Abstract
In this study, the allelochemical-induced effects of the leaf water extract of hemp (Cannabis sativa) on seed germination and seedling growth were investigated in durum wheat (Triticum durum) cv. Mongibello and barley (Hordeum vulgare) cv. Alamo. The seeds were imbibed in 25%, 50%, 75%, or 100% solution concentrations, obtained from a 2% hemp leaf extract, and they were germinated at 20 and 10 °C. The 2% extract stock solution (i.e., 100% solution concentration) was obtained by extracting 20 g of leaf powder in 1 L distilled water (2%, w/v). A control of seeds imbibed in distilled water was considered. Germination was scored daily and at the end of the germination test, the final germination percentage (FGP), mean germination time (MGT), germination index (GI), and vigor index (VI) were calculated. The radicles and shoots of the germinated seeds of both species were also measured for their length and weight. The analyses revealed the presence of phenols in the hemp leaf extract, which were the highest in the 100% concentration (13.14 ± 0.20 mg gallic acid equivalent-GAE g−1 extract). At 20 °C, increasing the concentration had either minor (in wheat) or no effects (in barley) on the seed germination. At 10 °C, the germination was reduced by the increasing concentration, suggesting a potential allelopathic effect of the extract. However, at 100% concentration, the seeds germinated significantly better that those at 75% concentration. At 20 °C, increasing concentration progressively depressed both the radicle and shoot elongation in both species. At 10 °C, the radicle elongation was depressed by the extract, irrespective of the concentration, whilst the shoots were as long as those of the control in wheat, and smaller at the 50 and 75% concentrations in barley. We can conclude that the water extract of hemp leaves can have a negative impact on wheat and barley seed germination and seedling growth, particularly at temperatures below optimal. However, the high concentration of phenols in the extract may stimulate seed germination and seedling growth. Thus, in semi-arid areas, such as those of the Mediterranean regions, sowings of wheat and barley after hemp at an optimal (i.e., 20 °C) or nearly optimal temperature are recommended in order to minimize the combined stressing effects of the temperature and allelopathic leaf extract upon plant establishment, which may limit a successful crop establishment.
1. Introduction
Allelopathy is the process by which a plant species stimulates or inhibits the growth and development of other plant species through certain secondary metabolites (so-called allelochemicals) [1]. Allelopathic compounds may be released by plants or plant residues into the surrounding environment through leaching, volatilization, and root exudation [2], thus affecting the seed germination and seedling growth of the nearest plants, including crops. These compounds mainly exert their effects upon cell division, membrane permeability, plant hormones production, photosynthesis, respiration, and enzymatic activity [3].
Allelopathy is a property generally recognized in weeds. Lots of studies report the negative impact of weed extracts on the seed germination and seedling growth of plants, including crops [4]. In turn, allelopathy can play an important role in sustainable agriculture as a strategy for plant defense [5]. Indeed, recent agronomic research has addressed the identification of new sustainable biological methods for replacing the more impacting synthetic products in weed and pest control [6]. Plant residues left in soil might inhibit weed emergence and growth. The efficacy of the allelopathic weed control has been extensively investigated [7,8,9].
However, crop residues may exhibit seedling allelopathy in crop rotation systems, thus negatively affecting the subsequent crops. Hundreds of secondary metabolites are recognized as phytotoxic [1]. Some of them may cause inhibition in seed germination and seedling growth [10]. Drops in crop yields have often been attributed to allelopathic effects [10].
Phenolics are a group of bioactive compounds that have been recognized to be present in large amounts in the leaves of plants [9,11]. Phenolics are the most widespread class of water-soluble compounds with allelopathic effects [12]. Phenolics released in the soil by plant residues through decomposition, leaching, root exudation, etc., are reported to be one of the main causes of allelopathy for plant growth in many crops and weeds [9].
Hemp (Cannabis sativa L.) is a textile plant that, in the past, was cultivated primarily as a raw material for industrial application [13]. In recent years, its cultivation has grown extensively in Europe, and the area dedicated to this crop in the EU has increased from approximately 20,000 ha in 2015 to over 34,000 ha in 2019 (Eurostat). The recent growing interest towards this plant is also associated with its multipurpose products, as seeds for oil and food destinations [14], or secondary metabolites, such as cannabinoids and volatile terpenes for biomedical uses, from its inflorescences [15].
However, this plant contains a great number of alleochemicals [16], which may be released into the soil by leaching, through rainfall or irrigation, thus greatly affecting seed germination and seedling growth in the subsequent crop [17].
At present, the validity of introducing hemp into cropping systems of semi-arid cultivation areas in the Mediterranean regions is under consideration [18,19]. Therefore, it is necessary to investigate the potential phytotoxic effects of its residues for other crop plants in rotation.
Studies on allelopathy are generally conducted with different parts of plants, with leaves being the largest source of allelochemicals [20].
In the framework of the EU project, Marginal lands for Growing Industrial Crops (MAGIC), the aim of the present study was that to assess the allelochemical-induced effects of the leaf water extract of hemp upon seed germination and seedling growth in two important crops, durum wheat (Triticum durum) and barley (Hordeum vulgare) that are potentially in the crop rotation systems with hemp in the Mediterranean area.
Wheat and barley are among the crops that possess strong allelopathic activity on weeds or other crops [21]. However, studies on the allelopathic effects of hemp extract on these two crops are still lacking. The two species have been selected according to their wide diffusion and economic importance in the Mediterranean regions.
