
Nitrogen Hotspots on the Farm—A Practice-Oriented Approach




Abstract
:1. Introduction—Food Gap and Sustainable Agriculture
2. Nitrogen Sources
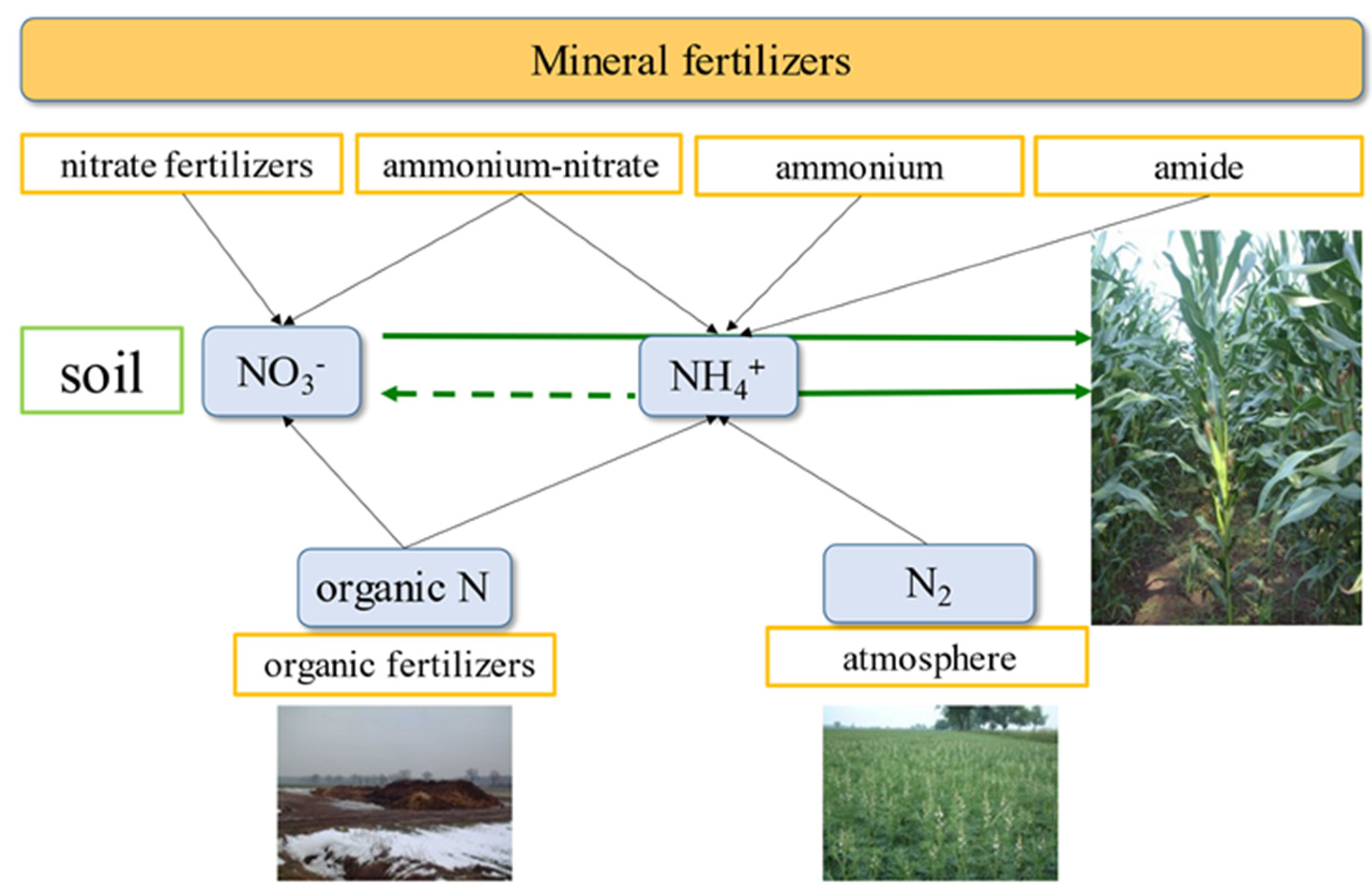
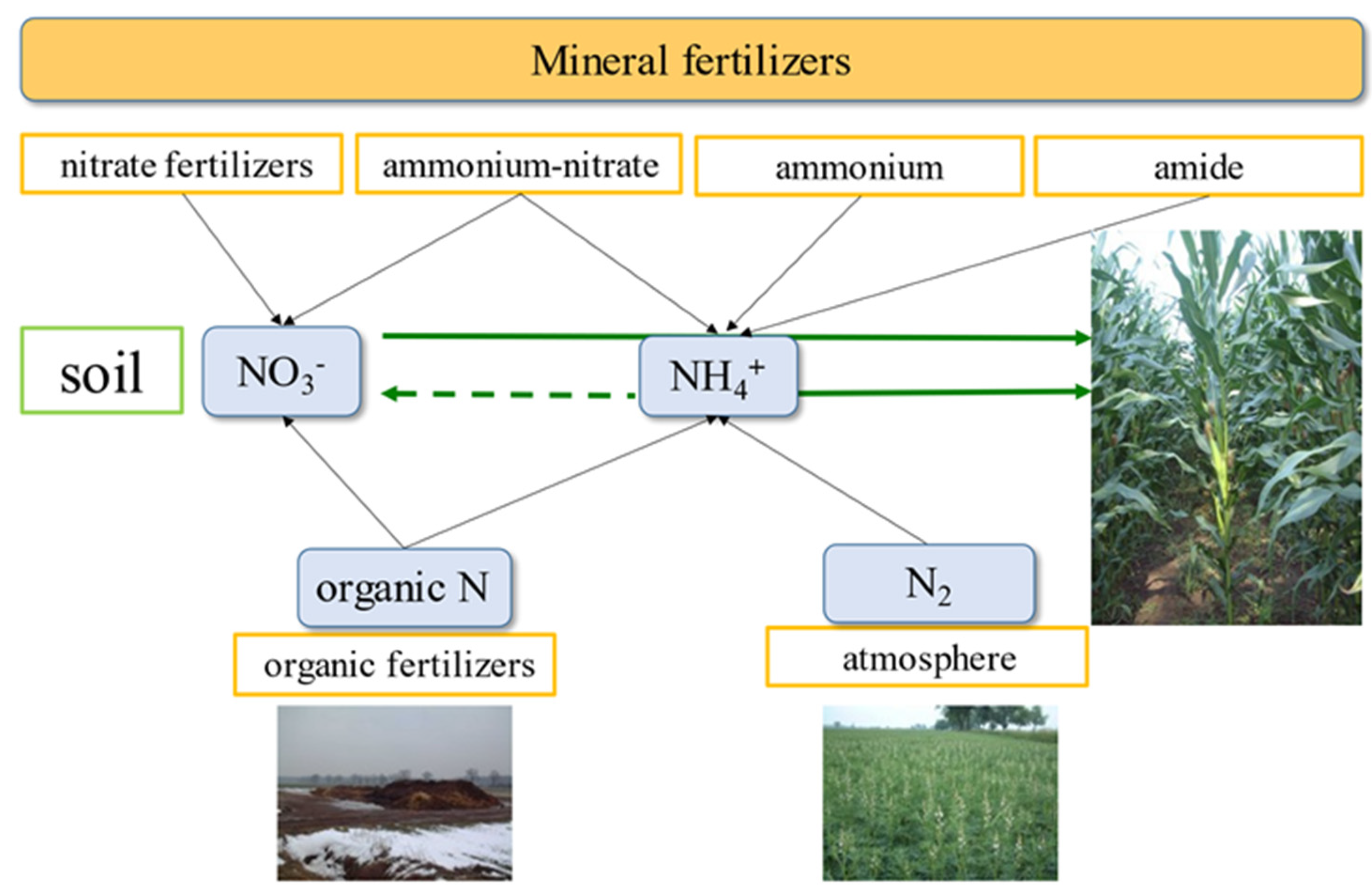
2.1. Simplified Diagram of the N Cycle on the Farm
2.2. Biological Nitrogen Fixation (BNF)—The Primary Source of N for Plants
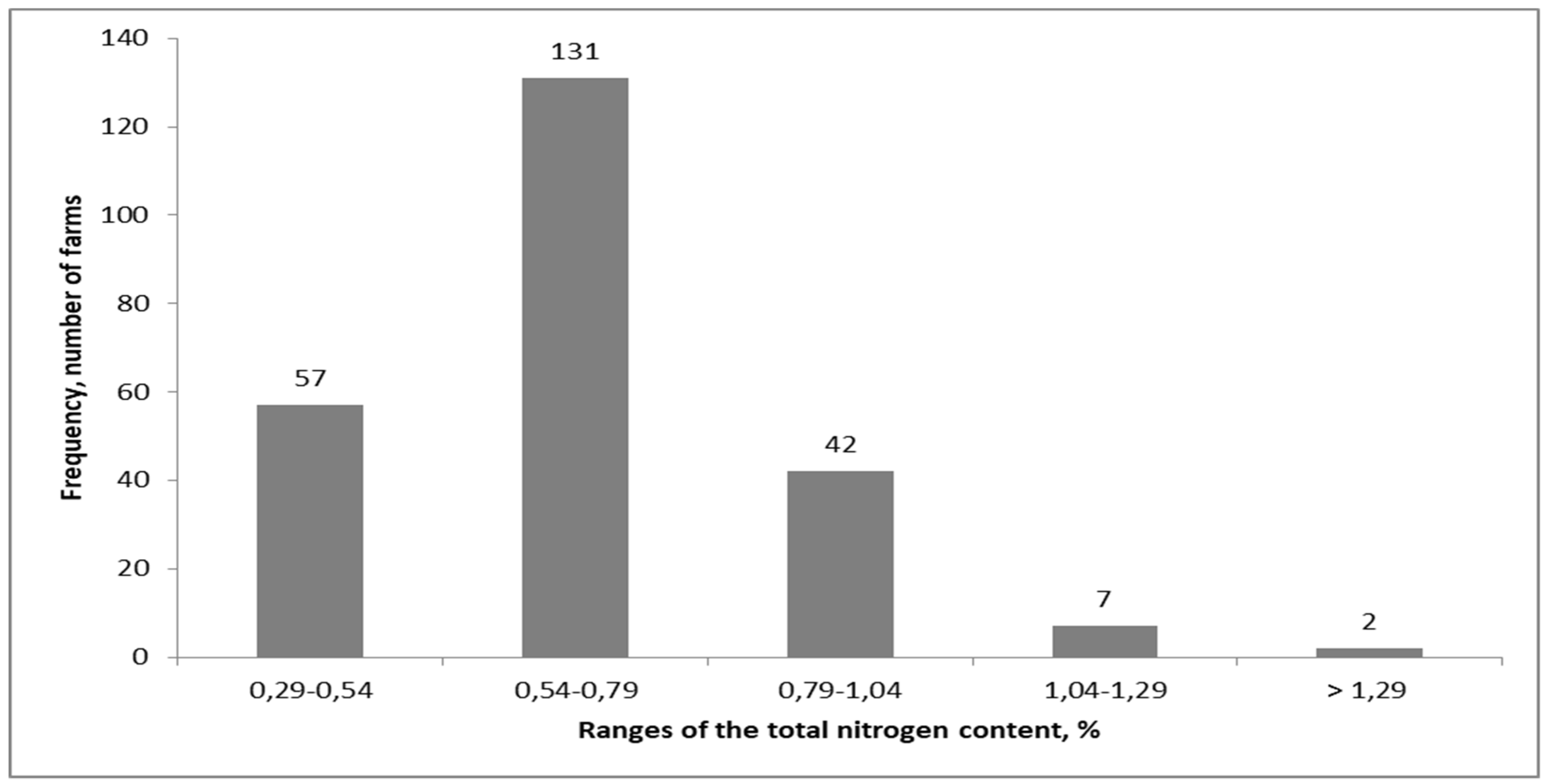
2.3. Recycled Sources of Nitrogen
- C0—initial percentage of C mass incorporated into the soil, at t = 0, kg C × ha−1;
- C(1)—percentage of residual C, at time t1, kg C × ha−1;
- t—time, days (months, years);
- k—specific rate constant, day−1, (month−1, year−1);
- e—constant of natural logarithm, 2.718.
