
A Gene Encoding a SHINE1/WAX INDUCER1 Transcription Factor Controls Cuticular Wax in Barley


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Cuticle Integrity Measurements
2.3. Genotyping and Candidate Gene Sequencing
2.4. Gene Expression
2.5. Wax Quantification
2.6. Sequence Retrieval and Phylogeny
2.7. Haplotype Analysis
3. Results
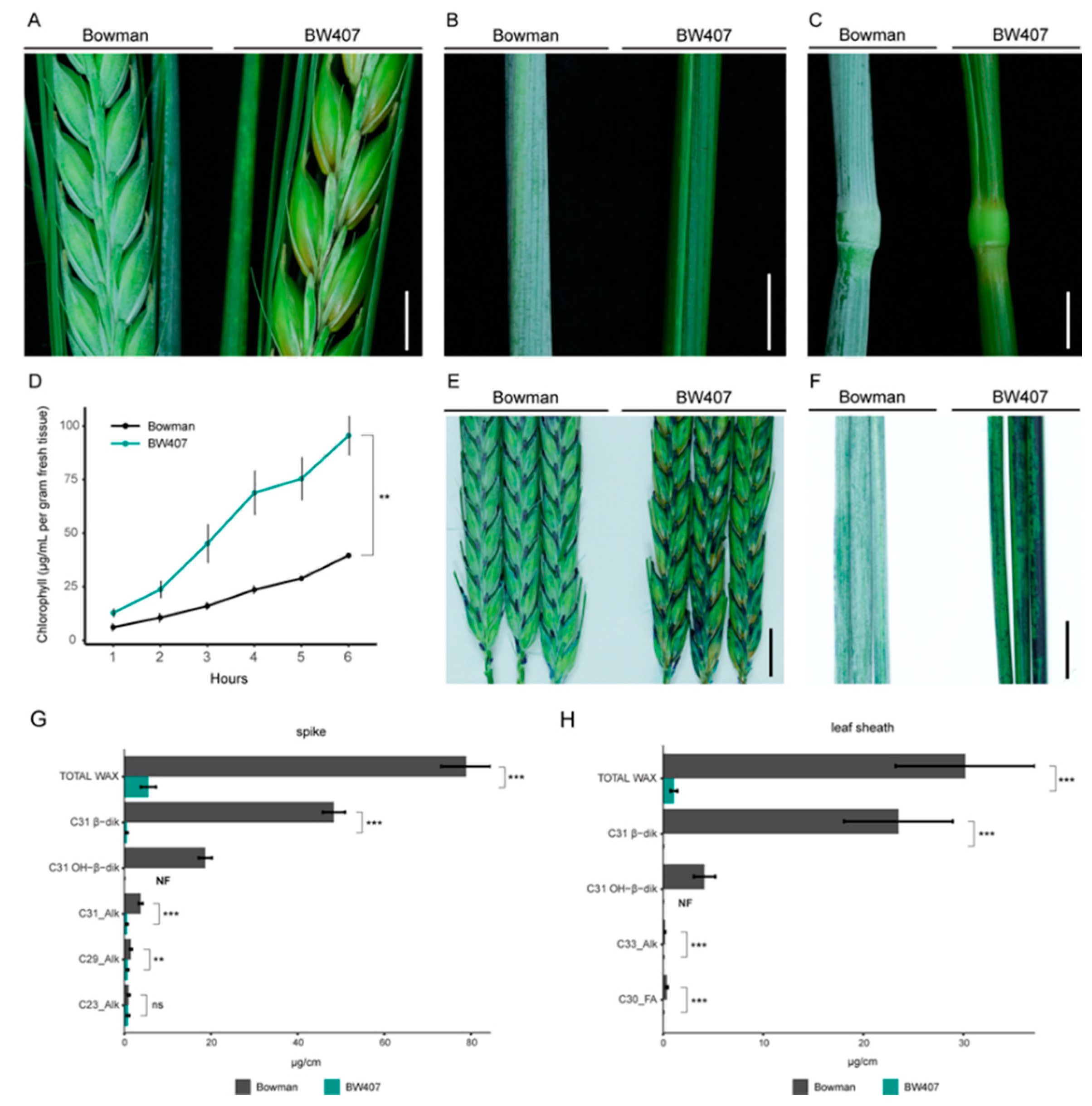
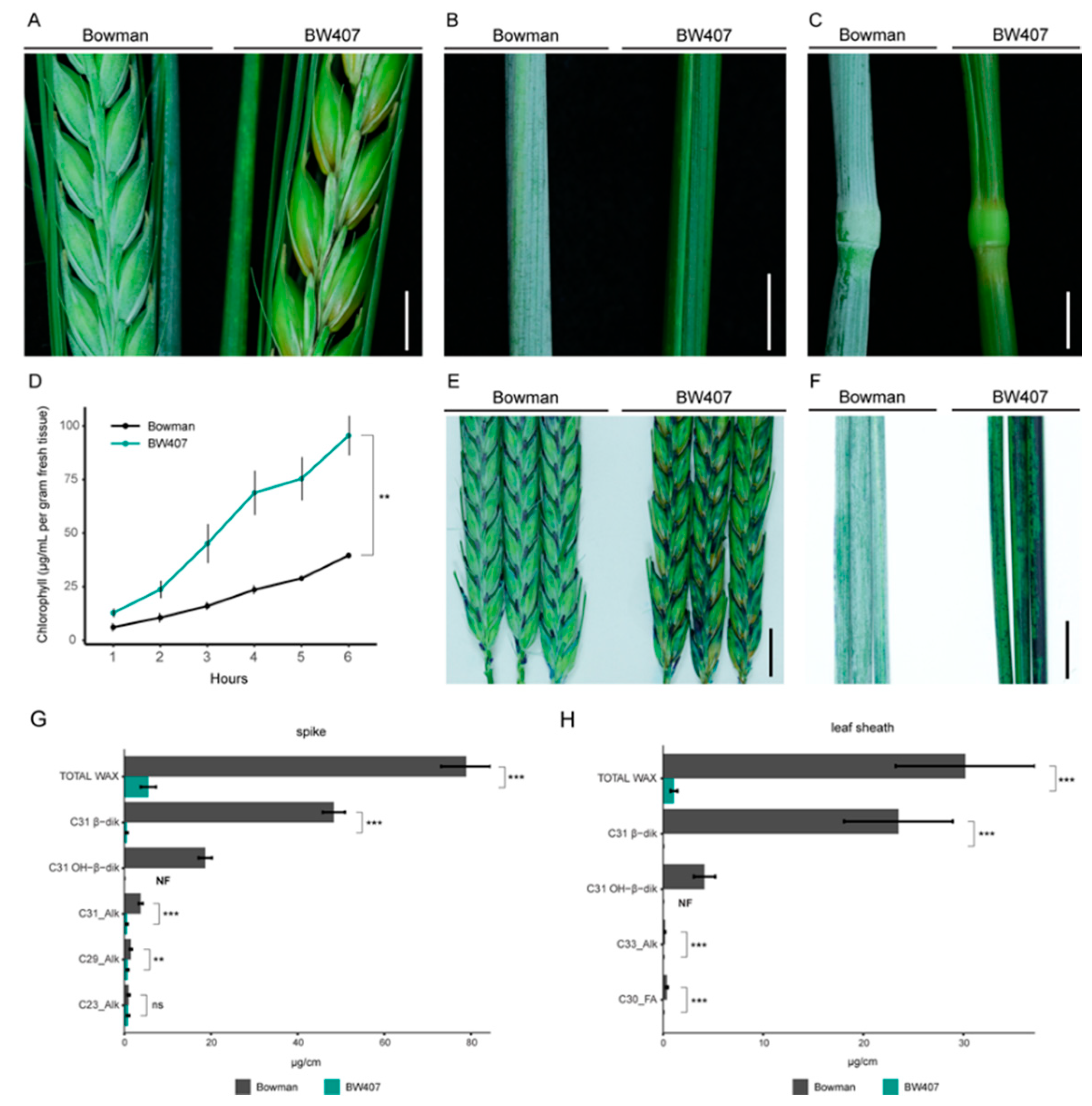
3.1. CER-X Controls Cuticular Integrity and Epicuticular Wax Composition
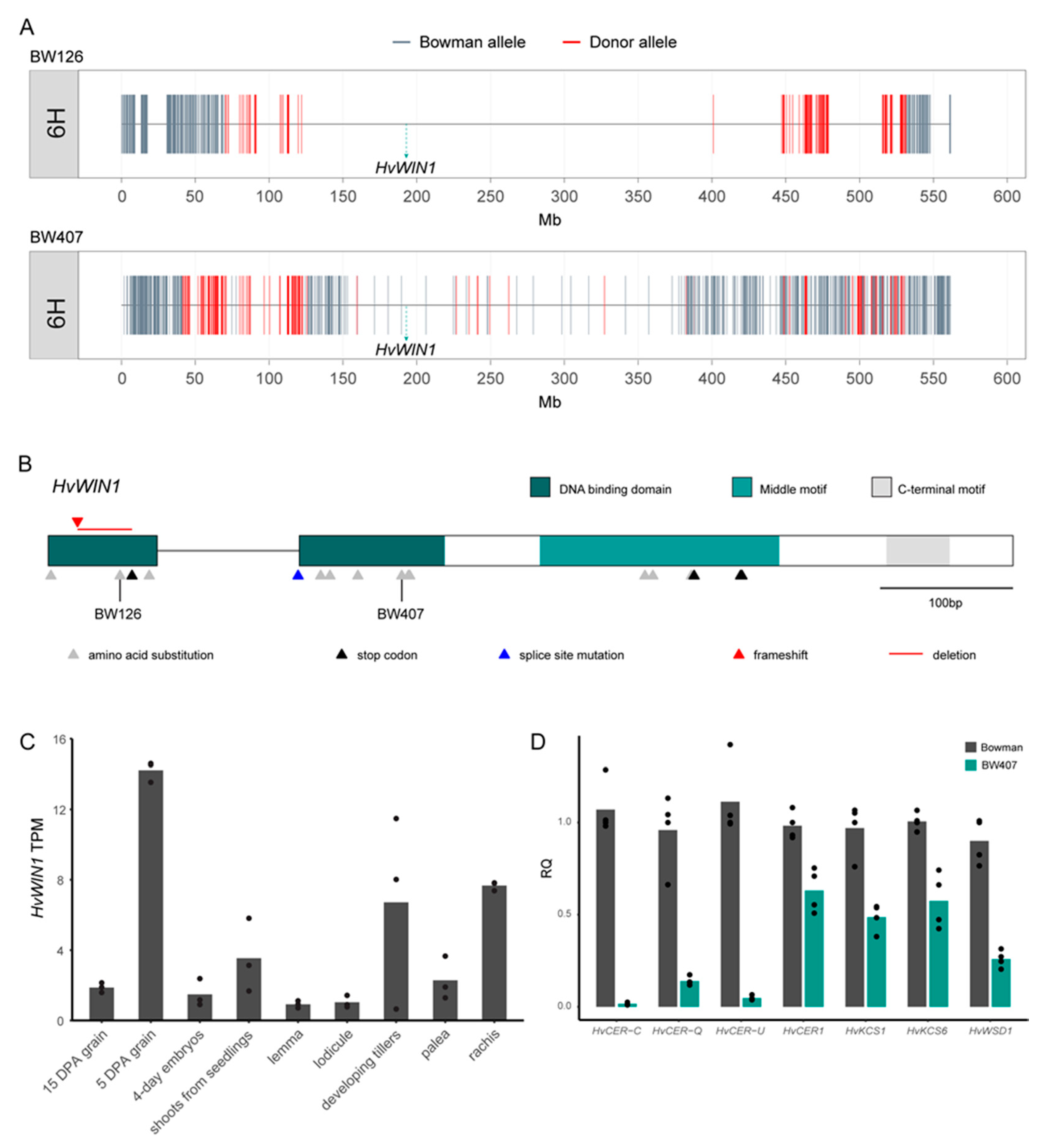
3.2. CER-X Is HvWIN1
3.3. HvWIN1 Influences Gene Expression Associated with Cuticle Development
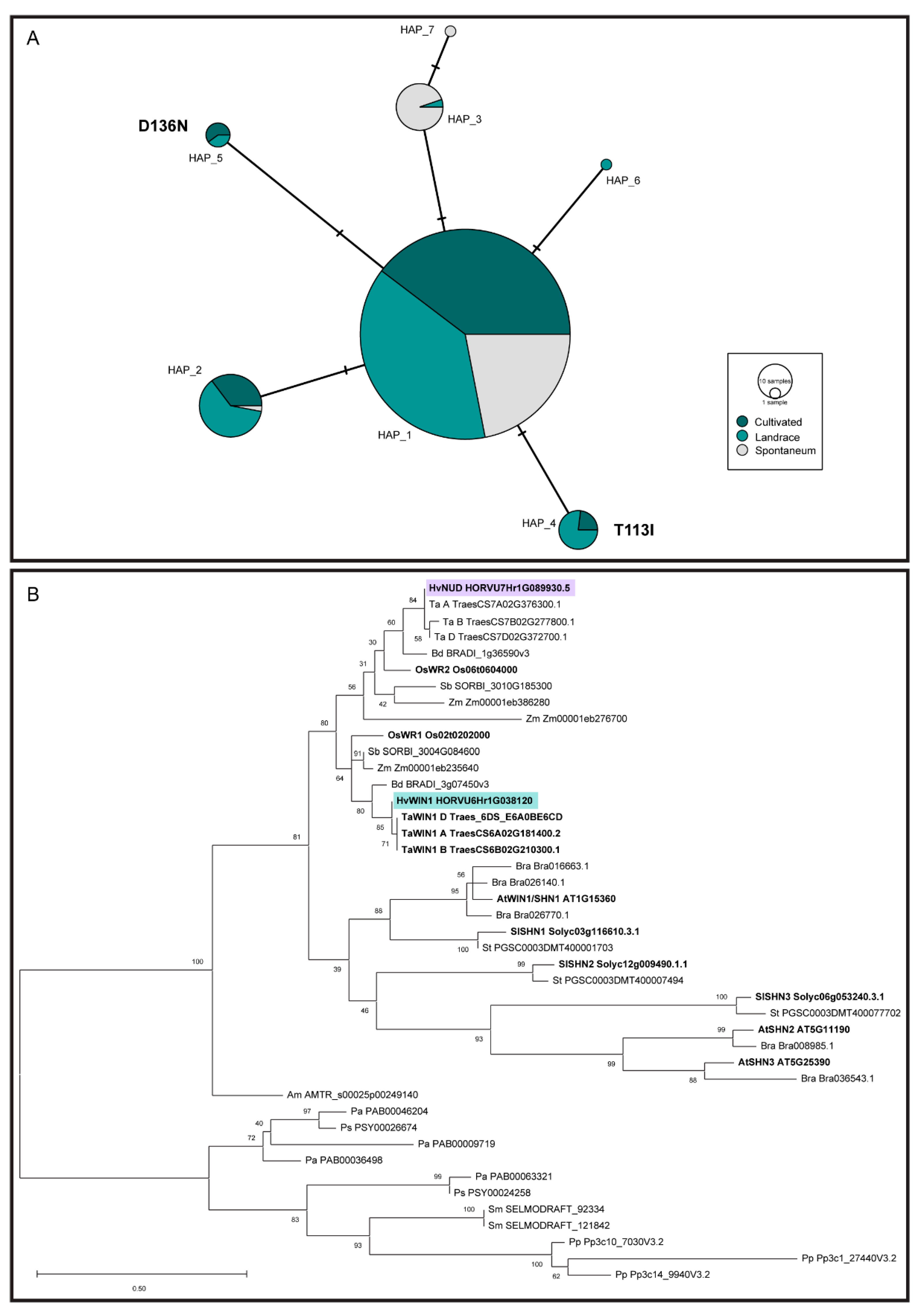
3.4. HvWIN1 Is Part of a Highly Conserved Gene Family
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yeats, T.H.; Rose, J.K.C. The Formation and Function of Plant Cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffree, C.E. The Fine Structure of the Plant Cuticle. Ann. Plant Rev. 2006, 23, 11–125. [Google Scholar]
- Shepherd, T.; Wynne Griffiths, D. The Effects of Stress on Plant Cuticular Waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, P.; Fernández, V.; García, M.L.; Khayet, M.; Fernández, A.; Gil, L. Localization of Polysaccharides in Isolated and Intact Cuticles of Eucalypt, Poplar and Pear Leaves by Enzyme-Gold Labelling. Plant Physiol. Biochem. 2014, 76, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuels, L.; DeBono, A.; Lam, P.; Wen, M.; Jetter, R.; Kunst, L. Use of Arabidopsis Eceriferum Mutants to Explore Plant Cuticle Biosynthesis. J. Vis. Exp. 2008, 16, e709. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. (Ed.) Kolattukudy PE Biochemistry of Plant Waxes. In Chemistry and Biochemistry of Natural Waxes; Elsevier: Amsterdam, The Netherlands, 1976; pp. 289–347. [Google Scholar]
- Xue, D.; Zhang, X.; Lu, X.; Chen, G.; Chen, Z.-H. Molecular and Evolutionary Mechanisms of Cuticular Wax for Plant Drought Tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef]
- Edwards, D.; Kenrick, P. The Early Evolution of Land Plants, from Fossils to Genomics: A Commentary on Lang (1937) “On the Plant-Remains from the Downtonian of England and Wales”. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140343. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, E.; Heredia-Guerrero, J.A.; Heredia, A. The Plant Cuticle: Old Challenges, New Perspectives. J. Exp. Bot. 2017, 68, 5251–5255. [Google Scholar] [CrossRef]