2. Materials and Methods
2.1. Plant Material
Fresh, fully expanded leaves of hemp cv. Futura 75, were collected randomly from plants at the flowering stage (late July). The hemp plants were those of an open field experiment conducted during the summer season of 2018 in a flat site of the eastern coast of Sicily (South Italy, 10 m a.s.l., 37°24′35.8″ N Lat, 15°03′31.7″ E Long).
2.2. Leaf Extract
An adaptation of the method proposed by Khan et al. [6] was applied in the leaf extraction. The hemp leaves (approx. 100 g) were dried at room temperature (approx. 20 °C ± 1 °C) for 2 weeks, then they were grinded finely using an electric grinder (A11 Basic, IKA Works, Willmington, NC, USA) and sieved through a 1 mm sieve. Twenty grams of leaf powder was mixed with 1 L distilled water (2%, w/v) at room temperature and stirred for 24 h in the dark. After that, the aqueous solution was filtered through a 0.45 μm paper filter and used as a stock solution. The 2% aqueous extract stock solution (20 g L−1, 100% concentration) was diluted in distilled water to prepare three further solutions, having one of the following concentrations: 25%, 50%, 75% (respectively, 5, 10, 15 g L−1 of aqueous hemp leaf extract). These were stored at −20 °C until use. The distilled water was used as the control (0% extract).
2.3. Germination Tests
Seeds of durum wheat cv. Mongibello, and barley cv. Alamo were used for the germination test. The seeds of the two species were produced in 2018 at the experimental field of the University of Catania (Catania, Italy). The mean 100-seed weight was 5.89 ± 0.08 g, in the wheat and 5.24 ± 0.06 in the barley.
Thirty seeds of each species were placed in 90 mm Petri dishes containing a single paper sheet and moistened with 7 mL of one of the above-mentioned solutions from the hemp leaves extract. The Petri dishes were sealed with Parafilm to prevent water losses; therefore, no solution replenishment was required during the germination test. The Petri dishes were incubated in a thermostatically controlled incubator, in the dark, in a completely randomized experimental design with four replicates. The seeds were germinated at two constant temperatures: 20 °C (optimal, control) (ISTA, International rules for seed testing, 1996) and 10 °C (suboptimal). Seed germination was scored daily (radicle at least 2 mm long) until no further visible germination occurred for 72 h. At the end of the tests, the final percentage germination (FGP, %), mean germination time (MGT, days), germination index (GI), and seed vigor index (VI) were calculated. For the calculations, the following formulas were used [22]:
where FGP is the final germination (%), Σn is total number of seeds germinated, Tn is total number of seeds left to germinate, MGT is mean germination time (days), n is the number of seeds germinated on day d, GI is germination index, D is the number of days from the first to the last seed germination, VI is vigor index, and L is the length of shoot (mm) from the same seed lot.
FGP = (Σn/Tn) × 100
MGT = Σdn/Σn
GI = Σ(n/d)/D
VI = FGP × L
2.4. Radicle and Shoot Measurements
The radicles and shoots of the germinated seeds of both species were measured for their length and weight. To this end, five representative seedlings per each Petri dish were chosen among those germinated at the first stage that displayed a normal radicle growth. The radicles and shoots were excised from the seedlings 6 days after the start of the germination test and measured for their length (mm), in a caliper measure tool (Vernier caliper, 503 Series, Roder srl, Oglianico, Italy), and for their fresh weight (FW) (mg); they were oven-dried at 65 °C for 48 h for the dry weight (DW) (mg) measurement.
2.5. Total Phenolic Content Measurement on Leaf Extract
The total phenolic content (TPC) was estimated for the leaf extracts solutions using the Folin-Ciocalteau reagent [23]. The extract aliquots (125 μL) were mixed with 125 μL of the Folin-Ciocalteau reagent (Sigma-Aldrich, Milan, Italy) and, 10′ later, 1.25 mL of Na2CO3 (7%, w/v) (Sigma-Aldrich, Milan, Italy) was added. The final mixture was diluted to 3 mL with distilled water and incubated at 20 °C for 90 min in the dark.
The absorbance was read at 760 nm in a spectrophotometer (UV-30 Scan, Onda Spectrophotometer, Carpi, Italy). The measurements were compared to a gallic acid calibration curve (0, 20, 50, 75, 100 μg mL−1) (R2 = 0.9937). The results were expressed as mg gallic acid equivalent (GAE) g−1 extract DW. The analyses were conducted in triplicate.
2.6. Data Analysis
The time course of the cumulative values of the seed germination was described by a nonlinear iterative regression method (SIGMAPLOT® 11.0 software; Systat Software Inc., San Jose, CA, USA) using the following sigmoidal logistic equation with three parameters:
where a is the maximal value of y (i.e., maximum germination), x is the time (days) starting from seed imbibition, x0 is the time (days) to reach 50% of maximal germination, b is a fitting parameter of the curve. The x value on the curve corresponding to 50% of cumulative germination on the y axis was assumed to be the theoretical median germination time (t50) or theoretical time to 50% germination [24].
y = a/(1 + (x/x0)b)
The data of the total phenolic content (TPC) were statistically analyzed by a one-way ANOVA. The means were separated for significance at p < 0.05 level according to the Tukey’s test.
The data of the final germination percentage, previously arcsine transformed, those of the MGT, and those of the radicles and shoots length, fresh and dry weight, were statistically analyzed by a two-way ANOVA, separately within each species, considering ‘extract concentration’ and ‘germination temperature’ as the fixed factors. The main effects and interactions were evaluated for significance at p < 0.05 level according to the Tukey’s test.