- (1)
- Half-C decomposition constant, t1/2 = 0.693/k; refers to the time needed for 50% of C0 decomposition.
- (2)
- C decomposition constant, t0.05 = 3/k; refers to the time taken for 95% of C0 decomposition.
- (1)
- Fresh organic matter (FOM); k > 3.0 and t0.05 < 1.0 year.
- (2)
- Active humus (AH, ≈ 5% of the total humus content); 3.0 > k > 0.6 and t0.05 < 5.0 years.
- (3)
- Labile humus (LH, 60–85%); 0.6 > k > 0.03 and t0.05 < 100 years.
- (4)
- Stable humus (SH, 10–40%); k < 0.03 and t0.05 ≈ 100–10,000 years.
2.3.1. Crop Residues
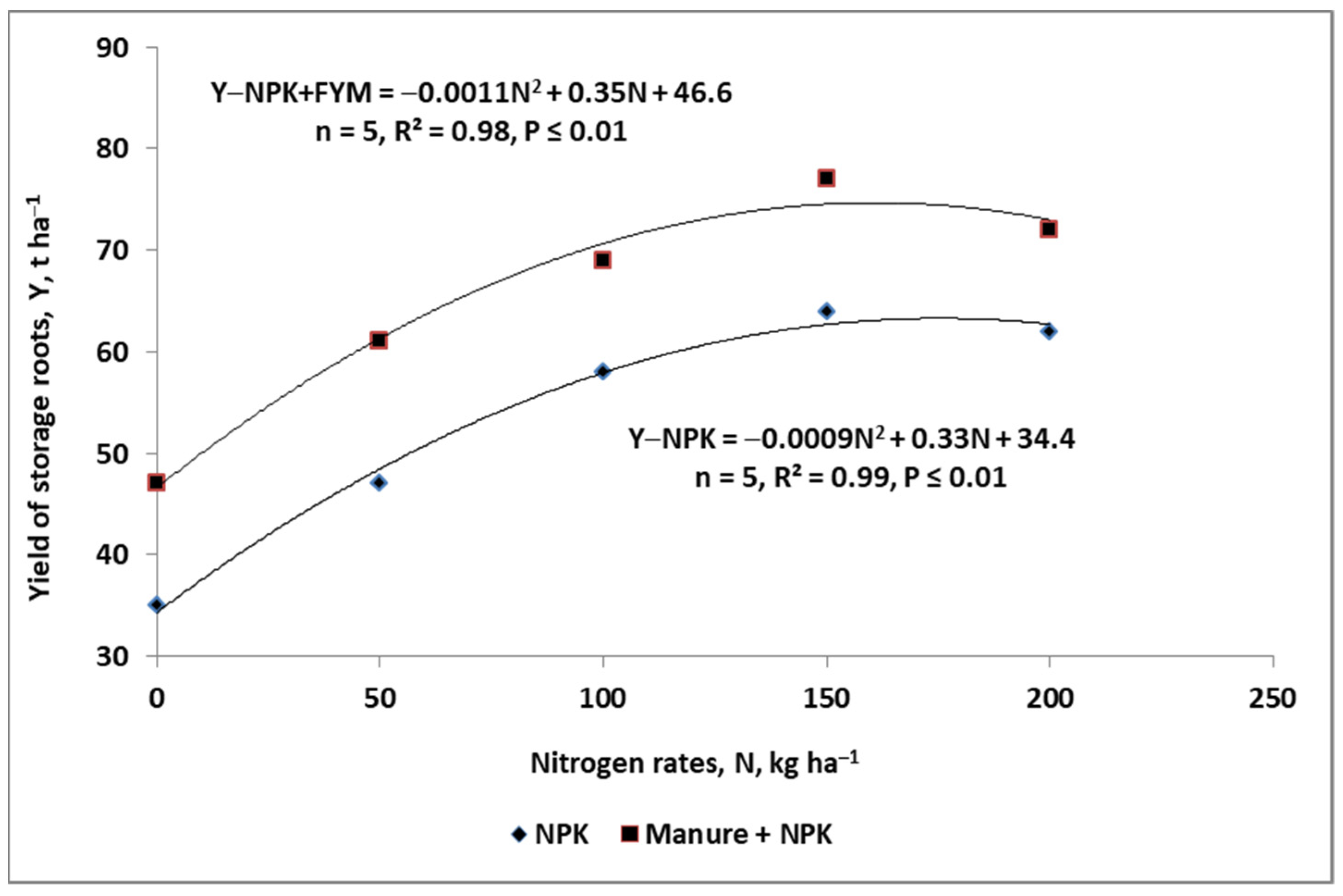
2.3.2. Manure
- The highest, primary fertilizing value of the livestock excreta to be maintained. The goal of the action is to increase the effectiveness of the use of nutrients applied as fertilizer.
- The loss of dry matter and nutrients to the environment to be minimized. The purpose of the applied solutions is to reduce the negative impact of animal production on the environment.
- TEd—daily amount of excreta, kg day−1 cow−1;
- Milk—milk daily production, kg day−1 cow−1;
- Urine—urine daily production, dm3 day−1;
- BW—cow body weight, kg;
- TNd—daily amount of N in excreta, g day−1 cow−1;
- DMI—dry matter intake, kg day−1 cow−1;
- Ccp—concentration of crude proteins, g 100 g−1 of dry fed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method of Manure Fermentation | N Losses, % of Initial | Source |
|---|---|---|
| Straw–loose pile–liquid outflow | 30 | [102] |
| 30–50 | ||
| 45–55 | [103] | |
| Cut straw–mechanically compacted pile | 18 | [102] |
| 25 | ||
| 23 | [103] | |
| Straw–a pile flooded with liquid slurry or water | 10 | [102] |
2.3.3. Transformation of Organic Amendments in the Soil
- (1)
- Biodegradability:
- Total N and C content;
- C:N ratio;
- Chemical composition;
- Lignin content.
- (2)
- Soil conditions:
- One-time dose of fertilizer introduced into the soil;
- Soil temperature;
- Soil pH;
- Other factors influencing soil fauna and microorganism activity.
- (1)
- Direct N mineralization. Conditions for this trend are:
- N content > 1.8% DW;
- C:N ratio < than 22.2: 1.
- (2)
- Fluctuation in N immobilization/mineralization processes up to 1–2 years after the incorporation of FOM into the soil. Conditions are:
- N content in the range of 1.2–1.8% DW;
- C:N ratio in the range of 22.2–33.3: 1.
- (3)
- Direct N immobilization. Conditions are:
- N content < 1.2% DW,
- C:N ratio > than 33.3: 1.
- (1)
- Direct effect of the N released from the applied fertilizer or CRs;
- (2)
- Increase in the content of soil available nutrients others than N;
- (3)
- Overall improvement in soil fertility (humus content, soil structure, water content).
- (1)
- Soil mineral N;
- (2)
- Livestock slurry;
- (3)
- Digestate from agricultural biogas plants;
- (4)
- Mineral N fertilizer.
- Nd—added N, kg ha−1;
- CRB—biomass of crop residues incorporated into the soil, t ha−1;
- NCRB—N content in CRs, kg t−1 DW;
- 12—critical N content in OM (1.2% DW), recalculated into kg t−1 DW of CRs.
3. Nitrogen—Driver of Crop Production
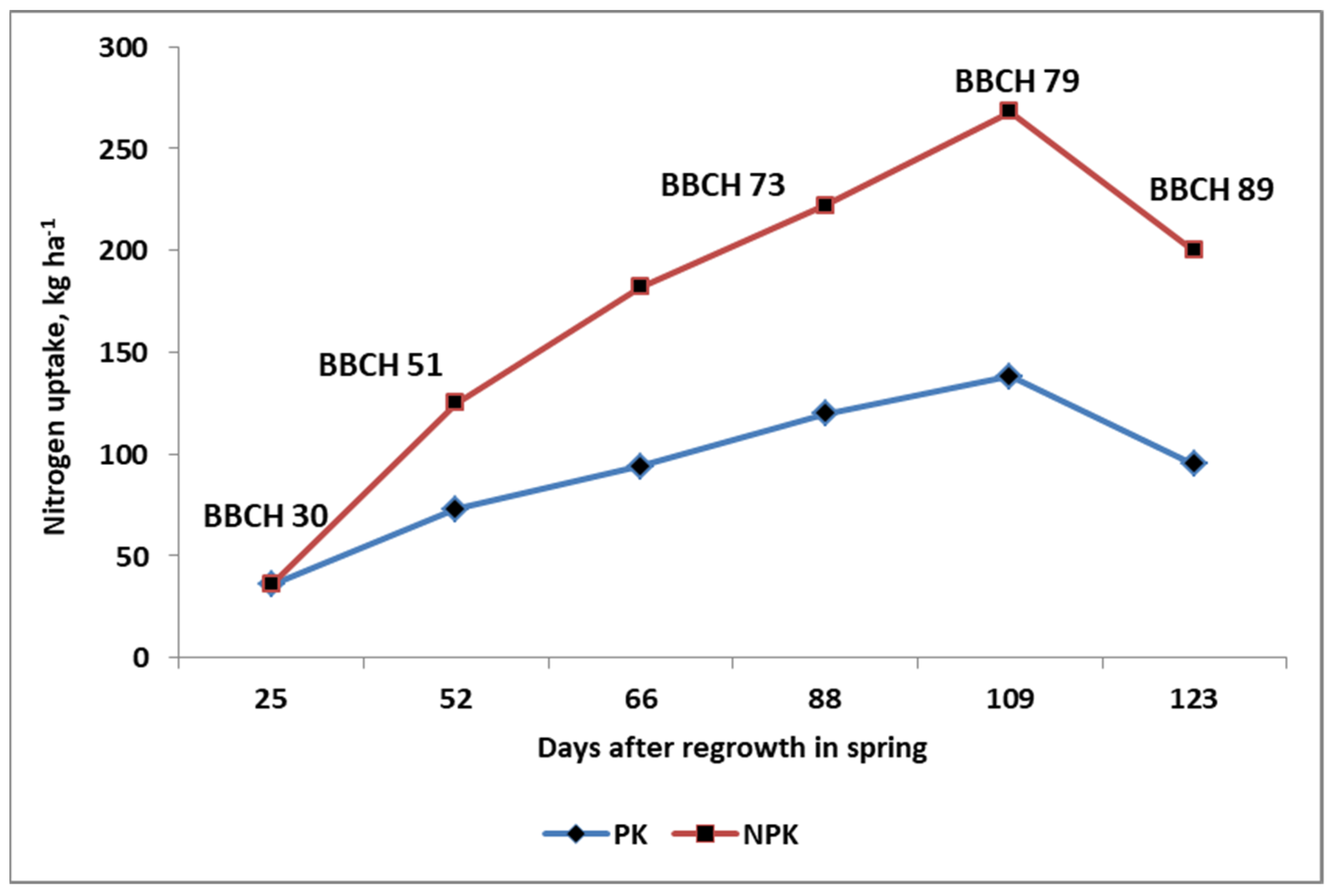
3.1. Nitrogen Uptake by Plants
- (1)
- Necessary condition—the concentration of N-NO3 in the soil solution, or N-NH4 both in the soil solution and in the soil exchange complex;
- (2)
- Sufficient condition: (i) water content in the air—vapor pressure deficit, and (ii) root density.
