
The Decreased Availability of Soil Moisture and Canopy Conductance Dominate Evapotranspiration in a Rain-Fed Maize Ecosystem in Northeastern China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Evapotranspiration and Auxiliary Measurements
2.2.1. Eddy Covariance Measurements
2.2.2. Meteorological Variable Measurements
2.2.3. Biological Variable Measurements
2.3. Statistical Analyses
2.3.1. ET Partitioning with the Shuttleworth–Wallace Model
2.3.2. Structural Equation Modeling (SEM)
3. Results
3.1. Allocation of Available Energy into Sensible and Latent Heat Fluxes
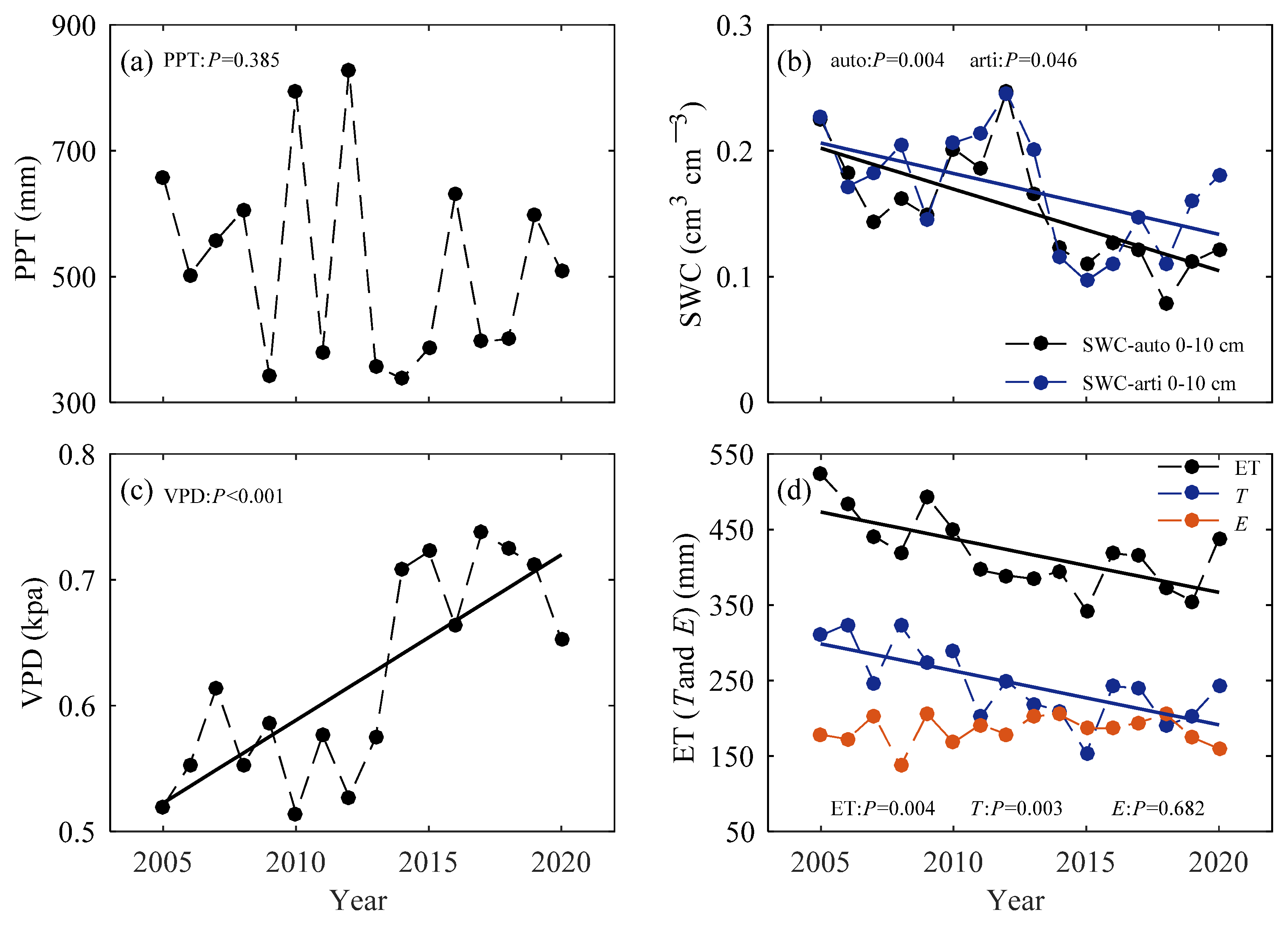
3.2. Allocation of Available Water to Soil Water and Evapotranspiration
3.3. Environmental Control on Evapotranspiration and Its Components
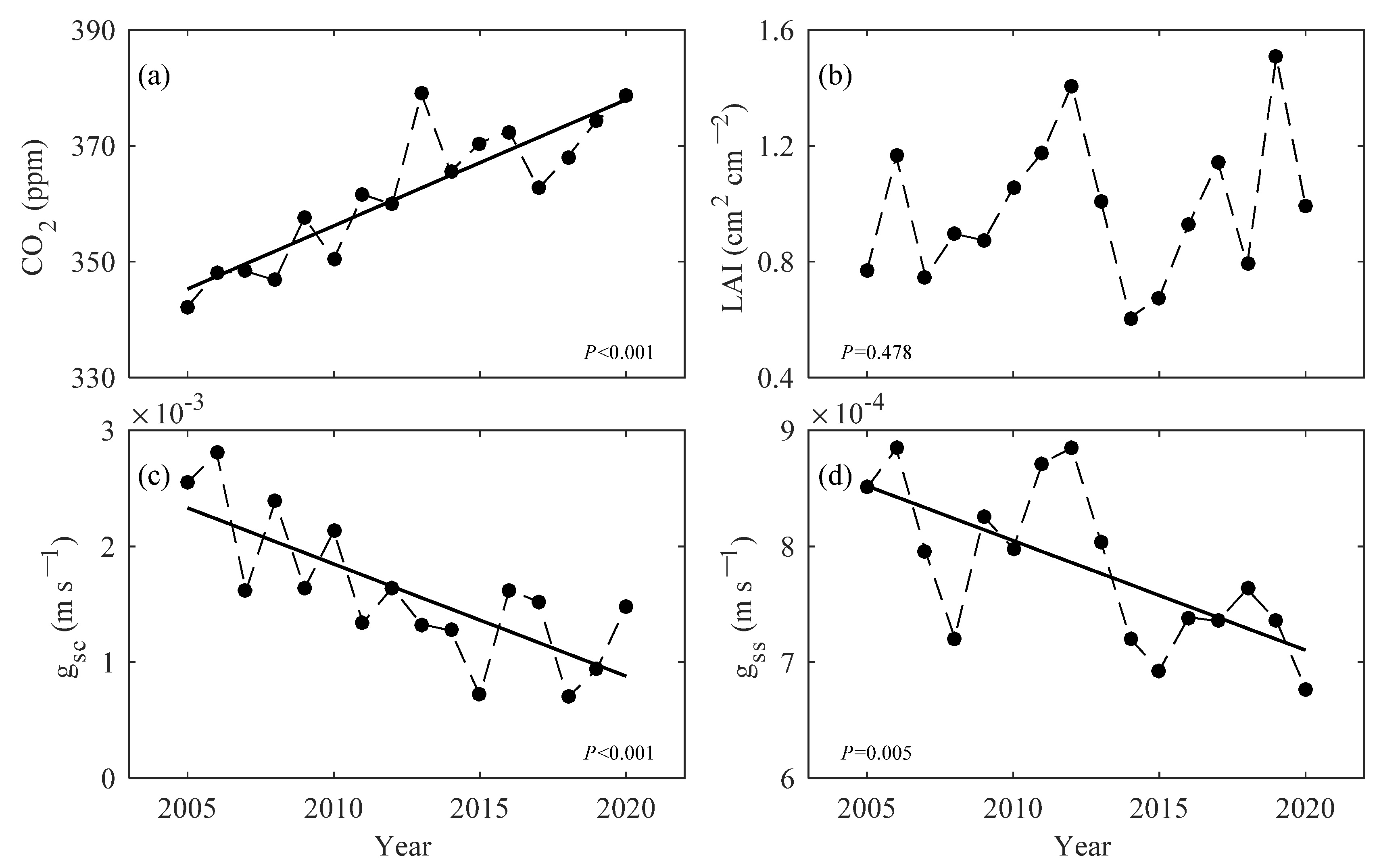
3.4. Biological Control on Evapotranspiration and Its Components
4. Discussion
4.1. Joint Limitation of Atmospheric Demand and Soil Moisture on ET
4.2. Physiological Adaptability of Plants to Climate Change
4.3. Implications for Farmland Water Management
4.4. Limitation and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, M.; Reichstein, M.; Ciais, P.; Seneviratne, S.I.; Sheffield, J.; Goulden, M.L.; Bonan, G.; Cescatti, A.; Chen, J.Q.; de Jeu, R.; et al. Recent decline in the global land evapotranspiration trend due to limited moisture supply. Nature 2010, 467, 951–954. [Google Scholar] [CrossRef]
- Fu, Z.; Ciais, P.; Feldman, A.F.; Gentine, P.; Makowski, D.; Prentice, I.C.; Stoy, P.C.; Bastos, A.; Wigneron, J.-P. Critical soil moisture thresholds of plant water stress in terrestrial ecosystems. Sci. Adv. 2022, 8, eabq7827. [Google Scholar] [CrossRef] [PubMed]
- Pascolini-Campbell, M.; Reager, J.T.; Chandanpurkar, H.A.; Rodell, M. A 10 per cent increase in global land evapotranspiration from 2003 to 2019. Nature 2021, 593, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, M.M.; Hoekstra, A.Y. The green, blue and grey water footprint of crops and derived crop products. Hydrol. Earth Syst. Sci. 2011, 15, 1577–1600. [Google Scholar] [CrossRef]
- Rockström, J.; Barron, J.; Fox, P. Water productivity in rain-fed agriculture: Challenges and opportunities for smallholder farmers in drought-prone tropical agroecosystems. Water Product. Agric. Limits Oppor. Improv. 2003, 1, 145–162. [Google Scholar] [CrossRef]
- Parent, A.C.; Anctil, F. Quantifying evapotranspiration of a rainfed potato crop in South-eastern Canada using eddy covariance techniques. Agric. Water Manag. 2012, 113, 45–56. [Google Scholar] [CrossRef]
- FAO. Faostat. Verified, 3 March 2019. 2019. Available online: http://www.fao.org/faostat/en/#data (accessed on 22 October 2023).
