Abstract
Secondary nutrient (e.g., calcium, magnesium, sulfur) deficiencies in crop plants disturb the nitrogen balance in the plants, thus reducing the overall yield. This hypothesis was analyzed based on the physiological functions of these nutrients, in relation to the uptake and utilization of N, in crop plants. Nitrogen uptake by plants requires a well-developed root system, the size of which depends on the supply of calcium. This process is largely controlled by the content of toxic aluminum in the soil, which can be mitigated through the application of lime and/or gypsum. In humid climates, the excessive uptake of calcium by plants occurs during water shortages; this process significantly interferes with N uptake. Magnesium, which affects plant growth throughout the growing season, can effectively control excessive calcium uptake. Magnesium deficiency can be ameliorated with soil- or foliar-applied fertilizers. These stages define the timing of plant sampling and determination of the N:S ratio, as an indicator of plant nutritional status. The application of Mg, S, or MgS facilitates higher productivity of fertilizer N by narrowing the N:Mg and N:S ratios in plants. The use of secondary nutrients can allow farmers to obtain high yields while reducing both production costs and environmental risks.
1. Introduction
The demand for food is expected to constantly grow in the coming years, which clearly indicates the need to increase the yields of basic staple crops [1]. However, this goal cannot be achieved without increased Nf consumption [2]. The dilemma is how to increase yields while, at the same time, reducing the pressure of the active N on the environment [3]. Various solutions have been considered, such as changing people’s eating habits [4]. These theoretical solutions may be interesting, but eating habits so far are consistent with Engel’s law. This law states that as household income increases, the structure of consumption changes. In the first phase of this process, the consumption of starch products (cereals, tubers, roots) decreases in favor of meat and dairy products. In the second stage, a significant decrease in the share of expenditure on food is noticeable. Currently this phenomenon is most intense in Asia and South America [5,6].
Modern crop plant varieties, as a result of genetic modification (dwarf genes), have high yield potential. However, the higher yields that can be harvested by farmers require the optimal water conditions during the growing season and application of nitrogen fertilizer (Nf) at high doses. The second condition entails full pesticide protection of plants throughout their entire life-cycle (i.e., the growing season) [7]. This strategy has come under severe criticism, since the Nf production efficiency is low (e.g., well below 50% for cereals) [8]. Such a low recovery of Nf by plants during the single growing season, in fact, indicates the high potential threat of the residual Nf to the environment [9].
Increased nitrogen use efficiency (NUE) has been of great interest to scientists for about 40 years [10]. NUE is an aggregate index of N productivity, consisting of two units—Nitrogen Uptake Efficiency (NUpE) and Nitrogen Utilization Efficiency (NUtE)—as follows:
NUE = NUpE × NUtE
The first sub-unit, NUpE, expresses the efficiency of N uptake by the currently grown plant, and is defined as the ratio of the amount of N accumulated in the plant to the amount of its available forms in the soil during the growing season. The question is: what nutrients (apart from N) are necessary for efficient uptake of inorganic N from the soil solution. The nutrient uptake process is energy-dependent, determined by the ATP-ase activity in the root cells. In fact, the efficiency of the proton pump—that is, the rate of proton (H+) transfer from the cell cytoplasm to its apoplast—is conditioned by the nutritional status of the plant, in terms of phosphorus (P) and magnesium (Mg). Moreover, the uptake of nitrate ions (NO3−) requires the presence of accompanying cations. Potassium (K+) is the dominant cation, followed by Mg2+ or Ca2+, as indicated by their content and accumulation in a crop biomass [11,12]. The second NUE sub-unit, NUtE, is defined as the ratio of the yield (main or total) to the amount of N accumulated by the grown crop at harvest [10,13]. The question is: what are the roles and significance of other nutrients in the process of the re-mobilization and translocation of organic compounds to the harvestable part of the plant? All of these processes require energy, which depends of the activity of ATP-ase. These processes depend on numerous nutrients, including calcium, sulfur, and micronutrients [11,14].
In agriculture, there are two main strategies for increasing NUE. In the case of N uptake by plants, two solutions are currently discussed. The first is to increase the depth of the plant root system growth in the soil, as proposed by breeders [15]. In recent decades, they have recognized opportunities and made efforts to develop methods for modifying the architecture of the root system of the crop plants. The so-called steep, cheap, and deep concept assumes that increased vertical root growth in the soil profile results in higher N uptake. This means that the inorganic N that is leached into the lower soil layers can be captured by the plant, provided deeper plant root penetration in the soil profile. The second approach, called foraging, focuses on modifying the root system in such a way as to increase the uptake of N from soil patches artificially enriched with Nf [16,17].
Environmental policy-makers, in light of the negative impacts of the applied Nf on the environment, have attempted to focus the attention of both advisors and farmers on two other solutions. The first group, however, presents a reductionist vision of food production. These methods involve a sharp decrease in the applied Nf dose. The assumed effect of these activities is, of course, a reduction in Nf pressure on the environment. Unfortunately, this strategy often leads to a decrease in yield [18]. The second group—the modern one—focuses on optimization of the applied Nf dose by adjusting it to the temporally and spatially different requirements of the plant canopy for Nf [19,20,21].
The current problems facing the food market—resulting directly and indirectly from the war in Ukraine—have led to a drastic increase in the prices of fertilizers, especially nitrogenous ones [22]. Consequently, this situation has resulted in a significant reduction in Nf consumption. This unexpected situation in the world food market clearly indicates the urgent need to find a different set of solutions to maintain food production in main regions of the World, including Europe [23]. In the current situation, in this basic food production sector, farmers should pay more attention to the use of nutrients from the so-called secondary group, consisting of calcium (Ca), magnesium (Mg), and sulfur (S) [24]. The effects of this set of nutrients on NUE, despite a good knowledge of their physiological functions in plants, is poorly understood. There are several reasons for this state of knowledge and limitations in practical application. The first is the insufficient—or even lack of—knowledge regarding the production functions of these nutrients in crops. The second weakness is their poorly understood impact on Nf management by crop. This applies to both the role of these nutrients in the uptake of inorganic N compounds and the efficiency of N transformation in the plant into yield. The third is a lack of sufficient data on their impact on the formation of yield components. The latter two points are closely related to each other. Therefore, the key objective of this conceptual review is to answer the following question: to what extent can secondary nutrients affect the productivity of nitrogen fertilizer? The phenomena and processes discussed in the article concern the natural conditions of plant growth. An essential component of the so-called rain-fed agriculture is the sum and distribution of rainfall during the growing season.
2. Uptake and Accumulation of Calcium and Magnesium by Crop Plants
Transport of Ca and Mg ions in the soil solution towards the root surface is carried out by interception and mass flow [11]. These processes take place in the juvenile, apical zone of the root, in which the basic tissues have not yet reached maturity. A fully developed endoderm with a well-formed casparian band serves as a strong barrier to the flow of water and multi-valent cations, including Ca and Mg, into the xylem [25]. The mature endoderm is probably the reason why the amount of both of these nutrients, arriving at the root apoplast, is several times greater than that which the real plant needs [26]. It seems logical that efficient supply of the currently grown plant with Ca and Mg requires the continuous growth of new roots in the soil. However, it cannot be rejected that the root is a temporary storage plant organ for both cations. This hypothesis can be confirmed by the fact that plants from Fabaceae and Brassicaceae families contain most of the accumulated Ca in pods [27,28]. The growth of plant roots at the Grain-Filling Period (GFP) is strongly reduced, but not stopped, as shown in Figure 1. The stage of the maximum standing weight in winter rye roots has been found to depend on the type of fertilization. On a control plot, which was not fertilized for about 30 years, the maximum root mass was recorded before flowering whereas, for plants fertilized with NPK (an annual Nf dose = 90 kg ha−1), it was at the beginning of the milk phase. The most dynamic period of root growth extended from the beginning of the stem elongation phase up to flowering. This period covers the yield construction mega-phase (YCC) of seed plant growth [29]. In cereals, the supply of N to plants, especially during the booting stage, is decisive for the grain density (GD), which is the key yield component [30]. The observed greater investment of plants grown on the absolute control into root biomass, as compared to those fertilized with NPK, likely resulted from a shortage of available N in the growth medium [31]. It is necessary to add that, in a presented case, the grain yield on an NPK plot was about two times greater than on the absolute control [32]. The period of winter rye growth—extending from the beginning of heading to the beginning of milk development—is characterized by a very stable standing root mass. New root growth has also been observed during the GFP; however, it was much slower, compared to the rate of death of older roots. The main reason for this phenomenon is the appearance of grains as a new, much stronger physiological sink. The presented example clearly shows that cereals produce new roots, even under the dominance of the reproductive organs. This information indicates that seed plants can actively take up nutrients such as Ca and Mg from the soil solution during the GFP. However, it seems logical that a major part of the Ca present in pods of Fabaceae and Brassicaceae at maturity originate from its resources accumulated in its roots during the vegetative stages of growth.
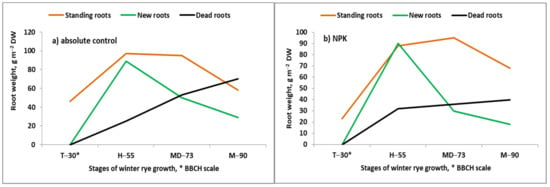
Figure 1.
Effects of soil fertility on patterns of roots dynamics in winter rye during the growing season (based on Grzebisz [33]). Legend: T, stem elongation phase; H, heading; MD, milk development; M, maturity. * stages of winter rye growth according to the BBCH scale [34].
The hypothesis presented above regarding the net uptake of Ca and Mg during the GFP undoubtedly confirms their significant accumulation in seed crops during this period. As shown in Figure 2, the amount of both nutrients increased in winter wheat during GFP by 24.5% for Ca, and by 38% for Mg. Three conclusions can be drawn by analyzing this figure. First, the greater amount of both nutrients at wheat maturity was accumulated in stems, where the degree of increase was more pronounced for Ca. Second, the translocation efficiency—expressed by the nutrient harvest index (HI)—was four times higher for Mg, achieving 11.4% for Ca and 44.8% for Mg. Third, leaves appeared as the plant organ depleted with both nutrients, but ears were the net source only for Mg.
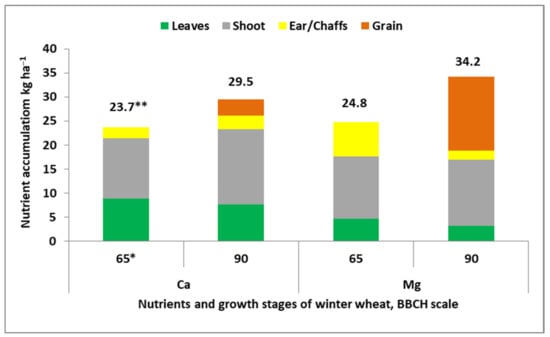
Figure 2.
Accumulation and partitioning of Ca and Mg between winter wheat parts at the beginning and end of the grain-filling phase (based on Nowicki, [35]). Legend: * 65–, 90: growth stages of winter wheat, according to the BBCH scale; **, total accumulation.
A low level of Ca accumulation in seeds or grain is typical for seed plants. The harvest index for Ca (CaHI) in seed plants is low, usually below 10% [36,37]. As shown in Figure A1, the total amount of Ca accumulated by winter wheat at harvest increased in accordance with applied Nf rates. The recorded trend resulted mainly from the uptake of nitrates, for which Ca2+ is an accompanying cation. An inverse relationship was observed for ammonia [38]. On the other hand, CaHI responded inversely to increasing Nf rates, decreasing from 12% for plants grown on the N control plot to 10% for plants fertilized with high Nf dose (240 kg N ha−1). Such a small difference in the value of the index between the extreme treatments indicates its conservative character. Moreover, the amount of Ca in stems was in accordance with increasing N rates while, in grain, it reached a maximum at the Nf rate of 228 kg N ha−1.
The pattern of Mg accumulation by crop plants in response to increasing Nf rates differs. As shown in Figure A2, its total accumulation by winter wheat increased gradually with the Nf dose. The Mg harvest index (MgHI) showed the same pattern, increasing from 40% for plants grown on the N control plot to 47% on plots fertilized with 200 kg N ha−1. The recorded trends confirmed the strong relationship between Mg and nitrates [38]. The Mg accumulated in grain, in contrast to Ca, increased linearly with the applied Nf doses. On the other hand, for straw, the maximum Mg in wheat biomass was recorded at Nf = 232 kg ha−1. This trend indicates that Mg is relatively well re-mobilized from the vegetative wheat parts during GFP.
The pattern of in-season Ca accumulation by root crops has been obtained for sugar beets (as shown in Figure 3). Its total amount in crop biomass depends, to a great extent, on the biomass of leaves. In the presented case, the maximum uptake of Ca was recorded around the 110th day after sowing of sugar beets. This critical date is defined by the date of its maximum rate of daily accumulation in the leaves [39]. The accumulation of Ca by sugar beets in storage roots increases gradually during the growing season. The maximum daily rate of Ca accumulation reaches its maximum before the maximum recorded for the leaves [39]. Its downward trend (as shown in Figure A3) indicates a dilution effect on the Ca content, resulting from the progressively increased root biomass. The CaHI, affected by the gradual storage root biomass growth, increases progressively during the growing season.