- Gosney, B.J.; Potts, B.M.; O’Reilly-Wapstra, J.M.; Vaillancourt, R.E.; Fitzgerald, H.; Davies, N.W.; Freeman, J.S. Genetic Control of Cuticular Wax Compounds in Eucalyptus Globulus. New Phytol. 2016, 209, 202–215. [Google Scholar] [CrossRef]
- Jetter, R.; Kunst, L.; Samuels, A.L. Composition of Plant Cuticular Waxes. In Annual Plant Reviews Volume 23: Biology of the Plant Cuticle; Wiley: Hoboken, NJ, USA, 2006; pp. 145–181. [Google Scholar]
- Buschhaus, C.; Jetter, R. Composition Differences between Epicuticular and Intracuticular Wax Substructures: How Do Plants Seal Their Epidermal Surfaces? J. Exp. Bot. 2011, 62, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.; Kovalchuk, N.; Langridge, P.; Tricker, P.J.; Lopato, S.; Borisjuk, N. The Impact of Drought on Wheat Leaf Cuticle Properties. BMC Plant Biol. 2017, 17, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, W.; Li, W. Genetic Interactions Underlying the Biosynthesis and Inhibition of β-Diketones in Wheat and Their Impact on Glaucousness and Cuticle Permeability. PLoS ONE 2013, 8, e54129. [Google Scholar]
- Laskoś, K.; Czyczyło-Mysza, I.M.; Dziurka, M.; Noga, A.; Góralska, M.; Bartyzel, J.; Myśków, B. Correlation between Leaf Epicuticular Wax Composition and Structure, Physio-Biochemical Traits and Drought Resistance in Glaucous and Non-Glaucous near-Isogenic Lines of Rye. Plant J. 2021, 108, 93–119. [Google Scholar] [CrossRef]
- Guo, J.; Xu, W.; Yu, X.; Shen, H.; Li, H.; Cheng, D.; Liu, A.; Liu, J.; Liu, C.; Zhao, S.; et al. Cuticular Wax Accumulation Is Associated with Drought Tolerance in Wheat Near-Isogenic Lines. Front. Plant Sci. 2016, 7, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, R.S.; Poveda, M.M. Environmental Gradients and Population Divergence Contribute to Variation in Cuticular Wax Composition in Juniperus Communis. Biochem. Syst. Ecol. 2003, 31, 1257–1270. [Google Scholar] [CrossRef]
- Tulloch, A.P.; Hoffman, L.L. Epicuticular Wax of Panicum Virgatum. Phytochemistry 1980, 19, 837–839. [Google Scholar] [CrossRef]
- Richards, R.A.; Rawson, H.M.; Johnson, D.A. Glaucousness in Wheat: Its Development and Effect on Water-Use Efficiency, Gas Exchange and Photosynthetic Tissue Temperatures. Funct. Plant Biol. 1986, 13, 465–473. [Google Scholar] [CrossRef]