- Shen, Y.J.; Zhang, Y.C.; Scanlon, B.R.; Lei, H.M.; Yang, D.W.; Yang, F. Energy/water budgets and productivity of the typical croplands irrigated with groundwater and surface water in the North China Plain. Agric. For. Meteorol. 2013, 181, 133–142. [Google Scholar] [CrossRef]
- Ojara, M.A.; Lou, Y.S.; Ongoma, V.; Mumo, L.; Akodi, D.; Ayugi, B.; Ogwang, B.A. Projected changes in East African climate and its impacts on climatic suitability of maize production areas by the mid-twenty-first century. Environ. Monit. Assess. 2021, 193, 831. [Google Scholar] [CrossRef]
- Gao, X.; Mei, X.R.; Gu, F.X.; Hao, W.P.; Gong, D.Z.; Li, H.R. Evapotranspiration partitioning and energy budget in a rainfed spring maize field on the Loess Plateau, China. Catena 2018, 166, 249–259. [Google Scholar] [CrossRef]
- de Oliveira, G.; Brunsell, N.A.; Sutherlin, C.E.; Crews, T.E.; DeHaan, L.R. Energy, water and carbon exchange over a perennial Kernza wheatgrass crop. Agric. For. Meteorol. 2018, 249, 120–137. [Google Scholar] [CrossRef]
- Nazari, M.; Sadeghi, S.M.M.; Van Stan, J.T.; Chaichi, M.R. Rainfall interception and redistribution by maize farmland in central Iran. J. Hydrol. Reg. Stud. 2020, 27, 100656. [Google Scholar] [CrossRef]
- Wang, X.W.; Huo, Z.L.; Shukla, M.K.; Wang, X.H.; Guo, P.; Xu, X.; Huang, G.H. Energy fluxes and evapotranspiration over irrigated maize field in an arid area with shallow groundwater. Agric. Water Manag. 2020, 228, 105922. [Google Scholar] [CrossRef]
- Yue, P.; Zhang, Q.; Ren, X.; Yang, Z.; Li, H.; Yang, Y. Environmental and biophysical effects of evapotranspiration in semiarid grassland and maize cropland ecosystems over the summer monsoon transition zone of China. Agric. Water Manag. 2022, 264, 107462. [Google Scholar] [CrossRef]
- Condon, L.E.; Atchley, A.L.; Maxwell, R.M. Evapotranspiration depletes groundwater under warming over the contiguous United States. Nat. Commun. 2020, 11, 873. [Google Scholar] [CrossRef] [PubMed]
- Pan, R. Plant Physiology; Higher Education Press: Beijing, China, 2012. [Google Scholar]
- Zhou, Y.C.; Cheng, X.L.; Fan, J.W.; Harris, W. Patterns and controls of foliar nitrogen isotope composition on the Qinghai-Tibet Plateau, China. Plant Soil 2016, 406, 265–276. [Google Scholar] [CrossRef]
- Fu, Z.; Ciais, P.; Makowski, D.; Bastos, A.; Stoy, P.C.; Ibrom, A.; Knohl, A.; Migliavacca, M.; Cuntz, M.; Šigut, L.; et al. Uncovering the critical soil moisture thresholds of plant water stress for European ecosystems. Glob. Chang. Biol. 2022, 28, 2111–2123. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. N. Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef]