3.2. Critical Stages of Nitrogen Accumulation by Plants
- (1)
- Crop Foundation Period—CFP;
- (2)
- Yield Formation Period—YFP;
- (3)
- Yield Realization Period—YRP.

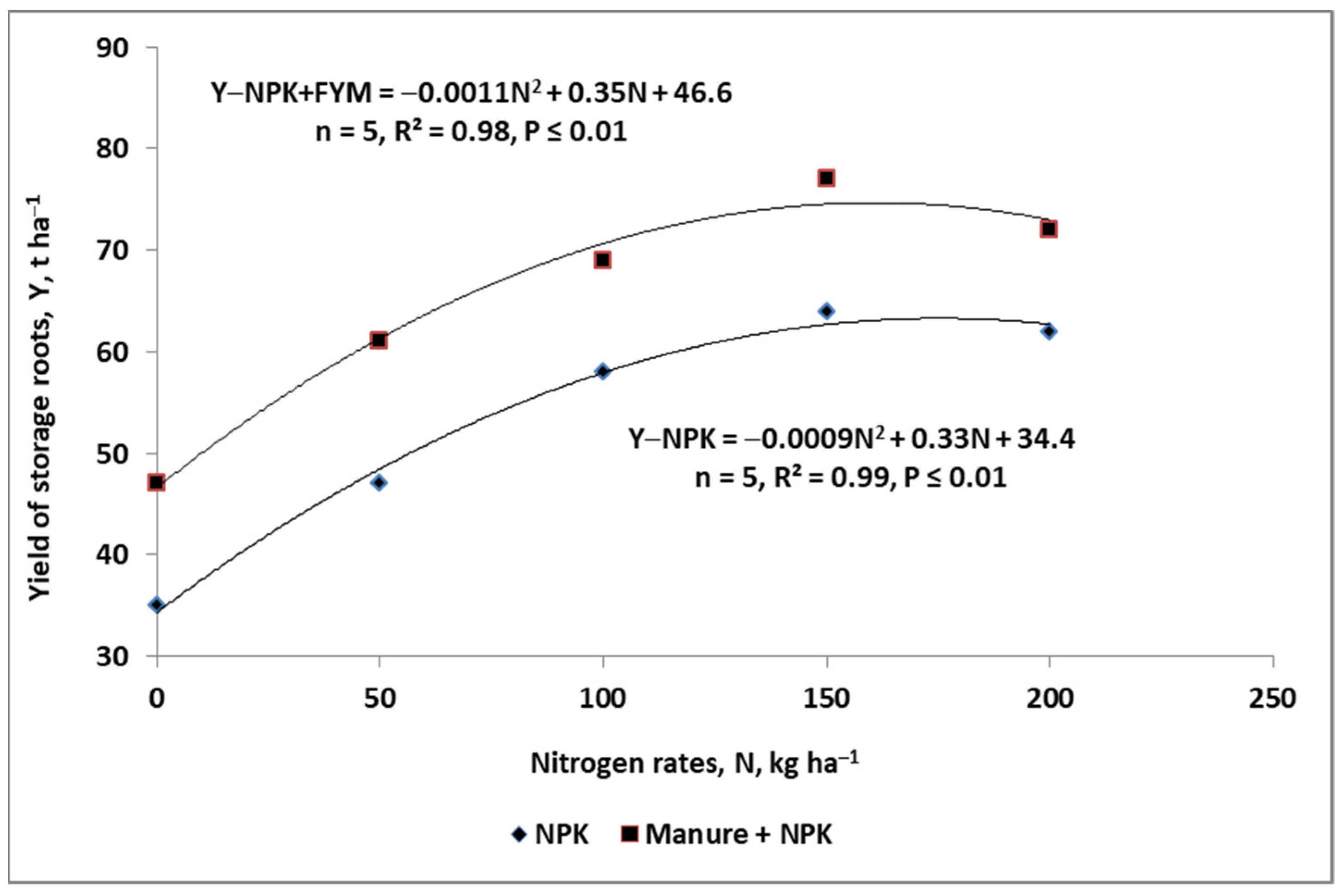
3.3. Crop Response to Nitrogen Fertilizer
- (1)
- Maximum yield of the currently grown crop variety;
- (2)
- Actual N plant status at critical stages of yield component(s) formation;
- (3)
- Amount of available N in soil resources:
- At the beginning of the growing season,
- Released from soil resources during the growing season.
- (1)
- Optimum Nf rate (Nfop):
- (2)
- Maximum achievable yield:
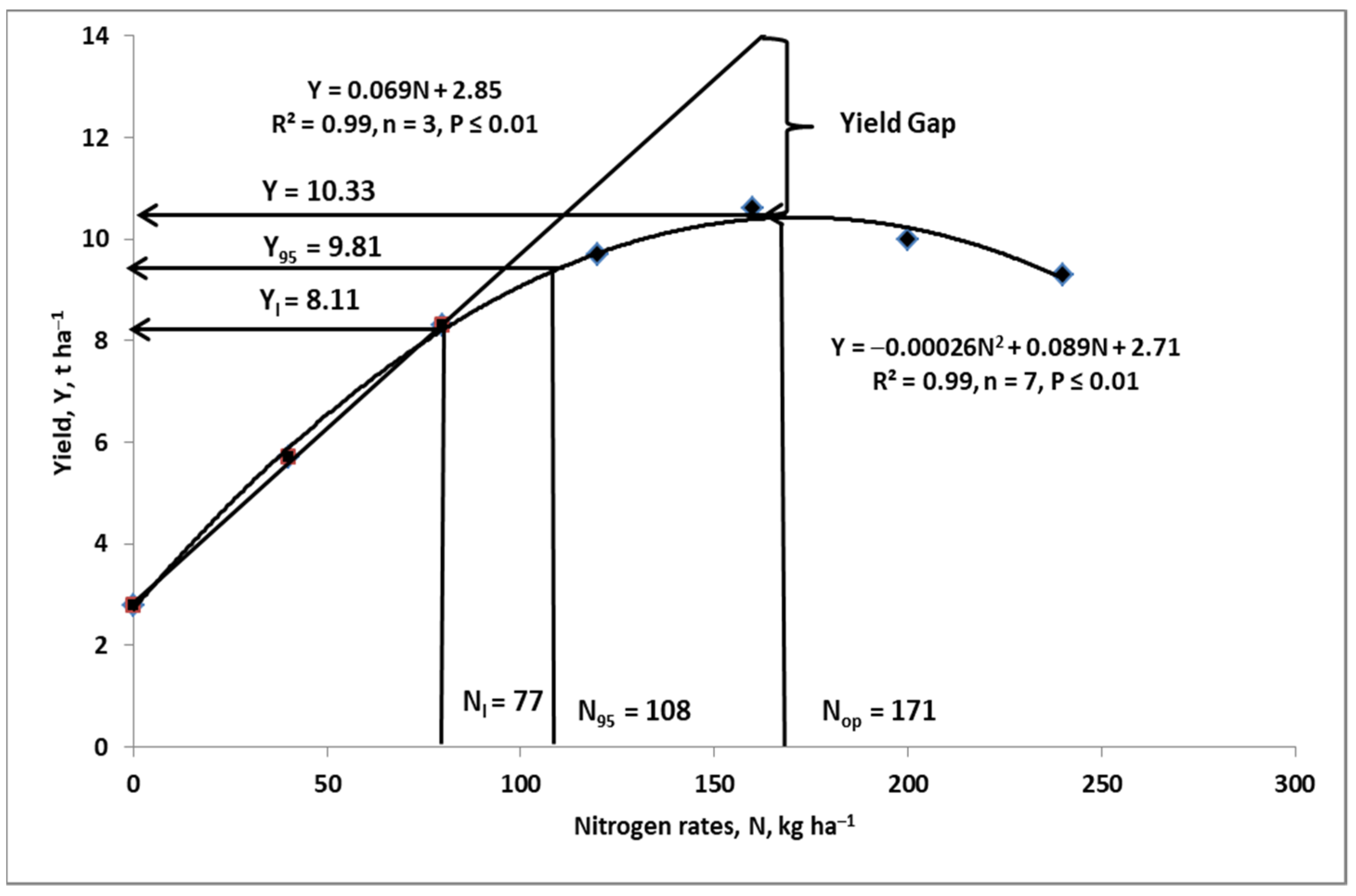
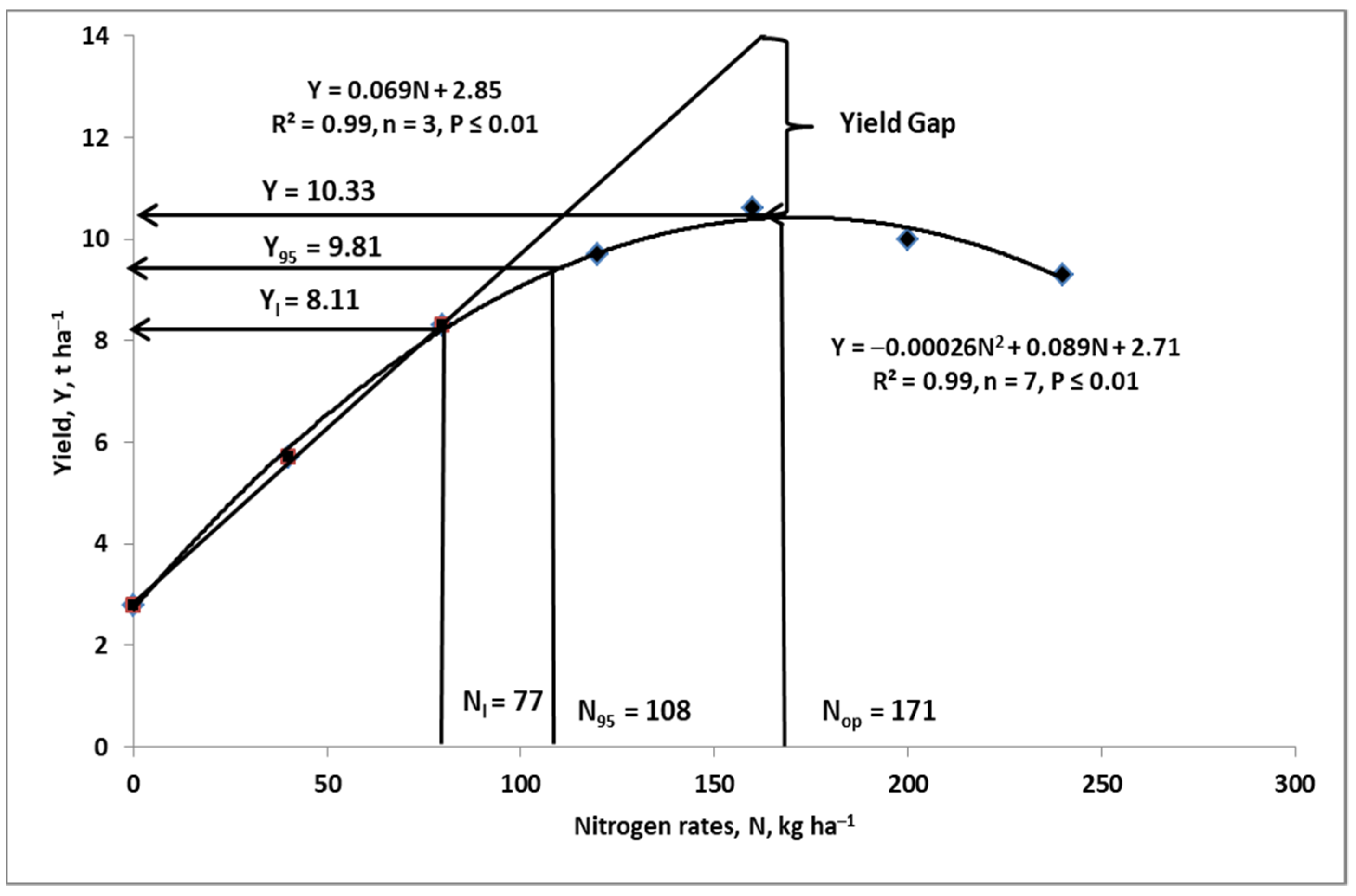
4. Nitrogen Budgeting on the Farm
4.1. Nitrogen Cycle—Crop Succession Approach
- (1)
- Farm Gate Balance (FGB);
- (2)
- Soil Surface Balance (SSU);
- (3)
- Soil System Balance (SSB).
