
Marker-Assisted Backcross Breeding for Improving Bruchid (Callosobruchus spp.) Resistance in Mung Bean (Vigna radiata L.)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parental Materials
2.2. Marker-Assisted Backcross (MABC) Breeding Strategy for Bruchid Resistance
2.3. DNA Extraction and Polymerase Chain Reaction (PCR) Amplification
2.4. RNA Extraction and cDNA Synthesis
2.5. Evaluation of Bruchid Resistance
- (1)
- Highly resistant (HR): 0% to 10%.
- (2)
- Resistant (R): 10% to 35%.
- (3)
- Moderately resistant (MR): 35% to 65%.
- (4)
- Susceptible (S): 65% to 90%.
- (5)
- Highly susceptible: 90% to 100%.
2.6. Investigation of Agronomic Traits
2.7. Genetic Background Analysis
3. Results
3.1. Development of Bruchid-Resistant Lines through MABC Breeding
3.2. R67-11, R67-22, and R92-15 Lines All Showed Resistance to Bruchid
3.3. The Agronomic Traits of Bruchid-Resistant Lines
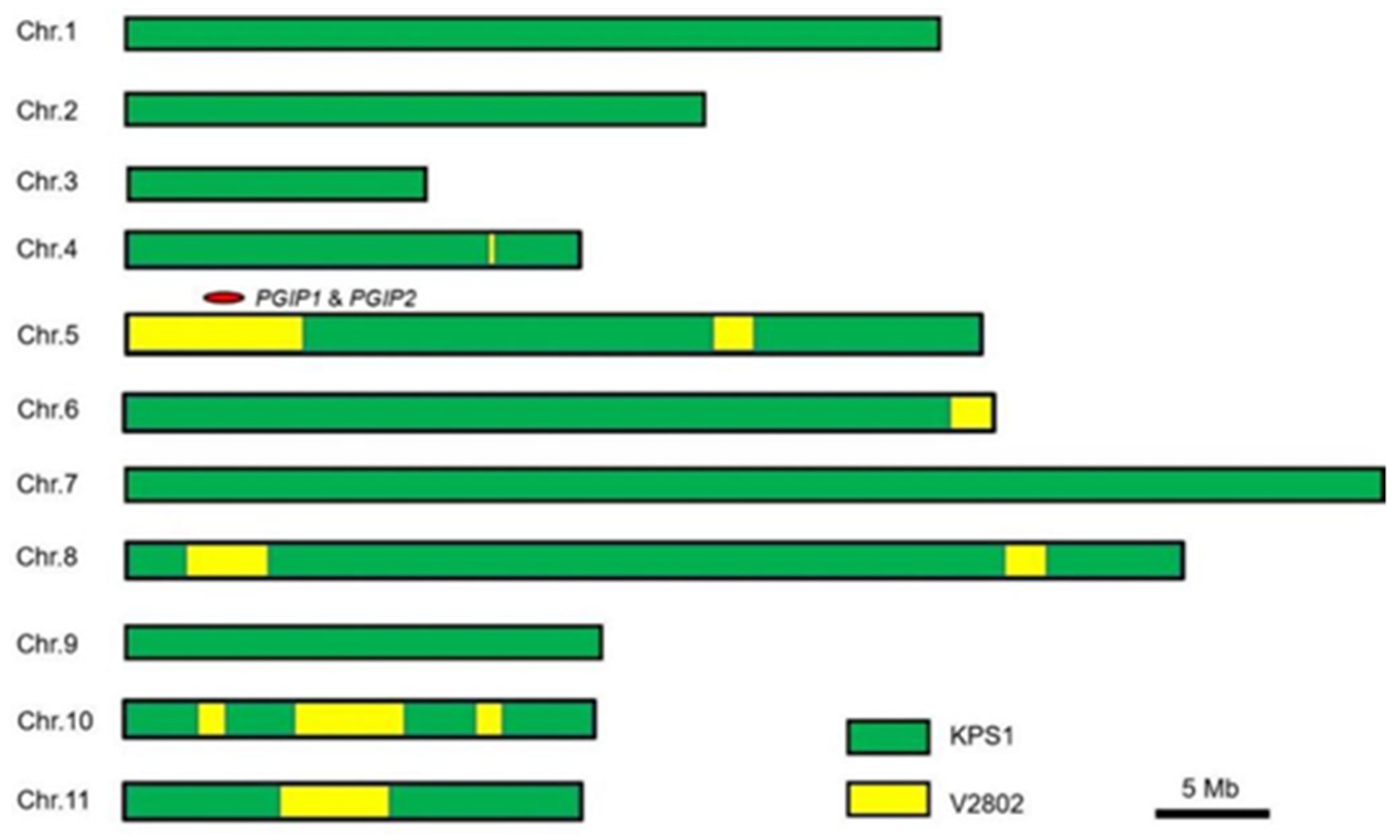
3.4. The Genetic Background of R67-22 Showed Highly Similarity to KPS1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomooka, N.; Vaughan, D.A.; Moss, H.; Maxted, N. The Asian Vigna: Genus Vigna Subgenus Ceratotropis Genetic Resources; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Nair, R.; Schreinemachers, P. Global Status and Economic Importance of Mungbean. In The Mungbean Genome; Nair, R.M., Schafleitner, R., Lee, S.-H., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–8. [Google Scholar]
- Somta, P.; Srinives, P. Genome research in mungbean (Vigna radiata (L.) Wilczek) and blackgram (V. mungo (L.) Hepper). ScienceAsia 2007, 33, 69–74. [Google Scholar] [CrossRef]
- Lambrides, C.J.; Godwin, I.D. Mungbean. In Pulses, Sugar and Tuber Crops. Genome Mapping & Molecular Breeding in Plants; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 3, pp. 69–90. [Google Scholar]
- Wang, L.X.; Wang, S.H.; Luo, G.L.; Zhang, J.T.; Chen, Y.H.; Chen, H.L.; Cheng, X.Z. Evaluation of the production potential of mung bean cultivar “Zhonglv 5”. Agronomy 2022, 12, 707. [Google Scholar] [CrossRef]
- Mandal, B.K.; Dhara, M.C.; Mandal, B.B.; Das, S.K.; Nandy, R. Rice, mungbean, soybean, peanut, ricebean, and blackgram yields under different intercropping systems. Agron. J. 1990, 82, 1063–1066. [Google Scholar] [CrossRef]
- Laosatit, K.; Somta, P.; Xin, C.; Srinives, P. Approaches to Biotic Stresses. In The Mungbean Genome; Nair, R.M., Schafleitner, R., Lee, S.-H., Eds.; Springer: Cham, Switzerland, 2020; pp. 133–167. [Google Scholar]