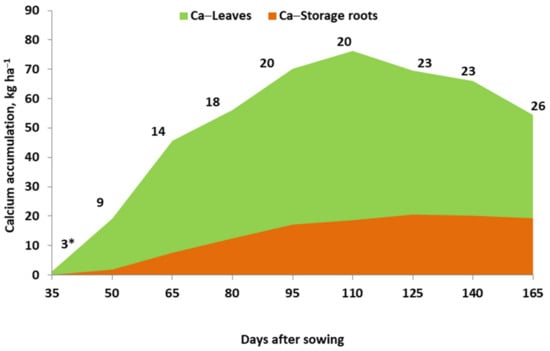
Figure 3.
Pattern of calcium accumulation and partitioning between storage roots and leaves of sugar beet during the growing season. (Grzebisz, W., unpublished). *, calcium harvest index (%).
Uptake of Ca and Mg by plants from the soil is often disturbed by many factors. The most important ones that require the attention of farmers are:
- (1)
- The low content of available nutrients in the soil rooting zone. This is typical for sandy soils and in regions of the world with high precipitation [40];
- (2)
- Soil acidity and the excessive content of Al3+, which is a strong Ca2+ antagonist [41];
- (3)
- (4)
- Excessive fertilization with nitrogen fertilizers containing ammonium (N–NH4). Ammonium exerts a strong negative effect on the uptake of both nutrients from the soil solution [11];
- (5)
- Weather conditions, such as drought leading to a shortage of water in the soil.
Under conditions of water shortage in the soil, antagonism between Ca and Mg may occur. The main reason is the excessive accumulation of Ca in the plant in critical stages of yield formation. This phenomenon has been observed in maize, spelt wheat, winter oilseed rape, and potatoes [36,42,43,44]. This excessive accumulation of Ca can significantly disturb the N management by the crop, as has been observed for spelt wheat [42]. In this particular case, the excess of Ca—which first appeared before flowering—resulted in a reduction in grain density and, thus, in yield. The application of Mg has emerged as an agronomic factor mitigating—at least in part—these negative processes [42]. The same phenomenon has been observed in potato plants. In this case, excessive accumulation of Ca in stems in the early stages of potato growth resulted in narrowing of the Ca:N ratio. Consequently, the marketable tuber yield (MTY) was significantly reduced [43]. As can be calculated using the equation below, the lowest tuber yield of 29.2 t ha−1 was obtained for a Ca:N ratio of 5:10. At a Ca:N of 1:10, the MTY was 66.3 t ha−1.
The relationship between successive pairs of nutrients, such as Ca × K, Ca × Mg, and Mg × K, assessed based on the amount of these nutrients in plant biomass at harvest, indicates the occurrence of three stages: synergism, neutrality, and antagonism. Therefore, each pro-productive assessment of the plant nutritional status of a currently grown plant should be carried out before the critical phase of yield component formation. In farming practice, there are rarely problems with the correction of the Mg in the crop, which are sometimes raised [45]. Foliar fertilization of crops with Mg in strictly defined phases of growth can significantly reduce the deficiency of this nutrient [46,47]. The often-discussed antagonism between K and Ca manifests mainly under water shortage, as has been documented for potato by Potarzycki and Grzebisz [44]. Magnesium fertilization significantly controls Ca accumulation by crop plants, mitigating its negative impact on N management and yield [42,48].
One of the most important pieces of information needed by the farmer to effectively manage a currently grown plant is the amount of a nutrient necessary for the crop to produce the intended yield. The calculation is based on an index called unit nutrient accumulation (UNA). Table 1 shows the ranges for only the index values of N and secondary nutrients. One of the basic principles affecting the accumulation of secondary nutrients in a crop is the increase in the UNA index in accordance with the yield. This is due to the faster increase of the by-product than the main yield [48]. The only exception is N, whose content affects the main yield, while in the by-product its content decreases with an increase in the main yield [49].

Table 1.
Unit nutrient accumulation for selected crop plants (kg t−1 main yield 1).
3. Physiological Basis for Use of Calcium and Magnesium in Crop Production
3.1. Calcium
The content of Ca in crop plants is very wide, ranging from 0.1 to 4.4% DW [61]. Thus, the proper functioning of the plant depends on the supply of Ca, but its requirement varies between species. The recorded concentrations of Ca in indicative plant parts are usually low for cereals (2–5 g kg−1 DW); much greater for root crops, such as sugar beets and cassava (5–15 g kg−1 DW); and the highest for cruciferous plants, such as cabbage and oilseed rape [62,63]. For these reasons, dicots are considered as more sensitive than monocots (including cereals) to Ca supply.
The functions of Ca in the plant are currently recognized in two main groups of processes. The first refers to its roles in the development and stability of the plant structure at each level of its organization. The second role, as a secondary messenger, is currently considered crucial in the control of physiological processes in the plant [64]. The formation and stability of plant tissues requires Ca, as a component of the middle lamella—the plant anatomical unit that connects the plant cells together. The basis for this specific Ca function is the fact that the hydrated Ca2+ cation, when undergoing dehydration in the polysaccharide matrix of the emerging cell wall, reacts with pectins. The second important structural part of the plant is the cell membrane. The Ca2+ cation, by creating bonds with phospholipids, stabilizes the structure of the lipid double layer. In this way, Ca controls the permeability of the plasmalemma (flow) to substances of different origin, including nutrients [63]. However, the role of Ca in the mitotic division of meristematic buds (of both roots and above-ground shoots) cannot be overlooked. The initiation of both the root and shoot systems is crucial for the start of the plant’s life-cycle [65].
The concentration of Ca in the cytoplasm of a plant cell in a non-stressed plant is typically at a level of 0.1 μM. Such a low concentration of Ca in the cytosol, compared to its total concentration in the plant, is due to two main reasons. The first is the limited transport of Ca2+ ions through the plasmalemma (the so-called symplastic route). The second is the internal control mechanisms of the cell, maintaining the Ca2+ concentration in balance with other components of the cell. The key reason for the low Ca concentration is the high potential of Ca2+ for rapid dehydration, thus binding to anions present in the cytosol, such as phosphate (PO43−), as well as its competition with Mg2+. Excess Ca in the plant cell is translocated to the vacuole, forming several compounds (Ca-oxalate, Ca-citrate, and so on) [11,63]. This process is accelerated in plants fed with nitrates [11]. Despite the existing control mechanisms, the cytosolic Ca2+ concentration is highly sensitive to both abiotic stimuli, such as environmental (drought, salinity, elevated temperature) and nutritional (nutrient deficiencies); and biotic stimuli, including developmental (morphogenesis) and attack (e.g., pathogens or wounding). In response to any of these stimuli, the concentration of Ca2+ in the cytosol increases rapidly. The sources of Ca transported to the cytosol are its resources in the apoplast and in intracellular organelles, mainly in the vacuole [65]. A sudden increase in Ca2+ concentration in the cytosol is one of the earliest signals of stress, triggering a cascade of physiological processes aimed at maintaining the plant growth homeostasis [64,66]. At this point of analyzing the function of Ca in plants, an important fact must not be omitted: the content of Ca in the plant apoplast is many times higher than in the cytoplasm of the cell. An important message for the farmer is the need to keep the plant in a favorable state of Ca nutrition. Only under such conditions is the crop plant is able to efficient, quickly respond to any stresses. However, it should be remembered that “favorable Ca nutritional status” involves its balance with other nutrients [11].
Two classic examples help us to understand the function of Ca in the plant as a secondary messenger. The first is the plant’s response to water shortage. Increasing transpiration increases the uptake of Ca from the soil solution, which is the first signal of abiotic stress. In the first stage of water stress, the uptake of Ca is increased by the plant, resulting in its higher concentration in guard cells. This process is associated with a simultaneous increase in the concentration of the abscisic acid (ABA). The increasing concentration of ABA induces an influx of Ca into the stomata, leading to a simultaneous efflux of K+ ions into the apoplast. Consequently, the leaf turgor decreases and the plant’s stomata close [67]. The key negative aspect of these water-saving processes is the reduced supply of CO2 which, in turn, slows down the growth rate of the plant, ultimately leading to decreased yield [11,42]. The second example of cytosolic Ca2+ signaling is the plant’s response to pathogenic attack. The actual infection of the plant occurs when the pathogen hyphae penetrate the barrier; that is, the plasmalemma. At this critical stage of pathogenesis, three nutrients play important roles; namely, K, Cl, and Ca. Damage of the plasmalemma causes the leakage of K+ and Cl− from the cytosol into the apoplast. This process is associated with the reverse transportation of H+ and Ca2+ [68,69]. The nutritional status of the plant, in terms of K and Ca, facilitates the plant’s defense mechanisms against the pathogen, through determining the strength of the transmitted stress signal. A decreasing K concentration in the plant cell stimulates the activity of NADPH oxidase, leading to the increased production of reactive oxygen species. A change in the Ca2+ concentration in the cytosol activates local and systemic signaling in the plant. Signal carriers are hormones such as salicylic acid, jasmonic acid, and ethylene [68].
Ca plays a crucial role in the initial stages of the symbiosis between the legume plant and Rhizobium bacteria. According to the calcium oscillation theory, signal molecules excreted by Rhizobium bacteria (Nod factors) induce oscillations of Ca2+ in the nucleus of the root hair of the host plant [70]. In response, the bacteria attach to the root hair of the host plant using a Ca-fixation protein [71]. The growth of the root nodule requires a good supply of Ca, which is needed to build up its cellular structure.
The concentration, as well as the partitioning of Ca between of the plant organs at harvest, is a conserved trait of certain crops:
- Maize → leaves > stem > cob-covering leaves, grain > cob core [36];
- Potato → leaves > stem > stolons + roots > tubers [44].
The dominance of leaves over other plant organs for both traits clearly highlights the low rate of Ca re-mobilization to the growing reproductive and/or storage organs of the crop. The rate of Ca re-mobilization from leaves and stems is extremely low, mainly due to its poor transportation in the phloem. Thus, the most-affected plant parts by Ca deficiency are those which present a low transpiration rate (e.g., buds, young leaves), as well as fruits, in which the supply of Ca through the phloem is limited [38,72]. In a consequence, the demand of growing tissues for Ca depends on efficiency of its uptake by roots from the soil solution [73].
In general, Ca deficiency in classic field crops occurs only under extremely bad weather conditions, usually prolonged drought or in acidic soils. On sandy soils, which are naturally poor in total and available Ca, the low supply of Ca to the actually grown crop is often overlooked by farmers. The lower density of plants or a weak root system is, as a rule, considered to be caused by Al3+ toxicity (see Section 5), water shortages, and possibly low supply of N, K or other nutrients. The key set of factors contributing to Ca deficiency were presented in the previous chapter. A deficiency of Ca in the soil results in a less-developed root system, leading to decreased supply of nutrients to the plant [74]. Consequently, the plant growth rate decreases (Figure 4a). Moreover, during the growing season, some plants may fall out of the field. An exceptional situation (as shown in Figure 4b) is the drooping of inflorescences. The key reason for this is probably the low saturation of the plant vascular tissue with Ca.

Figure 4.
Winter oilseed rape response to deficiency of calcium: (a) tipburn and stunted stature of the plant grown in acid soil (Rumex acetos, dried sorrel); and (b) drooping of the rapeseed inflorescences (photos by Witold Grzebisz).
The functions of Ca in the plant, as specified above, constitute the physiological basis for the performance of yield-forming functions in crop plants. The growth rate of the plant, the formation of the yield components and, finally, the yield are affected as the result of Ca having an impact on:
- (1)
- Conditions for nutrient uptake, both by stimulating the growth of the primary root (seminal roots) and initiating the lateral roots;
- (2)
- Signaling the disorder in plant growth by increasing its content in the plant indicative parts;
- (3)
- The relationship between the supply of N and other nutrients, being decisive for their uptake and utilization by the plant;
- (4)
- Plant health, as a result of both a balanced nutritional status and indirect reduction of pathogen pressure;
- (5)
- Intensification of N2 fixation as a result of greater root nodulation of legumes in soil rich in available Ca;
- (6)
- The quality of the products of grown crops—this applies mainly to the market quality of fruits, leafy vegetables, and potato tubers.
3.2. Magnesium
The content of Mg in crop plants ranges from 0.1 to 1.5% DW, and is conditioned by numerous factors (as discussed in Section 2). All of these factors are responsible not only for the total Mg content, but also for its content in edible plant parts [75]. The key problem of modern intensive agriculture, based on new, very productive varieties and high doses of applied Nf, is the so-called syndrome of breeding-induced Mg dilution, observed in grain, seeds, vegetables, and fruits [76,77]. In cereal grains, the basic source of staple food for humans, the Mg content increases from about 600 (1000) mg kg−1 for rice, rye, soft wheat, and triticale to 1400 (1800) mg kg−1 DW for corn, oats, and especially spelt wheat. Legume seeds—an important source of protein—are usually much richer in Mg, the content of which can reach 2000 mg kg−1 DW. Fruits, as a rule, are a poor source of this nutrient. Vegetables, especially those representing cruciferous plants, as well as tomato, lettuce, and carrot, are a good source of Mg (>1300 mg kg−1 DW). The best field carriers of Mg are pumpkin (3000 mg kg−1 DW), buckwheat, radish (>2500 mg kg−1 DW), and green beans (>2000 mg kg−1 DW) [78].
