini, R.; Palese, A.M.; Piccolo, A.; Celano, G.; Zaccardelli, M. On-farm compost: A useful tool to improve soil quality under intensive farming systems. Appl. Soil Ecol. 2016, 107, 13–23. [Google Scholar] [CrossRef]
- Celik, I.; Ortas, I.; Kilic, S. Effects of compost, mycorrhiza, manure and fertilizer on some physical properties of a Chromoxerert soil. Soil Tillage Res. 2004, 78, 59–67. [Google Scholar] [CrossRef]
- Duong, T.T.T.; Penfold, C.; Marschner, P. Differential effects of composts on properties of soils with different textures. Biol. Fertil. Soils 2012, 48, 699–707. [Google Scholar] [CrossRef]
- Toundou, O.; Pallier, V.; Feuillade-Cathalifaud, G.; Tozo, K. Impact of agronomic and organic characteristics of waste composts from Togo on Zea mays L. nutrients contents under water stress. J. Environ. Manag. 2021, 285, 112158. [Google Scholar] [CrossRef]
- Luo, X.; Liu, G.; Xia, Y.; Chen, L.; Jiang, Z.; Zheng, H.; Wang, Z. Use of biochar-compost to improve properties and productivity of the degraded coastal soil in the Yellow River Delta, China. J. Soils Sediments 2017, 17, 780–789. [Google Scholar] [CrossRef]
- Donn, S.; Wheatley, R.E.; McKenzie, B.M.; Loades, K.W.; Hallett, P.D. Improved soil fertility from compost amendment increases root growth and reinforcement of surface soil on slopes. Ecol. Eng. 2014, 71, 458–465. [Google Scholar] [CrossRef]
- Siedt, M.; Schäffer, A.; Smith, K.E.; Nabel, M.; Roß-Nickoll, M.; van Dongen, J.T. Comparing straw, compost, and biochar regarding their suitability as agricultural soil amendments to affect soil structure, nutrient leaching, microbial communities, and the fate of pesticides. Sci. Total. Environ. 2021, 751, 141607. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Piccolo, A.; Spaccini, R.; Celano, G.; Villecco, D.; Zaccardelli, M. Agricultural waste-based composts exhibiting suppressivity to diseases caused by the phytopathogenic soil-borne fungi Rhizoctonia solani and Sclerotinia minor. Appl. Soil Ecol. 2013, 65, 43–51. [Google Scholar] [CrossRef]
- Gao, X.; Tan, W.; Zhao, Y.; Wu, J.; Sun, Q.; Qi, H.; Xie, X.-Y.; Wei, Z. Diversity in the Mechanisms of Humin Formation during Composting with Different Materials. Environ. Sci. Technol. 2019, 53, 3653–3662. [Google Scholar] [CrossRef] [PubMed]
- Cossu, A.; Degl’Innocenti, S.; Agnolucci, M.; Cristani, C.; Bedini, S.; Nuti, M. Assessment of the Life Cycle Environmental Impact of the Olive Oil Extraction Solid Wastes in the European Union. Open Waste Manag. J. 2013, 6, 12–20. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Zhang, Z.; Wang, Q.; Shen, F.; Li, R.; Li, D.-S.; Ren, X.; Wang, M.; Chen, H.; Zhao, J. New insight with the effects of biochar amendment on bacterial diversity as indicators of biomarkers support the thermophilic phase during sewage sludge composting. Bioresour. Technol. 2017, 238, 589–601. [Google Scholar] [CrossRef]
- Santos, J.; Nunes, L.; de Melo, W.J.; Araújo, A. Tannery sludge compost amendment rates on soil microbial biomass of two different soils. Eur. J. Soil Biol. 2011, 47, 146–151. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Wang, H.; Lu, Q.; Cao, Z.; Cui, H.; Zhu, L.; Wei, Z. An optimized regulating method for composting phosphorus fractions transformation based on biochar addition and phosphate-solubilizing bacteria inoculation. Bioresour. Technol. 2016, 221, 139–146. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, Y.; Zhang, W.; Zhou, H.; Chen, X.; Li, Y.; Wei, D.; Wei, Z. Roles of bacterial community in the transformation of organic nitrogen toward enhanced bioavailability during composting with different wastes. Bioresour. Technol. 2019, 285, 121326. [Google Scholar] [CrossRef]
- Karadag, D.; Özkaya, B.; Ölmez, E.; Nissilä, M.E.; Çakmakçı, M.; Yıldız, S.; Puhakka, J.A. Profiling of bacterial community in a full-scale aerobic composting plant. Int. Biodeterior. Biodegrad. 2013, 77, 85–90. [Google Scholar] [CrossRef]
- Wang, K.; Mao, H.; Li, X. Functional characteristics and influence factors of microbial community in sewage sludge composting with inorganic bulking agent. Bioresour. Technol. 2018, 249, 527–535. [Google Scholar] [CrossRef] [PubMed]
- De Gannes, V.; Eudoxie, G.; Hickey, W.J. Insights into fungal communities in composts revealed by 454-pyrosequencing: Implications for human health and safety. Front. Microbiol. 2013, 4, 164. [Google Scholar] [CrossRef] [PubMed]
- Siles, J.; García-Sánchez, M.; Gómez-Brandón, M. Studying Microbial Communities through Co-Occurrence Network Analyses during Processes of Waste Treatment and in Organically Amended Soils: A Review. Microorganisms 2021, 9, 1165. [Google Scholar] [CrossRef]
- Li, X.; Shi, X.-S.; Lu, M.-Y.; Zhao, Y.-Z.; Li, X.; Peng, H.; Guo, R.-B. Succession of the bacterial community and functional characteristics during continuous thermophilic composting of dairy manure amended with recycled ceramsite. Bioresour. Technol. 2019, 294, 122044. [Google Scholar] [CrossRef]
- Pankratov, T.A.; Ivanova, A.O.; Dedysh, S.; Liesack, W. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environ. Microbiol. 2011, 13, 1800–1814. [Google Scholar] [CrossRef]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, Q.; Wei, Y.; Cui, H.; Zhang, X.; Wang, X.; Shan, S.; Wei, Z. Effect of actinobacteria agent inoculation methods on cellulose degradation during composting based on redundancy analysis. Bioresour. Technol. 2016, 219, 196–203. [Google Scholar] [CrossRef]
- Quinn, G.A.; Banat, A.M.; Abdelhameed, A.M.; Banat, I.M. Streptomyces from traditional medicine: Sources of new innovations in antibiotic discovery. J. Med. Microbiol. 2020, 69, 1040–1048. [Google Scholar] [CrossRef]
