


Deciphering Early Movements and Domestication of Coffea arabica through a Comprehensive Genetic Diversity Study Covering Ethiopia and Yemen

 ,
,
Abstract
1. Introduction
2. Materials and Methods
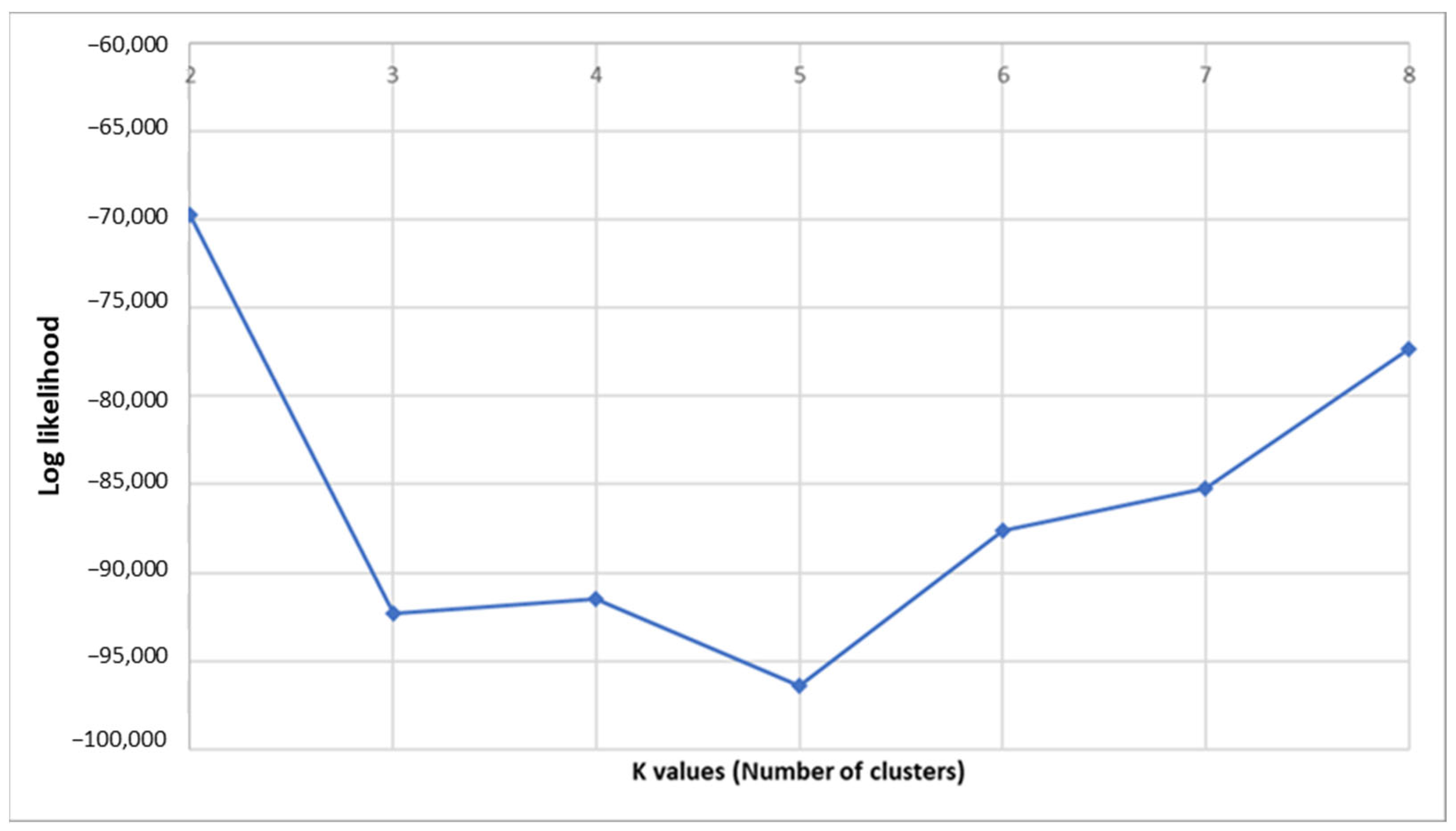
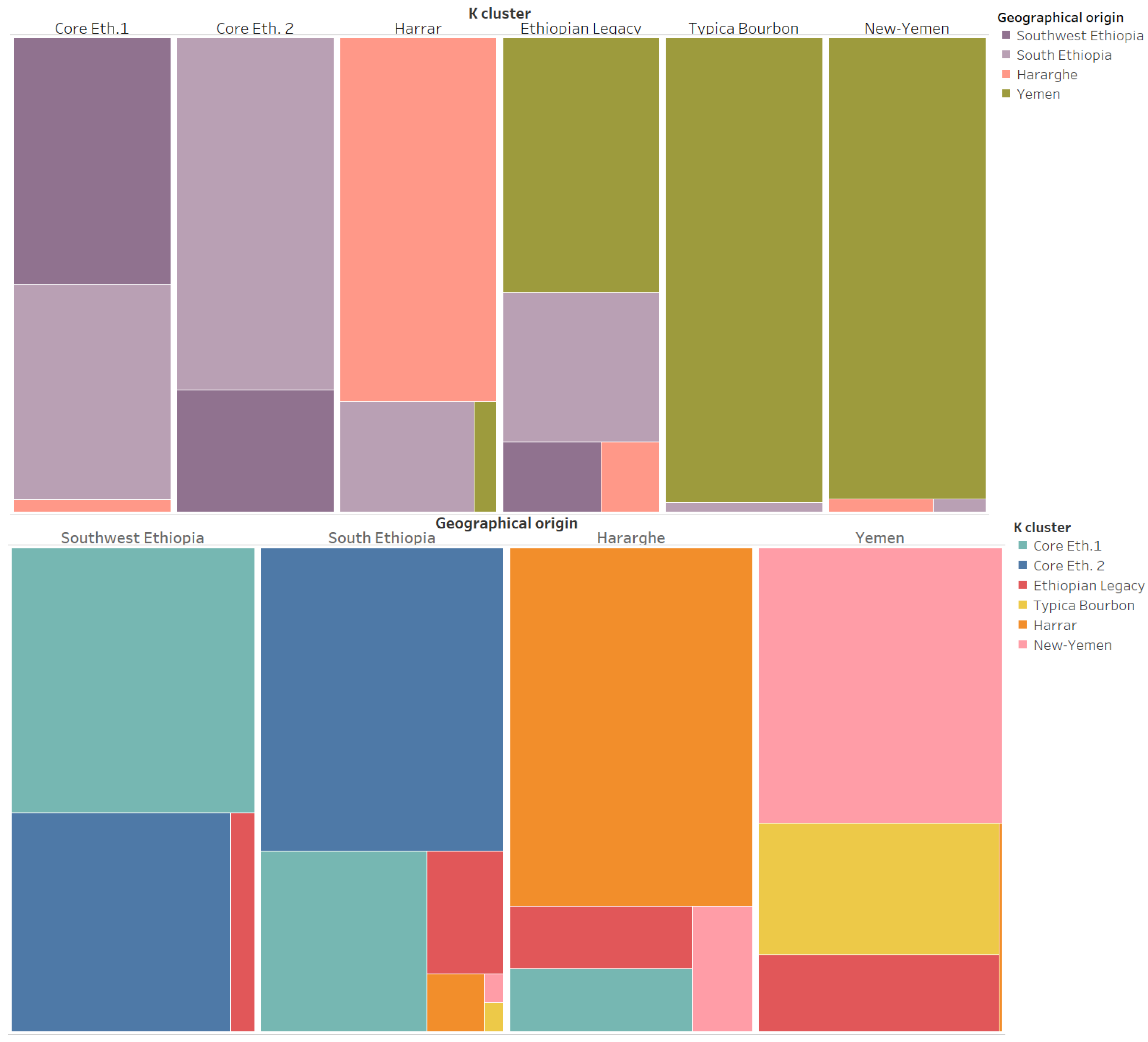
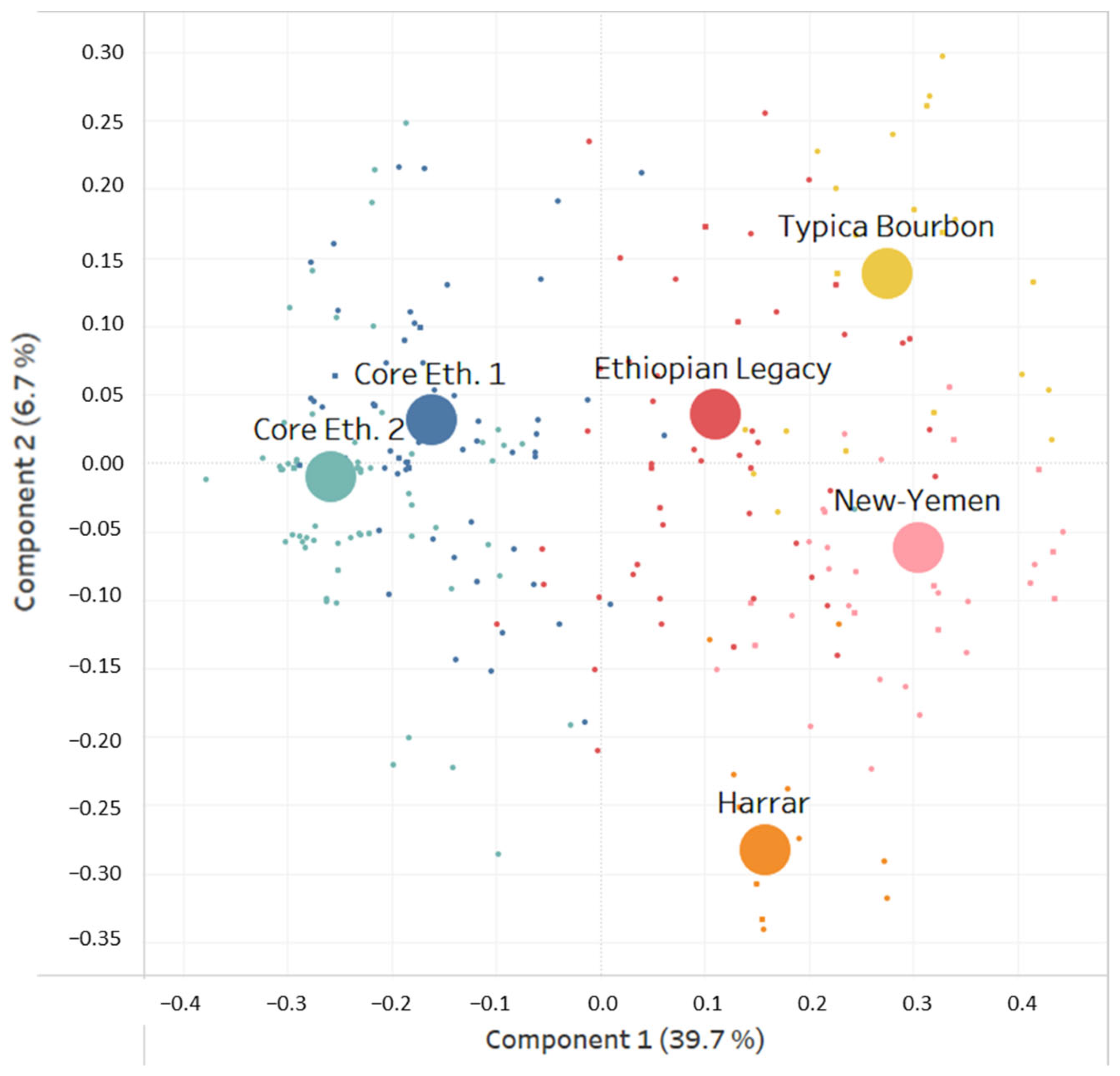
3. Results
4. Discussion