- González, A.; Ayerbe, L. Effect of Terminal Water Stress on Leaf Epicuticular Wax Load, Residual Transpiration and Grain Yield in Barley. Euphytica 2010, 172, 341–349. [Google Scholar] [CrossRef]
- Febrero, A.; Fernández, S.; Molina-Cano, J.L.; Araus, J.L. Yield, Carbon Isotope Discrimination, Canopy Reflectance and Cuticular Conductance of Barley Isolines of Differing Glaucousness. J. Exp. Bot. 1998, 49, 1575–1581. [Google Scholar] [CrossRef]
- Hen-Avivi, S.; Savin, O.; Racovita, R.C.; Lee, W.-S.; Adamski, N.M.; Malitsky, S.; Almekias-Siegl, E.; Levy, M.; Vautrin, S.; Bergès, H.; et al. A Metabolic Gene Cluster in the Wheat W1 and the Barley Cer-Cqu Loci Determines β-Diketone Biosynthesis and Glaucousness. Plant Cell 2016, 28, 1440–1460. [Google Scholar] [CrossRef] [Green Version]
- Nice, L.M.; Steffenson, B.J.; Brown-Guedira, G.L.; Akhunov, E.D.; Liu, C.; Kono, T.J.Y.; Morrell, P.L.; Blake, T.K.; Horsley, R.D.; Smith, K.P.; et al. Development and Genetic Characterization of an Advanced Backcross-Nested Association Mapping (AB-NAM) Population of Wild × Cultivated Barley. Genetics 2016, 203, 1453–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundqvist, U.; Wettstein, D. Induction of Eceriferum Mutants in Barley by Ionizing Radiations and Chemical Mutagens. Hereditas 1962, 48, 342–362. [Google Scholar] [CrossRef]
- Lundqvist, U.; Lundqvist, A. Mutagen Specificity in Barley for 1580 Eceriferum Mutants Localized to 79 Loci. Hereditas 1988, 108, 1–12. [Google Scholar] [CrossRef]
- Li, C.; Haslam, T.M.; Kr�ger, A.; Schneider, L.M.; Mishina, K.; Samuels, L.; Yang, H.; Kunst, L.; Schaffrath, U.; Nawrath, C.; et al. The β-Ketoacyl-CoA Synthase HvKCS1, Encoded by Cer-Zh, Plays a Key Role in Synthesis of Barley Leaf Wax and Germination of Barley Powdery Mildew. Plant Cell Physiol. 2018, 59, 811–827. [Google Scholar] [CrossRef]
- Weidenbach, D.; Jansen, M.; Franke, R.B.; Hensel, G.; Weissgerber, W.; Ulferts, S.; Jansen, I.; Schreiber, L.; Korzun, V.; Pontzen, R. Evolutionary Conserved Function of Barley and Arabidopsis 3-KETOACYL-CoA SYNTHASES in Providing Wax Signals for Germination of Powdery Mildew Fungi. Plant Physiol. 2014, 166, 1621–1633. [Google Scholar] [CrossRef] [Green Version]