- Batzer, J.C.; Singh, A.; Rairdin, A.; Chiteri, K.; Mueller, D.S. Mungbean: A preview of disease management challenges for an alternative U.S. cash crop. J. Integr. Pest. Manag. 2022, 13, 4. [Google Scholar] [CrossRef]
- Basu, P.S.; Pratap, A.; Gupta, S.; Sharma, K.; Tomar, R.; Singh, N.P. Physiological traits for shortening crop duration and improving productivity of greengram (Vigna radiata L. Wilczek) under high temperature. Front. Plant Sci. 2019, 10, 1508. [Google Scholar] [CrossRef] [Green Version]
- Southgate, B.J. Biology of the Bruchidae. Annu. Rev. Entomol. 1979, 24, 449–473. [Google Scholar] [CrossRef]
- Lambrides, C.J.; Imrie, B.C. Susceptibility of mungbean varieties to the bruchid species Callosobruchus maculatus (F.), C.phaseoli (Gyll.), C.chinensis (L.), and Acanthoscelides obtectus (Say.) (Coleoptera: Chrysomelidae). Aust. J. Agric. Res. 2000, 51, 85–89. [Google Scholar] [CrossRef]
- Somta, C.; Somta, P.; Tomooka, N.; Ooi, P.a.C.; Vaughan, D.A.; Srinives, P. Characterization of new sources of mungbean (Vigna radiata (L.) Wilczek) resistance to bruchids, Callosobruchus spp. (Coleoptera: Bruchidae). J. Stored Prod. Res. 2008, 44, 316–321. [Google Scholar] [CrossRef]
- Somta, P. Genetics and breeding of resistance to bruchids (Callosobruchus spp.) in Vigna Crops: A review. NU. Int. J. Sci. 2007, 4, 1–17. [Google Scholar]
- Mishra, S.K.; Macedo, M.L.R.; Panda, S.K.; Panigrahi, J. Bruchid pest management in pulses: Past practices, present status and use of modern breeding tools for development of resistant varieties. Ann. Appl. Biol. 2018, 172, 4–19. [Google Scholar] [CrossRef]
- Fujii, K.; Miyazaki, S. Infestation resistance of wild legumes (Vigna Sublobata) to azuki bean weevil, Callosobruchus Chinensis (L.) (Coleoptera, Bruchidae) and its relationship with cytogenetic classification. Appl. Entomol. Zool. 1987, 22, 229–230. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Ishimotob, M.; Yamanakaa, N.; Miyazaki, S.; Hiramatsu, M.; Nakajima, Y.; Hirano, K.; Krtamura, K.; Miyazaki, S. Effects of bruchid-resistant mungbean meal on growth and blood-biochemical values in mice. JIRCAS J. 1996, 3, 23–31. [Google Scholar]
- Watanasit, A.; Pichitporn, S. Improvement of mung-bean for resistance to bruchids. In Mungbean Germ-Plasm: Collection, Evaluation and Utilization Forbreeding Program; Srinives, P., Kitbamroong, C., Miyazaki, S., Eds.; Japan International Research Center for Agricultural Sciences: Tsukuba, Japan, 1996; pp. 67–71. [Google Scholar]
- Talekar, N.S.; Lin, C.P. Characterization of Callosobruchus Chinensis (Coleoptera, Bruchidae) resistance in mungbean. J. Econ. Entomol. 1992, 85, 1150–1153. [Google Scholar] [CrossRef]