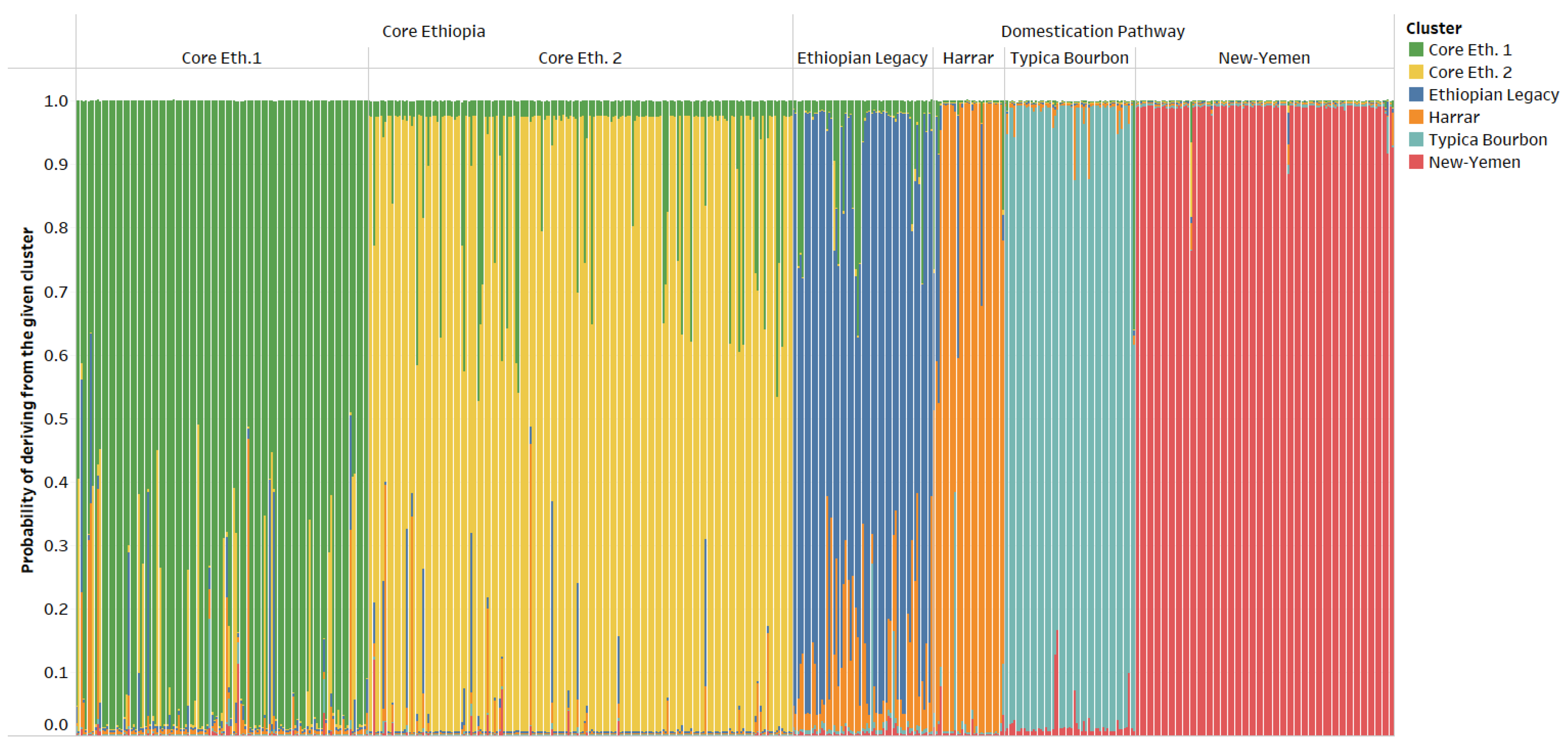
- C. arabica was introduced from Ethiopia to Yemen with seeds from wild populations located in the southern part of Ethiopia, namely in the regions of Gedio (including Yirgachefe) and Guji.
- The descendants of these populations formed the “Ethiopian Legacy” cluster found in these regions of Ethiopia, Hararghe and in some regions of Yemen.
- In Yemen, the “Ethiopian Legacy” cluster is preponderant in Ibb, which was probably a key location for the introduction of coffee into Yemen.
- From this original population, two genetic clusters derived in Yemen, most likely through seed movements and isolation. This gave rise to the “Typica/Bourbon” and “New-Yemen” genetic clusters. The former is spreading from Ibb to Dhamar and the latter prospering up in the northern coffee growing areas of Sanaa and Mawhit.
- The varieties that conquered the world from Yemen are derived from two clusters or mother populations: “Ethiopian Legacy” (SL34, SL09, K-7 or SL17 for instance) and “Typica/Bourbon” (Typica, Bourbon, SL-28 or Kent for instance).