- Wang, C.; Dong, D.; Wang, H.; Müller, K.; Qin, Y.; Wang, H.; Wu, W. Metagenomic analysis of microbial consortia enriched from compost: New insights into the role of Actinobacteria in lignocellulose decomposition. Biotechnol. Biofuels 2016, 9, 22. [Google Scholar] [CrossRef]
- Hashmi, I.; Bindschedler, S.; Junier, P. Firmicutes. In Beneficial Microbes in Agro-Ecology; Academic Press: Cambridge, MA, USA, 2020; pp. 363–396. [Google Scholar]
- Parkes, R.J.; Sass, H. Deep sub-surface. In Encyclopedia of Microbiology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Ryckeboer, J.; Mergaert, J.; Vaes, K.; Klammer, S.; De Clercq, D.; Coosemans, J.; Insam, H.; Swings, J. A survey of bacteria and fungi occurring during composting and self-heating processes. Ann. Microbiol. 2003, 53, 349–410. [Google Scholar]
- Yang, Y.; Awasthi, M.K.; Bao, H.; Bie, J.; Lei, S.; Lv, J. Exploring the microbial mechanisms of organic matter transformation during pig manure composting amended with bean dregs and biochar. Bioresour. Technol. 2020, 313, 123647. [Google Scholar] [CrossRef] [PubMed]
- Biyada, S.; Merzouki, M.; Dėmčėnko, T.; Vasiliauskienė, D.; Ivanec-Goranina, R.; Urbonavičius, J.; Marčiulaitienė, E.; Vasarevičius, S.; Benlemlih, M. Microbial community dynamics in the mesophilic and thermophilic phases of textile waste composting identified through next-generation sequencing. Sci. Rep. 2021, 11, 23624. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Duan, Y.; Awasthi, S.K.; Liu, T.; Zhang, Z. Effect of biochar and bacterial inoculum additions on cow dung composting. Bioresour. Technol. 2020, 297, 122407. [Google Scholar] [CrossRef] [PubMed]
- López, M.J.; Jurado, M.M.; López-González, J.A.; Estrella-González, M.J.; Martínez-Gallardo, M.R.; Toribio, A.; Suárez-Estrella, F. Characterization of Thermophilic Lignocellulolytic Microorganisms in Composting. Front. Microbiol. 2021, 12, 697480. [Google Scholar] [CrossRef]
- Moreno, J.; López-González, J.; Arcos-Nievas, M.; Suárez-Estrella, F.; Jurado, M.; Estrella-González, M.; López, M. Revisiting the succession of microbial populations throughout composting: A matter of thermotolerance. Sci. Total. Environ. 2021, 773, 145587. [Google Scholar] [CrossRef]
- Xu, S.; Lu, W.; Liu, Y.; Ming, Z.; Liu, Y.; Meng, R.; Wang, H. Structure and diversity of bacterial communities in two large sanitary landfills in China as revealed by high-throughput sequencing (MiSeq). Waste Manag. 2017, 63, 41–48. [Google Scholar] [CrossRef]
- Yin, Y.; Gu, J.; Wang, X.; Song, W.; Zhang, K.; Sun, W.; Zhang, X.; Zhang, Y.; Li, H. Effects of Copper Addition on Copper Resistance, Antibiotic Resistance Genes, and intl1 during Swine Manure Composting. Front. Microbiol. 2017, 8, 344. [Google Scholar] [CrossRef]
- Gavande, P.V.; Basak, A.; Sen, S.; Lepcha, K.; Murmu, N.; Rai, V.; Mazumdar, D.; Saha, S.P.; Das, V.; Ghosh, S. Functional characterization of thermotolerant microbial consortium for lignocellulolytic enzymes with central role of Firmicutes in rice straw depolymerization. Sci. Rep. 2021, 11, 3032. [Google Scholar] [CrossRef]
- Pepper, I.L.; Gentry, T.J. 2015. Earth Environments. In Environmental Microbiology; Gerpa, C.P., Ed.; Academic Press: Amsterdam, The Netherlands, 2015; pp. 59–88. [Google Scholar]
- Boubekri, K.; Soumare, A.; Mardad, I.; Lyamlouli, K.; Ouhdouch, Y.; Hafidi, M.; Kouisni, L. Multifunctional role of Actinobacteria in agricultural production sustainability: A review. Microbiol. Res. 2022, 261, 127059. [Google Scholar] [CrossRef]
- Kinkel, L.L.; Schlatter, D.C.; Bakker, M.; Arenz, B.E. Streptomyces competition and co-evolution in relation to plant disease suppression. Res. Microbiol. 2012, 163, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Ngalimat, M.S.; Mohd Hata, E.; Zulperi, D.; Ismail, S.I.; Ismail, M.R.; Mohd Zainudin, N.A.I.; Said, I.N.B.; Yusof, M.T. Plant Growth-Promoting Bacteria as an Emerging Tool to Manage Bacterial Rice Pathogens. Microorganisms 2021, 9, 682. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, G.; García-De-La-Fuente, R.; Abad, M.; Fornes, F. Isolation and identification of actinomycetes from a compost-amended soil with potential as biocontrol agents. J. Environ. Manag. 2010, 95, S280–S284. [Google Scholar] [CrossRef]
- Kausar, H.; Sariah, M.; Saud, H.M.; Alam, M.Z.; Ismail, M.R. Isolation and screening of potential actinobacteria for rapid composting of rice straw. Biodegradation 2010, 22, 367–375. [Google Scholar] [CrossRef]
- Insam, H.; de Bertoldi, M. Microbiology of the composting process. Waste Manag. Ser. 2007, 8, 25–48. [Google Scholar] [CrossRef]
- Rawat, S.; Johri, B.N. Role of Thermophilic Microflora in Composting. In Thermophilic Microbes in Environmental and Industrial Biotechnology; Satyanarayana, T., Littlechild, J., Kawarabayasi, Y., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 137–169. [Google Scholar] [CrossRef]
- Kersters, K.; De Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg,, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 3–37. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef]
- Mignot, T. The elusive engine in Myxococcus xanthus gliding motility. Cell. Mol. Life Sci. 2007, 64, 2733–2745. [Google Scholar] [CrossRef]
- Ishaq, S.L. Plant-microbial interactions in agriculture and the use of farming systems to improve diversity and productivity. AIMS Microbiol. 2017, 3, 335–353. [Google Scholar] [CrossRef]