- Somta, P.; Ammaranan, C.; Ooi, P.a.C.; Srinives, P. Inheritance of seed resistance to bruchids in cultivated mungbean (Vigna radiata, L. Wilczek). Euphytica 2007, 155, 47–55. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B 2008, 363, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Ghosh, S.; Chatterjee, M.; Das, P.; Lahari, T.; Maji, A.; Mondal, N.; Pradhan, K.K.; Bhattacharyya, S. Molecular markers linked with bruchid resistance in Vigna radiata var. sublobata and their validation. J. Plant Biochem. Biotechnol. 2011, 20, 155–160. [Google Scholar] [CrossRef]
- Chotechung, S.; Chankaew, S.; Srinives, P.; Somta, P. Identification of DNA markers associated with bruchid resistance in mungbean. Khon Khan Agric. J. 2011, 39, 221–226, (In Thai with English Abstract). [Google Scholar]
- Chotechung, S.; Somta, P.; Chen, J.; Yimram, T.; Chen, X.; Srinives, P. A gene encoding a polygalacturonase-inhibiting protein (PGIP) is a candidate gene for bruchid (Coleoptera: Bruchidae) resistance in mungbean (Vigna radiata). Theor. Appl. Genet. 2016, 129, 1673–1683. [Google Scholar] [CrossRef]
- Kaewwongwal, A.; Chen, J.B.; Somta, P.; Kongjaimun, A.; Yimram, T.; Chen, X.; Srinives, P. Novel alleles of two tightly linked genes encoding polygalacturonase-inhibiting proteins (VrPGIP1 and VrPGIP2) associated with the Br. locus that confer bruchid (Callosobruchus spp.) resistance to mungbean (Vigna radiata) accession V2709. Front. Plant Sci. 2017, 8, 1692. [Google Scholar] [CrossRef]
- Kaewwongwal, A.; Liu, C.Y.; Somta, P.; Chen, J.B.; Tian, J.; Yuan, X.X.; Chen, X. A second VrPGIP1 allele is associated with bruchid resistance (Callosobruchus spp.) in wild mungbean (Vigna radiata var. sublobata) accession ACC41. Mol. Genet. Genom. 2020, 295, 275–286. [Google Scholar] [CrossRef]
- Zhang, Q.X.; Yan, Q.; Yuan, X.X.; Lin, Y.; Chen, J.B.; Wu, R.R.; Xue, C.C.; Zhu, Y.L.; Chen, X. Two polygalacturonase-inhibiting proteins (VrPGIP) of Vigna radiata confer resistance to bruchids (Callosobruchus spp.). J. Plant Physiol. 2021, 258–259, 153376. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.; Melchinger, A.E. Selection theory for marker-assisted backcrossing. Genetics 2005, 170, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Cheng, X.Z.; Fan, B.J.; Wang, L.X.; Liu, J.J.; Liu, C.Y.; Wang, S.H.; Cao, Z.M.; Chen, H.L.; Wang, Y.; et al. Current situation and development trend of mung bean varieties in China. Crops 2021, 6, 15–21. (In Chinese) [Google Scholar]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular-weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Laosatit, K.; Chen, J.B.; Yuan, X.X.; Wu, R.R.; Amkul, K.; Chen, X.; Somta, P. Mapping and functional characterization of Stigma Exposed 1, a DUF1005 gene controlling petal and stigma cells in mungbean (Vigna radiata). Front. Plant Sci. 2020, 11, 575922. [Google Scholar] [CrossRef]