- (1)
- Residual in-organic N may be partially taken up by the next crop;
- (2)
- Production effect of manure N is longer than one growing season;
- (3)
- N in crop residues significantly impacts the N flow within more than one growing season.
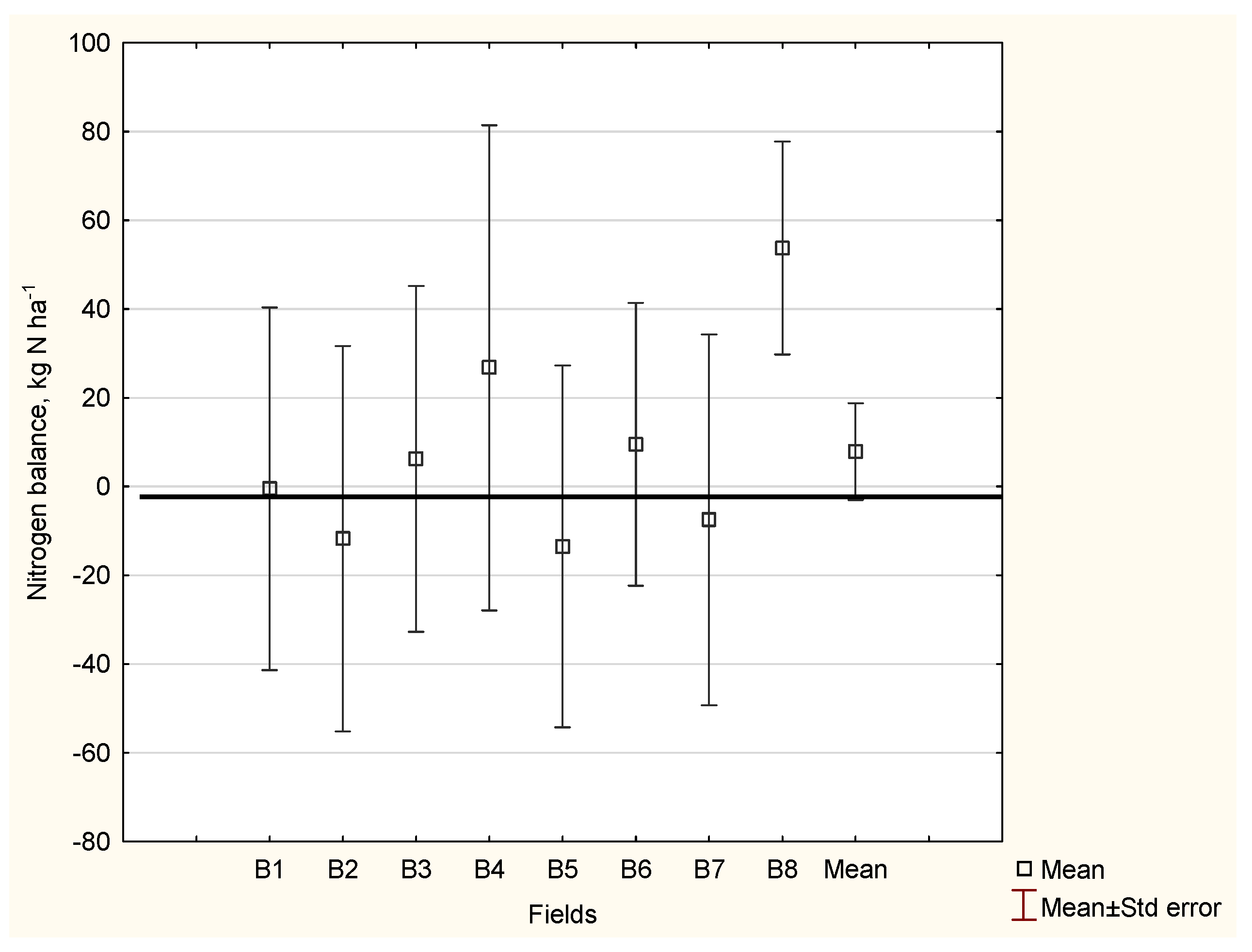
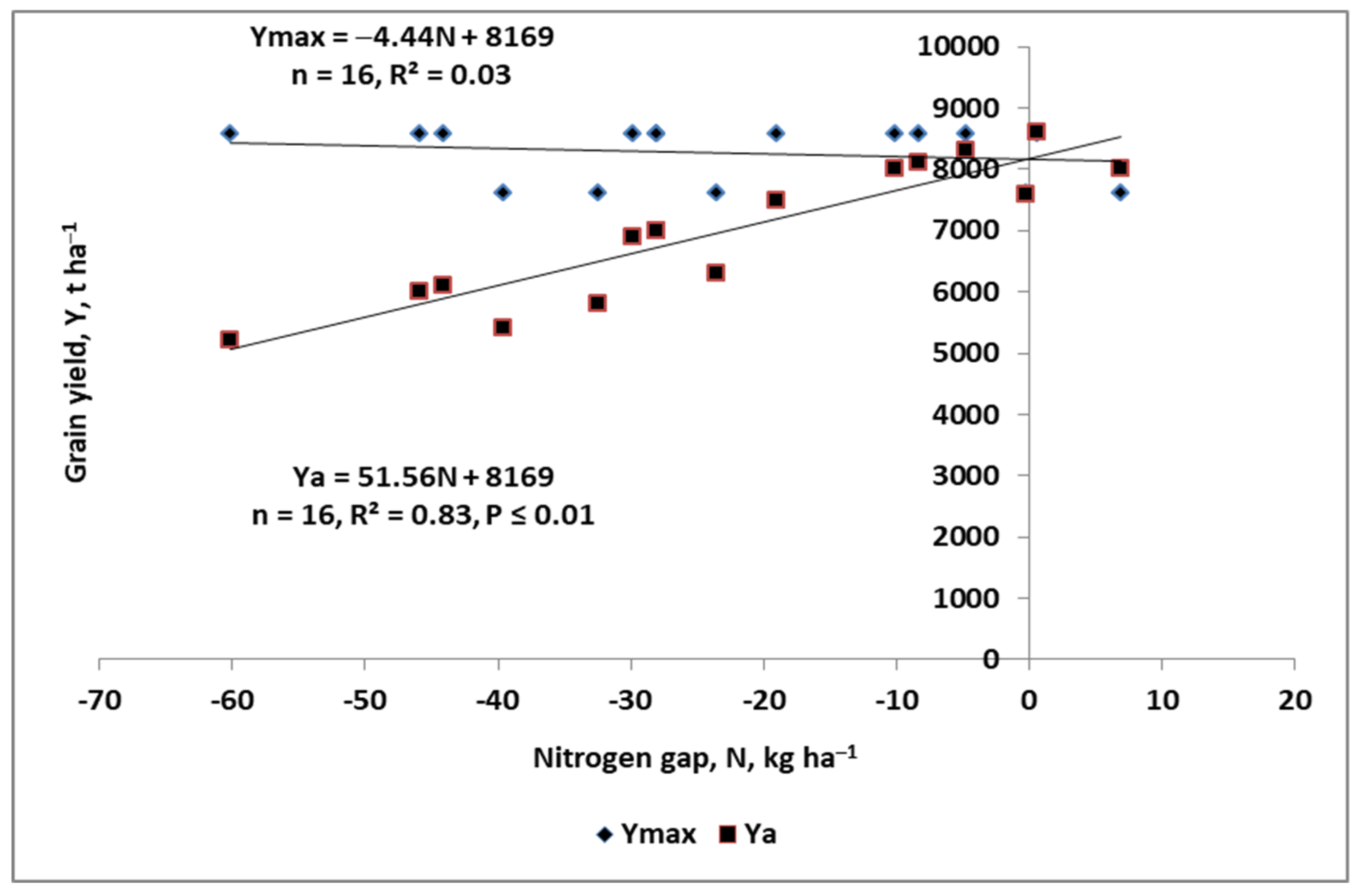
4.2. Nitrogen Gap
- Ya—actual yield of a current growing crop, t ha−1;
- Nf—amount of applied fertilizer N, kg ha−1;
- PFP-Nf—partial factor of productivity of Nf, kg grain/seeds, tubers etc. per kg Nf;
- Yattmax—maximum attainable yield, t ha−1;
- YG—yield gap, t ha−1;
- NG—nitrogen gap, kg N ha−1.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
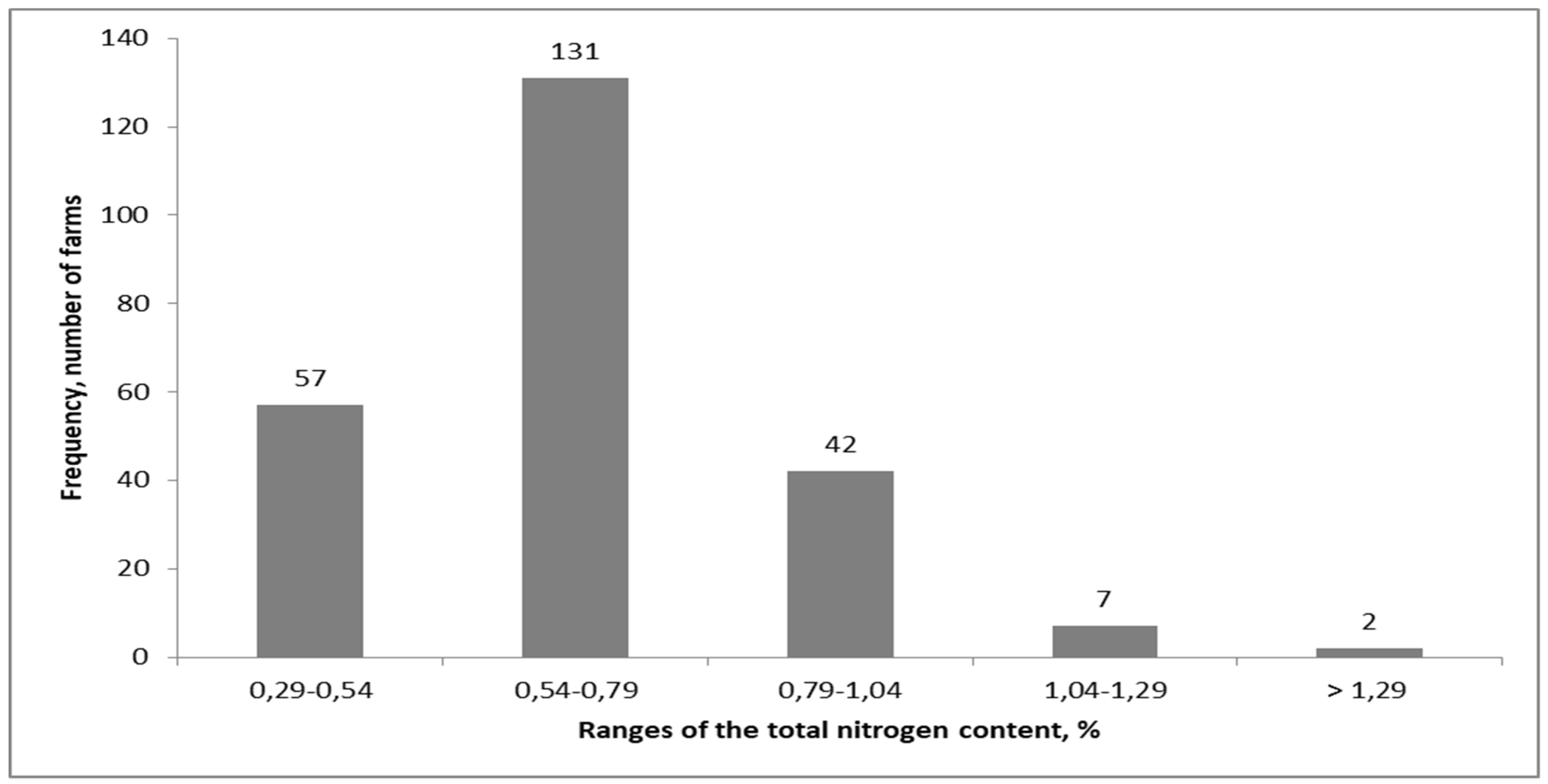
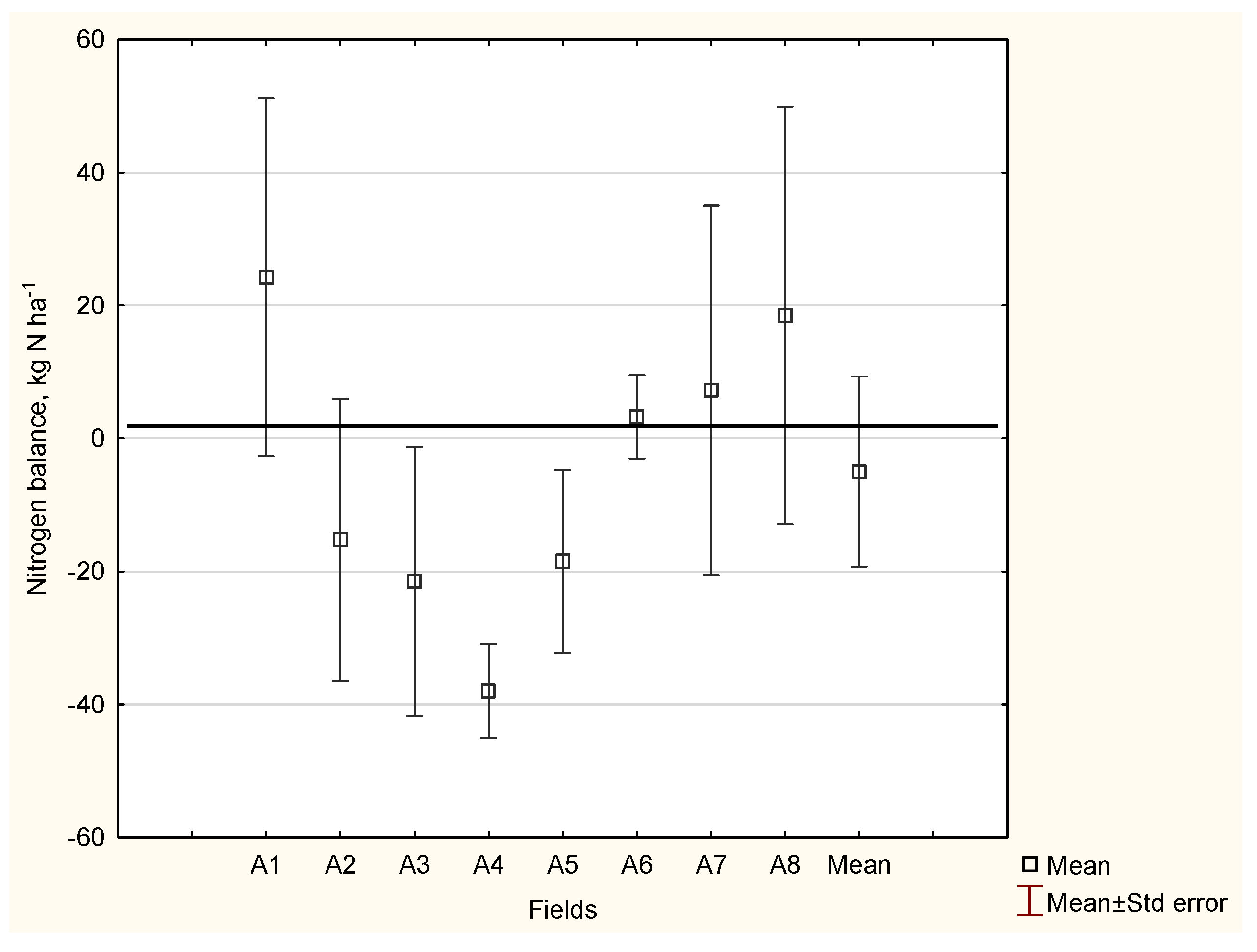
Appendix A
| Field Characteristics | Field Number → Decreasing Nitrogen Gap | |||
|---|---|---|---|---|
| 4 | 13 | 3 | 11 | |
| Nitrogen gap, kg N ha−1 | −61 | −46 | −20 | +6 |
| Yield Gap, kg ha−1 | −3414 | −2614 | −1114 | +343 |
| Soil usability class | Low | Medium/low | Medium | Medium |
| Fore-crop | Low | Low | Low | Low |
| Variety | Medium intensive | Medium intensive | Medium intensive | Intensive |
| Sowing term | Adequate | Adequate | Adequate | Adequate |
| Manure | Lack | Lack | Lack | Lack |
| Soil reaction (pH) | Acidic | Slightly acid | Neutral | Neutral |
| Phosphorus content—class | Low | Medium | Very high | High |
| Potassium content—class | Medium | Low | Medium | Medium |
| Magnesium content class | Low | Medium | High | High |
| Fungicide protection | Medium | Medium | Medium | Medium |
References
- Le Mouël, C.; Forslund, A. How can we feed the world in 2050? A review of the responses from global scenario studies. Eur. Rev. Agric. Econ. 2017, 44, 541–591. [Google Scholar] [CrossRef]
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. BioScience 2017, 67, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Beltran-Peña, A.; Rosa, L.; D’Odorico, P. Global food self-sufficiency in the 21st century under sustainable intensification of agriculture. Environ. Res. Lett. 2020, 15, 095004. [Google Scholar] [CrossRef]
- Keating, B.A.; Herrero, M.; Carberry, P.S.; Gardner, J.; Cole, N.B. Food wedges: Framing the global food demand and supply challenge towards 2050. Glob. Food Sec. 2014, 3, 125–132. [Google Scholar] [CrossRef]
- Röös, E.; Bajželj, B.; Smith, P.; Patel, M.; Little, D.; Garnett, T. Greedey or needy? Land use and climate impacts of food in 2050 under different livestock futures. Glob. Environ. Chang. 2017, 47, 1–12. [Google Scholar] [CrossRef]
- Alexander, P.; Brown, C.; Ameth, A.; Finnigan, J. Human appropriation for land and food: The role of diet. Glob. Environ. Chang. 2016, 41, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Berners-Lee, M.; Kennelly, C.; Watson, R.; Hewitt, C.N. Current global food production is sufficient to meet human nutritional needs in 2050 provided there is radical societal adaptation. Elem. Sci. Anth. 2018, 5, 52. [Google Scholar] [CrossRef]
- Kummu, M.; Fader, M.; Gerten, D.; Guillaume, J.H.A.; Jalava, M.; Jägermeyr, J.; Pfister, S.; Porkka, M.; Siebert, S.; Varis, O. Bringing it al together: Linking measures to secure nations’ food supply. Curr. Opin. Environ. Sustain. 2017, 29, 98–117. [Google Scholar] [CrossRef] [Green Version]
- Harvey, M.; Pilgrim, S. The new competition for land: Food, Energy, and climate change. Food Policy 2011, 36, S40–S50. [Google Scholar] [CrossRef]
- Smith, P.; Gregory, P.J.; van Vuuren, D.; Obersteiner, M.; Havlik, P.; Rounsevell, M.; Woods, J.; Stehfest, E.; Bellarby, J. Competition for land. Phil. Trans. R. Soc. B 2010, 365, 2941–2957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomiero, T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenza, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Inter. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L. Agriculture, plant physiology, and human population growth: Past, present, and future. Theor. Exp. Plant Physiol. 2013, 25, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Erisman, J.W.; Leach, A.; Bleeker, A.; Atwell, B.; Cattaneo, L.; Galloway, J. An integrated approach to a nitrogen use efficiency (NUE) indicator for the food production-consumption chain. Sustainability 2018, 10, 925. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conijn, J.G.; Bindraban, P.S.; Schröder, J.J.; Jongschaap, R.E.E. Can our global food system meet food demand within planetary boundries? Agric. Ecosyst. Environ. 2018, 251, 244–256. [Google Scholar] [CrossRef]
- Pradhan, P.; Fischer, G.; van Velthuizen, H.; Reusser, D.E.; Kropp, J.P. Closing yield gaps: How sustainable can we be? PLoS ONE 2015, 10, e0129487. [Google Scholar] [CrossRef] [Green Version]
- The Royal Society. Reaping the Benefits: Science and the Sustainable Intensification of Global Agriculture; RS Policy Document; The Royal Society: London, UK, 2009; p. 86. [Google Scholar]
- Grafton, R.Q.; Williams, J.; Jiang, Q. Food and water gaps to 2050: Preliminary results from the global food and water systems (GFWS) platform. Food Secur. 2015, 7, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Larson, J.A.; Stefanini, M.; Yin, X.; Boyer, C.N.; Lambert, D.M.; Zhou, X.V.; Tubaña, B.S.; Scharf, P.; Varco, J.J.; Dunn, D.J.; et al. Effects of landscape, soils, and weather on yields, nitrogen use, and profitability with sensor-based variable rate nitrogen management in cotton. Agronomy 2020, 10, 1858. [Google Scholar] [CrossRef]
- Grzebisz, W.; Łukowiak, R. Nitrogen gap amelioration is a core for sustainable intensification of agriculture—A concept. Agronomy 2021, 11, 419. [Google Scholar] [CrossRef]
- Bodirsky, B.L.; Popp, A.; Lotze-Campen, H.; Dietrich, J.P.; Rolinski, S.; Weindl, I.; Smitz, C.; Müller, C.; Bonsch, M.; Humpenöder, F.; et al. Reactive nitrogen requirements to feed the world in 2050 and potential to mitigate nitrogen pollution. Nat. Commun. 2014, 5, 3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassaletta, L.; Billen, G.; Garnier, J.; Bouwman, L.; Velazquez, E.; Muleller, N.D.; Gerber, J.S. Nitrogen use in the global system: Past trends and future trajectories of agronomic performance, pollution, trade, and dietary demand. Environ. Res. Lett. 2016, 11, 095007. [Google Scholar] [CrossRef]
- Haene, K.D.; Salomez, J.; De Neve, S.; De Waele, J.; Hofman, G. Environmental performance of nitrogen fertilizer limits imposed by the EU Nitartes Directive. Agric. Ecosyst. Environ. 2014, 192, 67–79. [Google Scholar] [CrossRef]
- Oenema, O.; Pietrzak, S. Nutrient management in food production. Achieving agronomic and environmental targets. Ambio 2003, 31, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, G.; Jeuffroy, M.-H.; Gastal, F. Diagnosis tool for plant an crop N status in vegetative stage: Theory and practices for crop N management. Eur. J. Agron. 2008, 28, 181–190. [Google Scholar] [CrossRef]