The core function of Mg in plant production is its content in the chlorophyll molecule, in which it is bound to the porphyrin ring [79]. The inherent interaction between N and Mg acts in the plant as the photosynthetic engine; as such, any energy transformation process in a plant requires a certain amount not only of P, but also Mg [80]. Other key physiological functions of Mg include loading of the phloem and the transport of sugar from the leaves to the reproductive plant organs (e.g., seeds, grains, fruits). In addition, the effects of Mg on the activity of about 350 plant enzymes involved in stress signaling have been detailed in various textbooks and scientific articles [81].
The harvest index of Mg is much lower, compared to N and P, being in the range of 40–60% [78]. This can be explained by the Mg concentration in plant tissues of seed plants at maturity. A classic example is maize, where the order of maize organs at physiological maturity, with respect to Mg concentration, is significantly affected by the weather [36]:
- (1)
- Optimal weather conditions → leaves > grain > stem > cob-covering leaves > cob core;
- (2)
- Water shortage → leaves > cob core = stem = grain > cob-covering leaves.
The first place of leaves, regardless of the weather during the growing season, clearly indicates a moderate Mg remobilization potential during grain growth, especially in years characterized by drought. The increase in yield in response to Mg fertilization for most crops is about 10% or less, and is directly related to the increase in N productivity [82]. The effect of applying Mg on yield in seed crops is mediated through its impact on GD, as has been observed for cereals, maize, winter oilseed rape, and sugar beets [83,84,85,86]. Another important aspect in the response of crops to the application of Mg is increasing its content in the edible parts of plants. This is the easiest way to overcome Mg deficiencies in food, mainly due to the high yields of modern crops [75].
Mg deficiency in crop plants, regardless of the Nf dose, causes significant growth disturbances in crop plants. This nutrient, unlike Ca, is mobile in both the xylem and phloem [11]. Thus, its deficiency during the intensive growth of the plant can be compensated for by physiological and morphological adaptation. It is important for the farmer to recognize these stages early enough to mitigate any potential reduction in plant growth and, consequently, the yield [87]. The stages of crop adaptation to the Mg deficiency are as follows [78]:
3.2.1. Early Adaptation
Deficiency of available Mg in the soil induces plant activity, which manifests in the increased growth of new roots and accelerated activity of Mg transporters in the root apical zone [88]. A practical solution is to apply (before sowing or during the early stages of the growing season) fertilizers containing Mg in water-soluble form [87].
3.2.2. Advanced Adaptation
At this stage of Mg deficiency, the plant uses Mg resources stored in the root. It has been well-documented that the amount of Mg2+ accumulated in the root is several times greater than its actual demand [26]. Indirect information regarding a probable Mg deficiency is the low level of available Mg in the soil. A prophylactic solution is top-dressing and/or foliar fertilization of the currently grown crop [46].
3.2.3. Critical Adaptation
Plant growth depends on internal Mg resources (i.e., that accumulated in older leaves). Visual deficiency symptoms are not yet visible, but the nutritional status is insufficient. Determining the deficiency (which is called hidden at this stage) requires analysis of the plant’s indicatory organs. The current state of Mg deficiency is the basis for the farmer to determine the dose and selection of fertilizer for foliar spraying [46].
3.2.4. Growth Rate Degradation
The Mg nutritional plant status with visible symptoms indicates a deep disturbance of plant metabolism. The decrease in yield may even reach 30% [88]. The only solution to minimize the level of yield reduction is immediate foliar fertilization [85,87].
Control of the Mg nutritional status of the plants grown on the farm is necessary in the early stages of plant growth. At the latest, it should be carried out just before the cardinal phase of yield component formation [87]. Symptoms of Mg deficiency in winter wheat, regardless of the supply of Nf, are more specific, but less destructive to the yield than a shortage of N, as shown for wheat in Figure 5a. The early symptoms of Mg deficiency on maize may significantly interfere with N uptake and, consequently, plant growth and yield (Figure 5b). The highest requirements of plants for Mg supply are mostly just before flowering, as has been documented for maize, winter oilseed rape, and winter wheat [83,85,89].

Figure 5.
Symptoms of magnesium deficiency: (a) wheat during the stem elongation phase; and (b) maize at the fifth leaf stage (photos by Witold Grzebisz).
Foliar fertilization of plants with Epsom salt (MgSO4 × 7H2O) in the growth stages preceding the critical phases of yield formation in seed plants significantly affected both grain density and grain protein content [46]. A recent study has clearly shown that foliar fertilization of winter wheat with Mg significantly increased the grain yield [85]. The achieved result was a consequence of the increase in GD, conditioned by a higher amount of N in wheat at the beginning of flowering. The same response has been observed for sugar beet fertilized with Epsom salt [86]. A farmer can expect such an effect, provided a well-balanced ratio of the content of both nutrients in the indicative parts of the plant. For example, in sugar beet, an N:Mg ratio of 7:1 can guarantee the maximum yield of storage roots [46], while a narrower or wider ratio will lead to decreased yield. It is necessary to assume that this ratio is species-specific [46,78]. Another significant interaction of Mg concerns its relationship with K and ammonium (NH4+). As has been explained by Grzebisz et al. [12], the relationship between the accumulation of Mg and WOSR yield is consistent with a quadratic regression model while, for K, the relationship is linear. This discrepancy suggests antagonism between these two elements. High content of K in a plant may interfere with the uptake and function of Mg [36,45]. The effect of NH4+ on the content of Mg2+ is one-way antagonistic [11]. Therefore, the use of only ammonium fertilizers as a source of N may disturb plant growth and yield.
4. Roles of Sulfur in Plant Crop Nutritional Homeostasis
4.1. Sulfur Uptake and Assimilation
The physiological functions of sulfur in plants are well-known and have been described in various academic textbooks [11]. The question is: to what extent can these functions be exploited by farmers to improve the yield formation by the grown crops? The plant demand for sulfur is variable during plant growth. Its uptake (Su) depends on two main factors. The first is the rate of plant growth, which can be expressed as the crop growth rate (CGR, kg ha−1 × day−1), and the second is the required content of S in the plant in the particular stage of growth (S, %; or g kg−1 DW):
A plant’s demand for S is not constant throughout its life-cycle. As shown in Figure 6, the highest rate of S accumulation by spring barley occurs in the early stages of grain yield formation (full flowering to the early milk stage). In this particular period, the amount of S in spring barley almost doubled. This also applies to crops with a much higher requirement for S, such as winter oilseed rape, which is one the most sensitive plants with respect to S supply. This crop showed the highest demand for S from the middle to the end of flowering [90]. It can, therefore, be concluded that the presented pattern of S accumulation is typical for all seed plants. Four questions should be formulated at this point, which are important for the efficient management of S in the currently grown crop by the farmer:
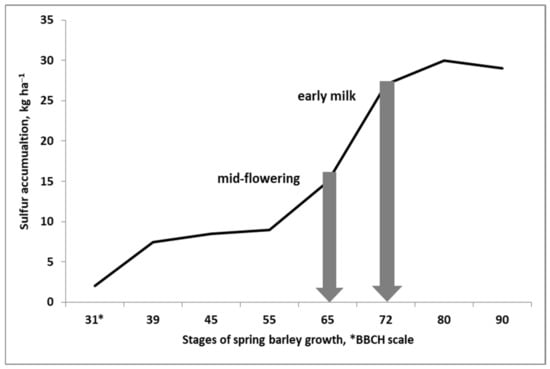
Figure 6.
The pattern of sulfur accumulation in spring barley during the growing season [Przygocka-Cyna, K., unpublished].
- (1)
- What is the source of S (or, where does the plant get the required amount of S from)?
- (2)
- Does the farmer have the operational measures to correct the nutritional status of S before the critical stages of yield formation?
- (3)
- To what extent does S deficiency:
- disturb the N economy of the plant?
- limit the formation of yield components and, ultimately, yield?
The optimal S content in winter oilseed rape just before flowering is in the range 0.55–0.65% DW. A content less than 0.35% DW indicates deep, visible S stress (Figure 7a). The range of 0.35–0.55% DW describes a hidden S deficiency, which is difficult to identify by the farmer but may result in significant yield reduction [90]. The critical S content in cereals is much lower, being in the range of 0.15–0.4% DW [91].

Figure 7.
Symptoms of sulfur deficiency: (a) winter oilseed rape at the inflorescence stage (sulfur pandemia); and (b) potato before flowering (photos by Witold Grzebisz).
The plant can use three chemical forms of S, including sulfate (SO42−), SO2, and H2S. Sulfate is taken up by the roots from the soil solution, and the other two compounds are obtained from the air. Sulfates are the dominant carrier of S for crops, and its direct source is the soil. The leaves are also an absorption site, for sulfates applied in water spraying. The whole process of S compound utilization by the plant is carried out in three main steps. The first (initial) involves the transportation of SO42− anions from the soil solution through the root apoplast to the xylem, and then to the leaf. The latter part is also the place of absorption of gaseous S forms. In the leaf, the arriving molecules, regardless of chemical form, undergo transformation into cysteine. The third step of S assimilation by the plant refers to re-translocation of S metabolites (e.g., glutathione) to the new vegetative organs or the developing fruits [92,93].
The first stage—that is, SO42− uptake from the soil solution—begins with its transfer across the root cell membrane. This process is mediated by membrane proteins called H+/sulfate transporters, representing the family of H+/SULTR transporters [93]. Sulfate uptake occurs as co-transport (symport) with protons (H +) in the formula 3H+/SO42−. This formula clearly indicates a huge requirement for metabolic energy by the plant to absorb S-SO4 from the soil solution. The activity of the proton pump (H+-ATPase) is required to acidify the root apoplast, creating favorable conditions for the transfer of SO42− to the root [94]. The active absorption of S is carried out in distinct parts of the root. Their activation and efficiency in SO42− uptake depends on the current nutritional state of S in the shoot. The deficiency of S due to the increased sensitivity of growing buds or leaves to S supply induces the activity of high-affinity sulfate transporters in the root (SULTR 1;1; SULTR 1;2) [92]. The most efficient absorption sites are located at the root tip and in the root hair zone. Under conditions of optimal S status in the plant, the uptake of sulfates is regulated by low-affinity transporters, which are located in the cortex cells of the root. The absorbed SO42− ions then move within the plasmodesmata (intercellular channels connected the root cells) to the xylem. Then, they move upward, in the transpiration water stream, to the leaves [95].
The efficiency of S uptake at each stage of growth requires a large input of plant energy to load and then unload SO42− carriers. Therefore, the efficiency of these processes is related to the ATP resources in the plant which, in turn, depend on the nutritional plant status, in terms of P and Mg. In the leaf, sulfate ions flowing from the xylem—depending on the content of S metabolites—are further translocated to the chloroplasts or the vacuole. The latter leaf organelle acts as temporal storage for sulfates. The three key steps of SO42− ion transformation in the chloroplasts include: (i) its activation to APS (adenosine 5′-phosphosulfate), followed by reduction to sulfite (SO32−); (ii) the reduction of sulfite to sulfide (SH−); and (iii) the synthesis of cysteine. This particular amino acid is the first stable product of S in the plant metabolic S cycle [96]. The reduction of sulfate to sulfite is determined by the supply of glutathione (GHS), which acts as an electron donor. The reduction of sulfite to sulfide is limited by the content of ferredoxin, which also acts as an electron donor [97]. Thus, the efficiency of SO42− fixation is limited by the nutritional status of the plant, in terms of S and Fe. The synthesis of cysteine is also directly controlled by the supply of carbon–nitrogen skeleton; that is, O-acetylserine (ASA) [98]. In summary, it can be concluded that synthesis of cysteine in the plant, based on sulfate as a substrate, is a process limited by the supply of five nutrients: N, P, Mg, Fe, and S.
The uptake of S by plants from the soil is regulated by systemic plant signals, which indicate the current state of the S content in the shoot. Four groups of S uptake control mechanisms have been distinguished [92,99]. The first is the content of sulfate (S–SO4), which is mobile both in the xylem and the phloem. The second group contains a set of S metabolites, including sulfite, cysteine, and glutathione. These organic compounds are regarded to be potential indicators of the S nutritional status in crop plants [96]. The third group includes the content (or, rather, the rate) of ASA production by the plant. An excess of S compounds from the first two groups, or deficiency in ASA in the leaf, reduces S-SO4 uptake from the soil. The fourth controlling mechanism is the amount of gaseous S compounds absorbed by the plant directly from the air. These compounds affect the main stages of sulfate transformation in the plant. By influencing the activity of APS reductase, they indirectly limit the uptake of sulfate ions from the soil [100].
Re-mobilization of S from adult plant organs, despite accumulation of sulfates in the vacuole, is low. The rate of S re-mobilization from vegetative plant tissues is generally too low, in relation to the needs of intensively growing new plant parts or seeds/grains [101]. In seed plants, the key reason for this (as shown in Figure 6) is a sudden, high increase in demand by the intensively growing reproductive plant organs. The first symptoms of S deficiency are not readily recognizable, and are often mistaken for N or Fe deficiency symptoms. Deficiency of S manifests mainly in the group of plants sensitive to this nutrient. Example crops are cruciferous plants (see Figure 7a). In less S-demanding crops, such as cereals or potato, the symptoms of S deficiency are often overlooked by the farmer (Figure 7b). The deficiency revealed under conditions of strong and long-lasting S shortages pose a great threat to crops with high demand for this nutrient. A classic example is oilseed rape (Figure 8a). The consequence of S deficiency for this crop is a yellowish–whitish color of the flowers, which are not very attractive to bees [102]. As such, a consequence of S deficiency is self-pollination, resulting in a low number of seeds per pod (see Figure 8b).