- Lin, M.; Sadeghi, S.M.M.; Van Stan, J.T. Partitioning of Rainfall and Sprinkler-Irrigation by Crop Canopies: A Global Review and Evaluation of Available Research. Hydrology 2020, 7, 76. Available online: https://www.mdpi.com/2306-5338/7/4/76 (accessed on 22 October 2023). [CrossRef]
- Wu, C.; Wang, J.; Ciais, P.; Peñuelas, J.; Zhang, X.; Sonnentag, O.; Tian, F.; Wang, X.; Wang, H.; Liu, R.; et al. Widespread decline in winds delayed autumn foliar senescence over high latitudes. Proc. Natl. Acad. Sci. USA 2021, 118, e2015821118. [Google Scholar] [CrossRef]
- Pan, S.; Pan, N.; Tian, H.; Friedlingstein, P.; Sitch, S.; Shi, H.; Arora, V.K.; Haverd, V.; Jain, A.K.; Kato, E.; et al. Evaluation of global terrestrial evapotranspiration using state-of-the-art approaches in remote sensing, machine learning and land surface modeling. Hydrol. Earth Syst. Sci. 2020, 24, 1485–1509. [Google Scholar] [CrossRef]
- Lemordant, L.; Gentine, P.; Swann, A.S.; Cook, B.I.; Scheff, J. Critical impact of vegetation physiology on the continental hydrologic cycle in response to increasing CO2. Proc. Natl. Acad. Sci. USA 2018, 115, 4093–4098. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Piao, S.; Li, L.Z.X.; Wang, T.; Ciais, P.; Lian, X.; Yang, Y.; Mao, J.; Shi, X.; Myneni, R.B. Impact of earth greening on the terrestrial water cycle. J. Clim. 2018, 31, 2633–2650. [Google Scholar] [CrossRef]
- Yu, G.; Wang, Q. Ecophysiology of Plant Photosynthesis, Transpiration, and Water Use; Science Press: Beijing, China, 2010. [Google Scholar]
- Zhang, H.; Gao, Q.; Chang, S.; Jin, C.; Liang, W.; Cai, F. Interannual variability in carbon exchange in a rain-fed maize in Northeast China and its influencing factors. J. Appl. Meteorol. Sci. 2023, 34, 246–256. [Google Scholar] [CrossRef]
- Zhang, Y. Responses and Adapatation of Crop Production to Climate Change in Northeast China; Liaoning Science and Technology Publishing: Shenyang, China, 2016. [Google Scholar]
- Chu, Z.; Guo, J.; Zhao, J. Impacts of projected climate change on agricultural climate resources in Northeast China. Acta Geogr. Sin. 2017, 72, 1248–1260. [Google Scholar] [CrossRef]
- Shuttleworth, W.J.; Wallace, J.S. Evaporation from sparse crops-an energy combination theory. Q. J. R. Meteorol. Soc. 1985, 111, 839–855. [Google Scholar] [CrossRef]
- Hu, Z.; Yu, G.; Zhou, Y.; Sun, X.; Li, Y.; Shi, P.; Wang, Y.; Song, X.; Zheng, Z.; Zhang, L.; et al. Partitioning of evapotranspiration and its controls in four grassland ecosystems: Application of a two-source model. Agric. For. Meteorol. 2009, 149, 1410–1420. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.; Mi, N.; Ming, H.; Zhang, S.; Zhang, H.; Zhao, X. Maize (Zea mays L.) physiological responses to drought and rewatering, and the associations with water stress degree. Agric. Water Manag. 2020, 241, 106379. [Google Scholar] [CrossRef]