- In Hararghe, a specific genetic cluster—“Harrar”—is identified for the first time. This cluster likely originates from the same “Domestication Pathway” and is derived from the same “Ethiopian Legacy” cluster. In that sense, “Harrar”, “Typica/Bourbon”, and “New-Yemen” share a common genetic origin: The “Ethiopian Legacy” cluster.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lashermes, P.; Combes, M.C.; Robert, J.; Trouslot, P.; D’Hont, A.; Anthony, F.; Charrier, A. Molecular characterisation and origin of the Coffea arabica L. genome. Mol. Gen. Genet. 1999, 261, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Bawin, Y.; Ruttink, T.; Staelens, A.; Haegeman, A.; Stoffelen, P.; Mwanga Mwanga, J.C.I.; Roldan-Ruiz, I.; Honnay, O.; Janssens, S.B. Phylogenomic analysis clarifies the evolutionary origin of Coffea arabica. J. Syst. Evol. 2020, 59, 953–963. [Google Scholar] [CrossRef]
- Scalabrin, S.; Toniutti, L.; Di Gaspero, G.; Scaglione, D.; Magris, G.; Vidotto, M.; Pinosio, S.; Cattonaro, F.; Magni, F.; Jurman, I.; et al. A single polyploidization event at the origin of the tetraploid genome of Coffea arabica is responsible for the extremely low genetic variation in wild and cultivated germplasm. Sci. Rep. 2020, 10, 4642. [Google Scholar] [CrossRef]
- IFC. Performance Standard 6: Biodiversity Conservation and Sustainable Management of Living Natural Resources; International Finance Corporation: Washington, DC, USA, 2012; Available online: https://www.ifc.org/ps6 (accessed on 30 October 2022).
- Meyer, F.G. Notes on wild Coffea arabica from Southwestern Ethiopia, with some historical considerations. Econ. Bot. 1965, 19, 136–151. [Google Scholar] [CrossRef]
- Koehler, J. Where the Wild Coffee Grows: The Untold Story of Coffee from the Cloud Forests of Ethiopia to Your Cup; Bloomsbury Publishing: New York, NY, USA, 2017. [Google Scholar]
- Montagnon, C.; Bouharmont, P. Multivariate analysis of phenotypic diversity of Coffea arabica. Genet. Resour. Crop Evol. 1996, 43, 221–227. [Google Scholar] [CrossRef]
- Lashermes, P.; Trouslot, P.; Anthony, F.; Combes, M.C.; Charrier, A. Genetic diversity for RAPD markers between cultivated and wild accessions of Coffea arabica. Euphytica 1996, 87, 59–64. [Google Scholar] [CrossRef]
- Davis, A.P.; Gole, T.W.; Baena, S.; Moat, J. The impact of climate change on indigenous arabica coffee (Coffea arabica): Predicting future trends and identifying priorities. PLoS ONE 2012, 7, e47981. [Google Scholar] [CrossRef]
- Mairal, M.; Sanmartín, I.; Herrero, A.; Pokorny, L.; Vargas, P.; Aldasoro, J.J.; Alarcón, M. Geographic barriers and Pleistocene climate change shaped patterns of genetic variation in the Eastern Afromontane biodiversity hotspot. Sci. Rep. 2017, 7, 45749. [Google Scholar] [CrossRef]
- Thomas, A.S. The wild Arabica coffee on the Boma Plateau. Angle-Egypt. Sudan. Emp. J. Exp. Agric. 1942, 10, 207–212. [Google Scholar]
- Krishnan, S.; Pruvot-Woehl, S.; Davis, A.P.; Schilling, T.; Moat, J.; Solano, W.; Al Hakimi, A.; Montagnon, C. Validating South Sudan as a Center of Origin for Coffea arabica: Implications for Conservation and Coffee Crop Improvement. Front. Sustain. Food Syst. 2021, 5, 445. [Google Scholar] [CrossRef]
- Anthony, F.; Berthaud, J.; Guillaumet, J.L.; Lourd, M. Collecting wild Coffea species in Kenya and Tanzania. Plant Genet. Resour. Newsl. 1987, 69, 23–29. [Google Scholar]
- De La Roque, J. Voyage de l’Arabie Heureuse, par l’Océan Oriental, et le Détroit de la Mer Rouge: Fait par les François Pour la Première Fois, Dans les Années 1708, 1709 et 1710; André Cailleau: Paris, France, 1716; Available online: https://play.google.com/books/reader?id=3fsOAAAAQAAJ&hl=fr&num=10&printsec=frontcover&pg=GBS.PP7 (accessed on 30 October 2022).