- Ma, S.; Fang, C.; Sun, X.; Han, L.; He, X.; Huang, G. Bacterial community succession during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane under slight positive pressure. Bioresour. Technol. 2018, 259, 221–227. [Google Scholar] [CrossRef]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour. Technol. 2018, 256, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, G.; Fernández-González, A.J.; Lasa, A.V.; Aranda, E.; Torralbo, F.; González-Murua, C.; Fernández-López, M.; Benítez, E.; Bedmar, E.J. Involvement of the metabolically active bacteria in the organic matter degradation during olive mill waste composting. Sci. Total. Environ. 2021, 789, 147975. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Hehemann, J.-H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and Gut Bacteroidetes: The Food Connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed]
- Dodd, D.; Mackie, R.I.; Cann, I.K.O. Xylan degradation, a metabolic property shared by rumen and human colonic Bacteroidetes. Mol. Microbiol. 2011, 79, 292–304. [Google Scholar] [CrossRef]
- López-González, J.; Suárez-Estrella, F.; Vargas-García, M.; López, M.; Jurado, M.; Moreno, J. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity. Bioresour. Technol. 2015, 175, 406–416. [Google Scholar] [CrossRef]
- Islam, Z.F.; Cordero, P.R.F.; Feng, J.; Chen, Y.-J.; Bay, S.K.; Jirapanjawat, T.; Gleadow, R.M.; Carere, C.R.; Stott, M.B.; Chiri, E.; et al. Two Chloroflexi classes independently evolved the ability to persist on atmospheric hydrogen and carbon monoxide. ISME J. 2019, 13, 1801–1813. [Google Scholar] [CrossRef]
- Wan, J.; Wang, X.; Yang, T.; Wei, Z.; Banerjee, S.; Friman, V.-P.; Mei, X.; Xu, Y.; Shen, Q. Livestock Manure Type Affects Microbial Community Composition and Assembly During Composting. Front. Microbiol. 2021, 12, 621126. [Google Scholar] [CrossRef]
- Werner, K.A.; Poehlein, A.; Schneider, D.; El-Said, K.; Wöhrmann, M.; Linkert, I.; Hübner, T.; Brüggemann, N.; Prost, K.; Daniel, R.; et al. Thermophilic Composting of Human Feces: Development of Bacterial Community Composition and Antimicrobial Resistance Gene Pool. Front. Microbiol. 2022, 13, 824834. [Google Scholar] [CrossRef]
- Liang, B.; Wang, L.-Y.; Mbadinga, S.M.; Liu, J.-F.; Yang, S.-Z.; Gu, J.-D.; Mu, B.-Z. Anaerolineaceae and Methanosaeta turned to be the dominant microorganisms in alkanes-dependent methanogenic culture after long-term of incubation. AMB Express 2015, 5, 37. [Google Scholar] [CrossRef]
- Langarica-Fuentes, A.; Zafar, U.; Heyworth, A.; Brown, T.; Fox, G.; Robson, G.D. Fungal succession in an in-vessel composting system characterized using 454 pyrosequencing. FEMS Microbiol. Ecol. 2014, 88, 296–308. [Google Scholar] [CrossRef]
- Meng, Q.; Yang, W.; Men, M.; Bello, A.; Xu, X.; Xu, B.; Deng, L.; Jiang, X.; Sheng, S.; Wu, X.; et al. Microbial Community Succession and Response to Environmental Variables During Cow Manure and Corn Straw Composting. Front. Microbiol. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yin, X.; Mao, H.; Chu, C.; Tian, Y. Changes in structure and function of fungal community in cow manure composting. Bioresour. Technol. 2018, 255, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Bonito, G.; Isikhuemhen, O.S.; Vilgalys, R. Identification of fungi associated with municipal compost using DNA-based techniques. Bioresour. Technol. 2010, 101, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Hultman, J.; Vasara, T.; Partanen, P.; Kurola, J.; Kontro, M.; Paulin, L.; Auvinen, P.; Romantschuk, M. Determination of fungal succession during municipal solid waste composting using a cloning-based analysis. J. Appl. Microbiol. 2009, 108, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Lara, A.; Ros, M.; Cuartero, J.; Bustamante, M.; Moral, R.; Andreu-Rodríguez, F.J.; Fernández, J.A.; Egea-Gilabert, C.; Pascual, J.A. Bacterial and fungal community dynamics during different stages of agro-industrial waste composting and its relationship with compost suppressiveness. Sci. Total. Environ. 2021, 805, 150330. [Google Scholar] [CrossRef]
- Ryckeboer, J.; Mergaert, J.; Coosemans, J.; Deprins, K.; Swings, J. Microbiological aspects of biowaste during composting in a monitored compost bin. J. Appl. Microbiol. 2003, 94, 127–137. [Google Scholar] [CrossRef]
- Pavan, H.V.; Murthy, S.M.; Jogaiah, S. Explorations of Fungal Diversity in Extreme Environmental Conditions for Sustainable Agriculture Applications. In Biocontrol Agents and Secondary Metabolites; Jogaiah, S., Ed.; Woodhead Publishing: Sawston, UK, 2021; pp. 483–494. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G. An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Mori, A.S.; Isbell, F.; Fujii, S.; Makoto, K.; Matsuoka, S.; Osono, T. Low multifunctional redundancy of soil fungal diversity at multiple scales. Ecol. Lett. 2015, 19, 249–259. [Google Scholar] [CrossRef]
- Harpole, W.S.; Sullivan, L.L.; Lind, E.M.; Firn, J.; Adler, P.B.; Borer, E.T.; Chase, J.; Fay, P.A.; Hautier, Y.; Hillebrand, H.; et al. Addition of multiple limiting resources reduces grassland diversity. Nature 2016, 537, 93–96. [Google Scholar] [CrossRef]
- Tian, W.; Sun, Q.; Xu, D.; Zhang, Z.; Chen, D.; Li, C.; Shen, Q.; Shen, B. Succession of bacterial communities during composting process as detected by 16S rRNA clone libraries analysis. Int. Biodeterior. Biodegradation 2013, 78, 58–66. [Google Scholar] [CrossRef]
- Albrecht, R.; Périssol, C.; Ruaudel, F.; Le Petit, J.; Terrom, G. Functional changes in culturable microbial communities during a co-composting process: Carbon source utilization and co-metabolism. Waste Manag. 2010, 30, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cui, H.; Shi, J.; Zhao, X.; Zhao, Y.; Wei, Z. Relationship between bacterial diversity and environmental parameters during composting of different raw materials. Bioresour. Technol. 2015, 198, 395–402. [Google Scholar] [CrossRef]