- Mi, N.; Cai, F.; Zhang, Y.; Ji, R.; Zhang, S. Differential responses of maize yield to drought at vegetative and reproductive stages. Plant Soil Environ. 2018, 64, 260–267. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.; Ming, H.; Mi, N.; Zhang, S.; Zhang, H.; Xie, Y.; Zhao, X. Comparison of the roles of optimizing root distribution and the water uptake function in simulating water and heat fluxes within a maize agroecosystem. Water 2018, 10, 1090. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Y.; Jia, Q.; Li, R.; Zhou, M.; Zhou, G. Evapotranspiration over a rainfed maize field in northeast China: How are relationships between the environment and terrestrial evapotranspiration mediated by leaf area? Agric. Water Manag. 2019, 221, 538–546. [Google Scholar] [CrossRef]
- Ji, R. Handbook of Meteorological Services for Food Crops in Liaoning Provinece; Liaoning Science and Techinology Publishing House: Shenyang, China, 2022. [Google Scholar]
- Zhang, H.; Wen, X. Flux footprint climatology estimated by three analytical models over a subtropical soniferous plantation in Southeast China. J. Meteorol. Res. 2015, 29, 654–666. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, T.; Lyu, S.; Wu, H.; Yang, Y.; Wen, X. Interannual variability in net ecosystem carbon production in a rain-fed maize ecosystem and its climatic and biotic controls during 2005–2018. PLoS ONE 2021, 16, e0237684. [Google Scholar] [CrossRef] [PubMed]
- Pastorello, G.; Trotta, C.; Canfora, E.; Chu, H.; Christianson, D.; Cheah, Y.-W.; Poindexter, C.; Chen, J.; Elbashandy, A.; Humphrey, M.; et al. The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data. Sci. Data 2020, 7, 225. [Google Scholar] [CrossRef] [PubMed]
- Code for Agrometeorological Observation; Meteorology Press China Meteorological Administration: Beijing, China, 1993.
- Hu, Z.; Wu, G.; Zhang, L.; Li, S.; Zhu, X.; Zheng, H.; Zhang, L.; Sun, X.; Yu, G. Modeling and partitioning of pegional pvapotranspiration using a satellite-driven water-carbon coupling model. Remote Sens. 2017, 9, 54. [Google Scholar] [CrossRef]
- Hu, Z.; Li, S.; Yu, G.; Sun, X.; Zhang, L.; Han, S.; Li, Y. Modeling evapotranspiration by combing a two-source model, a leaf stomatal model, and a light-use efficiency model. J. Hydrol. 2013, 501, 186–192. [Google Scholar] [CrossRef]
- Euskirchen, E.S.; Edgar, C.W.; Bret-Harte, M.S.; Kade, A.; Zimov, N.; Zimov, S. Interannual and seasonal patterns of carbon dioxide, water, and energy fluxes from ecotonal and thermokarst-impacted ecosystems on carbon-rich permafrost soils in Northeastern Siberia. J. Geophys. Res. Biogeosci. 2017, 122, 2651–2668. [Google Scholar] [CrossRef]