- von Wettstein-Knowles, P. The Polyketide Components of Waxes and the Cer-Cqu Gene Cluster Encoding a Novel Polyketide Synthase, the β-Diketone Synthase, DKS. Plants 2017, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.; von Wettstein-Knowles, P. Structure of Epicuticular Waxes on Spikes and Leaf Sheaths of Barley as Revealed by a Direct Platinum Replica Technique. Carlsberg Res. Commun. 1980, 45, 465–481. [Google Scholar] [CrossRef] [Green Version]
- Schneider, L.M.; Adamski, N.M.; Christensen, C.E.; Stuart, D.B.; Vautrin, S.; Hansson, M.; Uauy, C.; von Wettstein-Knowles, P. The Cer-Cqu Gene Cluster Determines Three Key Players in a β-Diketone Synthase Polyketide Pathway Synthesizing Aliphatics in Epicuticular Waxes. J. Exp. Bot. 2016, 67, 2715–2730. [Google Scholar] [CrossRef] [Green Version]
- von Wettstein-Knowles, P. Genetic Control of β-Diketone and Hydroxy-β-Diketone Synthesis in Epicuticular Waxes of Barley. Planta 1972, 106, 113–130. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE Clade of AP2 Domain Transcription Factors Activates Wax Biosynthesis, Alters Cuticle Properties, and Confers Drought Tolerance When Overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.X.; Malitsky, S.; De Oliveira, S.; Branigan, C.; Franke, R.B.; Schreiber, L.; Aharoni, A. SHINE Transcription Factors Act Redundantly to Pattern the Archetypal Surface of Arabidopsis Flower Organs. PLoS Genet. 2011, 7, e1001388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, Y.; Shikata, M.; Koyama, T.; Ohtsubo, N.; Mitsuda, N.; Ohme-Takagi, M. MIXTA-Like Transcription Factors and WAX INDUCER1/SHINE1 Coordinately Regulate Cuticle Development in Arabidopsis and Torenia Fournieri. Plant Cell 2013, 25, 1609–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannangara, R.; Branigan, C.; Liu, Y.; Penfield, T.; Rao, V.; Mouille, G.; Höfte, H.; Pauly, M.; Riechmann, J.L.; Broun, P. The Transcription Factor WIN1/SHN1 Regulates Cutin Biosynthesis in Arabidopsis Thaliana. Plant Cell 2007, 19, 1278–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broun, P.; Poindexter, P.; Osborne, E.; Jiang, C.-Z.; Riechmann, J.L. WIN1, a Transcriptional Activator of Epidermal Wax Accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 4706–4711. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wan, L.; Zhang, L.; Zhang, Z.; Zhang, H.; Quan, R.; Zhou, S.; Huang, R. An Ethylene Response Factor OsWR1 Responsive to Drought Stress Transcriptionally Activates Wax Synthesis Related Genes and Increases Wax Production in Rice. Plant Mol. Biol. 2012, 78, 275–288. [Google Scholar] [CrossRef]