- Oshima, T.; Moriya, T. A Preliminary Analysis of Microbial and Biochemical Properties of High-Temperature Compost. Ann. N. Y. Acad. Sci. 2008, 1125, 338–344. [Google Scholar] [CrossRef]
- Yu, Z.; Tang, J.; Liao, H.; Liu, X.; Zhou, P.; Chen, Z.; Rensing, C.; Zhou, S. The distinctive microbial community improves composting efficiency in a full-scale hyperthermophilic composting plant. Bioresour. Technol. 2018, 265, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hou, Y.; Li, Z.; Yu, Z.; Tang, J.; Wang, Y.; Zhou, S. Hyperthermophilic composting of sewage sludge accelerates humic acid formation: Elemental and spectroscopic evidence. Waste Manag. 2020, 103, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, W.; Xing, R.; Xie, S.; Yang, X.; Cui, P.; Lü, J.; Liao, H.; Yu, Z.; Wang, S.; et al. Enhanced in situ biodegradation of microplastics in sewage sludge using hyperthermophilic composting technology. J. Hazard. Mater. 2020, 384, 121271. [Google Scholar] [CrossRef] [PubMed]
- Wen, P.; Tang, J.; Wang, Y.; Liu, X.; Yu, Z.; Zhou, S. Hyperthermophilic composting significantly decreases methane emissions: Insights into the microbial mechanism. Sci. Total. Environ. 2021, 784, 147179. [Google Scholar] [CrossRef]
- Cui, P.; Chen, Z.; Zhao, Q.; Yu, Z.; Yi, Z.; Liao, H.; Zhou, S. Hyperthermophilic composting significantly decreases N2O emissions by regulating N2O-related functional genes. Bioresour. Technol. 2019, 272, 433–441. [Google Scholar] [CrossRef]
- Liao, H.; Lu, X.; Rensing, C.; Friman, V.P.; Geisen, S.; Chen, Z.; Yu, Z.; Wei, Z.; Zhou, S.; Zhu, Y. Hyperthermophilic Composting Accelerates the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements in Sewage Sludge. Environ. Sci. Technol. 2018, 52, 266–276. [Google Scholar] [CrossRef]
- Zhu, N.; Zhu, Y.; Kan, Z.; Li, B.; Cao, Y.; Jin, H. Effects of two-stage microbial inoculation on organic carbon turnover and fungal community succession during co-composting of cattle manure and rice straw. Bioresour. Technol. 2021, 341, 125842. [Google Scholar] [CrossRef]
- Mejias, L.; Komilis, D.; Gea, T.; Sánchez, A. The effect of airflow rates and aeration mode on the respiration activity of four organic wastes: Implications on the composting process. Waste Manag. 2017, 65, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Qasim, W.; Moon, B.E.; Okyere, F.G.; Khan, F.; Nafees, M.; Kim, H.T. Influence of aeration rate and reactor shape on the composting of poultry manure and sawdust. J. Air Waste Manag. Assoc. 2019, 69, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Yin, H.; Shen, X.; Liu, N.; Ge, J.; Han, L.; Huang, G. Effect of aeration interval on oxygen consumption and GHG emission during pig manure composting. Bioresour. Technol. 2018, 250, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Li, G.; Tang, Q.; Ma, X.; Wang, G.; Schuchardt, F. Effects of aeration method and aeration rate on greenhouse gas emissions during composting of pig feces in pilot scale. J. Environ. Sci. 2015, 31, 124–132. [Google Scholar] [CrossRef]
- Su, J.-Q.; Wei, B.; Ou-Yang, W.-Y.; Huang, F.-Y.; Zhao, Y.; Xu, H.-J.; Zhu, Y.-G. Antibiotic Resistome and Its Association with Bacterial Communities during Sewage Sludge Composting. Environ. Sci. Technol. 2015, 49, 7356–7363. [Google Scholar] [CrossRef]
- Onwosi, C.O.; Igbokwe, V.C.; Odimba, J.N.; Eke, I.E.; Nwankwoala, M.O.; Iroh, I.N.; Ezeogu, L.I. Composting technology in waste stabilization: On the methods, challenges and future prospects. J. Environ. Manag. 2017, 190, 140–157. [Google Scholar] [CrossRef]
- Zhang, D.; Luo, W.; Yuan, J.; Li, G.; Luo, Y. Effects of woody peat and superphosphate on compost maturity and gaseous emissions during pig manure composting. Waste Manag. 2017, 68, 56–63. [Google Scholar] [CrossRef]
- Paradelo, R.; Moldes, A.B.; Barral, M.T. Evolution of organic matter during the mesophilic composting of lignocellulosic winery wastes. J. Environ. Manag. 2013, 116, 18–26. [Google Scholar] [CrossRef]
- Yu, H.; Xie, B.; Khan, R.; Shen, G. The changes in carbon, nitrogen components and humic substances during organic-inorganic aerobic co-composting. Bioresour. Technol. 2019, 271, 228–235. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, X. Influence of bulking agents on physical, chemical, and microbiological properties during the two-stage composting of green waste. Waste Manag. 2016, 48, 115–126. [Google Scholar] [CrossRef]
- Chen, R.; Wang, Y.; Wang, W.; Wei, S.; Jing, Z.; Lin, X. N2O emissions and nitrogen transformation during windrow composting of dairy manure. J. Environ. Manag. 2015, 160, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.K.; Shafiq, T.; Ahmed, K. Characterization of bulking agents and its effects on physical properties of compost. Bioresour. Technol. 2010, 101, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Yu, K.; Ahmed, I.; Gin, K.; Xi, B.; Wei, Z.; He, Y.; Zhang, B. Key factors driving the fate of antibiotic resistance genes and controlling strategies during aerobic composting of animal manure: A review. Sci. Total. Environ. 2021, 791, 148372. [Google Scholar] [CrossRef]
- Echeverria, M.C.; Pellegrino, E.; Nuti, M. The Solid Wastes of Coffee Production and of Olive Oil Extraction: Management Perspectives in Rural Areas. In Solid Waste Management in Rural Areas; Mihai, F.C., Ed.; IntechOpen: Rijeka, Croatia, 2017; ISBN 978-953-51-3486-2. [Google Scholar] [CrossRef]
- Kim, S.M.; Dien, B.S.; Tumbleson, M.E.; Rausch, K.D.; Singh, V. Improvement of sugar yields from corn stover using sequential hot water pretreatment and disk milling. Bioresour. Technol. 2016, 216, 706–713. [Google Scholar] [CrossRef]