- Córdova, C.; Barrera, J.A.; Magna, C. Spatial variation in nitrogen mineralization as a guide for variable application of nitrogen fertilizer to cereal crops. Nutr. Cycl. Agroecosyst. 2018, 110, 83–88. [Google Scholar] [CrossRef]
- Schilizzi, S.; Pannel, D.J. The economics of nitrogen fixation. Agronomie 2001, 21, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Gutser, R.; Ebertseder, T.; Weber, A.; Schraml, M.; Schmidhalter, U. Short-term and residual availability of nitrogen after long-term application of organic fertilizers on arable land. J. Plant Nutr. Soil. Sci. 2005, 168, 439–446. [Google Scholar] [CrossRef]
- Bussink, D.W.; Oenema, O. Ammonia volatilization from dairy farming systems in temperate areas. A review. Nutr. Cycl. Agroecosyst. 1998, 51, 19–33. [Google Scholar] [CrossRef]
- Mary, B.; Recous, S.; Darwis, D.; Robin, D. Interactions between decomposition of plant residues and nitrogen cycling in soil. Plant Soil 1996, 181, 71–82. [Google Scholar] [CrossRef]
- Smil, V. Nitrogen in crop production: An account of global flows. Glob. Biogeochem. Cycles 1999, 13920, 647–662. [Google Scholar] [CrossRef] [Green Version]
- de Vries, W.; Schulte-Uebbing, L.; Kros, H.; Voogd, J.C.; Louwagie, G. Spatially explicit boundaries for agricultural nitrogen inputs in the European Union to meet air and water quality target. Sci. Total Environ. 2021, 786, 147283. [Google Scholar] [CrossRef] [PubMed]
- Luce, M.S.; Whalen, J.K.; Ziadi, N.; Zebarth, B.J. Nitrogen dynamics and indices to predict soil nitrogen supply in humid temperate soils. Adv. Agron. 2011, 112, 55–102. [Google Scholar]
- Grzebisz, W.; Łukowiak, R.; Sassenrath, G. Virtual nitrogen as a tool for assessment of nitrogen at the field scale. Field Crops Res. 2018, 218, 182–184. [Google Scholar] [CrossRef]
- Olfs, H.-W.; Blankenau, K.; Brentrup, F.; Jasper, J.; Link, A.; Lammel, J. Soil- and plant-based nitrogen-fertilizer recommendations in arable farming. J. Plant Nutr. Soil Sci. 2005, 168, 414–431. [Google Scholar] [CrossRef]
- Barłóg, P.; Łukowiak, R.; Grzebisz, W. Predicting the content of soil mineral nitrogen based on the content of calcium chloride-extractable nutrients. J. Plant Nutr. Soil Sci. 2017, 180, 624–635. [Google Scholar] [CrossRef]
- Dobermann, A. Nitrogen use efficiency—State of the art. In Proceedings of the IFA International Workshop on Enhanced Efficiency Fertilizers, Frankfurt, Germany, 28–30 June 2005; pp. 1–16. [Google Scholar]
- Liu, Y.; Wu, L.; Baddeley, J.A.; Watson, C.A. Models of biological nitrogen fixation of legumes. Sustain. Agric. 2011, 2, 883–905. [Google Scholar]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995; 899p. [Google Scholar]
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by rhizobia—The roots of a success story. Curr. Opin. Plant Biol. 2018, 44, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Soumare, A.; Giedhiou, A.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kousni, L. Exploiting biological nitrogen fixation: A route towards sustainable agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Masson-Boivin, C.; Giraud, E.; Perret, X.; Batut, J. Establishing nitrogen-fixing symbiosis with legumes: How many rhizobium recipes? Trends Microbiol. 2009, 17, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Davies-Barnard, T.; Friedlingstein, P. The Global Distribution of Biological Nitrogen Fixation in Terrestrial Natural Ecosystems. Glob. Biogeochem. Cycles 2020, 34, e2019GB006387. [Google Scholar] [CrossRef]
- Bever, J.D.; Simms, E.L. Evolution of nitrogen fixation in spatially structured populations of Rhizobium. Heredity 2000, 85, 366–372. [Google Scholar] [CrossRef]
- McFall-Ngai, M.J.; Gordon, J.I. Experimental Models of Symbiotic Host-Microbial Relationships: Understanding the Underpinnings of Beneficence and the Origins of Pathogenesis. In Evolution of Microbial Pathogens; Steven, H., DiRita, V.J., Eds.; Jon Wiley & Sons Inc.: New York, NY, USA, 2006; Volume 9, pp. 147–166. [Google Scholar]
- Voisin, A.S.; Salon, C.; Jeudy, C.; Warembourg, F.R. Symbiotic N2 fixation activity in relation to C economy of Pisum sativum L. as a function of plant phenology. J. Exp. Bot. 2003, 54, 2733–2744. [Google Scholar] [CrossRef] [Green Version]
- Kahindi, J.H.; Karanja, N.K. Essentials of Nitrogen Fixation Biotechnology. Biotechnol.-Vol. VIII Fundam. Biotechnol. 2009, 8, 54–80. [Google Scholar]
- Sujkowska, M. The course of the infection process in the symbiotic system of legumes-Rhizobium. Bot. News 2009, 53, 35–53. [Google Scholar]
- Martyniuk, S. Production of microbial preparations: Symbiotic bacteria of legumes as an ex ample. J. Res. Appl. Agric. Eng. 2010, 55, 20–23. [Google Scholar]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G.; Valori, F. Microbial diversity and microbial activity in the rhizosphere. Cienc. Del Suelo 2007, 25, 89–97. [Google Scholar]
- Pathak, J.; Rajneesh; Maurya, P.K.; Singh, S.P.; Häder, D.-P.; Sinha, R.P. Cyanobacterial farming for environment friendly sustainable agriculture practices: Innovations and perspectives. Front. Environ. Sci. 2018, 6, 7. [Google Scholar] [CrossRef]
- Waraczewska, Z.; Niewiadomska, A. Diazotroph—Characteristics of the symbiotic legume—Rhizobium. In Leguminous Plants in Polish Agriculture: Genetics, Breeding, Cultivating and Using; Poznan University of Life Science: Poznań, Poland, 2017; pp. 83–94. [Google Scholar]
- Zahran, H.H. Rhizobia from wild legumes: Diversity, taxonomy, ecology, nitrogen fixation and biotechnology. J. Biotech. 2001, 91, 143–153. [Google Scholar] [CrossRef]
- Graham, P.H. Stress tolerance in Rhizobium and Bradyrhizobium, and nodulation under adverse soil conditions. Can. J. Microbiol. 1992, 38, 475–484. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Aldasoro, J.; Arrese-Igor, C.; Erice, G.; Sanz-Sáez, Á. How Does High Temperature Affect Legume Nodule Symbiotic Activity? In Legume Nitrogen Fixation in a Changing Environment; Sulieman, S., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Fageria, N.K.; Nascente, A.S. Management of soil acidity of South American soils for sustainable crop production. Adv. Agron. 2014, 128, 221–275. [Google Scholar]
- Martinze-Hidalgo, P.; Hirsch, A.M. The nodule microbiome: N2-fixing Rhizobia do not live alone. Phytobiomes 2017, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Kochian, L.V.; Hoekenga, O.A.; Piñeros, M.A. How do crop plants tolerate acid soils? Mechanism of aluminum tolerance and phosphorus efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Waraczewska, Z.; Budka, A. The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation. Agronomy 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Weisany, W.; Raei, Y.; Allahverdipoor, K.H. Role of some of mineral nutrients in biological nitrogen fixation. Bulletin of Environment. Pharmacol. Life Sci. 2013, 2, 77–84. [Google Scholar]