- Djemal, R.; Khoudi, H. Isolation and Molecular Characterization of a Novel WIN1/SHN1 Ethylene-Responsive Transcription Factor TdSHN1 from Durum Wheat (Triticum Turgidum. L. Subsp. Durum). Protoplasma 2015, 252, 1461–1473. [Google Scholar] [CrossRef]
- Li, X.; Liu, N.; Sun, Y.; Wang, P.; Ge, X.; Pei, Y.; Liu, D.; Ma, X.; Li, F.; Hou, Y. The Cotton GhWIN2 Gene Activates the Cuticle Biosynthesis Pathway and Influences the Salicylic and Jasmonic Acid Biosynthesis Pathways. BMC Plant Biol. 2019, 19, 379. [Google Scholar] [CrossRef]
- Taketa, S.; Amano, S.; Tsujino, Y.; Sato, T.; Saisho, D.; Kakeda, K.; Nomura, M.; Suzuki, T.; Matsumoto, T.; Sato, K.; et al. Barley Grain with Adhering Hulls Is Controlled by an ERF Family Transcription Factor Gene Regulating a Lipid Biosynthesis Pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 4062–4067. [Google Scholar] [CrossRef] [Green Version]
- Lundqvist, U.; Franckowiak, J.D. BGS 354, Glossy Sheath4, Gsh4. Barley Genet. Newsl. 2011, 41, 146–148. [Google Scholar]
- von Wettstein-Knowles, P. Biosynthetic Relationships between β-Diketones and Esterified Alkan-2-Ols Deduced from Epicuticular Wax of Barley Mutants. Mol. Gen. Genet. MGG 1976, 144, 43–48. [Google Scholar] [CrossRef]
- Kumar, A.; Yogendra, K.N.; Karre, S.; Kushalappa, A.C.; Dion, Y.; Choo, T.M. WAX INDUCER1 (HvWIN1) Transcription Factor Regulates Free Fatty Acid Biosynthetic Genes to Reinforce Cuticle to Resist Fusarium Head Blight in Barley Spikelets. J. Exp. Bot. 2016, 67, 4127–4139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druka, A.; Franckowiak, J.; Lundqvist, U.; Bonar, N.; Alexander, J.; Houston, K.; Radovic, S.; Shahinnia, F.; Vendramin, V.; Morgante, M.; et al. Genetic Dissection of Barley Morphology and Development. Plant Physiol. 2011, 155, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Warren, C.R. Rapid Measurement of Chlorophylls with a Microplate Reader. J. Plant Nutr. 2008, 31, 1321–1332. [Google Scholar] [CrossRef]
- Lampinen, J.; Raitio, M.; Perälä, A.; Oranen, H.; Harinen, R.-R. Microplate Based Pathlength Correction Method for Photometric DNA Quantification Assay. Thermo Fisher Scientific. 2012. Available online: https://docslib.org/doc/8198812/microplate-based-pathlength-correction-method-for-photometric (accessed on 7 March 2022).