- Reyes-Torres, M.; Oviedo-Ocaña, E.; Dominguez, I.; Komilis, D.; Sánchez, A. A systematic review on the composting of green waste: Feedstock quality and optimization strategies. Waste Manag. 2018, 77, 486–499. [Google Scholar] [CrossRef]
- Neher, D.A.; Weicht, T.R.; Bates, S.T.; Leff, J.W.; Fierer, N. Changes in Bacterial and Fungal Communities across Compost Recipes, Preparation Methods, and Composting Times. PLoS ONE 2013, 8, e79512. [Google Scholar] [CrossRef]
- Li, Z.; Lu, H.; Ren, L.; He, L. Experimental and modeling approaches for food waste composting: A review. Chemosphere 2013, 93, 1247–1257. [Google Scholar] [CrossRef]
- Agnolucci, M.; Cristani, C.; Battini, F.; Palla, M.; Cardelli, R.; Saviozzi, A.; Nuti, M. Microbially-enhanced composting of olive mill solid waste (wet husk): Bacterial and fungal community dynamics at industrial pilot and farm level. Bioresour. Technol. 2013, 134, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, M.; Nandal, M.; Khosla, B. Microbes as vital additives for solid waste composting. Heliyon 2020, 6, e03343. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.K.; Pandey, A.K.; Khan, J.; Bundela, P.S.; Wong, J.W.; Selvam, A. Evaluation of thermophilic fungal consortium for organic municipal solid waste composting. Bioresour. Technol. 2014, 168, 214–221. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, C.; Wang, X.; Hai, L. Increased abundance of nitrogen transforming bacteria by higher C/N ratio reduces the total losses of N and C in chicken manure and corn stover mix composting. Bioresour. Technol. 2020, 297, 122410. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, M.; Cardelli, R.; Bedini, S.; Colombini, A.; Incrocci, L.; Castagna, A.; Agnolucci, M.; Cristani, C.; Ranieri, A.; Saviozzi, A.; et al. Microbially-enhanced composting of wet olive husks. Bioresour. Technol. 2012, 104, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, R.; Jouzani, G.S.; Karimi, E.; Ghanavati, H.; Kowsari, M. A bioprocess for rapid compost production from rice straw using C/N ratio improvers and indigenous microorganisms at the lab and pilot scales. Waste Biomass Valorization 2013. [Google Scholar] [CrossRef]
- Mc Carthy, G.; Lawlor, P.G.; Coffey, L.; Nolan, T.; Gutierrez, M.; Gardiner, G.E. An assessment of pathogen removal during composting of the separated solid fraction of pig manure. Bioresour. Technol. 2020, 102, 9059–9067. [Google Scholar] [CrossRef]
- Millner, P.; Ingram, D.; Mulbry, W.; Arikan, O.A. Pathogen reduction in minimally managed composting of bovine manure. Waste Manag. 2014, 34, 1992–1999. [Google Scholar] [CrossRef]
- Yoshii, T.; Moriya, T.; Oshima, T. Bacterial and Biochemical Properties of Newly Invented Aerobic, High-Temperature Compost. In Thermophilic Microbes in Environmental and Industrial Biotechnology: Biotechnology of Thermophiles; Satyanarayana, T., Littlechild, J., Kawarabayasi, Y., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 119–135. [Google Scholar] [CrossRef]
- Erickson, M.C.; Liao, J.; Boyhan, G.; Smith, C.; Ma, L.; Jiang, X.; Doyle, M.P. Fate of manure-borne pathogen surrogates in static composting piles of chicken litter and peanut hulls. Bioresour. Technol. 2010, 101, 1014–1020. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, J.; Li, J.; Xin, Y.; Hao, Z.; Chen, C.; Li, H.; Wang, B.; Ding, M.; Li, W.; et al. Insights into bacterial diversity in compost: Core microbiome and prevalence of potential pathogenic bacteria. Sci. Total. Environ. 2020, 718, 137304. [Google Scholar] [CrossRef]
- Soobhany, N.; Mohee, R.; Garg, V.K. Inactivation of bacterial pathogenic load in compost against vermicompost of organic solid waste aiming to achieve sanitation goals: A review. Waste Manag. 2017, 64, 51–62. [Google Scholar] [CrossRef]
- Storino, F.; Arizmendiarrieta, J.S.; Irigoyen, I.; Muro, J.; Aparicio-Tejo, P.M. Meat waste as feedstock for home composting: Effects on the process and quality of compost. Waste Manag. 2016, 56, 53–62. [Google Scholar] [CrossRef]
- Pandey, P.K.; Cao, W.; Biswas, S.; Vaddella, V. A new closed loop heating system for composting of green and food wastes. J. Clean. Prod. 2016, 133, 1252–1259. [Google Scholar] [CrossRef]
- Wu, H.; Lai, C.; Zeng, G.; Liang, J.; Chen, J.; Xu, J.; Dai, J.; Li, X.; Liu, J.; Chen, M.; et al. The interactions of composting and biochar and their implications for soil amendment and pollution remediation: A review. Crit. Rev. Biotechnol. 2016, 37, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, Y.; Jiang, S.; Deng, Y.; Christie, P.; Murray, P.J.; Li, X.; Zhang, J. Arbuscular mycorrhizal fungi in soil and roots respond differently to phosphorus inputs in an intensively managed calcareous agricultural soil. Sci. Rep. 2016, 6, 24902. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zeng, G.; Huang, D.; Lai, C.; Xu, P.; Zhang, C.; Cheng, M.; Wan, J.; Hu, L.; Zhang, Y. Effect of Phanerochaete chrysosporium inoculation on bacterial community and metal stabilization in lead-contaminated agricultural waste composting. Bioresour. Technol. 2017, 243, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Nakasaki, K.; Hirai, H. Temperature control strategy to enhance the activity of yeast inoculated into compost raw material for accelerated composting. Waste Manag. 2017, 65, 29–36. [Google Scholar] [CrossRef]
- Rastogi, M.; Nandal, M.; Nain, L. Seasonal variation induced stability of municipal solid waste compost: An enzyme kinetics study. SN Appl. Sci. 2019, 1, 849. [Google Scholar] [CrossRef]
- Karnchanawong, S.; Nissaikla, S. Effects of microbial inoculation on composting of household organic waste using passive aeration bin. Int. J. Recycl. Org. Waste Agric. 2014, 3, 113–119. [Google Scholar] [CrossRef]