Figure 8.
Sulfur deficiency in winter oilseed rape: (a) at full flowering; and (b) at the stage of pod growth (photos by Dorota, Karolina Grzebisz and Witold Grzebisz).
Unlike N, S is not taken by crop plants in excess [103]. It is necessary to take into account the extremely important fact that the content of available S in the soil increases with the depth of the soil profile [104]. It has been recently documented that a deficiency of S does not inhibit the growth of primary roots [105]. However, the uptake of S can be seriously disturbed by several factors of varying origin [93,103,106,107,108,109]. These factors should be first considered, and then controlled by the farmer:
- (1)
- The low content of available S:
- sandy soil;
- low content of organic matter, as the key S source in arable soils.
An agronomic solution in such cases is to increase the content of humus and/or use sulfur fertilizers.
- (2)
- The limited access of roots to the soil available S pool, due to the reduced depth of the plant rooting zone:
- toxicity of Al3+;
- low content and supply of mineral nitrogen;
- soil compaction;
- excess water content.
Such threats to root growth require action by the farmer to eliminate them.
- (3)
- Agronomic:
- lack or insufficient dose of S fertilizers;
- too-high frequency of S-sensitive plants in the crop rotation;
- too-high biomass of harvest residues poor in S, resulting in immobilization of S-SO4.
The multi-functional effect of S on the crop—and especially on N management—requires systematic control of factors influencing the content of available S in the soil.
4.2. Sulfur Functions in Crop Plants
In plant production, S plays multiple roles. For the farmer, three groups are important [97,106,109,110,111,112]:
- (1)
- The Fe–S clusters, impacting the management of C and N:
- chlorophyll synthesis, photosynthesis;
- nitrogenase (N2 reduction);
- nitrate reductase (NO2− → NH3);
- sulfite reductase (SO32− → SH−)l
- synthesis of storage proteins, sulfur amino-acids (cysteine, cysteine, methionine), and components limiting the nutritional quality of harvested products.
- (2)
- Synthesis of S metabolites:
- cysteine, as a precursor of methionine, coenzyme A, glutathione (GSH), biotin, glucosinolates;
- derivatives of methionine, such as lignins, flavonoids, and ethylene.
- (3)
- Crop yield and quality of main product (seeds, grain, tubers, roots, fruits):
- increased yield of crops;
- increased harvest index of seed crops;
- enhanced content of gluten in wheat grain, leading to higher bread-making quality;
- increased synthesis of organo-sulfurous compounds (a classical example is allicin);
- biofortification of edible plant parts with essential micronutrients.
In plant production, S plays multiple functions; for example, as a nutrient or fungicide. One of its important nutritional functions is regulation of the N balance. Improving the N:S ratio results in better formation of yield components, higher yield, and higher protein content and quality in broad range of crops [107,113]. An increase in the N:S ratio leads to an increase in the content of low molecular N compounds. This state, caused mainly by S deficiency in crops, results in deterioration of the quality of crop products [114]. During the growing season, S deficiency concomitant with N excess creates a favorable environment for pathogenic attack. On the other hand, a balanced supply of N and S enhances synthesis of proteins, which results in lower pressure of pathogens [115].
The use of inorganic sulfur as a fungicide has been practiced for hundreds of years [116]. The protective functions of S in the plant are related to both the action of its carriers (inorganic fertilizers) and secondary S metabolites [117,118,119]. This broad spectrum of compounds involves, among others:
- (1)
- Mineral S compounds, such as hydrogen sulfide; elemental sulfur, sulfur dioxide, CS2, H2S, K2S, thiosulfates;
- (2)
- Phenols and polyphenols;
- (3)
- Lignins, as organic building materials of cell walls, providing a biophysical barrier in leaves and other plant-covering tissues against pathogens and insects;
- (4)
- Plant-specific S defense compounds, such as glutathione, glucosinolates, phytoalexins, S-rich proteins;
- (5)
- Plant hormones, such as ethylene and jasmonic acid.
Balanced fertilization of crops with S allows a double production effect to be reached. First, as mentioned in the previous section, it results in higher efficiency of applied Nf. Secondly, S increases the content of defense compounds in the plant, thereby increasing the level of defense to the attack by pathogens. This phenomenon is called sulfur-induced resistance (SIR) in crop plants [120]. The fungicidal effect of S is due to the presence of mineral and organic S compounds in the plant. A large part of the S absorbed by the plant is reduced to H2S or dimethyl sulfides, which are released into the atmosphere through the leaves. Their presence in the leaf phyllosphere reduces the pressure of pathogens [120]. An example is Leptosphaeria maculans, a pathogen causing dry rot of brassicas. The sensitivity of oilseed rape to this pathogen decreases with improvement of the S nutritional status of the plant. Plant hormones, such as salicylic acid (SA), jasmonic acid (JA), and ethylene (ETY) play important roles in transmitting the signal of infection. The synthesis of these organic signal substances increases in accordance with increased S status in the crop [121]. The inhibitory effect of S0 results directly from its toxicity, as well as the toxicity of its oxidized (SO2) or reduced (H2S) products. One of the effects of S0 is the inhibition of fungal spore germination [117,122]. It has been observed that the foliar fertilization of cereals with S0 significantly decreased the infestation of plants by powdery mildew and stem rust [119,123]. A low concentration of S in potato plants may result in increased sensitivity to pathogen pressure [124].
5. Secondary Nutrients and Nitrogen Fertilizer Productivity
5.1. Amelioration of Factors Limiting Nitrogen Uptake Use Efficiency
Understanding the importance of secondary nutrients in Nf productivity requires knowledge of their roles in the processes responsible for the uptake of inorganic N from the soil, as well as its subsequent utilization by the plant. The uptake of inorganic N by the currently grown plant depends on its content in the soil solution, as a necessary condition, and the presence of roots, as a sufficient condition. The first condition depends on a series of natural processes involved in mineralization of soil organic N, and as well as application of Nf [125,126].
The root system architecture (RSA), although encoded in the plant genome, is highly sensitive to environmental factors, including the content of nitrates (N–NO3). The ability of roots to penetrate the soil is driven by several factors, which interact with each other. The key set of factors includes: (i) physical conditions (soil texture and structure, compaction, spatial distribution of moisture); (ii) temperature; (iii) acidity; (iv) content of available nutrients; and (v) the nutritional status of the plant [104,127]. As a result of geochemical processes taking place in the soil, the content of nutrients in the soil profile with depth increases in the order P > K > Ca > Mg > Na = Cl = S-SO4 [128]. The solubility of secondary nutrients in water, which determines their availability to the plant root (an extended version of the necessary condition), depends on the content of water and soil pH [40]. The mobility of a given nutrient in the soil (the value of the effective diffusion coefficient) determines the current water content, which depends on precipitation [26,129]. The response of plant roots to the deficiency of Ca, Mg, and S is poorly understood. Studies on Arabidopsis thaliana have indicated a decrease in total plant biomass in response to reduced Mg supply, but the shoot to root ratio did not change much [130]. S deficiency increased the length of the primary root and density of lateral roots, but only at the root tip [131].
The destructive effect of Al3+ on plant architecture results from disturbances to the root system architecture (RSA), including plant rooting depth, root length density, and so on [132]. A reduced root system due to excessive content of Al3+ indirectly increases the risk of nitrate leaching [17]. The consequences of Al toxicity are very often related to shortages of Mg and Ca, resulting in plant dwarfism, poor formation of yield components and, finally, plant death [133]. The limit ranges for the content of Al3+ in arable soils are well-recognized [41]. Morphological changes in the RSA interfere not only with the uptake of mobile nutrients, such as nitrates, but even more with immobile ones (e.g., P and K) [134]. Therefore, the control of Al3+ activity is necessary in both humid and tropical world regions [133]. The plant production functions of Ca are, in fact, reduced by the impact of soil Al3+ control.
The role of Ca in the RSA for plant production results from its (direct and indirect) impact on root growth and proliferation. Its deficiency in the growth medium is manifested by a drastic reduction in the length of primary or seminal roots, along with enhanced proliferation of secondary roots [131]. Inhibition of root growth occurs immediately after a decrease in the supply of Ca ions to the plant [11]. The indirect effect of Ca deficiency is a result of Al3+ toxicity. A gradual increase in soil acidification, associated with increased content of Al3+ cations in the soil, leads to a reduction in root growth into the soil profile [74,135]. Consequently, the rooting zone is reduced, and a part of the applied Nf is leached into the sub-soil. Thus, the probability of uptake by the currently grown plant is diminished. Moreover, the uptake of S-SO4, Ca, and Mg is also disturbed, negatively affecting the N:S ratio in the plant. The deficiency of Mg negatively affects the production of energy carriers by the plant (ATP). The negative effects of these cumulative processes include not only the loss of plants from the field but, in turn, a loss of yield (Figure 9). As shown in Figure 9a, the excessive content of Al3+ in soil results in the potential leaching of nitrates. Barley, as a plant sensitive to Al3+ toxicity, can be used by the farmer as a plant indicator of soil acidity (Figure 9b). The final result of this series of processes is low efficiency of the used production measures, including Nf, as well as a threat posed to the environment.

Figure 9.
Response of crop plants to progressive gradient of soil acidity—a case considering winter barley. (a) reduced stems and roots along with increased content of toxic Al3; and (b) squares of plants falling out of the canopy due to Al3+ toxicity (photos by Witold Grzebisz).
The farmer’s corrective actions aimed at adjusting the current pH level of the soil to the level required for plants grown in a particular crop rotation are typically based on the use of lime or, alternatively, gypsum [136]. The expected production and environmental effects of these soil amendments result from secondary processes taking place in the soil. The neutralization of Al3+ toxicity, as well as the increased Ca resources in the soil, restores the conditions for the growth of plant roots in the soil [135]. At present, gypsum (CaSO4 × 2H2O) is used as a soil amendment in many countries, in the form of the FGD gypsum (flue gas desulfurization). The contents of Ca and S in this nutrient carrier are 23.3% and 18.6%, respectively. Gypsum does not de-acidify the soil, but neutralizes the activity of Al3+ [137]. The simplified control mechanisms—more specifically, regarding the exclusion of Al3+ from the soil solution—are detailed below:
The key factor affecting the release of both nutrients from gypsum is the concentration of their dissolution products in the water [138]. Assuming a naturally low content of SO42− ions in the soil solution, the key factor for gypsum dissolution is the amount of water in the soil. In rain-fed areas of plant production, the content of sulfates in the soil varies during the growing season. This state results from the cyclic release and precipitation of Ca2+ SO42− in the soil solution. The use of gypsum and/or FDG gypsum significantly affects the physical, chemical, and biological properties of arable soils. The re-generated soil structure creates conditions for efficient infiltration of water into the soil profile, as well as root ingrowth into the sub-soil. Thus, the increased soil volume exploited by plant roots leads to the stronger resistance of grown plants to abiotic stresses, maintaining higher yields [139,140]. The effects of using CaSO4 × 2H2O in agriculture, as expected by the farmer, are associated with an increase in the contents of Ca2+ and SO42− in the soil solution, along with a simultaneous decrease in the content of exchangeable Al3+ [141]. As shown in Figure 10, increased doses of agricultural gypsum (phosphogypsum), applied to highly acidic tropical soil, significantly increased the content of available sulfate. This is a necessary condition for fixation of exchangeable Al3+, thus creating favorable conditions for the growth of roots [142]. The second expected result of gypsum application is the increase (and, at the same time the change the percentage structure) of the cation exchange complex (CEC). In acid soil, the key geochemical aim of applied CaSO4 × 2H2O is to exchange the Al3+ present in the CEC with Ca2+ [137]. This effect is achievable both in sandy and clay soils. As shown in Figure 11, the CEC in sandy soil increases linearly in response to the dose of FGD. In clay soil, this trend was much lower and reached the saturation point at an FGD rate of 900 kg ha−1.
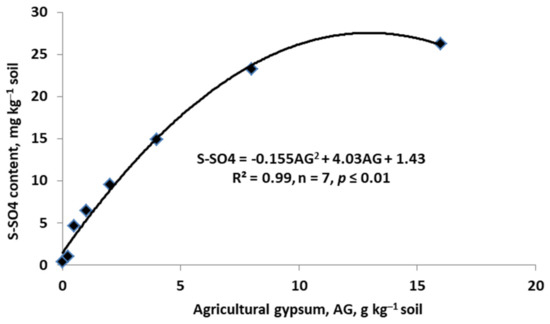
Figure 10.
Effect of agricultural gypsum on the content of available sulfate (based on Cuervo-Alzate and Osorio, [143]).
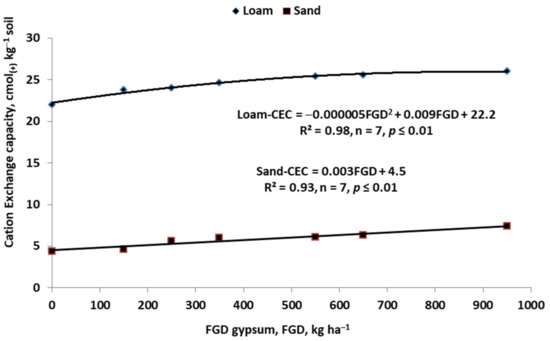
Figure 11.
Effect of FGD gypsum on cation exchange capacity of soil with contrastive texture (Grzebisz, W., unpublished).