- Babel, W.; Lüers, J.; Hübner, J.; Rebmann, C.; Wichura, B.; Thomas, C.K.; Serafimovich, A.; Foken, T. Long-Term Carbon and Water Vapour Fluxes; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Budyko, M.I. Climate and Life; Academic Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Donohue, R.J.; Roderick, M.L.; McVicar, T.R. On the importance of including vegetation dynamics in Budyko’s hydrological model. Hydrol. Earth Syst. Sci. 2007, 11, 983–995. [Google Scholar] [CrossRef]
- Liu, Y.; Qiu, G.; Zhang, H.; Yang, Y.; Zhang, Y.; Wang, Q.; Zhao, W.; Jia, L.; Ji, X.; Xiong, Y.; et al. Shifting from homogeneous to heterogeneous surfaces in estimating terrestrial evapotranspiration: Review and perspectives. Sci. China Earth Sci. 2022, 65, 197–214. [Google Scholar] [CrossRef]
- Yuan, W.P.; Zheng, Y.; Piao, S.L.; Ciais, P.; Lombardozzi, D.; Wang, Y.P.; Ryu, Y.; Chen, G.X.; Dong, W.J.; Hu, Z.M.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, 12. [Google Scholar] [CrossRef]
- Aubinet, M.; Vesala, T.; Papale, D. Eddy Covariance—A Practical Guide to Measurement and Data Analysis; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Mauder, M.; Foken, T.; Cuxart, J. Surface-energy-balance closure over Land: A review. Bound. Layer Meteorol. 2020, 177, 395–426. [Google Scholar] [CrossRef]
- Burba, G. Eddy Covariance Method-for Scientific, Regulatory, and Commercial Applications; LI-COR Biosciences: Lincoln, NE, USA, 2022. [Google Scholar]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef]
- Liu, L.; Gudmundsson, L.; Hauser, M.; Qin, D.; Li, S.; Seneviratne, S.I. Soil moisture dominates dryness stress on ecosystem production globally. Nat. Commun. 2020, 11, 4892. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Q.; Wang, C.Y.; Zhang, F.W.; He, Y.T.; Shi, P.L.; Guo, X.W.; Wang, J.B.; Zhang, L.M.; Li, Y.N.; Cao, G.M.; et al. Atmospheric water vapor and soil moisture jointly determine the spatiotemporal variations of CO2 fluxes and evapotranspiration across the Qinghai-Tibetan Plateau grasslands. Sci. Total Environ. 2021, 791, 148379. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, S.; Zhao, S.; Zhu, Y.; Feng, S.; Wang, Z.; Wu, Y.; Xiao, J.; Yuan, W.; Yan, W.; et al. Temporal dynamics of ecosystem, inherent, and underlying water use efficiencies of forests, grasslands, and croplands and their responses to climate change. Carbon Balanc. Manag. 2023, 18, 13. [Google Scholar] [CrossRef]
- Konings, A.G.; Williams, A.P.; Gentine, P. Sensitivity of grassland productivity to aridity controlled by stomatal and xylem regulation. Nat. Geosci. 2017, 10, 284–288. [Google Scholar] [CrossRef]