- Sadeghipour, O.; Abbasi, S. Soybean response to drought and seed inoculation. World Appl. Sci. J. 2012, 17, 55–60. [Google Scholar]
- Cooper, J.; Scherer, H. Nitrogen Fixation. In Mineral Nutrition of Higher Plants, 3rd ed.; Marchner, P., Ed.; Academic Press: London, UK, 2012; pp. 389–408. [Google Scholar]
- Roy, R.N.; Finck, A.; Blair, G.J.; Tandon, H.L.S. Plant nutrition for food security. A guide for integrated nutrient management. FAO Fertil. Plant Nutr. Bull. 2006, 16, 368. [Google Scholar]
- Bambara, S.; Ndakidemi, P.A. The potential roles of lime and molybdenum on the growth, nitrogen fixation and assimilation of metabolites in nodulated legumes: A special reference to Phaseolus vulgaris L. African J. Biotech. 2010, 8, 2482–2489. [Google Scholar]
- Bellaloui, N.; Mengistu, A.; Kasses, M.A.; Abel, C.A.; Zobiole, L.H.S. Role of boron nutrient in nodules growth and nitrogen fixation in soybean genotypes under water stress conditions. In Advance in Biology and Ecology of Nitrogen Fixation; Ohyama, T., Ed.; InTech: London, UK, 2014; pp. 237–258. [Google Scholar]
- Hu, X.; Wei, X.; Ling, J.; Chen, J. Cobalt: An essential micronutrient for plant growth. Front. Plant Sci. 2021, 16, 768523. [Google Scholar] [CrossRef]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Torma, S.; Vilček, J.; Lošák, T.; Kužel, S.; Martensson, A. Residual plant nutrients in crop residues—An importnat resource. Acta Agric. Scand. Sec. B Soil Plant Sci. 2017, 68, 358–366. [Google Scholar]
- Kumar, T.K.; Rana, D.S.; Nain, L. Legume residue and N management for improving productivity and N economy and soil fertility in wheat (Triticum aestivum)—Based cropping system. Natl. Acad. Sci. Lett. 2019, 42, 297–307. [Google Scholar] [CrossRef]
- Ludwig, B.; Geisseler, D.; Michel, K.; Joergensen, R.G.; Schulz, E.; Merbach, I.; Raupp, J.; Rauner, R.; Hu, K.; Niu, L.; et al. Effects of fertilization and soil management on crop yields and carbon stabilization in soils. A review. Agron. Sustain. Dev. 2011, 31, 361–372. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.; Black, A.L. Quantification of the effect of soil organic matter content on soil productivity. Soil Sci. Soc. Am. J. 1994, 58, 185–193. [Google Scholar] [CrossRef]
- Karlen, D.L.; Lal, R.; Follet, R.F.; Komble, J.M.; Hatfield, R.D.; Miranowski, J.M.; Cambardella, C.A.; Manale, A.; Anex, R.P.; Rice, C.W. Crop residues: The rest of the story. Environ. Sci. Technol. 2009, 43, 8011–8015. [Google Scholar] [CrossRef] [Green Version]
- Scotti, R.; Bonanomii, G.; Scelza, R.; Zoina, A.; Rao, M.A. Organic amendments as sustainable tool to recovery soil fertility in intensive agricultural systems. J. Soil Sci. Plant Nutr. 2015, 15, 333–352. [Google Scholar]
- Parton, W.; Schimel, D.; Cole, C.; Ojima, D. Analysis of factors controlling soil organic matter in great plains grasslands. Soil. Sci. Soc. Am. 1987, 51, 1173–1179. [Google Scholar] [CrossRef]
- Stevenson, F.J. Cycles of Soil; John Wiley & Sons: New York, NY, USA, 1986; 380p. [Google Scholar]
- Bolinder, M.A.; Crotty, F.; Elsen, A.; Feąc, M.; Kismányoky, T.; Lipiec, J.; Tits, M.; Tóth, Z.; Kätterer, T. The effect of crop residues, cover crops, manures and nitrogen fertilziation on soil organic carbon changes in agroecosystems: A synthesis of reviews. Mitig. Adapt. Strateg. Glob. Chang. 2020, 25, 929–952. [Google Scholar] [CrossRef]
- Koishi, A.; Bragazza, L.; Maltas, A.; Guillaume, T.; Sinaj, S. Lon-term efefcts of organic amendments on soil organic matetr quantity and quality in conventional cropping systems in Switzerland. Agronomy 2020, 10, 1977. [Google Scholar] [CrossRef]
- Hiel, M.P.; Chélin, M.; Prvin, N.; Barbieux, S.; Degrune, F.; Lemtiri, A.; Colinet, G.; Degré, A.; Bodson, B.; Garré, S. Crop residue management in arable cropping systems under temperate climate. Part 2. Soil physical properties and crop production. A review. Biotechnol. Agron. Sci. Environ. 2016, 20, 2450256. [Google Scholar] [CrossRef]
- Wnuk, A.; Górny, A.G.; Bocianowski, J.; Kozak, M. Visualizing harvest index in crops. Comm. Biometry Crop Sci. 2013, 8, 48–59. [Google Scholar]
- Stepaniuk, M.; Głowacka, A. Yield of winter oilseed rape (Brassica napus L. var. bnapus) in a short-term monoculture and the macronutrient accumulation in relation to the dose and method of Sulphur application. Agronomy 2022, 12, 68. [Google Scholar] [CrossRef]
- Grzebisz, W. Crop Plants Fertilization. Part 2. Fertilizers and Fertilization Systems; PWRiL: Poznań, Poland, 2015; 396p. (In Polish) [Google Scholar]
- Zhao, X.; Yuan, G.; Wang, H.; Lu, D.; Chen, X.; Zhou, J. Effects of full straw incorporation on soil fertility and crop yield in rice-wheat rotation for silty clay loamy cropland. Agronomy 2019, 9, 133. [Google Scholar] [CrossRef] [Green Version]
- Venkatramanan, V.; Shah, S.; Rai, A.K.; Prasad, R. Nexus between Crop Residue Burning, Bioeconomy and Sustainable Development Goals Over North-Western India. Front. Energy Res. 2021, 392. [Google Scholar] [CrossRef]
- Kamusoko, R.; Jingura, R.M.; Parawira, W.; Chikwambi, Z. Strategies for valorization of crop residues into biofuels and other value-added products. Biofpr 2021, 15, 1950–1964. [Google Scholar] [CrossRef]
- Sarkar, S.; Skalicky, M.; Hossain, A.; Brestic, M.; Saha, S.; Gorai, S.; Ray, K.; Brahmachari, K. Management of crop residues for improving input use efficiency and agricultural sustainability. Sustainability 2020, 12, 9808. [Google Scholar] [CrossRef]
- Parmar, A.; Sturm, B.; Hensel, O. Crops that feed the world: Production and improvement of cassava for food, feed and industrial uses. Food Secur. 2017, 9, 907–927. [Google Scholar] [CrossRef]
- Whitemore, A.P.; Groot, J.J.R. The decomposition of sugar beet residues: Mineralization versus immobilization in contrasting soil types. Plant Soil 1997, 192, 237–247. [Google Scholar] [CrossRef]
- Tomaszewska, J.; Bieliński, D.; Binczarski, M.; Berlowska, J.; Dziugan, P.; Piotrowski, J.; Stanishevsky, A.; Witońska, I.A. Products of sugar beet processing as raw materials for chemical and biodegradable polymers. RSC Adv. 2018, 8, 3161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotz, C.A. Management to reduce nnitrogen losses in animal production. J. Anim. Sci. 2004, 82, E119–E137. [Google Scholar] [PubMed]
- Loyon, L. Overview of animal manure management for beef, pig, and poultry in France. Front. Sustain. Food Syst. 2018, 2, 36. [Google Scholar] [CrossRef]
- Sefeedpari, P.; Pudełko, R.; Jędrejek, A.; Kozak, M.; Borzęcka, M. To what extent is manure produced, distributed and potentially available for bioenergy? A step toward stimulating circular bio-economy in Poland. Energies 2020, 13, 6266. [Google Scholar] [CrossRef]