3. Results
3.1. Total Phenolic Content (TPC)
The total phenolic content (TPC) was the highest in the 20 g L−1 extract (i.e., 100% concentration) (13.14 ± 0.20 mg GAE g−1 extract) and the lowest in the 5.0 g L−1 extract (i.e., 25% concentration) (3.81 ± 0.04 mg GAE) (Table 1). The TPC proportionally increased with the increase in the extract concentration (R2 = 0.99). No phenols were detected in the distilled water (control).

Table 1.
Total phenolic content (TPC) in different solutions obtained from 2% (w/v) hemp leaves aqueous extract.
3.2. Germination Course
The course of cumulative seed germination for the two species during imbibition at 20 and 10 °C, under different extract concentrations of the hemp leaves, is illustrated in Figure 1. This course is well described (R2 ≥ 0.97) by a sigmoidal logistic equation, the trend of which indicates an initial phase of low germination, followed by a step rise in germination, up to a maximum (a parameter of the curve) (Table 2 and Table 3).
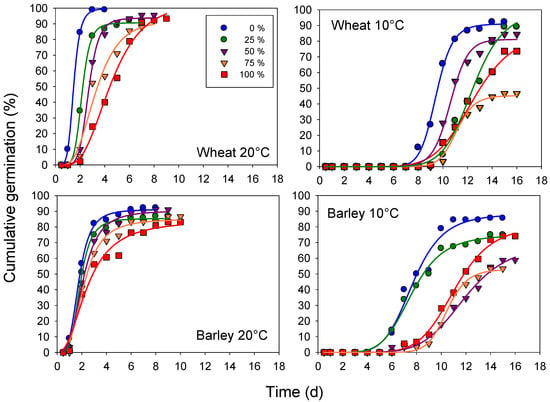
Figure 1.
Time course (solid curves) of cumulative seed germination at different hemp leaf extract concentrations of the imbibition solution at 20 and 10 °C, in wheat cv. Mongibello and barley cv. Alamo. Symbols represent the observed percentage with time.
Table 2.
Values of parameters of the sigmoidal logistic function interpolating data of seed germination at 20 and 10 °C in wheat cv. Mongibello under different hemp leaf extract concentrations.
Table 3.
Values of parameters of the sigmoidal logistic function interpolating data of seed germination at 20 and 10 °C in barley cv. Alamo under different hemp leaf extract concentrations.
The length of the initial phase of germination differed between the species, being longer in the seeds of wheat, and with the temperatures being negligible or null in the optimal thermal conditions. The thermal lowering from 20 to 10 °C slowed down the overall course of seed germination in both species.
The extract concentration of the imbibition solution affected both the start and the speed of seed germination. Increasing the concentration adversely affected germination, which was delayed and reduced to a maximum extent at the highest concentration (100%), at least in the seeds of barley germinated at 20 °C. In fact, in the wheat, the increase in the extract concentration at 20 °C only slowed down seed germination, which approached similar final values (a parameter of the curve) at all of the concentrations (when excluded under distilled water). In addition, interestingly, at the suboptimal temperature (10 °C), germination at the 100% concentration started earlier and was greater than that at the 75% concentration in wheat, and at 50 and 75% concentrations in barley.
3.3. FGP, MGT, t50, GI
The final germination percentage (FGP) was affected by the temperature, extract concentration, and their interaction in both species (Table 4 and Table 5).
Table 4.
Main effects of temperature and hemp leaf extract concentrations on final germination percentage (FGP), Mean germination time (MGT), and time to 50% germination (t50), in wheat cv. Mongibello.
Table 5.
Main effects of temperature and hemp leaf extract concentrations on final germination percentage (FGP), Mean germination time (MGT), and time to 50% germination (t50), in barley cv. Alamo.
Across the concentrations of the imbibition solution, the lowering of the temperature from 20 to 10 °C significantly altered the seed germination in both species (T, p < 0.001), which was reduced to a greater extent in the barley (−21% FGP) than the wheat (−17% FGP). Across the temperatures, a significant decline in the seed germination, from 95.6 and 88.9% (in wheat and barley, respectively) down to 70% in both species, was recorded as the extract concentration increased from 0% (distilled water) to 75%. After that, a further increase in the concentration to 100% led to a new rise in the seed germination, to 83.1% (in wheat) and 78.7% (in barley).
However, a significant C × T interaction (p < 0.05) in both species reveals a different effect of the extract concentration on seed germination at the two temperatures. Indeed, at the optimal temperature (20 °C), the FGP either did not change much (in wheat) or did not change at all (in barley) with the concentration of the imbibition solution, always exceeding 92% in wheat and 83% in barley (Figure 2). In contrast, at 10 °C, a clear effect of the imbibition solution was observed, and the seeds at the highest concentration (100%) germinated significantly better than those at the 75% concentration in both species.
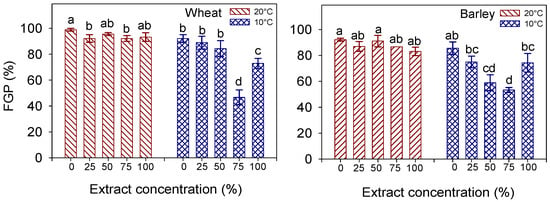
Figure 2.
Interaction effect of ‘temperature × extract concentration’ on final seed germination percentage (FGP, %) in wheat cv. Mongibello and barley cv. Alamo. Data are means ± se (n = 4). Different letters indicate significance at p < 0.05 according to Tukey’ test.