Source of Ca in agriculture is carbonates and their derivatives—known in farming practice as lime [54]. The dissolution of calcium carbonates in water is low, much lower than gypsum [40]. In acidic soils, carbonates undergo hydrolysis, the products of which have a double function: delivering Ca2+, as well as neutralization of Al3+. Both of these processes are very complex but, in a simplified version, can be presented as follows:
- Calcium carbonate hydrolysis:
- Neutralization of toxic aluminum:
Neutralization of Al3+ with lime or gypsum is the primary stage in the restoration of soil productivity [54]. As has been reported by Carmeis Filho et al. [136], after 14 years of successive application of phosphogypsum, lime, or both fertilizers together to tropical soil, significant changes in the main chemical, biological, and physical properties in the soil profile were noted, to a depth of 60 cm. First, the content of exchangeable Al presented a decrease. Its content decreased the most in the topsoil (more than 5×), but also (slightly) in the deepest soil layer. The opposite trend was recorded for the content of exchangeable Ca. At the same time, the mass of roots of two successive crops in the cropping sequence (i.e., wheat and common bean) increased significantly. The largest relative increase in root mass was recorded in the sub-soil (more than 7×). Third, the total shoot dry weight of both crops increased by 250%. A primary, and extremely important effect of soil amendment application was the change in the structure of soil aggregates, including greater stability and increased soil porosity. The observed effect of liming in such a difficult environment for crop growth is typical of acidic soil. A significant increase in the content of exchangeable Al3+ was noted when the pH of the soil under wheat fell below pH 5.0.
The yield increase in response to lime application to acid soils is well-recognized in agriculture [41,54]. The main reason for such a yield increase is the improvement of N management. In seed plants such as wheat, the critical period of grain density (number of grains per unit area) formation takes place in the stages directly preceding flowering [144]. Thus, the effect of soil liming on the N economy in the currently grown plant is revealed before this phase. All of these processes can be divided into two main groups, affecting: (i) the rate of mineralization of soil organic N, and (ii) uptake of inorganic N. As shown in Figure 12, the liming of two acid soils, using carbonate rocks of different geological origin, resulted in an additional amount and uptake of N, consequently leading to yield increases. The annual maximum N uptake by plants grown on loamy sand slightly exceeded 32 kg ha−1. This value corresponds to 1.0 t ha−1 of wheat grain of high milling value [145,146]. This amount of N could potentially be leached from the soil, posing a threat to the environment.
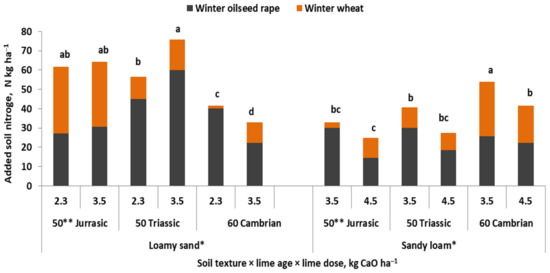
Figure 12.
The amount of nitrogen accumulated in the cropping sequence winter oilseed rape → winter wheat in response to the type and dose of lime of different geological origin (Grzebisz, W., unpublished). Legend: *, different fields; **, CaO content in the fertilizer; a–d similar letters denote a lack of significant difference using Tukey’s test.
5.2. Amelioration of Factors Limiting Nitrogen Utilization Use Efficiency
The effects of the interaction between secondary nutrients and nitrogen (i.e., for the pairs Mg × N and S × N, as well as Mg × S × N) is most reduced to the amount of extra yield [82,147,148]. This mode of analysis is both simple and useful, but only from a farm economic point of view. In fact, it is too general and insufficient to develop effective operational tools, mainly in terms of timing and dosage of a particular nutrient application. The basis for the effective application of Mg, S, or MgS fertilizers is the farmer’s knowledge regarding the critical stages of yield formation for the currently grown crops. However, it cannot be neglected that the basic causative factor of the yield is N; particularly the state of plant nutrition in terms of N [12]. In production practice, the use of S and Mg fertilizers is essentially based on three objectives. The first two are directly related to the formation of yield components. In seed plants, the yield is determined by the number of seeds/grains (SD, Seed Density; GD, Grain Density) and thousand seed/grain weight [148,149]. The third objective is to increase the content of N in seeds/grain, for which bread wheat is the classical example [146].
Körner’s [150] hypothesis assumes that yield is a function of the plant sink capacity. This means that the higher the number of seeds per plant per unit area (SD, GD), the higher the yield that can be expected. If the SD represents the sink capacity of the seed plant, the key goal of N application is maximization this yield component. The content of N in seeds/grains is a conservative plant trait [151]. Thus, the primary goal of secondary nutrient application is to increase SD. The increase in SD results in a greater mass of N in grain and, consequently, greater efficiency of N accumulated by the plant (i.e., nitrogen utilization efficiency; NUtE). Such a relationship has been noted for crops yielding at a very high level, such as winter oilseed rape and winter wheat [47,152,153]. Below, we present four equations developed for winter oilseed rape. They explain the above concept of NUtE dependence on secondary nutrient accumulation in a step-by-step manner [151]:
where: h and f denote harvest and flowering, respectively.
In the presented case, SD was limited by a shortage of Mg in seeds at harvest (Mgh). The critical phase of yield formation of winter oilseed rape extends from the formation of buds to full flowering [154]. The shortage of Mg—but not N—first manifests, as shown in Equation (8) (i.e., prior to flowering). The importance of Mg nutritional status in winter oilseed rape at this stage of growth has also been documented, by Szczepaniak et al. [155]. It is clear, from Equation (9), that the amount of Mg re-mobilized from the vegetative WOSR tissues during the seed filling period is not sufficient to cover the crop demand. Moreover, this set of equations indirectly informs us that seed plants should be treated with Mg before the beginning of the critical phase of yield component formation [46]. In cereals, as has been deeply documented for wheat, the booting/heading stages are crucial for grain density, which strongly depend on the supply of N to the growing ear [30,85]. The high—and, at the same time, more stable—increase in winter wheat yield can be secured by an adequate dose of Nf combined with Mg fertilizer application. The basis of this solution for winter wheat is soil-applied Kieserite and foliar fertilization using Epsom salt. The best stage in crop growth for this is the beginning of heading [46,151]. In this particular case, the NUtE linearly depends on the amount of Mg in winter wheat canopy at harvest:
The net grain yield increase under the dual method of winter wheat fertilization with Mg was 0.85–0.90 t ha−1 [85].
As has been documented by Maruyama-Nakashita et al. [93], the activity of sulfate transporters in the roots of Arabidopsis thaliana (SULTR1;1; SULTR1;2) depends on the relationship between the concentration of nitrates and sulfates in the growth medium. This relationship was indirectly confirmed in field experiments, in which the response of crop plants to doses of N and S was examined. As shown in Figure 13, the grain yield of maize during the grain-filling period showed a response to the interaction of N and S rates. This was manifested by a greater net accumulation of both nutrients by the crop. The partial factor productivity of Nf (PFP-Nf) indices for the plot fertilized with 120 kg N ha−1 were 116, 123, and 128 kg grain kg−1 Nf at S rates of 0, 8, and 16 kg ha−1, respectively. The same pattern of maize yield response to the interaction of both nutrients has been observed by Szulc et al. [156]. In this particular case, the maize yield on the S control plot was 7.62 t ha−1 at the optimal Nf rate of 111.1 kg ha−1. The application of 40 kg S ha−1 in the form of S0 resulted in a yield of 9.11 t ha−1, but the optimal Nf rate increased to 135 kg ha−1. The PFP-Nf index was almost the same for both treatments, amounting to 68.6 and 67.7 kg grain kg−1 Nf, respectively. In the discussed experiments, the key reason for the increase in yield was a higher Nf rate, which induced the action of the applied S fertilizer. Increased uptake of N in response to S application has also been observed for wheat. In the case of maize, the effect of applied S was revealed at higher Nf rates [157]. Based on the presented cases, despite their incompleteness (i.e., no data on yield components), it can be assumed that the increase in yield was the result of a higher GD or TGW. The increase in expression of both components was the result of better N supply to plants. Therefore, it can be concluded that the yield-forming effect of S may only become apparent when the resources of inorganic N in the soil are enriched with readily available Nf.
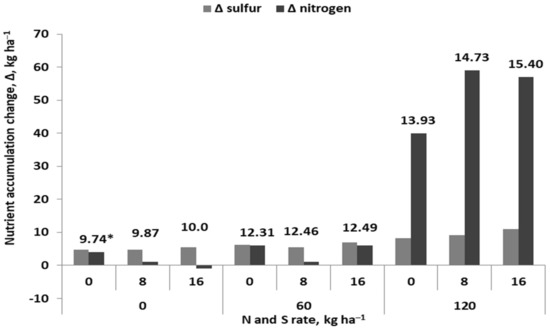
Figure 13.
Nitrogen and sulfur balance during the grain-filling period of maize growth and respective yield (based on Pagani et al., [158]); * grain yield.
The cases discussed above clearly indicate the need to fertilize the currently grown plant in such a way as to achieve the required constant relationship between the contents of N and S in the indicatory plant organ. Sulfur is not easily re-mobilized from older plant parts in response to its insufficient uptake from the soil. As a result, S deficiency disturbs N synthesis, manifested as the light green color of the young, but adult leaves; see Figure 7 and Figure 8. The appearance of visual symptoms of S deficiency indicates a loss of yield ([159], Figure 8). Despite the fact that the content of total S, sulfates, or glutathione reflects the S status in crop plants fairly well, the N:S ratio is most often used as an indicator of the expected relationship between both nutrients [103]. The indicative plant organs are young, fully developed leaves. The optimal date for the diagnosis of a plant’s nutritional status should precede the critical stage of yield formation. In cereals, this refers to upper leaves collected at the beginning of heading and, in oilseed rape, those at the beginning of budding [159]. In diagnostic practice, often the whole biomass of shoots is analyzed for the content of both nutrients [160]. The N:S range is a stable plant trait, but differs significantly between species. The N:S ratio is narrowest for Cruciferous plants, ranging from 5 to 7:1 [108]. The N:S ratio in cereals require in-depth discussion, and the published data ranges from 6 to 17: 1. The lowest ratio of 6–7:1 was recorded in Turkey for wheat [161]. The authors documented that increased doses of S resulted in narrowing of the N:S ratio, from eight on the S control to around five to plants fertilized with 80 kg S ha−1. However, for two tested wheat cultivars, the best N:S ratio was in the range of 6–7:1. The yield increase was due to an increase in yield components (i.e., the number of grains per ear and TGW). The same trend was observed for malt barley. The application of 50 kg S ha−1 at BBCH 37, in the form of S0, raised the S content to 0.4% in leaves, resulting in an N:S ratio of 8:1 and maximum yield of 5.0 t ha−1. This N:S ratio allowed the grain to obtain good technological parameters [162]. The most often published data on the N:S ratio for wheat is around 14–17:1 [163,164]. As reported by ReussiCalvo et al. [164], wheat yield response to applied S has been recorded only for plots fertilized with Nf. A narrower value of this index was recently documented by Sedlar et al. [165]. This study showed that the sulfur nutrition index of 1.0, resulting in an N:S ratio around 9.0 at BCCH 30, and 8.0 at BBCH 45 and 51, was optimal, with yield ranging from 7.0 to 11 t ha−1. The N:S index can be also used to determine the required S fertilizer dose; for example, an Nf of 160 kg N ha−1 is often recommended in Poland for winter wheat and winter oilseed rape [58], which should be balanced with S applied at a rate of 32 and 10 kg ha−1, respectively.
In agricultural practice, the carriers of Mg and S are also their sulfates, represented by Kieserite and Epsom Salt. The first fertilizer is applied to the soil, while the second is used for foliar fertilization [78]. Therefore, the yield-forming effect of N can be supported by the simultaneous action of both nutrients. The application of MgS to crop plants just before critical stages of yield formation results in higher recovery of N from applied fertilizer, uptake of nitrogen, and yield. This assumption has been documented for winter wheat, spring wheat, corn, and oilseed rape [36,42,83,106,166,167]. This response of crops to MgS application can be explained by the higher seed/grain density in seed plants. A classic example is shown in Figure 14. The PFP-Nf indices were significantly (as expected) lower on the plot fertilized with higher Nf doses. Two facts are important for agricultural practice. The first is the significant impact of S on Nf productivity, especially on plots with lower Nf rate. As for the second, yields on the plot fertilized with both Nf and MgS were the same as on the plot fertilized only with N, but applied in much higher dose. It can therefore be concluded that a lower dose of Nf supported with MgS fertilizer may lead to both lower production costs and a lower potential threat to the environment.
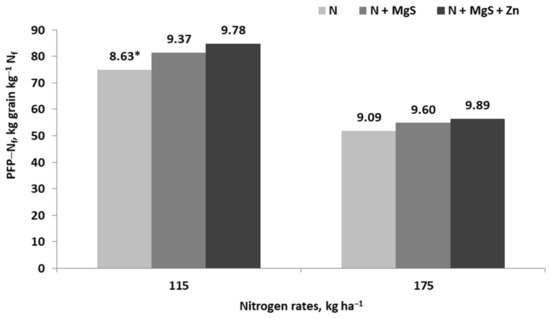
Figure 14.
Effect of magnesium sulfate application on values of the partial factor productivity of fertilizer nitrogen in maize (based on Potarzycki, [83]). *, grain yield, t ha−1.