- Perner, H.; Schwarz, D.; Bruns, C.; Mäder, P.; George, E. Effect of arbuscular mycorrhizal colonization and two levels of compost supply on nutrient uptake and flowering of pelargonium plants. Mycorrhiza 2007, 17, 469–474. [Google Scholar] [CrossRef]
- Copetta, A.; Bardi, L.; Bertolone, E.; Berta, G. Fruit production and quality of tomato plants (Solanum lycopersicum L.) are affected by green compost and arbuscular mycorrhizal fungi. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2011, 145, 106–115. [Google Scholar] [CrossRef]
- Tanwar, A.; Aggarwal, A.; Yadav, A.; Parkash, V. Screening and selection of efficient host and sugarcane bagasse as substrate for mass multiplication of Funneliformis mosseae. Biol. Agric. Hortic. 2013, 29, 107–117. [Google Scholar] [CrossRef]
- Cavagnaro, T.R. Biologically Regulated Nutrient Supply Systems: Compost and Arbuscular Mycorrhizas-A Review. Adv. Agron. 2015, 129, 293–321. [Google Scholar] [CrossRef]
- Kinet, R.; Destain, J.; Hiligsmann, S.; Thonart, P.; Delhalle, L.; Taminiau, B.; Daube, G.; Delvigne, F. Thermophilic and cellulolytic consortium isolated from composting plants improves anaerobic digestion of cellulosic biomass: Toward a microbial resource management approach. Bioresour. Technol. 2015, 189, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Manu, M.; Kumar, R.; Garg, A. Performance assessment of improved composting system for food waste with varying aeration and use of microbial inoculum. Bioresour. Technol. 2017, 234, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yang, Y.; Xia, Y.; Wu, T.; Zhu, J.; Wang, Z.; Yang, J. The succession pattern of bacterial diversity in compost using pig manure mixed with wood chips analyzed by 16S rRNA gene analysis. BioRxiv 2019, 674069. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Xia, J.; Chen, Y. Effect of microbial inoculation on physicochemical properties and bacterial community structure of citrus peel composting. Bioresour. Technol. 2019, 291, 121843. [Google Scholar] [CrossRef] [PubMed]
- Anli, M.; Symanczik, S.; El Abbassi, A.; Ait-El-Mokhtar, M.; Boutasknit, A.; Ben-Laouane, R.; Toubali, S.; Baslam, M.; Mäder, P.; Hafidi, M.; et al. Use of arbuscular mycorrhizal fungus Rhizoglomus irregulare and compost to improve growth and physiological responses of Phoenix dactylifera ‘Boufgouss’. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2020, 155, 763–771. [Google Scholar] [CrossRef]
- Jia, X.; Qin, X.; Tian, X.; Zhao, Y.; Yang, T.; Huang, J. Inoculating with the microbial agents to start up the aerobic composting of mushroom residue and wood chips at low temperature. J. Environ. Chem. Eng. 2021, 9, 105294. [Google Scholar] [CrossRef]
- Dang, Q.; Wang, Y.; Xiong, S.; Yu, H.; Zhao, X.; Tan, W.; Cui, D.; Xi, B. Untangling the response of fungal community structure, composition and function in soil aggregate fractions to food waste compost addition. Sci. Total. Environ. 2021, 769, 145248. [Google Scholar] [CrossRef]
- Guo, J.; Ling, N.; Chen, Z.; Xue, C.; Li, L.; Liu, L.; Gao, L.; Wang, M.; Ruan, J.; Guo, S.; et al. Soil fungal assemblage complexity is dependent on soil fertility and dominated by deterministic processes. New Phytol. 2020, 226, 232–243. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, N.; Ge, T.; Kuzyakov, Y.; Wang, Z.-L.; Li, Z.; Tang, Z.; Chen, Y.; Wu, C.; Lou, Y. Soil aggregation regulates distributions of carbon, microbial community and enzyme activities after 23-year manure amendment. Appl. Soil Ecol. 2016, 111, 65–72. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis; Elsevier: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Gianinazzi, S.; Gollotte, A.; Binet, M.-N.; Van Tuinen, D.; Redecker, D.; Wipf, D. Agroecology: The key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 2010, 20, 519–530. [Google Scholar] [CrossRef]
- Verbruggen, E.; Röling, W.F.M.; Gamper, H.; Kowalchuk, G.A.; Verhoef, H.A.; van der Heijden, M.G.A. Positive effects of organic farming on below-ground mutualists: Large-scale comparison of mycorrhizal fungal communities in agricultural soils. New Phytol. 2010, 186, 968–979. [Google Scholar] [CrossRef]
- Yang, W.; Gu, S.; Xin, Y.; Bello, A.; Sun, W.; Xu, X. Compost Addition Enhanced Hyphal Growth and Sporulation of Arbuscular Mycorrhizal Fungi without Affecting Their Community Composition in the Soil. Front. Microbiol. 2018, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Alguacil, M.; Torrecillas, E.; Caravaca, F.; Fernández, D.; Azcón, R.; Roldán, A. The application of an organic amendment modifies the arbuscular mycorrhizal fungal communities colonizing native seedlings grown in a heavy-metal-polluted soil. Soil Biol. Biochem. 2011, 43, 1498–1508. [Google Scholar] [CrossRef]
- Labidi, S.; Nasr, H.; Zouaghi, M.; Wallander, H. Effects of compost addition on extra-radical growth of arbuscular mycorrhizal fungi in Acacia tortilis ssp. raddiana savanna in a pre-Saharan area. Appl. Soil Ecol. 2007, 35, 184–192. [Google Scholar] [CrossRef]
- Valarini, P.J.; Curaqueo, G.; Seguel, A.; Manzano, K.; Rubio, R.; Cornejo, P.; Borie, F. Effect of Compost Application on Some Properties of a Volcanic Soil from Central South Chile. Chil. J. Agric. Res. 2009, 69, 416–425. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Dubois, D.; Ineichen, K.; Boller, T.; Wiemken, A. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 2004, 138, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Cavagnaro, T.R. Impacts of compost application on the formation and functioning of arbuscular mycorrhizas. Soil Biol. Biochem. 2014, 78, 38–44. [Google Scholar] [CrossRef]