- ASAE. Manure Production and Characteristics; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2014; Volume D384.1 DEC99 (R2014). [Google Scholar]
- Misselbrook, T.H.; Powell, J.M. Influence of bedding material on ammonia emissions from cattle excreta. J. Dairy Sci. 2005, 88, 4304–4312. [Google Scholar] [CrossRef]
- Kupper, T.; Hani, C.; Neftel, A.; Kincaid, C.; Buhler, M.; Amon, B.; Vander Zaag, A. Ammonia and greenhouse gas emissionsw from slurry storage—A review. Agric. Ecosyst. Environ. 2020, 300, 106963. [Google Scholar] [CrossRef]
- Whitehead, D.C.; Raistrick, N. Nitrogen in excreta if dairy cattle: Changes during short-term storage. J. Agric. Sci. 1993, 21, 73–81. [Google Scholar] [CrossRef]
- Paul, J.W.; Beauchamp, E.G. Nitrogen availability for corn in soils amended with urea, cattle slurry, and solid and composted manure. Can. J. Soil Sci. 1993, 73, 253–266. [Google Scholar] [CrossRef]
- Fangueiro, D.; Hjorth, M.; Gioelli, F. Acidification of animal slurry—A review. J. Environ. Manag. 2015, 149, 46–56. [Google Scholar] [CrossRef]
- Christensen, M.; Sommer, S. Manure characterization and inorganic chemistry. In Animal Manure Recycling: Treatment and Management; Sommer, S., Christensen, M., Schmidt, T., Jensen, L., Eds.; John Wiley & Sons. Ltd.: Chichester, UK, 2013. [Google Scholar]
- Boratyński, K. Organic Fertilizers; PWRiL: Warsaw, Poland, 1977; 231p. (In Polish) [Google Scholar]
- Dou, Z.; Koln, R.; Ferguson, J.; Boston, R.; Newbold, J. Managing nitrogen on dairy farms. An integrated approach. I. Model description. J. Dairy Sci. 1996, 79, 2071–2080. [Google Scholar] [CrossRef]
- Chen, B.; Liu, E.; Tian, Q.; Yan, C.; Zhang, Y. Soil nitrogen dynamics and crop residues. A review. Agron. Sustain. Dev. 2014, 34, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Grzyb, A.; Wolna-Maruwka, A.; Niewiadomska, A. Environmental factors affecting the mineralization of crop residues. Agronomy 2020, 10, 1951. [Google Scholar] [CrossRef]
- Hijbeek, R.; Ten Berge, H.F.; Whitemore, A.P.; Barkusky, D.; Schroder, J.J.; van Ittersum, M.K. Nitrogen fertiliser replacement values for organic amendments appear to increase with N application rates. Nutr. Cycl. Agroecosyst. 2018, 110, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Tarafdar, J.C.; Panwar, J.; Kathju, S. A rapid method for assessment of plant residue quality. J. Plant Nutr. Soil Sci. 2003, 166, 662–666. [Google Scholar] [CrossRef]
- Rahn, C.R.; Bending, G.D.; Turner, M.K.; Lillywhite, R.D. Management of N mineralization from crop residues of high N content using amendment materials of varying quality. Soil Use Manag. 2003, 19, 193–200. [Google Scholar] [CrossRef]
- Obalum, S.E.; Chibuike, G.U.; Peth, S.; Ouyang, Y. Soil organic matter as sole indicator of soil degradation. Environ. Monit. Assess. 2017, 189, 176. [Google Scholar] [CrossRef]
- Spychalski, W.; Grzebisz, W.; Diatta, J.; Kostarev, D. Humus stock degradation and impacts on phosphorus forms in arable soils—A case of the Ukrainian Forest Steppe Zone. Chem. Spec. Bioavail. 2018, 30, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Wadman, W.P. Effect of organic manure on crop yield in long-term field experiments. INTECOL Bull. 1987, 15, 83–89. [Google Scholar]
- Gross, A.; Glase, B. Meta-analysis on how manure application changes soil organic carbon storage. Sci. Rep. 2022, 11, 5516. [Google Scholar] [CrossRef]
- Przygocka-Cyna, K.; Grzebisz, W. The multifactorial effect of digestate on the availability of soil elements and grain yield and its mineral profile. Agronomy 2020, 10, 275. [Google Scholar] [CrossRef] [Green Version]
- Ötvös, K.; Marconi, M.; Vega, A.; O’brian, J.; Johnston, A.; Abualla, R.; Antonielli, L.; Montesinos, J.C.; Zhang, Y.; Yan, S.; et al. Modulation of plant root growth by nitrogen source-defined regulation of polar auxin transport. EMBO J. 2021, 40, e106862. [Google Scholar] [CrossRef] [PubMed]
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security. A review. Agron. Sustain. Dev. 2010, 30, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanism of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sondergaard, T.E.; Schulz, A.; Palmgreen, M.G. Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase. Plant Physiol. 2004, 136, 2475–2482. [Google Scholar] [CrossRef] [Green Version]
- Masoni, A.; Ercoli, L.; Mariotti, M.; Arduini, I. Post–anthesis accumulation and remobilization of dry matter, nitrogen and phosphorus in durum wheat as affected by soil type. Eur. J. Agron. 2007, 26, 179–186. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Y.; Xu, G. How does nitrogen shape plant architecture. J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef]
- Hu, Q.-Q.; Shu, J.-Q.; Lin, W.-M.; Zhang, G.-Z. Role of auxin and nitrate signaling in the development of root system architecture. Front. Plant Sci. 2021, 12, 690363. [Google Scholar] [CrossRef]
- Lynch, J.L. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma Membrane H+ -ATPase Regulation in the Center of Plant Physiology. Mol. Plant. 2016, 9, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Muratore, C.; Espen, L.; Prinsi, B. Nitrogen uptake in plants: The plasma membrane root transport systems from a physiological proteomic perpective. Plants 2021, 10, 681. [Google Scholar] [CrossRef]
- Marschner, H.; Kirkby, E.A.; Cakmak, J. Effect of mineral nutrition on shoot-root partitioning of photo-assimilates and cycling of mineral nutrients. J. Exp. Bot. 1996, 47, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.M.; Bryant, D.C.; Denison, R.F. Wheat yields, nitrogen uptake, and soil moisture following winter legume cover crop vs. Fallow. Agron. J. 1998, 90, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Barłog, P.; Grzebisz, W. Effect of timing and nitrogen fertilizer application on winter oilseed rape (Brassica napus L.). II. Nitrogen uptake dynamics and fertilizer efficiency. J. Agron. Crop Sci. 2004, 190, 314–323. [Google Scholar] [CrossRef]
- Barraclough, P.B. The growth and activity of winter wheat roots in the field: Nutrient uptakes of high-yielding crops. J. Agric. Sci. Camb. 1986, 106, 45–52. [Google Scholar] [CrossRef]
- Grzebisz, W.; Szczepaniak, W.; Grześ, S. Sources of nutrients for high-yielding winter oilseed rape (Brassica napus L.) during post-flowering growth. Agronomy 2020, 10, 626. [Google Scholar] [CrossRef]
- Sylvester-Bradley, R.; Lunn, G.; Foulkes, J.; Shearman, V.; Spink, J.; Ingram, J.; Management Strategies for Yield of Cereals and Oilseed Rape. HGCA Conference: Agronomic Intelligence: The Basis for Profitable Production. HGCA. (Vol. 18). Available online: www.hgca.com/publications (accessed on 14 November 2021).