- Ukers, M.A. All about Coffee; The Tea and Coffee Trade Journal: New York, NY, USA, 1922. [Google Scholar]
- Chevalier, A. Les Caféiers du Globe. I. Généralités sur les Caféiers. Encyclopédie Biologique; Paul Lechevalier: Paris, France, 1929. [Google Scholar]
- Cramer, P.J.S. A Review of Literature of Coffee Research in Indonesia (from about 1602 to 1945); IICA: Turrialba, Costa Rica, 1957. [Google Scholar]
- Haarer, A.E. Modern Coffee Production. Ebenezer Baylis and Son; The Trinity Press: London, UK, 1958. [Google Scholar]
- Tuchscherer, M. Commerce et production du café en mer Rouge au XVIe siècle. In Le Commerce du Café Avant L’ère des Plantations Coloniales; Tuchscherer, M., Ed.; Institut Français d’Archéologie Orientale, Cahier des Annales Ismalogiques: Cairo, Egypt, 2001; Volume 20, pp. 69–90. [Google Scholar]
- Berthaud, J. L’origine et la distribution des caféiers dans le monde. In Le Commerce du Café Avant L’ère des Plantations Coloniales; Tuchscherer, M., Ed.; Institut Français d’Archéologie Orientale, Cahier des annales ismalogiques: Cairo, Egypt, 2001; Volume 20, pp. 361–370. [Google Scholar]
- Montagnon, C.; Mahyoub, A.; Solano, W.; Sheibani, F. Unveiling a unique genetic diversity of cultivated Coffea arabica L. in its main domestication center: Yemen. Genet. Resour. Crop Evol. 2021, 68, 2411–2422. [Google Scholar] [CrossRef]
- Anthony, F.; Bertrand, B.; Quiros, O.; Wilches, A.; Lashermes, P.; Berthaud, J.; Charrier, A. Genetic diversity of wild coffee (Coffea arabica L.) using molecular markers. Euphytica 2001, 118, 53–65. [Google Scholar] [CrossRef]
- Silvestrini, M.; Junqueira, M.G.; Favarin, A.C.; Guerreiro-Filho, O.; Maluf, M.P.; Silvarolla, M.B.; Colombo, C.A. Genetic diversity and structure of Ethiopian, Yemen and Brazilian Coffea arabica L. accessions using microsatellites markers. Genet. Resour. Crop Evol. 2007, 54, 1367–1379. [Google Scholar] [CrossRef]
- Aga, E.; Bekele, E.; Bryngelsson, T. Inter-simple sequence repeat (ISSR) variation in forest coffee trees (Coffea arabica L.) populations from Ethiopia. Genetica 2005, 124, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, K.; Oljira, T.; Govers, K.; Belkele, E.; Borsh, T. Genetic diversity and population structure of wild Coffea arabica populations in Ethiopia using molecular markers. In Coffee Diversity and Knowledge; Adugna, G., Bellachew, B., Taye, E., Kufa, T., Eds.; Ethiopian Institute of Agricultural Research: Addis Ababa, Ethiopia, 2008; pp. 35–44. [Google Scholar]
- Dida, G.; Bantte, K.; Disasa, T. Molecular characterization of Arabica Coffee (Coffea arabica L.) germplasms and their contribution to biodiversity in Ethiopia. Plant Biotechnol. Rep. 2021, 15, 791–804. [Google Scholar] [CrossRef]
- Anthony, F.; Combes, M.C.; Astorga, C.; Bertrand, B.; Graziosi, G.; Lashermes, P. The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers. Theor. Appl. Genet. 2002, 104, 894–900. [Google Scholar] [CrossRef]
- FAO. FAO Coffee Mission to Ethiopia: 1964–1965; FAO: Rome, Italy, 1968. [Google Scholar]
- Charrier, A. Etude de la Structure et de la Variabilité Génétique des Caféiers: Résultats des Etudes et des Expérimentations Réalisées au Cameroun, en Côte d’Ivoire et à Madagascar sur l’espèce Coffea arabica L. Collectée en Ethiopie par une Mission Orstom en 1966; Bulletin IFCC n° 14; FRA: Paris, France, 1978. [Google Scholar]