The full list of Ca, Mg and S fertilizers is much wider. The classic source of Mg are carbonates, including dolomite or serpentynite [78]. A new carrier of Mg is struvite (MgNH4PO4 × 6H2O), a product recovered from wastewater effluents (urine). Despite its poor solubility, it can be considered as a potential future source of P and Mg for crops [168]. A well-known but quite new source of the nutrients discussed in this article is Polysulfate fertilizers. Their natural source is polyhalite (K2Ca2Mg(SO4)4 × 2H2O), deposits of which have been discovered in many countries, including Poland [169].
6. Conclusions
The effective use of Nf by a growing crop depends on its efficient uptake from the soil, as well as its subsequent efficient transformation in the plant. The farmer must remember that the yield is a result of the formation of the yield components, which, in turn, is affected by the supply of N. The uptake of N is limited by the availability of calcium, beginning from the initial stages of plant growth. The key competitor of Ca in the soil is toxic aluminum (Al3+), which limits root growth. This limitation can be effectively controlled by Al3+ neutralization using lime or gypsum. Both soil amendments affect a wide spectrum of soil properties, determining both the pool of inorganic N and its uptake by plants, as well as that in those grown in subsequent years (crop rotation). The excessive uptake of calcium, which generally occurs in areas where drought is frequent, can significantly reduce N uptake by plants, adversely affecting the formation of yield components and, consequently, yield. Magnesium can be treated, in agricultural practice, as a nutrient necessary for the plant throughout its life-cycle. Its deficiency should be first mitigated through soil-applied fertilizer. The foliar application of magnesium can significantly control calcium, provided that is carried out before the critical stages of yield formation in the crop. Deficiencies of sulfur and magnesium are most manifested at the beginning of the reproductive phase of seed plants. This indicates that the critical requirements for both nutrients occur just before critical stages of yield formation. Thus, this stage (or stages) of plant growth defines the timing of plant sampling and determination of its nutritional status in terms of N, Mg, and S. The application of Mg, S, or MgS, in order to reduce the ratio of N:Mg and/or N:S, creates conditions for higher productivity of Nf. Carriers of both nutrients, applied to the soil, typically show a residual effect, lasting for several years. It should be emphasized that good management of secondary nutrients means not only an increase in efficiency of Nf, but also a real reduction in production costs, while decreasing the potential threat of excess nitrates to the environment.
Author Contributions
Conceptualization, W.G. and W.Z.; methodology, W.Z.; software, K.P.-C.; validation, W.G., W.Z. and K.P.-C.; formal analysis, K.P.-C.; investigation, W.Z.; resources, W.Z.; data curation, K.P.-C.; writing—original draft preparation, W.Z. and K.P.-C.; writing—review and editing, W.G.; visualization, W.Z.; supervision, W.G.; project administration, K.P.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Figure A1.
Calcium accumulation and partitioning in by winter wheat in response to increasing N doses. *, Calcium harvest index (based on Nowicki, [35]).

Figure A2.
Magnesium accumulation and partitioning in by winter wheat in response to increasing N doses. *, Magnesium harvest index (based on Nowicki, [35]).

Figure A3.
Trends in the content of calcium in sugar beet plants during the growing season. (Grzebisz, W., unpublished). Legend: Ca-LE, Ca-SR, leaves and storage roots, respectively.
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Conijn, J.G.; Bindraban, P.S.; Schröder, J.J.; Jongschaap, R.E.E. Can our global food system meet food demand within planetary boundries? Agric. Ecosys. Environ. 2018, 251, 244–256. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Almaraz, M.; Aneja, V.; Austin, V.T.; Bai, E.; Cassman, K.G.; Compton, J.E.; Davidson, E.A.; Erisman, J.W.; Galloway, J.N.; et al. A world co-benefits: Solving the global nitrogen challenge. Earth’s Future 2019, 7, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Röös, E.; Bajželj, B.; Smith, P.; Patel, M.; Little, D.; Garnett, T. Greedy or needy? Land use and climate impacts of food in 2050 under different livestock futures. Glob. Environ. Chang. 2017, 47, 1–12. [Google Scholar] [CrossRef]
- Cirera, X.; Masset, E. Income distribution trends and future food demand. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2821–2834. [Google Scholar] [CrossRef]
- Mapes, B.R.; Prager, S.D.; Bene, C.; Gonzalez, C.E. Healthy and sustainable diets from today to 2050–the role of international trade. PLoS ONE 2022, 17, e0264729. [Google Scholar] [CrossRef]
- Taiz, L. Agriculture, plant physiology, and human population growth: Past, present, and future. Theor. Exp. Plant Physiol. 2013, 25, 167–181. [Google Scholar] [CrossRef]
- Omara, P.; Aula, L.; Oyebiyi, F.; Raun, W.R. World cereal nitrogen use efficiency trends: Review and current knowledge. Agrosys. Geosci. Environ. 2019, 2, 1–8. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Elsevier Ltd., Academic Press: London, UK, 1995; 899p. [Google Scholar]
- Grzebisz, W.; Diatta, J.; Barłóg, P.; Biber, M.; Potarzycki, J.; Łukowiak, R.; Przygocka-Cyna, K.; Szczepaniak, W. Soil fertility clock–crop rotation as a paradigm in nitrogen fertilizer productivity control. Plants 2022, 11, 2841. [Google Scholar] [CrossRef]
- Weih, M.; Hamner, K.; Pourazari, F. Analyzing plant nutrient uptake and utilization efficiencies: Comparison between crops and approaches. Plant Soil 2018, 430, 7–21. [Google Scholar] [CrossRef]
- Pandey, N. Role of Plant Nutrients in Plant Growth and Physiology. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 51–93. [Google Scholar]
- Lynch, J.P.; Wojciechowski, T. Opportunities and challenges in the subsoil: Pathways to deeper rooted crops. J. Exp. Bot. 2015, 66, 2199–2210. [Google Scholar] [CrossRef]
- Kell, D.B. Breeding crop plants with deep roots: Their role in sustainable carbon, nutrient and water sequestration. Ann. Bot. 2011, 108, 407–418. [Google Scholar] [CrossRef]
- Lynch, J.L. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef]
- Stewart, W.M.; Roberts, T.L. Food security and the role of fertilizer in supporting it. Procedia Engine 2012, 46, 76–82. [Google Scholar] [CrossRef]
- Kahabka, J.E.; van Es, H.M.; McClenahan, E.J.; Cox, W.J. Spatial analysis of maize response to nitrogen fertilizer in Central New York. Precision Agric. 2004, 5, 463–476. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bali, S.K. A review of methods to improve nitrogen use efficiency in agriculture. Sustainability 2018, 10, 51. [Google Scholar] [CrossRef]
- Song, L.; Wang, S.; Ye, W. Establishment and application of critical nitrogen dilution curves for rice based on leaf dry matter. Agronomy 2020, 10, 367. [Google Scholar] [CrossRef]
- FAO. Impact of the Ukraine-Russia Conflict on Global Food Security and Related Matters under the Mandate of the Food and Agriculture Organization of the United Nation (FAO). CL 170/6. May 2022. Available online: https://www.fao.org/3/nj164en/nj164en.pdf (accessed on 20 August 2022).
- Strubenhoff, H. The War in Ukraine Triggered a Global Food Shortage. 2022. Available online: www.brookings.edu/blog/future-development/2022/06/14/the-war-in-ukraine-triggered-a-global-food-shortage/ (accessed on 20 August 2022).
- Plant Health Association California. Western Fertilizer Handbook, 10th ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2022; 371p. [Google Scholar]
- Reid, R.; Hayes, J. Mechanisms and control of nutrient uptake by plants. Inter. Rev. Cytol. 2003, 229, 73–114. [Google Scholar]
- Clarkson, D.T. Nutrient interception and transport by root system. In Physiological Processes Limiting Plant Productivity; Johnson, C.B., Ed.; Butterworths: London, UK, 2013; pp. 307–330. [Google Scholar]
- Quintana, J.M.; Harrisosn, H.C.; Nienhuis, J.; Palta, J.P.; Kmiecik, K. Differences in pod calcium concentration in eight snap bean and dry bean cultivars. HortScience 1999, 34, 932–934. [Google Scholar] [CrossRef]
- Zhao, F.J.; Evans, E.J.; Bilsborrow, P.E.; Syers, J.K. Sulphur uptake and redistribution in double and single low varieties of oilseed rape (Brassica napus L.). Plant Soil 1993, 150, 69–76. [Google Scholar] [CrossRef]
- Grzebisz, W.; Łukowiak, R. Nitrogen gap amelioration is a core for sustainable intensification of agriculture—A concept. Agronomy 2021, 11, 419. [Google Scholar] [CrossRef]
- Meng, Q.; Yue, S.; Chen, X.; Cui, Z.; Ye, Y.; Ma, W.; Tong, Y.; Zhang, F. Understanding dry matter and nitrogen accumulation with time-course for high-yielding wheat production in China. PLoS ONE 2013, 8, e68783. [Google Scholar] [CrossRef] [PubMed]
- Van Noordwijk, M.; Cadisch, G. Access and excess problems in soil nutrition. Plant Soil 2002, 247, 25–40. [Google Scholar] [CrossRef]
- Blecharczyk, A. The reaction of winter rye and spring barley to the plant crop rotation system and fertilization in a long-term experiment. Rocz. Akad. Rol. Pozn. 2002, 236, 128, (In Polish with English summary). [Google Scholar]
- Grzebisz, W. Effect of soil fertility on roots dynamics of winter rye during the growing season. In Root Ecology and its Practical Application; Kutschera., L., Hubl, E., Lichtenegger, E., Person, H., Sobotik, M., Eds.; University of Natural Resources and Life Sciences: Wien, Austria, 1992; pp. 385–388. [Google Scholar]
- Meier, U. BBCH Monograph. In Growth Stages of Mono- and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Center for Agriculture and Forestry: Berlin, Germany, 2001; Available online: http://www.jki.bund.de/fileadmin/dam_uploads/_veroeff/bbch/BBCH-Skala_Englisch.pdf (accessed on 14 November 2021).