- Wongkiew, S.; Chaikaew, P.; Takrattanasaran, N.; Khamkajorn, T. Evaluation of nutrient characteristics and bacterial community in agricultural soil groups for sustainable land management. Sci. Rep. 2022, 12, 7368. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Gao, B.; Hu, X.-K.; Lu, X.; Well, R.; Christie, P.; Bakken, L.R.; Ju, X.-T. Ammonia-oxidation as an engine to generate nitrous oxide in an intensively managed calcareous Fluvo-aquic soil. Sci. Rep. 2014, 4, 3950. [Google Scholar] [CrossRef] [PubMed]
- Viti, C.; Tatti, E.; Decorosi, F.; Lista, E.; Rea, E.; Tullio, M.; Sparvoli, E.; Giovannetti, L. Compost Effect on Plant Growth-Promoting Rhizobacteria and Mycorrhizal Fungi Population in Maize Cultivations. Compos. Sci. Util. 2010, 18, 273–281. [Google Scholar] [CrossRef]
- Strachel, R.; Wyszkowska, J.; Baćmaga, M. The Role of Compost in Stabilizing the Microbiological and Biochemical Properties of Zinc-Stressed Soil. Water Air Soil Pollut. 2017, 228, 349. [Google Scholar] [CrossRef]
- Zhen, Z.; Liu, H.; Wang, N.; Guo, L.; Meng, J.; Ding, N.; Wu, G.; Jiang, G. Effects of Manure Compost Application on Soil Microbial Community Diversity and Soil Microenvironments in a Temperate Cropland in China. PLoS ONE 2014, 9, e108555. [Google Scholar] [CrossRef]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial interactions in the rhizosphere: Beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Jian, L.; Bai, X.; Zhang, H.; Song, X.; Li, Z. Promotion of growth and metal accumulation of alfalfa by coinoculation with Sinorhizobium and Agrobacterium under copper and zinc stress. Peerj 2019, 7, e6875. [Google Scholar] [CrossRef]
- Thakur, S.; Kapila, S. Seasonal changes in antioxidant enzymes, polyphenol oxidase enzyme, flavonoids and phenolic content in three leafy liverworts. Lindbergia 2017, 5, 39–44. [Google Scholar] [CrossRef]
- Arif, M.S.; Akhtar, M.J.; Asghar, H.N.; Ahmad, R. Bio-resource efficacy of compost and PGPR inoculation in improving sunflower plant growth and soil properties. In Proceedings of the 2010 International Conference on Environmental Engineering and Applications, Singapore, 10–12 September 2010; pp. 184–187. [Google Scholar] [CrossRef]
- Ahmad, I.; Akhtar, M.J.; Mehmood, S.; Akhter, K.; Tahir, M.; Saeed, M.F.; Hussain, M.B.; Hussain, S. Combined application of compost and Bacillus sp. CIK-512 ameliorated the lead toxicity in radish by regulating the homeostasis of antioxidants and lead. Ecotoxicol. Environ. Saf. 2018, 148, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Benidire, L.; Madline, A.; Pereira, S.; Castro, P.; Boularbah, A. Synergistic effect of organo-mineral amendments and plant growth-promoting rhizobacteria (PGPR) on the establishment of vegetation cover and amelioration of mine tailings. Chemosphere 2021, 262, 127803. [Google Scholar] [CrossRef]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: A critical review of microalgae as renewable resources for agriculture. Algal Res. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Hashem, A. Problems and prospects of cyanobacterial biofertilizer for rice cultivation. Funct. Plant Biol. 2001, 28, 881–888. [Google Scholar] [CrossRef]
- Valiente, E.F.; Ucha, A.; Quesada, A.; Leganés, F.; Carreres, R. Contribution of N2 fixing cyanobacteria to rice production: Availability of nitrogen from 15N-labelled cyanobacteria and ammonium sulphate to rice. Plant Soil 2000, 221, 107–112. [Google Scholar] [CrossRef]
- Mulbry, W.; Kondrad, S.; Pizarro, C.; Kebede-Westhead, E. Treatment of dairy manure effluent using freshwater algae: Algal productivity and recovery of manure nutrients using pilot-scale algal turf scrubbers. Bioresour. Technol. 2008, 99, 8137–8142. [Google Scholar] [CrossRef]
- Liu, X.; Shi, Y.; Kong, L.; Tong, L.; Cao, H.; Zhou, H.; Lv, Y. Long-Term Application of Bio-Compost Increased Soil Microbial Community Diversity and Altered Its Composition and Network. Microorganisms 2022, 10, 462. [Google Scholar] [CrossRef]
- Anastasi, A.; Varese, G.C.; Marchisio, V.F. Isolation and identification of fungal communities in compost and vermicompost. Mycologia 2005, 97, 33–44. [Google Scholar] [CrossRef]
- Goyer, C.; Neupane, S.; Zebarth, B.J.; Burton, D.L.; Wilson, C.; Sennett, L. Diverse compost products influence soil bacterial and fungal community diversity in a potato crop production system. Appl. Soil Ecol. 2021, 169, 104247. [Google Scholar] [CrossRef]
- González-González, S.; Astorga-Eló, M.; Campos, M.; Wick, L.; Acuña, J.; Jorquera, M. Compost Fungi Allow for Effective Dispersal of Putative PGP Bacteria. Agronomy 2021, 11, 1567. [Google Scholar] [CrossRef]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant Sci. 2015, 6, 815. [Google Scholar] [CrossRef]
- Piazza, G.; Ercoli, L.; Nuti, M.; Pellegrino, E. Interaction Between Conservation Tillage and Nitrogen Fertilization Shapes Prokaryotic and Fungal Diversity at Different Soil Depths: Evidence From a 23-Year Field Experiment in the Mediterranean Area. Front. Microbiol. 2019, 10, 2047. [Google Scholar] [CrossRef]
- Douds, D.D., Jr.; Nagahashi, G.; Hepperly, P.R. On-farm production of inoculum of indigenous arbuscular mycorrhizal fungi and assessment of diluents of compost for inoculum production. Bioresour. Technol. 2010, 101, 2326–2330. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Ashton, P.D.; Aziz, N.; Feng, G.; Nelson, M.; Dytham, C.; Fitter, A.H.; Helgason, T. Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing. New Phytol. 2011, 190, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Njeru, E.M.; Avio, L.; Bocci, G.; Sbrana, C.; Turrini, A.; Bàrberi, P.; Giovannetti, M.; Oehl, F. Contrasting effects of cover crops on ‘hot spot’ arbuscular mycorrhizal fungal communities in organic tomato. Biol. Fertil. Soils 2015, 51, 151–166. [Google Scholar] [CrossRef]