- Bonneuil, C.; Goffaux, R.; Bonnin, I.; Montalent, P.; Hamon, C.; Balfourier, F.; Goldringer, I. A new integrative indicator to assess crop genetic diversity. Ecol. Indic. 2012, 23, 280–289. [Google Scholar] [CrossRef]
- Gnapi, D.E.; Pokou, D.N.D.; Legnate, H.; Dapeng, Z.; Montagnon, C.; Bertrand, B.; N’guetta, A.S.P. Is the genetic integrity of wild Coffea canephora from Ivory Coast threatened by hybridization with introduced coffee trees from Central Africa? Euphytica 2022, 218, 62. [Google Scholar] [CrossRef]
- Pernès, J. Gestion des Ressources Genetiques des Plantes: Tome 2-Manuel; Lavoisier: Paris, France, 1984. [Google Scholar]
- Combes, M.C.; Andrzejewski, S.; Anthony, F.; Bertrand, B.; Rovelli, P.; Graziosi, G.; Lashermes, P. Characterization of microsatellite loci in Coffea arabica and related coffee species. Mol. Ecol. 2002, 9, 1178–1180. [Google Scholar] [CrossRef]
- Pruvot-Woehl, S.; Krishnan, S.; Solano, W.; Schilling, T.; Toniutti, L.; Bertrand, B.; Montagnon, C. Authentication of Coffea arabica Varieties through DNA Fingerprinting and its Significance for the Coffee Sector. J. AOAC Int. 2020, 103, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Wen, X.; Falush, D. 2010 Documentation for Structure Software: Version 2.3. Available online: https://web.stanford.edu/group/pritchardlab/structure_software/release_versions/v2.3.4/structure_doc.pdf (accessed on 9 December 2022).
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. 2006. Available online: http://darwin.cirad.fr/darwin (accessed on 30 October 2022).
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft: Paris, France, 2022; Available online: https://www.xlstat.com/fr (accessed on 30 October 2022).
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 238–244. [Google Scholar] [CrossRef]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- Montagnon, C.; Rossi, V.; Guercio, C.; Sheibani, F. Vernacular Names and Genetics of Cultivated Coffee (Coffea arabica) in Yemen. Agronomy 2022, 22, 1970. [Google Scholar] [CrossRef]
- Murray, D.G. Tableau Your Data!: Fast and Easy Visual Analysis with Tableau Software; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Arwa, A. Yemeni Coffee Trade. Master’s Thesis, University of Sanaa, Sana’a, Yemen, 2004. [Google Scholar]
- Morris, J. Coffee: A Global History; Reaktion Books: London, UK, 2019. [Google Scholar]
- Schaefer, C.G.H. Coffee unobserved: Consumption and Commoditization of Coffee in Ethiopia before the Eighteen century. In Le Commerce du Café Avant L’ère des Plantations Colonials; Tuchscherer, M., Ed.; Institut Français d’Archéologie Orientale, Cahier des annales ismalogiques: Cairo, Egypt, 2001; Volume 20, pp. 23–33. [Google Scholar]
- Sylvain, P.G. Ethiopian coffee—Its significance to world coffee problems. Econ. Bot. 1958, 12, 111–139. [Google Scholar] [CrossRef]
- Ahmed, W.M. History of Harar and the Hararis. Refined Version; Harari People Regional State Culture, Heritage and Tourism Bureau: Harar, Ethiopia, 2015. Available online: https://everythingharar.com/files/History_of_Harar_and_Harari-HNL.pdf (accessed on 30 October 2022).