- Nowicki, B. The Impact of Micronutrient Supplementary Foliar Feeding of Winter Wheat and Fungicide Protection on the Nitrogen Balance of Plants in Critical Stages of Crop Formation; Manuscript (in Polish). Library of the Poznan University of Life Sciences: Poznan, Poland, 2020; 254p. [Google Scholar]
- Szczepaniak, W.; Grzebisz, W.; Potarzycki, J.; Łukowiak, R.; Przygocka-Cyna, K. The magnesium and calcium mineral status of maize at physiological maturity as a tool for an evaluation of yield forming conditions. J. Elem. 2015, 21, 881–897. [Google Scholar] [CrossRef]
- Stepaniuk, M.; Głowacka, A. Yield of winter oilseed rape (Brassica napus L. var. napus) in a short−term monoculture and the macronutrient accumulation in relation to the dose and method of sulfur application. Agronomy 2021, 12, 68. [Google Scholar] [CrossRef]
- Wallace, A.; Mueller, R.T. Calcium uptake and distribution in plants. J. Plant Nutr. 1980, 2, 247–256. [Google Scholar] [CrossRef]
- De, M.; Moore, A.D.; Mikkelsen, R.L. In-season accumulation and partitioning of macronutrients and micronutrients in irrigated sugar beet production. J. Sugar Beet Res. 2019, 56, 54–78. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 15th ed.; Pearson Education: Boston, MA, USA, 2017; 1071p. [Google Scholar]
- Barłóg, P.; Grzebisz, W.; Łukowiak, R. Fertilizers and fertilization strategies mitigating soil factors constraining efficiency of nitrogen in plant production. Plants 2022, 11, 1855. [Google Scholar] [CrossRef]
- Grzebisz, W.; Barłog, P.; Kryszak, J.; Łukowiak, R. Pre-anthesis nutritional status of spelt wheat as a toll for predicting the attainable grain yield. Agronomy 2019, 9, 558. [Google Scholar] [CrossRef]
- Grzebisz, W.; Frąckowiak, K.; Potarzycki, J.; Diatta, J.; Szczepaniak, W. The unexploited potential of nutrient analysis at the onset of tuberization for tuber yield production. Agronomy 2020, 10, 103. [Google Scholar] [CrossRef]
- Potarzycki, J.; Grzebisz, W. The in-season variability in the calcium concentration in potato organs and its relationships with the tuber yield. J. Elem. 2020, 25, 107–124. [Google Scholar] [CrossRef]
- Xie, K.; Cakmak, I.; Wang, I.; Zhang, F.; Guo, S. Synergistic and antagonistic interactions between potassium and magnesium in higher plants. Crop J. 2021, 9, 249–256. [Google Scholar] [CrossRef]
- Grzebisz, W. Crop response to magnesium fertilization as affected by nitrogen supply. Plant Soil 2013, 368, 23–39. [Google Scholar] [CrossRef]
- Potarzycki, J.; Grzebisz, W.; Szczepaniak, W. Magnesium fertilization increases nitrogen use efficiency in winter wheat (Triticum aestivum L.). Plants 2022, 11, 2600. [Google Scholar] [CrossRef]
- Grzebisz, W.; Szczepaniak, W.; Łaczny, S.; Łański, M.; Zawieja, A. Indices of total and unit nutrient uptake by high yielding winter oilseed rape under conditions of intensive magnesium fertilization. Fragm. Agron. 2018, 35, 23–36. [Google Scholar]
- Heckman, J.R.; Sims, J.T.; Beegle, D.B.; Coale, F.J.; Herbert, S.J.; Bruulsema, T.W.; Bamka, W.J. Nutrient removal by corn grain harvest. Agron. J. 2003, 95, 587–591. [Google Scholar] [CrossRef]
- Grzebisz, W. Technology of Crop Plants Fertilization–Yield Physiology. Part II. Cereals and Maize; PWRiL: Poznań, Poland, 2012; 279p. (In Polish) [Google Scholar]
- Costa Crusciol, C.A.; Fernandes, A.M.; de Almeida Carmeis Filho, A.C.; de Cassia Felix Alvarez, R. Macronutrient uptake and removal by upland rice cultivars with different plant architecture. Rev. Bras. Cienc. Solo 2016, 40, e0150115. [Google Scholar] [CrossRef]
- Bender, R.R.; Haegele, J.W.; Ruffo, M.L.; Below, F.E. Nutrient uptake, partitioning, and remobilization in modern transgenic insect-protected maize hybrids. Agron J. 2013, 105, 161–170. [Google Scholar] [CrossRef]
- Hirzel, J.; Undurraga, P. Nutritional Management of cereals cropped under irrigation conditions. In Crop Production; Goyal, A., Asil, M., Eds.; InTech: London, UK, 2013; pp. 99–130. [Google Scholar]
- Jones, J.B., Jr. Agronomic Handbook; CRC Press LLC: Boca Raton, FL, USA, 2003; 450p. [Google Scholar]
- Neto, M.E.; Moreira Lara, L.; de Oliveira, S.M.; dos Santos, R.F.; Braccini, A.L.; Inou, T.T.; Batista, M.A. Nutrient removal by grain in modern soybean varieties. Front. Plant Sci. 2021, 12, 615019. [Google Scholar] [CrossRef] [PubMed]
- Canadian Fertilizer Institute. Nutrient Uptake and Removal by Field Crops Western Canada; Canadian Fertilizer Institute: Ottawa, Canada, 2001; 10p. [Google Scholar]
- Aulakh, M.S. Content and uptake of nutrients by pulses and oilseed crops. Ind. J. Ecol. 1985, 12, 238–242. [Google Scholar]
- Grzebisz, W. Technology of Crop Plants Fertilization–Yield Physiology. Part II. Oil, Roots, Legumes; PWRiL: Poznań, Poland, 2011; 414p. (In Polish) [Google Scholar]
- Westermann, D.T. Nutritional requirements of potatoes. Am. J. Potato Res. 2005, 82, 301–307. [Google Scholar] [CrossRef]
- International Fertilizer Industry Association. World Fertilizer Use Manual; IFA: Paris, France, 1992; 632p. [Google Scholar]
- Knez, M.; Stangoulis, C.R. Calcium bio-fortification of crops-challenges and projected benefits. Front. Plant Sci. 2021, 12, 669053. [Google Scholar] [CrossRef] [PubMed]
- Barłóg, P.; Grzebisz, W.; Diatta, J.B. Effect of timing and nitrogen fertilizers on nutrients content and uptake by winter oilseed rape. Part II. Dynamics of nutrient uptake. Chem. Agric. 2005, 6, 113–123. [Google Scholar]
- White, P.J. Calacium. In Handbook of Plant Nutrition, 2nd ed.; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press, Taylor and Francis Group: Bocan Raton, FL, USA, 2015; pp. 165–198. [Google Scholar]
- Thor, K. Calcium–Nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- White, P.J. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Saito, S.; Uozumu, N. Calcium-regulated phosphorylation systems controlling uptake and balance of plant nutrients. Front. Plant Sci. 2020, 11, 44. [Google Scholar] [CrossRef]
- Huang, S.; Waadt, R.; Nuhkat, M.; Kollist, H.; Hedrich, R.; Roelfsema, M.R. Calcium signals in guard cells enhance the efficiency by abscisic acid triggers stomatal closure. New Phytol. 2019, 224, 177–187. [Google Scholar] [CrossRef]
- Somssich, I.; Hahlbrock, K. 1998. Pathogen defense in plants—a paradigm of biological complexity. Trends Plant Sci. 1998, 3, 86–90. [Google Scholar] [CrossRef]
- Lecourieux, D.; Ranyeva, R.; Pugin, A. Calcium in plant defense-signalling pathways. New. Phytol. 2006, 171, 249–269. [Google Scholar] [CrossRef]
- Granqvist, E.; Sun, J.; Op den camp, R.; Pujic, P.; Hill, L.; Normand, P.; Morris, R.J.; Downie, J.A.; Geurts, R.; Oldroyd, G.E.D. Bacterial-induced calcium oscillations are common to nitrogen-fixing associations of noduling legumes and non-legumes. New Phytol. 2015, 207, 551–558. [Google Scholar] [CrossRef]
- Smith, G.; Swart, S.; Lugtenberg, B.J.J.; Kijne, J.W. Molecular mechanisms of attachment of Rhizobium bacteria to plant roots. Mol. Microbiol. 1992, 6, 2897–2903. [Google Scholar] [CrossRef]
- De Freitas, S.T.; Talamani Do Amarante, C.V.; Mitcham, E.J. Calcium disorders in plants. In Postharvest Ripening Physiology of Crops; Pareek, S., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 477–512. [Google Scholar]
- White, P. The pathways of calcium movement to the xylem. J. Exp. Bot. 2001, 52, 891–899. [Google Scholar] [CrossRef]
- Marschner, H. 1991. Mechanism of adaptation of plants to acid soils. Plant Soil 1991, 134, 1–20. [Google Scholar] [CrossRef]
- Gerendás, J.; Führs, H. The significance of magnesium for crop quality. Plant Soil 2013, 368, 101–128. [Google Scholar] [CrossRef]
- Fan, M.-S.; Zhao, F.-J.; Fairweather-tait, S.; Poulton, P.; Dunham, S.; McGrath, S. Evidence of decreasing mineral density in wheat grain over the last 160 years. J. Trace Elements Med. Biol. 2008, 22, 315–324. [Google Scholar] [CrossRef]
- Davis, D. Declining fruit and vegetable nutrient composition: What is the evidence? HortSci. 2009, 44, 15–19. [Google Scholar] [CrossRef]
- Grzebisz, W. Magnesium. In Handbook od Plant Nutrition, 2nd ed.; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2015; pp. 199–260. [Google Scholar]
- Shaul, O. Magnesium transport and function in plants: The tip of the iceberg. Biometals 2002, 15, 309–323. [Google Scholar] [CrossRef]
- Weih, M.; Liu, H.; Colombi, T.; Keller, T.; Jäck, O.; Vallenback, P.; Westerbergh, A. Evidence for magnesium–phosphorus synergism and co-limitation of grain yield in wheat agriculture. Sci. Rep. 2021, 11, 9012. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, A.H.; Nayab, S.; Hussainn, S.B.; Ali, M.; Pan, Z. Current understanding on magnesium deficiency and future outlooks for sustainable agriculture. Int. J. Mol. Sci. 2021, 22, 1819. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hassan, M.U.; Nadeem, F.; Wu, L.; Zhang, F.; Li, X. Magnesium fertilization improves crop yield in most production systems: A meta–analysis. Front. Plant Sci. 2020, 10, 1727. [Google Scholar] [CrossRef] [PubMed]
- Potarzycki, J. Effect of magnesium or zinc supplementation at the background of nitrogen rate on nitrogen management by maize canopy cultivated in monoculture. Plant Soil Environ. 2011, 57, 19–25. [Google Scholar] [CrossRef]
- Szczepaniak, W.; Barłóg, P.; Łukowiak, R.; Przygocka-Cyna, K. Effect of balanced nitrogen fertilization in four-year rotation on plant productivity. J. Central Europ. Agric. 2013, 14, 64–77. [Google Scholar] [CrossRef]
- Grzebisz, W.; Potarzycki, J. Effect of magnesium fertilization systems on grain yield formation by winter wheat (Triticum aestivum L.) during the grain filling period. Agronomy 2022, 12, 12. [Google Scholar] [CrossRef]
- Pogłodziński, R.; Barłóg, P.; Grzebisz, W. Effect of nitrogen and magnesium sulfate application on sugar beet yield and quality. Plant Soil Environ. 2021, 67, 507–517. [Google Scholar] [CrossRef]
- Grzebisz, W.; Przygocka-Cyna, K.; Szczepaniak, W.; Diatta, J.; Potarzycki, J. Magnesium as a nutritional tool of nitrogen efficient management—Pant production and environment. J. Elem. 2010, 15, 771–788. [Google Scholar]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef]
- Szczepaniak, W.; Grzebisz, W.; Potarzycki, J.; Łukowiak, R.; Przygocka-Cyna, K. Nutritional status of winter oilseed rape in cardinal stages of growth as yield indicator. Plant Soil Environ. 2015, 61, 291–296. [Google Scholar] [CrossRef]
- Schnug, E.; Haneklaus, S. Sulfur deficiency in Brassica. Lanbauforschung Völkenrode 1994, 144, 31. [Google Scholar]
- Grant, C.; Hawkesford, M.J. Sulfur. In Handbook od Plant Nutrition, 2nd ed.; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2015; pp. 261–304. [Google Scholar]
- Takahashi, H. Sulfate transport systems in plant: Functional diversity and molecular mechanisms underlying regulatory coordination. J. Exp. Bot. 2019, 70, 4075–4087. [Google Scholar] [CrossRef]
- Maruyama-Nakashita, A.; Nakamura, Y.; Yamaya, T.; Takahashi, H. Regulation of high-affinity sulphate transporters in plants: Towards systematic analysis of Sulphur signaling and regulation. J. Exp. Bot. 2004, 55, 1843–1849. [Google Scholar] [CrossRef]
- Sondergaard, T.E.; Schulz, A.; Palmgren, M.G. Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase. Plant Physiol. 2004, 136, 2475–2482. [Google Scholar] [CrossRef]
- Buchner, P.; Takahashi, H.; Hawkesford, M.J. Plant sulphate transporters; co-ordination of uptake, intracellular and long-distance transport. J. Exp. Bot. 2004, 55, 1765–1773. [Google Scholar] [CrossRef]
- De Kok, L.J.; Castro, A.; Durenkamp, M.; Koralewska, A.; Posthumus, F.S.; Stuiver, C.E.E.; Ynag, L.; Stulen, I. Pathways of plant sulfur uptake and metabolism—An overview. Landbauforsch. Volkenrode 2005, 282, 5–13. [Google Scholar]
- Lu, Y. Assembly and transfer of iron-sulfur clusters in the plastid. Front. Plant Sci. 2018, 9, 336. [Google Scholar] [CrossRef]
- Saito, K. Sulfur assimilatory metabolism. The long and smelling road. Plant Physiol. 2004, 136, 2443–2450. [Google Scholar] [CrossRef]
- Hawkesford, M.J. Plant responses to Sulphur deficiency and the genetic manipulation of sulphate transporters to improve S-Utilization efficiency. J. Exp. Bot. 2000, 51, 131–138. [Google Scholar] [CrossRef]
- Davidian, J.C.; Kopriva, S. Regulation of sulfate uptake and assimilation—the same or not the same. Plant 2010, 392, 314–325. [Google Scholar] [CrossRef]
- Blake-Kalff, M.M.A.; Harrison, K.R.; Hawkesford, M.J.; Zhao, F.J.; McGrath, S.P. Distribution of sulfur within oilseed rape leaves in response to sulfur deficiency during vegetative growth. Plant Physiol. 1998, 118, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Haneklaus, S.; Brauer, A.; Bloem, E.; Schnug, E. Relationship between sulfur deficiency in oilseed rape (Brassica napus L.) and its attractiveness for honeybees. Landbauforschung Völkenrode Sonderheft 2004, 283, 24–37. [Google Scholar]