- Lolle, S.J.; Berlyn, G.P.; Engstrom, E.M.; Krolikowski, K.A.; Reiter, W.-D.; Pruitt, R.E. Developmental Regulation of Cell Interactions in the Arabidopsis Fiddlehead-1 Mutant: A Role for the Epidermal Cell Wall and Cuticle. Dev. Biol. 1997, 189, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Bayer, M.M.; Rapazote-Flores, P.; Ganal, M.; Hedley, P.E.; Macaulay, M.; Plieske, J.; Ramsay, L.; Russell, J.; Shaw, P.D.; Thomas, W.; et al. Development and Evaluation of a Barley 50k ISelect SNP Array. Front. Plant Sci. 2017, 8, 1792. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Wicker, T.; Jenkins, J.; Plott, C.; Lux, T.; Koh, C.S.; Ens, J.; Gundlach, H.; Boston, L.B.; Tulpová, Z.; et al. Long-Read Sequence Assembly: A Technical Evaluation in Barley. Plant Cell 2021, 33, 1888–1906. [Google Scholar] [CrossRef]
- Duan, R.; Xiong, H.; Wang, A.; Chen, G. Molecular Mechanisms Underlying Hull-Caryopsis Adhesion/Separation Revealed by Comparative Transcriptomic Analysis of Covered/Naked Barley (Hordeum vulgare L.). Int. J. Mol. Sci. 2015, 16, 14181–14193. [Google Scholar] [CrossRef] [Green Version]
- Shoesmith, J.; Solomon, C.; Yang, X.; Wilkinson, L.; Sheldrick, S.; van Eijden, E.; Couwenberg, S.; Pugh, L.; Eskan, M.; Stephens, J.; et al. APETALA2 Functions as a Temporal Factor Together with BLADE-ON-PETIOLE2 and MADS29 to Control Flower and Grain Development in Barley. Development 2021, 148, dev194894. [Google Scholar] [CrossRef]
- Milne, L.; Bayer, M.; Rapazote-Flores, P.; Mayer, C.-D.; Waugh, R.; Simpson, C.G. EORNA, a Barley Gene and Transcript Abundance Database. Sci. Data 2021, 8, 90. [Google Scholar] [CrossRef]
- Rapazote-Flores, P.; Bayer, M.; Milne, L.; Mayer, C.-D.; Fuller, J.; Guo, W.; Hedley, P.E.; Morris, J.; Halpin, C.; Kam, J.; et al. BaRTv1.0: An Improved Barley Reference Transcript Dataset to Determine Accurate Changes in the Barley Transcriptome Using RNA-Seq. BMC Genom. 2019, 20, 968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, M.; Shepherd, T.; Mitchell, S.; Topp, C.F.E.; Hoad, S.P. Husk to Caryopsis Adhesion in Barley Is Influenced by Pre- and Post-Anthesis Temperatures through Changes in a Cuticular Cementing Layer on the Caryopsis. BMC Plant Biol. 2017, 17, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl Plants: Integrating Tools for Visualizing, Mining, and Analyzing Plant Genomics Data BT—Plant Bioinformatics: Methods and Protocols; Edwards, D., Ed.; Springer: New York, NY, USA, 2016; pp. 115–140. ISBN 978-1-4939-3167-5. [Google Scholar]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, S.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H.; et al. The Sol Genomics Network (SGN)—from Genotype to Phenotype to Breeding. Nucleic Acids Res. 2015, 43, D1036–D1041. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A Comparative Platform for Green Plant Genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a Mixture Model by Expectation Maximization to Discover Motifs in Biopolymers. In Proceedings of the 2nd International Conference on Intelligent Systems for Molecular Biology, Stanford, CA, USA, 14–17 August 1994; ISCB: Leesburg, VA, USA, 1994; Volume 2, pp. 28–36. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Russell, J.; Mascher, M.; Dawson, I.K.; Kyriakidis, S.; Calixto, C.; Freund, F.; Bayer, M.; Milne, I.; Marshall-Griffiths, T.; Heinen, S.; et al. Exome Sequencing of Geographically Diverse Barley Landraces and Wild Relatives Gives Insights into Environmental Adaptation. Nat. Genet. 2016, 48, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- Bustos-Korts, D.; Dawson, I.K.; Russell, J.; Tondelli, A.; Guerra, D.; Ferrandi, C.; Strozzi, F.; Nicolazzi, E.L.; Molnar-Lang, M.; Ozkan, H.; et al. Exome Sequences and Multi-Environment Field Trials Elucidate the Genetic Basis of Adaptation in Barley. Plant J. 2019, 99, 1172–1191. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.W.; Bryant, D. Popart: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- South, A. Rworldmap: A New R Package for Mapping Global Data. R J. 2011, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Jayakodi, M.; Padmarasu, S.; Haberer, G.; Bonthala, V.S.; Gundlach, H.; Monat, C.; Lux, T.; Kamal, N.; Lang, D.; Himmelbach, A.; et al. The Barley Pan-Genome Reveals the Hidden Legacy of Mutation Breeding. Nature 2020, 588, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, D.C.; Lambert, J.W. Inheritance of the Glossy-Sheath Character in Barley, Hordeum vulgare L. 1. Crop Sci. 1965, 5, 251–253. [Google Scholar] [CrossRef]