With the exception of a few cases, the values of t50, i.e., those of 50% seed germination on the interpolation curve, match with good approximation to those of the calculated MGT. Overall, germination proceeded faster under the optimal thermal conditions (20 °C), taking approximately three days (MGT) to be completed. Across the extract concentrations, the lowering of the temperature to 10 °C extended the germination time to 11 and 10 days, respectively, in wheat and barley. The significant interaction T × C (p < 0.001) in barley reveals that, in contrast to what occurred at 20 °C, where the effect of the increasing extract concentration on the germination speed was minor, at 10 °C, the seeds took longer (MGT approx. 11 days) to complete germination at concentrations ≥ 50% (Figure 3).
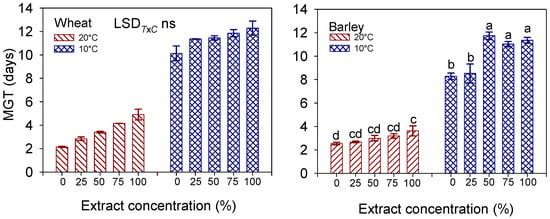
Figure 3.
Interaction effect of ‘temperature × extract concentration’ on mean germination time (MGT, days) in wheat cv. Mongibello and barley cv. Alamo. Data are means ± se (n = 4). Different letters in barley indicate significance at p < 0.05 according to Tukey’ test (T × C wheat, not significant-ns).
3.4. Radicle and Shoot Traits
In both species, the thermal lowering from 20 °C to 10 °C adversely affected the radicle and shoot elongation (T, p < 0.001) (Table 6 and Table 7). The depressive effect exerted by the low temperature was more evident in the shoot, which was 81% and 77% shorter at 10 °C in wheat and barley, respectively. Increasing the concentration of the extract also resulted in a significant reduction in the radicle and shoot length of both species and across temperatures (C, p < 0.001). In contrast to what occurred in response to the temperature, the shoots exhibited a lower sensitivity than the radicles to the increasing concentration of the hemp extract. Indeed, an increase in the concentration from 0 to 100% led to a −62% and −40% reduction in the shoot length, respectively, in wheat and barley, against the −79% and −62% decrease that occurred in the radicle length. However, the significant interaction T × C (p < 0.01) reveals a different effect of the extract concentration, depending on temperature (Figure 4). Indeed, at the optimal temperature (20 °C), an increase in the concentration progressively depressed the radicle elongation in both species. In contrast, at the suboptimal temperature (10 °C), the adverse effect of the hemp extract on this trait did not change with the concentration (excluding the control). As for the radicle, at 20 °C, the shoot elongation was progressively inhibited as the extract concentration increased. However, at 10 °C, the shoots at all concentrations were as long as those of the control in wheat (Figure 4C), whilst an inhibiting effect of the hemp extract was clear at the 50 and 75% (but not at 100%) concentrations in barley (Figure 4D).
Table 6.
Main effects of temperature and hemp leaf extract concentrations on radicle and shoot length, fresh (FW) and dry weight (DW), in wheat cv. Mongibello.
Table 7.
Main effects of temperature and hemp leaf extract concentrations on radicle and shoot length, fresh (FW) and dry weight (DW), in barley cv. Alamo.
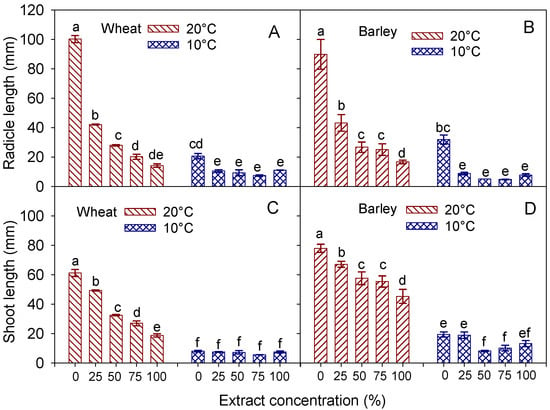
Figure 4.
Interaction effect of ‘temperature × extract concentration’ on the length of radicle and shoot, in wheat cv. Mongibello (A,C) and barley cv. Alamo (B,D). Data are means ± se (n = 4). Different letters in barley indicate significance at p < 0.05 according to Tukey’ test.
The germination temperature also affected the radicles and shoots in their fresh and dry weight (T, p < 0.001). As observed for length, in both species, the shoots were more sensitive than the radicles to thermal lowering from 20 to 10 °C, reducing their FW and DW to a greater extent. As for length, the effects of a progressive increase in the extract concentration were stronger on the radicles than on the shoots. Indeed, the FW and DW of the radicles were reduced, respectively, by 73 and 62% in wheat, and by the 68 and 58% in barley, while the FW and DW of the shoots were reduced by 64 and 54% in wheat, and by the 40 and 34% in barley, when the extract concentration of the imbibition solution was increased from 0 to 100%.
However, for these traits, the significant T × C interaction (0.05 > p < 0.001) revealed a different response to the extract concentration at the two temperatures.
In wheat, at 20 °C, both the FW and DW of the radicles progressively decreased with the rise in the concentration from 0 to 100%; at 10 °C, they decreased down to a minimum at 75% concentration, to rise again (at 100% concentration) to values similar to those of the control (distilled water) (Figure 5A,C).
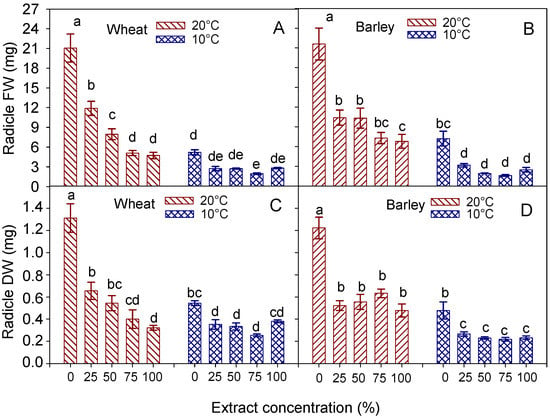
Figure 5.