- Guo, Q.; Hu, Z.; Li, S.; Li, X.; Sun, X.; Yu, G. Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: Effects of mean annual precipitation and its seasonal distribution. Glob. Chang. Biol. 2012, 18, 3624–3631. [Google Scholar] [CrossRef]
- Dennis, B.D.; Christoph, V.A.; Brad, H. Seasonal variation of energy and water vapor exchange rates above and below a boreal jack pine forest canopy. Agric. For. Meteorol. 1997, 102, 28939–28951. [Google Scholar] [CrossRef]
- Yue, P.; Zhang, Q.; Zhang, L.; Li, H.Y.; Yang, Y.; Zeng, J.; Wang, S. Long-term variations in energy partitioning and evapotranspiration in a semiarid grassland in the Loess Plateau of China. Agric. For. Meteorol. 2019, 278, 107671. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Still, C.J. 3 Trading water for carbon. Nat. Geosci. 2018, 11, 702–703. [Google Scholar] [CrossRef]
- Griebel, A.; Bennett, L.T.; Metzen, D.; Pendall, E.; Lane, P.N.J.; Arndt, S.K. Trading water for carbon: Maintaining photosynthesis at the cost of increased water loss during high temperatures in a temperate forest. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005239. [Google Scholar] [CrossRef]
- Wang, J.; Wen, X.; Zhang, X.; Li, S. The strategies of water–carbon regulation of plants in a subtropical primary forest on karst soils in China. Biogeosciences 2018, 15, 4193–4203. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Caemmerer, S.V.; Berry, J.A. A biochemical-model of photosynthetic CO2 assimilation in leaves of C-3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhao, T.; Ji, R.; Chang, S.; Gao, Q.; Zhang, G. The Decreased Availability of Soil Moisture and Canopy Conductance Dominate Evapotranspiration in a Rain-Fed Maize Ecosystem in Northeastern China. Agronomy 2023, 13, 2941. https://doi.org/10.3390/agronomy13122941
Zhang H, Zhao T, Ji R, Chang S, Gao Q, Zhang G. The Decreased Availability of Soil Moisture and Canopy Conductance Dominate Evapotranspiration in a Rain-Fed Maize Ecosystem in Northeastern China. Agronomy. 2023; 13(12):2941. https://doi.org/10.3390/agronomy13122941
Chicago/Turabian StyleZhang, Hui, Tianhong Zhao, Ruipeng Ji, Shuting Chang, Quan Gao, and Ge Zhang. 2023. "The Decreased Availability of Soil Moisture and Canopy Conductance Dominate Evapotranspiration in a Rain-Fed Maize Ecosystem in Northeastern China" Agronomy 13, no. 12: 2941. https://doi.org/10.3390/agronomy13122941
APA StyleZhang, H., Zhao, T., Ji, R., Chang, S., Gao, Q., & Zhang, G. (2023). The Decreased Availability of Soil Moisture and Canopy Conductance Dominate Evapotranspiration in a Rain-Fed Maize Ecosystem in Northeastern China. Agronomy, 13(12), 2941. https://doi.org/10.3390/agronomy13122941