- Cheng, X.Z.; Wang, S.H.; Wang, L.X. Descriptors and Data Standard for Mungbean [Vigna radiata (L.) Wilczek]; China Agriculture Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Lee, Y.H.; Moon, J.K.; Park, K.Y.; Ku, J.H.; Chung, M.N. A new mungbean cultivar with bruchid resistance, ‘Jangannogdu’. Korean J. Breed. 2000, 32, 296–297. [Google Scholar]
- Fox, C.W.; Bush, M.L.; Messina, F.J. Biotypes of the seed beetle Callosobruchus maculatus have differing effects on the germination and growth of their legume hosts. Agric. For. Entomol. 2010, 12, 353–362. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Nayak, D.K.; Pandit, E.; Behera, L.; Anandan, A.; Mukherjee, A.K.; Lenka, S.; Barik, D.P. Incorporation of bacterial blight resistance genes into lowland rice cultivar through marker-assisted backcross breeding. Phytopathology 2016, 106, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Rafii, M.Y.; Ismail, M.R.; Mahmood, M.; Rahim, H.A.; Alam, M.A.; Ashkani, S.; Malek, M.A.; Latif, M.A. Marker-assisted backcrossing: A useful method for rice improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef] [Green Version]
- Mallick, N.; Sharma, J.B.; Tomar, R.S.; Sivasamy, M.; Prabhu, K.V. Marker-assisted backcross breeding to combine multiple rust resistance in wheat. Plant Breed. 2015, 134, 172–177. [Google Scholar] [CrossRef]
- Yang, L.Q.; Wang, W.; Yang, W.P.; Wang, M.C. Marker-assisted selection for pyramiding the waxy and opaque-16 genes in maize using cross and backcross schemes. Mol. Breed. 2013, 31, 767–775. [Google Scholar] [CrossRef]
- Maranna, S.; Verma, K.; Talukdar, A.; Lal, S.K.; Kumar, A.; Mukherjee, K. Introgression of null allele of Kunitz trypsin inhibitor through marker-assisted backcross breeding in soybean (Glycine max L. Merr.). BMC Genet. 2016, 17, 106. [Google Scholar] [CrossRef] [Green Version]
- Papan, P.; Chueakhunthod, W.; Jinagool, W.; Tharapreuksapong, A.; Masari, A.; Kaewkasi, C.; Ngampongsai, S.; Girdthai, T.; Tantasawat, P.A. Improvement of Cercospora leaf spot and powdery mildew resistance of mungbean variety KING through marker-assisted selection. J. Agric. Sci. 2022, 159, 676–687. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Developmental Period (Day) | No. of Adults That Emerged | Percentage of Damaged Seeds (%) | Resistance Level |