Interaction effect of ‘temperature × extract concentration’ on fresh (FW) and dry (DW) weight of radicle, in wheat cv. Mongibello (A,C) and barley cv. Alamo (B,D). Data are means ± se (n = 4). Different letters in barley indicate significance at p < 0.05 according to Tukey’ test.
In barley, at both temperatures, the radicle FW and DW were reduced by the hemp extract at all concentrations. However, both weights did not change, or slightly decreased (FW at 20 °C), with the rise in the concentration of the extract from 25 to 100% (Figure 5B,D).
The drop of the FW and DW in response to the increasing extract concentration was less evident in the shoots, particularly in barley, whose shoot weight (both fresh and dry) was significantly reduced only at the highest concentrations (75 and 100%) at 20 °C, while no effect of the concentration was observed at 10 °C (Figure 6B,D). In addition, no effect of the extract concentration on the shoot weight was detected in wheat at the lowest temperature (Figure 6A,C).
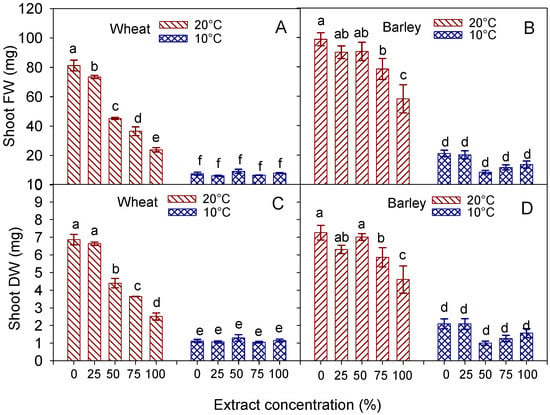
Figure 6.
Interaction effect of ‘temperature × extract concentration’ on fresh (FW) and dry (DW) weight of shoot, in wheat cv. Mongibello (A,C) and barley cv. Alamo (B,D). Data are means ± se (n = 4). Different letters in barley indicate significance at p < 0.05 according to Tukey’ test.
3.5. Germination Index (GI) and Vigor Index (VI)
The germination index (GI), calculated from the number of seeds germinated per each day and relative day of germination, changed with the temperature and extract concentration. The GI was reduced by 87% in both species, moving the seeds from the optimal to suboptimal temperature (Table 8 and Table 9). The GI was progressively reduced by increasing the concentration of the extract in wheat. At both temperatures, the effect of the extract did not change with the concentration (when excluding the control) (T × C, ns) in barley. Instead, the two factors significantly interacted in the wheat, where the effects of increasing the concentration of the hemp extract was significant at 20 °C but not at 10 °C.
Table 8.
Germination index (GI) and vigor index (VI) at two temperatures (20 and 10 °C), in wheat cv. Mongibello in relation to the hemp leaf extract concentrations. For the main effects (T = temperature, C = concentration), means followed by the same letter are not significantly different at p < 0.05 (Tukey’s test). LSD interaction value is also reported.
Table 9.
Germination index (GI) and vigor index (VI) at two temperatures (20 and 10 °C), in barley cv. Alamo in relation to the hemp leaf extract concentrations. For the main effects (T = temperature, C = concentration), means followed by the same letter are not significantly different at p < 0.05 (Tukey’s test). LSD interaction value is also reported (ns = not significant).
The vigor index (VI), calculated from the values of the FGP and length of the shoot, was influenced by the germination temperature and the extract concentration in both species. As expected, the VI was negatively affected (−84%, in wheat, −81%, in barley) by the thermal lowering from 20 to 10 °C (Table 7 and Table 8). Increasing the concentration of the hemp extract significantly reduced the VI, whilst the depressive effect progressively increased with the rise in the concentration up to 75%, with no further effects at 100% in wheat; no differences of the extract on the VI were ascertained at concentrations from 50% onward in barley. The significant T × C on the VI was highlighted in the ANOVA, which reveals how the extract concentration differed in its effect depending on temperature, affecting the VI to a greater extent at 20 °C in both species. Interestingly, at a low temperature, the VI at the maximum concentration (100%) did not differ from that of the control.
4. Discussion
When the seeds were germinated in distilled water (control), the lowering of the temperature from optimal to suboptimal had a minor effect on seed germination. Indeed, as microthermal species, the low thermal requirements for germination allow both plants to be preferably cultivated in the winter, at least in Mediterranean semi-arid regions, under rainfed conditions [25]. The thermal decrease only delayed the onset of germination; however, once this started, it proceeded more or less as at optimal temperature. Guo et al. [26] suggested that the occurrence of a secondary dormancy in seeds, when they are exposed to temperatures lower than optimum, as a mechanism of adaptation to stressful conditions.
In addition to temperature, the concentration of the extract affected all of the germination traits in both species. The aqueous extract of hemp leaves at different concentrations had varying levels of inhibition on seed germination in terms of the final percentage and speed. The inhibition was, overall, minimal at low concentrations in both species. Low inhibitory effects of the extract of Cassia tora at low concentrations were also reported for seed germination in mustard [11].