- Mikkelsen, J.D. Structure and Biosynthesis of β-Diketones in Barley Spike Epicuticular Wax. Carlsberg Res. Commun. 1979, 44, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Tanaka, H.; Machida, C.; Watanabe, M.; Machida, Y. A New Method for Rapid Visualization of Defects in Leaf Cuticle Reveals Five Intrinsic Patterns of Surface Defects in Arabidopsis. Plant J. 2004, 37, 139–146. [Google Scholar] [CrossRef]
- von Wettstein-Knowles, P. The molecular phenotypes of the eceriferum mutants. In Barley Genetics II; Nilan, R.A., Ed.; Washington State University Press: Pullman, WA, USA, 1971; pp. 146–193. [Google Scholar]
- Shahla, A.; Tsuchiya, T. Genetic Analysis in Six Telotrisomic Lines in Barley (Hordeum vulgare L.). J. Hered. 1990, 81, 127–130. [Google Scholar] [CrossRef]
- Lundqvist, U.; Franckowiak, J.D. BGS 351; Glossy Sheath 1. Barley Genet. Newsl. 1997, 26, 294–295. [Google Scholar]
- Li, F.; Wu, X.; Lam, P.; Bird, D.; Zheng, H.; Samuels, L.; Jetter, R.; Kunst, L. Identification of the Wax Ester Synthase/Acyl-Coenzyme A:Diacylglycerol Acyltransferase WSD1 Required for Stem Wax Ester Biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; et al. Overexpression of Arabidopsis ECERIFERUM1 Promotes Wax Very-Long-Chain Alkane Biosynthesis and Influences Plant Response to Biotic and Abiotic Stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Bessire, M.; Chassot, C.; Jacquat, A.; Humphry, M.; Borel, S.; Petétot, J.M.; Métraux, J.; Nawrath, C. A Permeable Cuticle in Arabidopsis Leads to a Strong Resistance to Botrytis Cinerea. EMBO J. 2007, 26, 2158–2168. [Google Scholar] [CrossRef]
- Schnurr, J.; Shockey, J.; Browse, J. The Acyl-CoA Synthetase Encoded by LACS2 Is Essential for Normal Cuticle Development in Arabidopsis. Plant Cell 2004, 16, 629–642. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente Cantó, C.; Russell, J.; Hackett, C.A.; Booth, A.; Dancey, S.; George, T.S.; Waugh, R. Genetic Dissection of Quantitative and Qualitative Traits Using a Minimum Set of Barley Recombinant Chromosome Substitution Lines. BMC Plant Biol. 2018, 18, 340. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.N.; Marshall, D.R. Variation in Epicuticular Wax Content in Wheat. Euphytica 1988, 38, 3–9. [Google Scholar] [CrossRef]
- Qi, P.; Pendergast, T.H.; Johnson, A.; Bahri, B.A.; Choi, S.; Missaoui, A.; Devos, K.M. Quantitative Trait Locus Mapping Combined with Variant and Transcriptome Analyses Identifies a Cluster of Gene Candidates Underlying the Variation in Leaf Wax between Upland and Lowland Switchgrass Ecotypes. Theor. Appl. Genet. 2021, 134, 1957–1975. [Google Scholar] [CrossRef] [PubMed]
- Würschum, T.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. Refining the Genetic Architecture of Flag Leaf Glaucousness in Wheat. Theor. Appl. Genet. 2020, 133, 981–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulloch, A.P.; Baum, B.R.; Hoffman, L.L. A Survey of Epicuticular Waxes among Genera of Triticeae. 2. Chemistry. Can. J. Bot. 1980, 58, 2602–2615. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Zhi, P.; Wang, X.; Xu, B.; Gong, Z.; Chang, C. Origins and Evolution of Cuticle Biosynthetic Machinery in Land Plants. Plant Physiol. 2020, 184, 1998–2010. [Google Scholar] [CrossRef]
- Jetter, R.; Riederer, M. Localization of the Transpiration Barrier in the Epi-and Intracuticular Waxes of Eight Plant Species: Water Transport Resistances Are Associated with Fatty Acyl Rather than Alicyclic Components. Plant Physiol. 2016, 170, 921–934. [Google Scholar] [CrossRef] [Green Version]
- Seufert, P.; Staiger, S.; Arand, K.; Bueno, A.; Burghardt, M.; Riederer, M. Building a Barrier: The Influence of Different Wax Fractions on the Water Transpiration Barrier of Leaf Cuticles. Front. Plant Sci. 2022, 12, 3170. [Google Scholar] [CrossRef]