|---|---|---|---|---|
| R67-11 | 44.33 ± 0.94 a | 9.67 ± 2.87 bc | 10 ± 3.27 c | HR |
| R67-22 | 44.67 ± 1.25 a | 3.33 ± 1.25 c | 6.67 ± 2.49 c | HR |
| R92-15 | 40.67 ± 1.25 b | 11.67 ± 5.31 bc | 13.33 ± 0.94 c | R |
| KPS1 | 25.67 ± 0.94 d | 125.67 ± 14.88 a | 100 ± 0 a | HS |
| V2802 | 32.33 ± 1.25 c | 24.33 ± 4.99 b | 42.67 ± 7.72 b | MR |
| Mean | 37.53 | 34.93 | 34.53 | - |
| CV | 20.68% | 137.76% | 106.04% | - |
| Genotype | Plant Height/cm | Branch Number | Main Stem Node Number | Seed Number per Pod | 100 Seed Weight/g | Seed Yield per Plant/g |
|---|---|---|---|---|---|---|
| R67-11 | 80.71 ± 3.93 b | 2.92 ± 0.64 b | 13.75 ± 1.09 b | 10.81 ± 0.52 b | 6.12 ± 0.30 b | 42.2 ± 5.04 c |
| R67-22 | 70.33 ± 5.32 c | 2.67 ± 0.47 bc | 13.5 ± 0.87 bc | 10.56 ± 0.59 bc | 6.59 ± 0.15 a | 47.1 ± 2.76 ab |
| R92-15 | 71.25 ±7.03 c | 2.50 ± 0.5 bc | 13.50 ± 0.87 bc | 10.21 ± 0.66 c | 6.23 ± 0.24 b | 39.1 ± 3.54 d |
| KPS1 | 71.63 ± 10.61 c | 2.42 ± 0.49 c | 12.17 ± 1.34 c | 10.48 ± 0.54 bc | 6.83 ± 0.34 a | 49.8 ± 4.49 a |
| V2802 | 118.92 ± 8.36 a | 4.67 ± 0.62 a | 20.17 ± 1.78 a | 12.17 ± 0.51 a | 4.37 ± 0.16 c | 34.5 ± 5.48 e |
| Mean | 82.58 | 3.03 | 14.62 | 10.85 | 6.03 | 42.40 |
| CV | 24.41% | 33.23% | 21.31% | 8.28% | 15.07% | 17.04% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, R.; Zhang, Q.; Lin, Y.; Chen, J.; Somta, P.; Yan, Q.; Xue, C.; Liu, J.; Chen, X.; Yuan, X. Marker-Assisted Backcross Breeding for Improving Bruchid (Callosobruchus spp.) Resistance in Mung Bean (Vigna radiata L.). Agronomy 2022, 12, 1271. https://doi.org/10.3390/agronomy12061271
Wu R, Zhang Q, Lin Y, Chen J, Somta P, Yan Q, Xue C, Liu J, Chen X, Yuan X. Marker-Assisted Backcross Breeding for Improving Bruchid (Callosobruchus spp.) Resistance in Mung Bean (Vigna radiata L.). Agronomy. 2022; 12(6):1271. https://doi.org/10.3390/agronomy12061271
Chicago/Turabian StyleWu, Ranran, Qinxue Zhang, Yun Lin, Jingbin Chen, Prakit Somta, Qiang Yan, Chenchen Xue, Jinyang Liu, Xin Chen, and Xingxing Yuan. 2022. "Marker-Assisted Backcross Breeding for Improving Bruchid (Callosobruchus spp.) Resistance in Mung Bean (Vigna radiata L.)" Agronomy 12, no. 6: 1271. https://doi.org/10.3390/agronomy12061271
APA StyleWu, R., Zhang, Q., Lin, Y., Chen, J., Somta, P., Yan, Q., Xue, C., Liu, J., Chen, X., & Yuan, X. (2022). Marker-Assisted Backcross Breeding for Improving Bruchid (Callosobruchus spp.) Resistance in Mung Bean (Vigna radiata L.). Agronomy, 12(6), 1271. https://doi.org/10.3390/agronomy12061271