However, the interaction between the two experimental factors (‘germination temperature’ and ‘extract concentration’) upon most of the germination traits revealed that the effect of the extract at the two temperatures was not the same. At the optimal temperature, in both species, the extract affected the germination speed but, not its final value, which was the same at all concentrations. Indeed, often, no allelopathic effects have been reported to occur on the final seed germination percentage, but have been reported to occur on the germination speed, thus affecting seedling establishment under natural conditions [27]. No significant effects of the hemp extract concentration were reported by Pudelko et al. [17] on the germination of wheat at the optimal temperature.
In this study, at suboptimal temperature, the inhibiting effect of the extract upon the germination percentage was clear at the highest concentrations, suggesting a potential allelopathy of the hemp plant [3].
Although it is commonly believed that allelopathy is not related to a single compound, but rather to a set of allelochemicals [9,28], phenolics have been also recognized as one of the main potential allelopathic compounds in the ecosystem [6].
The occurrence of phenolic compounds in hemp extract, as measured in the present study, may be the reason for the low germination performances observed in this experiment in the seeds of both wheat and barley when left to germinate in hemp leaf extract, particularly at high concentrations and at a low temperature. Earlier studies reported that phenolics are potent germination and growth inhibitors [4]. Germination inhibition was also reported in the seeds of three cultivars of wheat when germinated in an aqueous extract of Eucalyptus [10]. The authors suggested that phenolics were the main allelochemicals responsible for the inhibitory effects on seed germination. The same authors also measured increased peroxidase activity in wheat radicles, which denoted a detoxification process in response to stress conditions.
Some authors have suggested that the lowering in seed germination at high concentrations of aqueous extracts from leaves occurs as a result of reduced seed water absorption due to the presence of allelochemicals in the imbibition substrate [29]. Other authors have indicated that an alteration in the gibberellic activity that, as well known, regulates the amylase production during germination, may occur in the presence of allelochemicals [30]. Similarly, Knox et al. [31] proposed a possible phenolic-related inhibition of gibberellin activity, which regulates seed germination. However, we did not measure the amylase activity nor the sugar content in the seedlings.
The allelopathic effects of the leaf extract were also evident on the growth of the radicles and shoots in both species, and as for germination, this effect was concentration-dependent. Gulzar and Siddiqui [3] found that 8% (w/v) aqueous extract of Calotropis procera had effects on the germination traits and seedling growth of Brassica oleracea only at the highest concentrations (up to 100%), corroborating the results from the present study concerning the dose-dependent effect of aqueous leaf extract.
The radicle shoot elongation, more than shoot elongation, was affected by the increase in the extract concentration. A similar greater sensitivity of root than shoot elongation to increasing concentrations of aqueous leaf extract, in this case from leaves of Mikania micrantha, has been reported by Sahu and Devkota [12] in Raphanus sativus and Oryza sativa. Nishida et al. [32] reported that root growth is more susceptible to allelochemicals mainly because of the greater root tissue permeability than shoot tissue to these compounds.
Khan et al. [6] suggested an inhibitory effect of phenolics upon the mitotic activity of young cells, thus inhibiting root elongation and growth. In this regard, seedling growth, and root in particular, may be considered as a good indicator of plant sensitivity to allelopathy [33].
In the present experiment, the fresh weight decrease in both the radicles and shoots under increasing leaf extract concentrations was greater than that of the dry weight in both species. Ziaebrahimi et al. [10] attributed the greater reduction in the fresh weight to an imbalanced water uptake or a balanced osmosis in tissues due to allelochemical toxicity.
Slight differences were ascertained between the two species in response to the increasing concentration of the aqueous extract, with barley exhibiting less susceptibility to the presence of allelochemicals than wheat. Differences in the sensitivity to allelopathy were also reported between Raphanus sativus and Oryza sativa, with the former being more susceptible than the latter [12]. More resistant mechanisms have been evolved in some plants, which may support these different behaviors [10].
Interestingly, at the highest concentration (100%), the effect of the hemp leaf extract seemed to be less depressive upon seed germination and seedling growth, but somehow it was even stimulatory when compared to the lowest concentrations, particularly at the suboptimal temperature. This result was also supported by the values of the vigor index for the 100% extract at 10 °C in both species, which was higher than the values at lower concentrations. Vigor has been reported as an index of the physiological quality of the seed, the measurement of which makes it possible to evaluate the influence of the factors studied [34].
Thus, an inhibiting/stimulating effect was exerted by the leaf extract, which was concentration-dependent. Indeed, both stimulatory and inhibitory effects of allelochemicals have been reported in the literature [35]. Phenolics can stimulate seedling growth by increasing the level of phytormones, such as auxins, cytokinins, gibberellins, which are recognized as growth-promoters [36]. Authors have reported a stimulatory effect of the aqueous extract from plants of mugwort on seed germination in some vegetable crops. This effect, however, was not followed by the same stimulatory effects on seedling growth, which was instead reduced with the increasing extract concentration [37]. Similarly, an inhibitory effect of the aqueous extract of mugwort at a 25% (w/v) concentration on seed germination and plant growth was reported in Lolium multiflorum, but not in Triticum aestivum [35]. We may speculate that hemp leaf extract at a concentration of stock solution greater than that of the present study (2%, w/v) could have even greater stimulatory effects on the seed germination and seedling growth of durum wheat and barley, particularly at a low temperature.
In this regard, for each leaf plant extract, it is important to define the range of concentrations that inhibit a single process and that of the concentrations that, in turn, may stimulate the same process in plants [38].
5. Conclusions
Based on the results of this study, we can conclude that the phenols in the water extract of hemp leaves can have a negative impact on wheat and barley seed germination and seedling growth, particularly at temperatures below the optimal.