- Scherer, H.W. Sulphur in crop production—invited paper. Europ. J. Agron. 2001, 14, 81–111. [Google Scholar] [CrossRef]
- Giehl, R.F.H.; von Wirén, N. Root nutrient foraging. Plant Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, Z.; Amtmann, A. Food for thought: How nutrients regulate root system architecture. Curr. Opin. Plant Biol. 2017, 39, 8087. [Google Scholar] [CrossRef]
- Klikocka, H.; Marks, M. Sulphur and nitrogen fertilization as a potential means of agronomic biofortification to improve the content and uptake of microelements in spring wheat grain DM. J. Chem. 2018, 2018, 9326820. [Google Scholar] [CrossRef]
- Kulczycki, G. The effect of elemental sulfur fertilization on plant yields and soil properties. Adv. Agron. 2021, 167, 105–182. [Google Scholar]
- Zenda, T.; Liu, S.; Dong, A.; Duan, H. Revising Sulphur–the once neglected nutrient: It’s role in plant growth, metabolism, stress tolerance and crop production. Agriculture 2021, 11, 626. [Google Scholar] [CrossRef]
- Narayan, O.M.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 2022, e2030082-3. [Google Scholar] [CrossRef]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. 2006, 1112, 3–22. [Google Scholar] [CrossRef]
- Filipek-Mazur, B.; Tabak, M.; Gorczyca, O.; Lisowska, A. Effect of sulfur-containing fertilizers on the qunattity and quality of spring oilseed rape and winter wheat yield. J. Elem. 2019, 24, 1383–1394. [Google Scholar]
- Chorianopoulou, S.N.; Bouranis, D.L. The role of sulfur in agronomic biofortification with essesntial micronutrients. Plants 2022, 11, 1979. [Google Scholar] [CrossRef]
- Burkitbayev, M.; Bachilova, N.; Kurmanbayeva, M.; Tolnova, K.; Yerezhepova, N.; Zhumagul, M.; Mamurova, A.; Turysbek, B.; Demeu, G. Effect of sulfur-containing agrochemicals on growth, yield, and protein content of soybeans (Glycine max (L.) Merr). Saudi J. Biol. Sci. 2021, 28, 891–900. [Google Scholar] [CrossRef]
- Jamal, A.; Moon, Y.-S.; Abdin, M.Z. Sulphur—a general overview and interaction with nitrogen. Aust. J. Crop Sci. 2010, 4, 523–529. [Google Scholar]
- Datnoff, L.E.; Wade, H.E.; Huber, D.M. Mineral Nutrition and Plant Disease; Amer Phytopathological Society: Saint Paul, MN, USA, 2007; 278p. [Google Scholar]
- Tweedy, B.G. Inorganic sulfur as a fungicide. Residue Rev. 1981, 78, 43–68. [Google Scholar]
- Cooper, R.M.; Williams, J.S. Elemental sulfur as an induced antifungal substance in plant defense. J. Exp. Bot. 2004, 55, 1947–1953. [Google Scholar] [CrossRef]
- Nwachukwu, I.D.; Slusarenko, A.J.; Gruhlke, M.C.H. Sulfur and sulfur compounds in plant defense. Nat. Prod. Comm. 2012, 7, 395–400. [Google Scholar]
- Künstler, A.; Gullner, G.; Ádám, A.L.; Nagy, J.K.; Király, L. The versatile roles of sulfur-containing biomolecules in plant defense-A road to disease resistance. Plants 2020, 9, 1705. [Google Scholar] [CrossRef]
- Bloem, E.; Haneklaus, S.; Schnug, E. Milestones in plant sulfur research on sulfur-resistance (SIR) in Europe. Front. Plant Sci. 2015, 5, 779. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signalling crosstalk between silicic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Inter. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef]
- Vojtovic, D.; Luhova, L.; Petrivalsky, M. Something smells bad to plant pathogens: Production of hydrogen sulfide and its role in plant defense responses. J. Adv. Res. 2020, 27, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Haneklaus, S.; Bloem, E.; Schnug, E. Effect of foliar-applied elemental sulfur on Fusarium infection in barley. Landbauforsch. Völkenrode 2007, 57, 213–217. [Google Scholar]
- Klikocka, H.; Haneklaus, S.; Bloem, E.; Schnug, E. Influence of sulfur fertilization on infection of potato tubers with Rhizotania silani and Streptomyces scabies. J. Plant Nutr. 2005, 28, 1–14. [Google Scholar] [CrossRef]
- Luce, M.S.; Whalen, J.K.; Ziadi, N.; Zebarth, B.J. Nitrogen dynamics and indices to predict soil nitrogen supply in humid temperate soils. Adv. Agron. 2011, 112, 55–102. [Google Scholar]
- Grzebisz, W.; Niewiadomska, A.; Przygocka-Cyna, K. Nitrogen hotspots on the farm—A practice-oriented approach. Agronomy 2022, 12, 1305. [Google Scholar] [CrossRef]
- Rich, S.M.; Watt, M. Soil conditions and cereal root system architecture: Review and considerations for linking Darvin and Weaver. J. Exp. Bot. 2013, 64, 193208. [Google Scholar] [CrossRef]
- Jobbággy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Raynaud, X.; Leadley, P. Soil characteristics play a key role in modeling nutrient competition in plant communities. Ecology 2004, 85, 2200–2214. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation. Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Nibou, C.; Gibbs, D.J.; Coates, J.C. Branching out in new directions: The control of root architecture by lateral root formation. New Phytol. 2008, 179, 595–614. [Google Scholar] [CrossRef]
- Rout, G.R.; Samantaray, S.; Das, P. Aluminum toxicity in plants: A review. Agronomie 2001, 21, 3–21. [Google Scholar] [CrossRef]
- Meriño-Gergichevich, C.; Alberdi, M.; Ivanov, A.G.; Reyes-Diaz, R. Al3+—Ca2+ interaction in plants growing in acid soils: Al-phytotoxicity response to calcareous amendments. J. Soil Sci. Plant Butr. 2010, 10, 217–243. [Google Scholar]
- Tang, C.; Weligama, C.; Sale, P. Subsurface soil acidification in farming systems: Its possible causes and management options. Mol. Environ. Soil Sci. Prog. Soil Sci. 2013, 13, 20. [Google Scholar]
- Rahman, N.A.; Lee, S.-H.; Chung, H.; Kabir, A.H.; Jones, C.S.; Lee, K.-W. Importance of mineral nutrition for mitigating aluminum toxicity in plants on acid soils: Current status and opportunities. Int. J. Mol. Sci. 2018, 19, 3073. [Google Scholar] [CrossRef]
- Carmeis Filho, A.C.A.; Crusciol, C.A.C.; Guimaraes, T.N.; Calonego, J.C.; Mooney, S.C. Impact of amendments on the physical properties of soil under tropical long-term no till conditions. PLoS ONE 2016, 11, e0167564. [Google Scholar] [CrossRef]
- Watts, D.B.; Dikc, W.A. Sustainable use of FGD gypsum in agricultural systems: Introduction. J. Environ. Qual. 2014, 42, 246–252. [Google Scholar] [CrossRef]
- Lebedev, A.L. Kinetics of gypsum dissolution in water. Geochemistry Inter. 2015, 53, 811–824. [Google Scholar] [CrossRef]
- Chen, L.; Dick, W.A. Gypsum as an agricultural amendment: General use guidelines. Bull. Ohio State Univ. 2011, 945, 36. [Google Scholar]
- Zielewicz, W.; Swędrzyńska, D.; Swędrzyński, A.; Grzebisz, W.; Goliński, P. The influence of calcium sulfate and different doses of potassium on the soil enzyme activity and the yield of the sward with a misture of alfalfa and grasses. Agriculture 2022, 12, 485. [Google Scholar] [CrossRef]
- Shainberg, I.; Summer, M.E.; Miller, W.P.; Farina, M.P.; Pavan, M.A.; Fey, M.V. Use of gypsum on soils: A review. Adv. Soil Sci. 1989, 9, 111. [Google Scholar]
- Summer, M. Gypsum improves subsoil roots growth. In Proceedings of the International Symposium “Root Research and Applications” RootRAP, Vienna, Austria, 2–4 September 2009. [Google Scholar]
- Cuervo-Alzate, J.E.; Osorio, N.W. Gypsum incubation tests to evaluate its potential to effects on acidic soils of Colombia. Rev. Fac. Nac. Agron. Medellin 2020, 73, 9349–9359. [Google Scholar] [CrossRef]
- Pan, W.L.; Kidwell, K.K.; McCracken, V.A.; Bolton, R.P.; Allen, M. Economically optimal wheat yield, protein and nitrogen use component responses to varying N supply and genotype. Front. Plant Sci. 2019, 10, 1790. [Google Scholar] [CrossRef] [PubMed]
- Wooding, A.R.; Kavale, S.; MacRitchie, F.; Stoddard, F.L.; Wallace, A. Effects of nitrogen and sulfur fertilizer on protein composition, mixing requirements, and dough strength of four wheat cultivars. Cereal Che. 2000, 77, 798–807. [Google Scholar] [CrossRef]
- Stępniewska, S. Milling value of grain of selected cultivars from crop years 2012–2014. Acta Agrophysica 2016, 23, 105–117. [Google Scholar]
- Hlisnikovský, L.; Čermák, P.; Kunzová, E.; Barłog, P. The effect of application of potassium, magnesium, and Sulphur on wheat and barley grain yield and protein content. Agron. Res. 2019, 17, 1905–1917. [Google Scholar]
- Potarzycki, J.; Przygocka-Cyna, K.; Wendel, J.; Biniek, Ł.; Ridiger, B. The impact of Sulphur fertilization on yield of winter wheat. Fragm. Agron. 2015, 32, 63–72. [Google Scholar]
- Sadras, V.O.; Slafer, G.A. Environmental modulation of yield components in cereals: Heritability reveal a hierarchy of phenotypic plasticities. Field Crops Res. 2012, 127, 215–224. [Google Scholar] [CrossRef]
- Körner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef]
- Weymann, W.; Bottcher, U.; Sieling, K.; Kage, H. Effects of weather conditions during different growth phases on yield formation of winter oilseed rape. Field Crops Res. 2015, 173, 41–48. [Google Scholar] [CrossRef]
- Grzebisz, W.; Szczepaniak, W.; Grześ, S. Sources of nutrients for high-yielding winter oilseed rape (Brassica napus L.) during post-flowering growth. Agronomy 2020, 10, 626. [Google Scholar] [CrossRef]
- Schulte auf’m Erley, G.; Behrens, T.; Ulas, A.; Wiesler, F.; Horst, W.J. Agronomic traits contributing to nitrogen efficiency of winter oilseed rape cultivars. Field Crops Res. 2011, 124, 114–123. [Google Scholar] [CrossRef]
- Szczepaniak, W.; Grzebisz, W.; Barłóg, P.; Przygocka-Cyna, K. Mineral composition of winter oilseed rape (Brassica napus L.) seeds as a tool for oil seed prognosis. J. Cen. Europ. Agric. 2017, 18, 196–213. [Google Scholar] [CrossRef]
- Szulc, P.; Bocianowski, J.; Rybus-Zając, M. The effect of soil supplentation with nitrogen and elemental sulfur on chlorphyll content and grain yield of maize (Zea mays L.). Žemdirbystè=Agriculture 2012, 99, 247–254. [Google Scholar]
- Salviagotti, F.; Castellalrin, J.M.; Miralles, D.J.; Pedrol, H.M. Sulfur fertilization improves nitrogen use efficiency in wheat by increasing nitrogen uptake. Field Crops Res. 2009, 113, 170–177. [Google Scholar] [CrossRef]
- Pagani, A.; Echeverria, H.E.; Andrade, F.H.; Sainz Rozas, H.R. Effects of nitrogen and sulfur application on grain yield, nutrient accumulation, and harvest indexes in maize. J. Plant Nutr. 2012, 35, 1080–1097. [Google Scholar] [CrossRef]
- Schnug, E.; Haneklus, S. Diagnosis of Sulphur nutrition. In Sulphur in Agroecosystems; Schnug, E., Ed.; Kluwer Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 1–38. [Google Scholar]
- Haneklaus, S.; Fleckenstaein, J.; Schnug, E. Comaprative studies of plant and soil analysis for the Sulphur status of oilseed rape and winter wheat. Z. Pflanzenernähr. Bodenk. 1995, 158, 109–111. [Google Scholar] [CrossRef]
- Inal, A.; Günes, A.; Alpaslan, M.; Adak, M.S.; Taban, S.; Erasian, F. Diagnosis of sulfur deficiency and effects of sulfur on yield and yield components of wheat grown in Central Anatolia, Turkey. J. Plant Nutr. 2003, 26, 1483–1498. [Google Scholar] [CrossRef]
- Grzebisz, W.; Przygocka-Cyna, K. Spring malt barley response to elemental sulfur—the prognostic value of N and S concentrations in malt barley leaves. Plant Soil Environ. 2007, 53, 388–394. [Google Scholar] [CrossRef]
- Zhao, F.J.; Hawkesford, M.J.; Warrilow, A.G.S.; McGrath, S.P.; Clarkson, D.T. responses of two wheat varieties to Sulphur application and diagnosis of Sulphur deficiency. Plant Soil 1996, 181, 317–327. [Google Scholar] [CrossRef]
- Reussi Calvo, N.I.; Echeverria, H.E.; Sainz Rozas, H.R. Usefulness of foliar nitrogen-sulfur ratio in spring red wheat. J. Plant Nutr. 2008, 31, 1612–1623. [Google Scholar] [CrossRef]
- Sedlar, O.; Balik, J.; Kulhanek, M.; Cerny, J.; Suran, P. Sulphur nutrition index in relation to uptake and quality of winter wheat grain. Chilean J. Agric. Res. 2019, 79, 486–492. [Google Scholar] [CrossRef]
- Potarzycki, J. Influence of nitrogen and magnesium fertilization at the flag leaf stage of winter wheat development on the yield and grain quality. Fertil. Fertil. 2008, 32, 99–109. [Google Scholar]
- Grzebisz, W.; Szczepaniak, W.; Barłóg, P.; Przygocka-Cyna, K.; Potarzycki, J. Phosphorus sources for winter oilseed rape (Brassica napus L.) during reproductive growth–magnesium sulfate management impact on P use efficiency. Archiv. Agron. Soil Sci. 2018, 64, 1646–1662. [Google Scholar] [CrossRef]
- Potarzycki, J. Yield forming effect of combined application of magnesium, sulphur and zinc in maize fertilization. Fertilizers Fertilization 2010, 39, 44–59. [Google Scholar]
- Talboys, P.J.; Heppell, J.; Roose, T.; Healey, J.R.; Jones, D.L.; Withers, P.J.A. Struvite: A slow-release fertilizer for sustainable phosphorus management. Plant Soil 2016, 401, 109–123. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).