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wiren, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Mayer, U. BBCH Monograph. In Growth Stages of Mono- and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Center for Agriculture and Forestry: Berlin, Germany, 2001; Available online: http://www.jki.bund.de/fileadmin/dam_uploads/_veroeff/bbch/BBCH-Skala_Englisch.pdf (accessed on 14 March 2021).
- Yin, X.; Goudriaan, J.; Lantinga, E.A.; Vos, J.; Spiertz, H. A flexible sigmoid function of determinate growth. Ann. Bot. 2003, 91, 361–371. [Google Scholar] [CrossRef]
- Kurepa, J.; Smalle, J.A. Auxin/cytokinin antagonistic control of the shoot/root growth ration and its relevance for adaptation to growth and nutrient deficiency stress. Int. J. Mol. Sci. 2022, 23, 1933. [Google Scholar] [CrossRef]
- Koprna, R.; Humplik, J.F.; Spisek, Z.; Bryksova, M.; Zatloukal, M.; Mik, V.; Novak, O.; Nisler, J.; Dolezal, K. Improvement of tillering and grain yield by application of cytokinin derivatives in wheat and barley. Agronomy 2021, 11, 67. [Google Scholar] [CrossRef]
- Slafer, G.A.; Elia, M.; Savin, R.; Garcia, G.A.; Terrile, I.I.; Ferrante, A.; Miralles, D.J.; González, F.G. Fruiting efficiency: An alternative trait to further rise in wheat yield. Food Energy 2015, 4, 92–109. [Google Scholar] [CrossRef]
- Klepper, B.; Rickman, R.W.; Waldman, S.; Chevalier, P. The physiological life cycle of wheat: Its use in breeding and crop management. Euphytica 1998, 100, 341–347. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, D.; Schnurbusch, T. Plant and floret growth at distinct developmental stages during the stem elongation phase in wheat. Front. Plant. Sci. 2018, 9, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Preanthesis biomass accumulation and plant organs defines yield components in wheat. Eur. J. Agron. 2016, 81, 15–26. [Google Scholar] [CrossRef]
- Barłóg, P. Effect of magnesium and nitrogen fertilizers on the growth and alkaloid content of Lupinus angustifolius L. Aust. J. Agric. Res. 2002, 53, 671–676. [Google Scholar] [CrossRef]
- Kong, L.G.; Xie, Y.; Hu, L.; Feng, B.; Li, S.D. Remobilization of vegetative nitrogen to developing grain in wheat (Triticum aestivum L.). Field Crops Res. 2016, 196, 134–144. [Google Scholar] [CrossRef]
- Spiertz, J.; Vos, J. Grain Growth of Wheat and Its Limitation by Carbohydrate and Nitrogen Supply. In Wheat Growth and Modelling; Day, W., Atkin, R., Eds.; Plenum Press: New York, NY, USA, 1985. [Google Scholar]
- Bergmann, W. Nutritional Disorders of Plants; Verlag Gustav Fisher: Jena, Germany, 1992; p. 741. [Google Scholar]
- Elwali, A.; Gasho, G.; Summer, M. DRIS norms for 11 nutrients in con leaves. Agronomy J. 1985, 77, 506–508. [Google Scholar] [CrossRef]
- Matus, F.J.; Rodriguez, J. A simple method for estimating the contribution of nitrogen mineralization to nitrogen supply of crops from a stabilized pool of soil organic matter and recent organic input. Plant Soil 1994, 162, 259–271. [Google Scholar] [CrossRef]
- Iwańska, M.; Paderewski, J.; Stępień, M.; Rodrigues, P.C. Adaptation of winter wheat cultivars to different environments: A case study in Poland. Agronomy 2020, 10, 632. [Google Scholar] [CrossRef]
- Berbell, J.; Martinez-Dalmau, J. A simple agro-economic model for optimal farm nitrogen application under yield uncertainty. Agronomy 2021, 11, 1107. [Google Scholar] [CrossRef]
- Tabak, M.; Lepiarczyk, A.; Filipek–Mazur, B.; Lisowska, A. Efficiency of Nitrogen Fertilization of Winter Wheat Depending on Sulfur Fertilization. Agronomy 2020, 10, 1304. [Google Scholar] [CrossRef]
- Szczepaniak, W.; Nowicki, N.; Bełka, D.; Kazimierowicz, A.; Kulwicki, M.; Grzebisz, W. Effect of foliar application of micronutrients and fungicides on the nitrogen use efficiency in winter wheat. Agronomy 2022, 12, 257. [Google Scholar] [CrossRef]
- MvLellan, E.L.; Cassman, K.G.; Eagle, A.J.; Woodbury, P.B.; Sela, S.; Tonitto, C.; Marjerison, R.D.; van Es, H.M. The nitrogen balancing act: Tracking the environmental performance of food production. Bioscience 2018, 68, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.J.; Blicher-Mathiesen, G.; Rolighed, J.; Andersen, H.E.; Kronvang, B. Three decades of regulation of nitrogen losses: Experiences from the Danish Agricultural Monitoring program. Sci. Total Environ. 2021, 787, 147619. [Google Scholar] [CrossRef] [PubMed]
- Delphin, J.-E. Estimation of nitrogen mineralization in the field from an incubation test and from soil analysis. Agronomie 2000, 20, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-X.; Wang, Z.-H.; Miao, Y.-F.; Li, S.-Q. Soil organic nitrogen and its contribution to crop production. J. Integr. Agric. 2014, 139, 2061–2080. [Google Scholar] [CrossRef]
- Parris, K. Agricultural nutrient balances as agri-environmental indicators: An OECD perspective. Environ. Pollut. 1998, 102 (Suppl. S1), 219–225. [Google Scholar] [CrossRef]
- van Beek, C.L.; Brouver, L.; Oenema, O. The use of farmgate balances and soil surface balances an estimator for nitrogen leaching to surface water. Nutr. Cycl. Agroecosyst. 2003, 67, 233–244. [Google Scholar] [CrossRef]
- Oenema, O.; Kros, H.; de Vries, W. Approaches and uncertainties in nutrient budget: Implications for nutrient managment and environmental policies. Eur. J. Agron. 2003, 20, 3–16. [Google Scholar] [CrossRef]
- Blesh, J.; Drinkwater, L.E. The impact of nitrogen source and crop rotation on nitrogen mass balance in the Mississippi River Basin. Ecol. Appl. 2013, 23, 1017–1035. [Google Scholar] [CrossRef]
- Rayne, N.; Aula, L. Livestock manure and its impacts on soil health: A review. Soil Syst. 2020, 4, 64. [Google Scholar] [CrossRef]
- Ren, T.; Wang, J.; Chen, Q.; Zhang, F.; Lu, S. The effects of manure and nitrogen fertilizer applications on soil organic carbon and nitrogen in a high-input cropping system. PLoS ONE 2014, 9, e97732. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.T.; Fischer, R.A. Yield potential: Its definition, measurement, and significance. Crop Sci. Soc. Am. 1999, 39, 1544–1551. [Google Scholar] [CrossRef]
- Licker, R.; Johnston, M.; Foley, J.A.; Barford, C.; Kucharik, C.J.; Monfreda, C.; Ramankutty, N. Mind the gap: How do climate and agricultural management explain the “yield gap” of croplands around the world? Glob. Ecol. Biogeogr. 2010, 19, 769–782. [Google Scholar] [CrossRef]
- Rabbinge, R. The ecological background of food production. In Crop Production and Sustainable Agriculture; Rabbinge, R., Ed.; John Wiley and Sons: New York, NY, USA, 1993; pp. 2–29. [Google Scholar]
- Grzebisz, W.; Diatta, J. Constrains and solutions to maintain soil productivity, a case study from Central Europe. In Soil Fertility Improvement and Integrated Nutrient Management—A Global Perspective; Whalen, J., Ed.; InTech: London, UK, 2012; pp. 159–183. ISBN 978–953–307–945–5. [Google Scholar]


| Genus/Bacteria Species | Plant Species |
|---|---|
| Rhizobium leguminosarum biovar viciae | Pisum, Viciae, Lathyrus, Lens |
| Rhizobium trifolii | Trifolium |
| Rhizobium phaseoli | Phaseolus |
| Sinorhizobium meliloti | Medicago, Melilotus, Trigonella |
| Bradyrhizobium sp. | Glycine |
| Bradyrhizobium japonicum | Lupinus |
| Real Ranges | Commonly Used Ranges | |
|---|---|---|
| Free living organisms | ||
| Forests | 0–38 | 0–10 |
| Rice | 0–80 | 0–15 |
| Symbiotic microorganisms | ||
| Azolla | 10–150 | 10–50 |
| Fodder legumes | 15–680 | 50–250 |
| Seed legumes | 10–450 | 30–150 |
| Legumes as catch crop | 20–460 | 20–150 |
| Tree/bushes legume plants | 10–30 | 20–50 |
| Soil and Fresh Organic Matter | Carbon, C, % DW | Nitrogen, N, % DW | C:N Ratio |
|---|---|---|---|
| Soil microorganisms | 50 | 5–10 | 5(8):1 |
| Humus | 50 | 5.0 | 10:1 |
| Pig slurry, (6% DW | 50 | 8.0 | 6:1 |
| Dairy slurry (10% DW) | 50 | 4.0 | 12:1 |
| Farmyard manure (25% DW.) | 45 | 3.0 | 15:1 |
| Catch crops of legumes | 40 | 2.5 | 16:1 |
| Catch crops of cruciferous plants | 35 | 2.0 | 18:1 |
| Maize straw | 40 | 1.0 | 40:1 |
| Wheat straw | 40 | 0.5 | 80:1 |
| Biodegradability Classes | C:N | Lignin: N |
|---|---|---|
| High | <18 | <5–4 |
| Moderate | 19–27 | 5–7 |
| Low | 27–60 | 7–15 |
| Very low | > 60 | >15 |
| Soil N Pool—Acronyms | The Pool Description |
|---|---|
| Nmin-s | Mineral N at the beginning of the growing season, |
| Nmin-a | Mineral N at the end of the growing season |
| Nf | Fertilizer N |
| Nfyn1-3 | N released from the applied manure in consecutive years following year of application |
| Nfix | N2 fixed by microorganisms |
| Nrel | N released from soil resources during the growing season |
| NY | N in the main yield |
| NCRs | N in crop plant byproduct |
| Nuw | Un-workable N, a part of Nf which was not transferred into the yield |
| Nl | Nitrogen lost from the field |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzebisz, W.; Niewiadomska, A.; Przygocka-Cyna, K. Nitrogen Hotspots on the Farm—A Practice-Oriented Approach. Agronomy 2022, 12, 1305. https://doi.org/10.3390/agronomy12061305
Grzebisz W, Niewiadomska A, Przygocka-Cyna K. Nitrogen Hotspots on the Farm—A Practice-Oriented Approach. Agronomy. 2022; 12(6):1305. https://doi.org/10.3390/agronomy12061305
Chicago/Turabian StyleGrzebisz, Witold, Alicja Niewiadomska, and Katarzyna Przygocka-Cyna. 2022. "Nitrogen Hotspots on the Farm—A Practice-Oriented Approach" Agronomy 12, no. 6: 1305. https://doi.org/10.3390/agronomy12061305
APA StyleGrzebisz, W., Niewiadomska, A., & Przygocka-Cyna, K. (2022). Nitrogen Hotspots on the Farm—A Practice-Oriented Approach. Agronomy, 12(6), 1305. https://doi.org/10.3390/agronomy12061305