We may suppose that in humid and semi-humid areas of cultivation, abundant and frequent rain, through their leaching action, may allow the allelochemicals of hemp to be present in the soil at concentrations lower than those that have phytotoxic effects.
In contrast, in semi-arid areas, such as those in the Mediterranean regions, sowings of wheat and barley at optimal or nearly optimal temperature are recommended in order to minimize the combined stressing effects of temperature and allelopathic leaf extract upon plant establishment, which may limit a successful crop establishment after the presence of hemp in cropping systems.
In turn, taking into consideration the stimulatory effect of the hemp extract found at the highest concentration on seed germination and seedling growth, a long-term accumulation of these allelochemicals in the soil might be even beneficial for both species when late sowings are adopted. In this regard, further studies are required to explore the potential stimulatory effect of hemp leaf extract at higher concentrations for possible application as a biostimulant in agriculture.
Author Contributions
Conceptualization, C.P. and G.T.; methodology, C.P.; software, C.P.; validation, S.L.C.; formal analysis, C.P. and G.T.; investigation, C.P. and A.P.; resources, S.L.C. and G.T.; data curation, C.P.; writing—original draft preparation, C.P.; writing—review and editing, S.L.C.; visualization, C.P. and G.T.; supervision, S.L.C.; project administration, S.L.C. and G.T.; funding acquisition, S.L.C. and G.T. All authors have read and agreed to the published version of the manuscript.
Funding
This paper is part of a project that has received funding from the “European Union’s Horizon 2020 research and innovation program under grant agreement No 727698”.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Einhellig, F. Interactions involving allelopathy in cropping systems. Agron. J. 1996, 893, 886–893. [Google Scholar] [CrossRef]
- Sangeetha, C.; Baskar, P. Allelopathy in weed management: A critical review. Afr. J. Agric Res. 2015, 10, 1004–1015. [Google Scholar]
- Gulzar, A.; Siddiqui, M.B. Allelopathic effect of Calotropis procera (Ait.) R. Br. on growth and antioxidant activity of Brassica oleracea var. botrytis. J. Saudi Soc. Agric. Sci. 2017, 16, 375–382. [Google Scholar] [CrossRef]
- Madany, M.M.Y.; Saleh, A.M. Phytotoxicity of Euphorbia helioscopia L. on Triticum aestivum L. and Pisum sativum L. Annals Agric. Sci. 2015, 60, 141–151. [Google Scholar] [CrossRef]
- Kohli, R.K.; Batish, D.; Singh, H.P. Allelopathy and its implications in Agroecosystems. J. Crop. Prod. 1998, 1, 169–202. [Google Scholar] [CrossRef]
- Khan, A.U.; Ullah, F.; Mehmood, S.; Irshad, M.; Khan, F.U. Allelopathic effects of Jatropha curcas L. leaf aqueous extract on early seedling growth of Parthenium hysterophorus. Pakistan J. Agric. Res. 2017, 30, 45–54. [Google Scholar]
- Golisz, A.; Lata, B.; Gawronski, S.W.; Fujii, Y. Specific and total activities of the allelochemicals identified in buckwheat. Weed Biol. Manag. 2007, 7, 164–171. [Google Scholar] [CrossRef]
- Kalinova, J.; Vrchotova, N.; Triska, J. Exudation of allelopathic substances in buckwheat (Fagopyrum esculentum Moench.). J. Agric. Food Chem. 2007, 55, 6453–6459. [Google Scholar] [CrossRef]
- Szwed, M.; Mitrus, J.; Wiczkowski, W.; Dębski, H.; Horbowicz, M. If phenolic compounds in the soil with buckwheat residues affect the emergence and growth of weed seedlings? Acta Physiol. Plant. 2020, 42, 154. [Google Scholar] [CrossRef]
- Ziaebrahimi, L.; Khavari-Nejad, R.A.; Fahimi, H.; Nejadsatari, T. Effects of aqueous eucalyptus extracts on seed germination, seedling growth and activities of peroxidase and polyphenoloxidase in three wheat cultivar seedlings (Triticum aestivum L.). Pak. J. Biol. Sci. 2007, 10, 3415–3419. [Google Scholar] [CrossRef]
- Sarkar, E.; Chatterjee, S.N.; Chakraborty, P. Allelopathic effect of Cassia tora on seed germination and growth of mustard. Turk. J. Bot. 2012, 36, 488–494. [Google Scholar] [CrossRef]
- Sahu, A.; Devkota, A. Allelopathic effects of aqueous extract of leaves of Mikania micrantha H.B.K. on seed germination and seedling growth of Oryza sativa l. and Raphanus sativus L. Sci. World 2013, 11, 90–93. [Google Scholar] [CrossRef]
- Leoni, M.; Musio, S.; Croci, M.; Tang, K.; Magagnini, G.M.; Thouminot, C.; Müssig, J.; Amaducci, S. The effect of agronomic management of hemp (Cannabis sativa L.) on stem processing and fibre quality. Ind. Crops Prod. 2022, 188, 115520. [Google Scholar] [CrossRef]
- Höppner, F.; Menge-Hartmann, U. Yield and quality of fibre and oil of fourteen hemp cultivars in Northern Germany at two harvest dates. Landbauforsch. Volkenrode 2007, 57, 219–232. [Google Scholar]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Santini, G.; Fiorini, D.; Sutf, S.; Dall’Acqua, S.; Canale, A.; Maggi, F. The essential oil from industrial hemp (Cannabis sativa L.) by-products as an effective tool for insect pest management in organic crops. Ind. Crops Prod. 2018, 122, 308–315. [Google Scholar] [CrossRef]