- Merah, O.; Deléens, E.; Souyris, I.; Monneveux, P. Effect of Glaucousness on Carbon Isotope Discrimination and Grain Yield in Durum Wheat. J. Agron. Crop Sci. 2000, 185, 259–265. [Google Scholar] [CrossRef]
- Shi, J.X.; Adato, A.; Alkan, N.; He, Y.; Lashbrooke, J.; Matas, A.J.; Meir, S.; Malitsky, S.; Isaacson, T.; Prusky, D.; et al. The Tomato SlSHINE3 Transcription Factor Regulates Fruit Cuticle Formation and Epidermal Patterning. New Phytol. 2013, 197, 468–480. [Google Scholar] [CrossRef]
- Kong, L.; Chang, C. Suppression of Wheat TaCDK8/TaWIN1 Interaction Negatively Affects Germination of Blumeria Graminis f.Sp. Tritici by Interfering with Very-Long-Chain Aldehyde Biosynthesis. Plant Mol. Biol. 2018, 96, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jenks, M.A.; Liu, J.; Liu, A.; Zhang, X.; Xiang, J.; Zou, J.; Peng, Y.; Chen, X. Overexpression of Transcription Factor OsWR2 Regulates Wax and Cutin Biosynthesis in Rice and Enhances Its Tolerance to Water Deficit. Plant Mol. Biol. Rep. 2014, 32, 719–731. [Google Scholar] [CrossRef]
- Richardson, A.; Wojciechowski, T.; Franke, R.; Schreiber, L.; Kerstiens, G.; Jarvis, M.; Fricke, W. Cuticular Permeance in Relation to Wax and Cutin Development along the Growing Barley (Hordeum vulgare) Leaf. Planta 2007, 225, 1471–1481. [Google Scholar] [CrossRef] [PubMed]
- Tulloch, A.P. Composition of Leaf Surface Waxes of Triticum Species: Variation with Age and Tissue. Phytochemistry 1973, 12, 2225–2232. [Google Scholar] [CrossRef]
- Kosma, D.K.; Jenks, M.A. Eco-Physiological and Molecular-Genetic Determinants of Plant Cuticle Function in Drought and Salt Stress Tolerance BT—Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 91–120. ISBN 978-1-4020-5578-2. [Google Scholar]
- Djemal, R.; Khoudi, H. The Barley SHN1-Type Transcription Factor HvSHN1 Imparts Heat, Drought and Salt Tolerances in Transgenic Tobacco. Plant Physiol. Biochem. 2021, 164, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Djemal, R.; Mila, I.; Bouzayen, M.; Pirrello, J.; Khoudi, H. Molecular Cloning and Characterization of Novel WIN1/SHN1 Ethylene Responsive Transcription Factor HvSHN1 in Barley (Hordeum vulgare L.). J. Plant Physiol. 2018, 228, 39–46. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Germplasm ID | Background | Accession |
|---|---|---|
| Bowman | - | PI 483237 |
| BW407 | Bowman, gsh4.l in Gateway | NGB 20640, GSHO 2072 |
| BW126 | Bowman, cer-x.60 in Bonus | NGB 20532, GSHO 2073 |
| Tissue | Compound b | Bowman c µg/cm (% of Total) | BW407 c µg/cm (% of Total) | Significance d |
|---|---|---|---|---|
| Flag leaf sheath | C33_Alk | 0.21 (0.71%) | 0.02 (1.53%) | *** |
| C31 β-diketone | 23.5 (77.91%) | 0.01 (1.14%) | *** | |
| C30_FA | 0.39 (1.30%) | 0.02 (1.91%) | *** | |
| C31 OH-β-diketone | 4.14 (13.71%) | NF | - | |
| TOTAL WAX | 30.17 | 1.08 | *** | |
| Spike | C23_Alk | 1.02 (1.29%) | 0.88 (17.1%) | ns |
| C29_Alk | 1.51 (1.91%) | 0.78 (15.2%) | ** | |
| C31_Alk | 3.79 (4.81%) | 0.63 (12.22%) | *** | |
| C31 β-diketone | 48.35 (61.34%) | 0.58 (11.26%) | *** | |
| C31 OH-β-diketone | 18.67 (23.68%) | NF | - | |
| TOTAL WAX | 78.82 | 5.16 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McAllister, T.; Campoli, C.; Eskan, M.; Liu, L.; McKim, S.M. A Gene Encoding a SHINE1/WAX INDUCER1 Transcription Factor Controls Cuticular Wax in Barley. Agronomy 2022, 12, 1088. https://doi.org/10.3390/agronomy12051088
McAllister T, Campoli C, Eskan M, Liu L, McKim SM. A Gene Encoding a SHINE1/WAX INDUCER1 Transcription Factor Controls Cuticular Wax in Barley. Agronomy. 2022; 12(5):1088. https://doi.org/10.3390/agronomy12051088
Chicago/Turabian StyleMcAllister, Trisha, Chiara Campoli, Mhmoud Eskan, Linsan Liu, and Sarah M. McKim. 2022. "A Gene Encoding a SHINE1/WAX INDUCER1 Transcription Factor Controls Cuticular Wax in Barley" Agronomy 12, no. 5: 1088. https://doi.org/10.3390/agronomy12051088
APA StyleMcAllister, T., Campoli, C., Eskan, M., Liu, L., & McKim, S. M. (2022). A Gene Encoding a SHINE1/WAX INDUCER1 Transcription Factor Controls Cuticular Wax in Barley. Agronomy, 12(5), 1088. https://doi.org/10.3390/agronomy12051088