- Cozzolino, V.; Di Meo, V.; Monda, H.; Spaccini, R.; Piccolo, A. The molecular characteristics of compost affect plant growth, arbuscular mycorrhizal fungi, and soil microbial community composition. Biol. Fertil. Soils 2016, 52, 15–29. [Google Scholar] [CrossRef]
- Pellegrino, E.; Nuti, M.; Ercoli, L. Multiple Arbuscular Mycorrhizal Fungal Consortia Enhance Yield and Fatty Acids of Medicago sativa: A Two-Year Field Study on Agronomic Traits and Tracing of Fungal Persistence. Front. Plant Sci. 2022, 13, 814401. [Google Scholar] [CrossRef] [PubMed]
- Favoino, E.; Hogg, D. The potential role of compost in reducing greenhouse gases. Waste Manag. Res. J. Sustain. Circ. Econ. 2008, 26, 61–69. [Google Scholar] [CrossRef] [PubMed]




| Composting Stages | Dominant Phyla | Maximum Temperatures (°C) | References |
|---|---|---|---|
| Mesophilic | Saccharomycetales Tremellales | >25 °C | [68,69] |
| Thermophilic | Eurotiales Sordariales Hypocreales Microascales | >50 °C | [65] |
| Cooling | Eurotiales Sordariales | 50–40 °C | [65] |
| Maturation | 40 °C down to environment temperature |
| Substrate | Process Time (days) | Temperature (°C) | Bacteria Pathogens | Reference |
|---|---|---|---|---|
| Chicken manure + peanut hulls | 57 | 55 | E. coli | [114] |
| Salmonella spp. | ||||
| Listeria innocua | ||||
| Bovine manure | 28 | 70 | E. coli | [112] |
| Salmonella spp. | ||||
| Meat waste | 98 | 67 | E. coli | [117] |
| Salmonella spp. | ||||
| Food waste | 70 | 60 | E. coli | [118] |
| Salmonella spp. |
| Substrate | Inoculum | Effect on the Compost Process | Reference |
|---|---|---|---|
| Peat-based substrate | Three different commercial Arbuscular mycorrhizal fungi inocula | Improve nutrient status and flower development | [125] |
| Green compost | Arbuscular mycorrhizal fungal-rhizobacterial mixed inoculum | Improve plant growth on yield and quality of fruits in tomato | [126] |
| Sugarcane bagasse substrate (fresh, dry, and compost) | Arbuscular mycorrhizal fungus, Funneliformis mosseae (sensu Glomus mosseae) | Increase in plant growth | [127] |
| Food scraps and dry leaves | Mature and commercial compost | Lower C/N ratio, higher germination rate, higher compost fraction less than 12.5 mm, and higher volatile mass reduction. | [124] |
| Municipal green waste | Arbuscular mycorrhizal fungus | Increase plant growth and/or nutrition | [128] |
| Municipal solid waste | Clostridia sp. | Improved anaerobic digestion of cellulosic biomass | [129] |
| Food waste | Lactic acid bacteria, yeasts, and phototrophic bacteria. | Accelerate the decomposition process | [130] |
| Food waste | mesophilic yeast Pichia kudriavzevii | Promote the degradation of organic matter and accelerated the composting process. | [122] |
| Pig manure and corn straw | Acinetobacter pittii, Bacillus subtilis sub sp. Stercoris y Bacillus altitudinis | Prolonging thermophilic stage, increasing germination rate | [131] |
| Municipal solid waste, sawdust, brown matter, and grasses | Bacillus subtilis, B. amyloliquefaciens, B. nakamurai y B. velezensis | Shorter time to reach the thermophilic stage, lower C/N, higher cellulose hydrolysis | [123] |
| Citrus peels (CP) | Mixing of bacteria from high temperature CP compost samples and citrus orchard soil. | Increased pile temperature, accelerates cellulose and pectin degradation | [132] |
| Cow dung and wheat straw | Bacterial consortium | Inhibited gaseous emissions and enhanced carbon and nitrogen sequestration | [36] |
| Green and industrial waste | Arbuscular mycorrhizal fungus Rhizoglomus irregulare | Improve plant growth and development | [133] |
| Mushroom and wood chip residues | Aspergillus, Penicillium, Nocardia, Arthrobacter, Sphingobacterium, Streptomyces, Novosphingobium, Flavobacterium, Pseudomonas, Cladosporium y Bacillus | Heat the compost pile and thus initiate the composting process at low temperatures. Prolong the thermophilic stage and promote the decomposition of cellulosic biomass. | [134] |
| Olive wet husks | bacteria (Bacillus subtilis, Serratia marcescens), yeasts (Rhodotorula mucillaginosa, Candida butyrii), actinobacteria (Streptomyces drozdowiczii, S. griseofulvus, S. rochei), and microfungi (Phanerochaete chrysosporium, Chaetomium globosum, Trichoderma atroviride) | Faster start and shorter maturation time of the process of composting. Deeper humification rate. Better detoxification of the starting material. | [109] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Paredes, A.; Valdés, G.; Araneda, N.; Valdebenito, E.; Hansen, F.; Nuti, M. Microbial Community in the Composting Process and Its Positive Impact on the Soil Biota in Sustainable Agriculture. Agronomy 2023, 13, 542. https://doi.org/10.3390/agronomy13020542
Aguilar-Paredes A, Valdés G, Araneda N, Valdebenito E, Hansen F, Nuti M. Microbial Community in the Composting Process and Its Positive Impact on the Soil Biota in Sustainable Agriculture. Agronomy. 2023; 13(2):542. https://doi.org/10.3390/agronomy13020542
Chicago/Turabian StyleAguilar-Paredes, Ana, Gabriela Valdés, Nicole Araneda, Emky Valdebenito, Felipe Hansen, and Marco Nuti. 2023. "Microbial Community in the Composting Process and Its Positive Impact on the Soil Biota in Sustainable Agriculture" Agronomy 13, no. 2: 542. https://doi.org/10.3390/agronomy13020542
APA StyleAguilar-Paredes, A., Valdés, G., Araneda, N., Valdebenito, E., Hansen, F., & Nuti, M. (2023). Microbial Community in the Composting Process and Its Positive Impact on the Soil Biota in Sustainable Agriculture. Agronomy, 13(2), 542. https://doi.org/10.3390/agronomy13020542