- Mahmoodzadeh, H.; Ghasem, M.; Zanganeh, H. Allelopathic effect of medicinal plant Cannabis sativa L. on Lactuca sativa L. seed germination. Acta Agric. Slov. 2015, 105, 233–239. [Google Scholar] [CrossRef]
- Pudełko, K.; Majchrzak, L.; Narożna, D. Allelopathic effect of fibre hemp (Cannabis sativa L.) on monocot and dicot plant species. Ind. Crops Prod. 2014, 56, 191–199. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Testa, G.; Scordia, D.; Copani, V. Sowing time and prediction of flowering of different hemp (Cannabis sativa L.) genotypes in southern Europe. Ind. Crops Prod. 2012, 37, 20–33. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Riggi, E.; Testa, G.; Scordia, D.; Copani, V. Evaluation of European developed fibre hemp genotypes (Cannabis sativa L.) in semi-arid Mediterranean environment. Ind. Crops Prod. 2013, 50, 312–324. [Google Scholar] [CrossRef]
- Tanveer, A.; Rehman, A.; Javaid, M.M.; Abbas, R.N.; Sibtain, M.; Ahmad, A.U.; Ibin-I-Zamir, M.S.; Chaudhary, K.M.; Aziz, A. Allelopathic potential of Euphorbia helioscopia L. against wheat (Triticum aestivum L.), chickpea (Cicer arietinum L.) and lentil (Lens culinaris Medic.). Turk. J. Agric. Forest. 2010, 34, 75–81. [Google Scholar] [CrossRef]
- Jabran, K. Manipulation of allelopathic crops for weed control. In SpringerBriefs in Plant Science; Springer: Berlin/Heidelberg, Germany, 2017; p. 87. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor Determination in Soybean Seed by Multiple Criteria. Crop. Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Patanè, C.; Cavallaro, V.; Cosentino, S.L. Germination and radicle growth in unprimed and primed seeds of sweet sorghum as affected by reduced water potential in NaCl at different temperatures. Ind. Crops Prod. 2009, 30, 1–8. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Sanzone, E.; Testa, G.; Patanè, C.; Anastasi, U.; Scordia, D. Does post-anthesis heat stress affect plant phenology, physiology, grain yield and protein content of durum wheat in a semi-arid Mediterranean environment? J. Agron. Crop. Sci. 2019, 205, 309–323. [Google Scholar] [CrossRef]
- Guo, C.; Shen, Y.; Shi, F. Effect of temperature, light, and storage time on the seed germination of Pinus bungeana Zucc. ex Endl.: The role of seed-covering layers and abscisic acid changes. Forests 2020, 11, 300. [Google Scholar] [CrossRef]
- Chaves, N.; Sosa, T.; Escudero, J.C. Plant growth inhibiting fl avonoids in exudate of Cistus ladanifer and in associated soils. J. Chem. Ecol. 2001, 27, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Tawaha, A.M.; Turk, M.A. Allelopathic effects of black musard (Brassica nigra) on germination and growth of wild barley (Hordeum spontaneum). J. Agron. Crop. Sci. 2003, 189, 298–303. [Google Scholar] [CrossRef]
- Chandler, P.M.; Zucar, J.A.; Jacobson, J.V.; Higgins, T.J.V.; Inglis, A.S. The effect of gibberellic acid and abscisic acid on α-amylase mRNA levels in barley aleurone layers studies using an α-amylase c DNA clone. Plant Mol. Biol. 1984, 3, 407–408. [Google Scholar] [CrossRef]
- Knox, J.A.I.; Jaggi, D.; Paul, M.S. Allelopathic effect of selected weeds on biochemical activity of Parthenium hysterophorus. Curr. Res. J. Biol. Sci. 2014, 2, 238–240. [Google Scholar]
- Nishida, N.; Tamotsu, S.; Negata, N.; Saito, C.; Sakai, A. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. J. Chem. Ecol. 2005, 31, 1187–1203. [Google Scholar] [CrossRef] [PubMed]
- Mondal, F.M.; Asaduzzaman, M.; Asao, T. Adverse effects of allelopathy from legume crops and its possible avoidance. Am. J. Plant Sci. 2015, 6, 804–810. [Google Scholar] [CrossRef]
- Troyjack, C.; Pimentel, J.; Dubai Padilha, I.; Veliz Escalera, R.; Acosta Jaque, L.; Koch, F.; Monteiro, M.; Demari, G.; Szareski, V.; Carvalho, I.; et al. Nitrogen fertilization on maize sowing: Plant growth and seed vigor. Am. J. Plant Sci. 2018, 9, 83–97. [Google Scholar] [CrossRef]
- Foy, C.L.; Inderjit. Understanding the role of allelopathy in weed interference and declining plant diversity. Weed Technol. 2001, 15, 837–878. [Google Scholar] [CrossRef]
- Abdalla, M. The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. Sativa) plants. Int. J. Plant Physiol. Biochem. 2013, 5, 42–49. [Google Scholar]
- Pannacci, E.; Baratta, S.; Falcinelli, B.; Farneselli, M.; Tei, F. Mugwort (Artemisia vulgaris L.). Aqueous extract: Hormesis and biostimulant activity for seed germination and seedling growth in vegetable crops. Agriculture 2022, 12, 1329. [Google Scholar] [CrossRef]
- Pannacci, E.; Masi, M.; Farneselli, M.; Tei, F. Evaluation of Mugwort (Artemisia vulgaris L.) Aqueous extract as a potential bioherbicide to control Amaranthus retroflexus L. in maize. Agriculture 2020, 10, 642. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).