- Niebuhr, C. Beschreibung von Arabien. Auseigenen Beobachtungen und im Landeselbstgesammleten Nachrichten; Nicolaus Müller: Kopenhagen, Denmark, 1772; Available online: https://digi.ub.uni-heidelberg.de/diglit/niebuhr1772 (accessed on 30 October 2022). [CrossRef]
- Benti, T. Progress in Arabica Coffee breeding in Ethiopia: Achievements, challenges and prospects. Int. J. Sci. Basic Appl. Res. 2017, 33, 15–25. [Google Scholar]
- Hill, T.; Bekele, G. A Reference Guide to Ethiopian Coffee Varieties; Counter Culture Coffee: Durham, NC, USA, 2018. [Google Scholar]
- Benti, T.; Gebre, E.; Tesfaye, K.; Berecha, G.; Lashermes, P.; Kyallo, M.; Kouadio Yao, N. Genetic diversity among commercial arabica coffee (Coffea arabica L.) varieties in Ethiopia using simple sequence repeat markers. J. Crop Improv. 2021, 35, 147–168. [Google Scholar] [CrossRef]
- Labouisse, J.P.; Bellachew, B.; Kotecha, S.; Bertrand, B. Current status of coffee (Coffea arabica L.) genetic resources in Ethiopia: Implications for conservation. Genet. Resour. Crop Evol. 2008, 55, 1079–1093. [Google Scholar] [CrossRef]
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Lughadha, E.N. High extinction risk for wild coffee species and implications for coffee sector sustainability. Sci. Adv. 2019, 5, eaav3473. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Data. 2022. Available online: https://www.fao.org/faostat/en/#home (accessed on 30 October 2022).
- Berecha, G.; Aerts, R.; Vandepitte, K.; Van Glabeke, S.; Muys, B.; Roldán-Ruiz, I.; Honnay, O. Effects of forest management on mating patterns, pollen flow and intergenerational transfer of genetic diversity in wild Arabica coffee (Coffea arabica L.) from Afromontane rainforests. Biol. J. Linn. Soc. 2014, 112, 76–88. [Google Scholar] [CrossRef]
- Bellachew, B.; Atero, B.; Tefera, F.; Ayano, A.; Benti, T. Genetic diversity and heterosis in arabica coffee. In Coffee Diversity and Knowledge; Adugna, G., Bellachew, B., Taye, E., Kufa, T., Eds.; Ethiopian Institute of Agricultural Research: Addis Ababa, Ethiopia, 2008; pp. 50–57. [Google Scholar]
- Alemayehu, D. Review on genetic diversity of coffee (Coffea arabica L.) in Ethiopia. Int. J. For. Hortic. 2017, 3, 18–27. [Google Scholar] [CrossRef]
- Bertrand, B.; Alpizar, E.; Lara, L.; Santacreo, R.; Hidalgo, M.; Quijano, J.M.; Montagnon, C.; Georget, F.; Etienne, H. Performance of Coffea arabica F1 hybrids in agroforestry and full-sun cropping systems in comparison with American pure line cultivars. Euphytica 2011, 181, 147–158. [Google Scholar] [CrossRef]
- Georget, F.; Marie, L.; Alpizar, E.; Courtel, P.; Bordeaux, M.; Hidalgo, J.M.; Marraccini, P.; Breitler, J.C.; Déchamp, E.; Ponçon, C.; et al. Starmaya: The first Arabica F1 coffee hybrid produced using genetic male sterility. Front. Plant Sci. 2019, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Marie, L.; Abdallah, C.; Campa, C.; Courtel, P.; Bordeaux, M.; Navarini, L.; Lonzarich, V.; Bosselmann, A.S.; Turreira-Garcia, N.; Alpizar, E.; et al. G × E interactions on yield and quality in Coffea arabica: New F1 hybrids outperform American cultivars. Euphytica 2020, 216, 78. [Google Scholar] [CrossRef]
- Bertrand, B.; Villegas Hincapie, A.M.; Marie, L.; Breitler, J.C. Breeding for the main agricultural farming of arabica coffee. Front. Sustain. Food Syst. 2021, 5, 709901. [Google Scholar] [CrossRef]
- Gebreselassie, H.; Atinafu, G.; Degefa, M.; Ayano, A. Arabica Coffee (Coffea arabica L.) Hybrid Genotypes Evaluation for Growth Characteristics and Yield Performance under Southern Ethiopian Growing Condition. Acad. Res. J. Agric. Sci. Res. 2018, 6, 89–96. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Jimenez, J.G.P.; Montagnon, C.; Schilling, T. Multiclass classification of agro-ecological zones for Arabica coffee: An improved understanding of the impacts of climate change. PLoS ONE 2015, 10, e0140490. [Google Scholar] [CrossRef]
- Breitler, J.C.; Etienne, H.; Léran, S.; Marie, L.; Bertrand, B. Description of an Arabica coffee ideotype for agroforestry cropping systems: A guideline for breeding more resilient new varieties. Plants 2022, 11, 2133. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
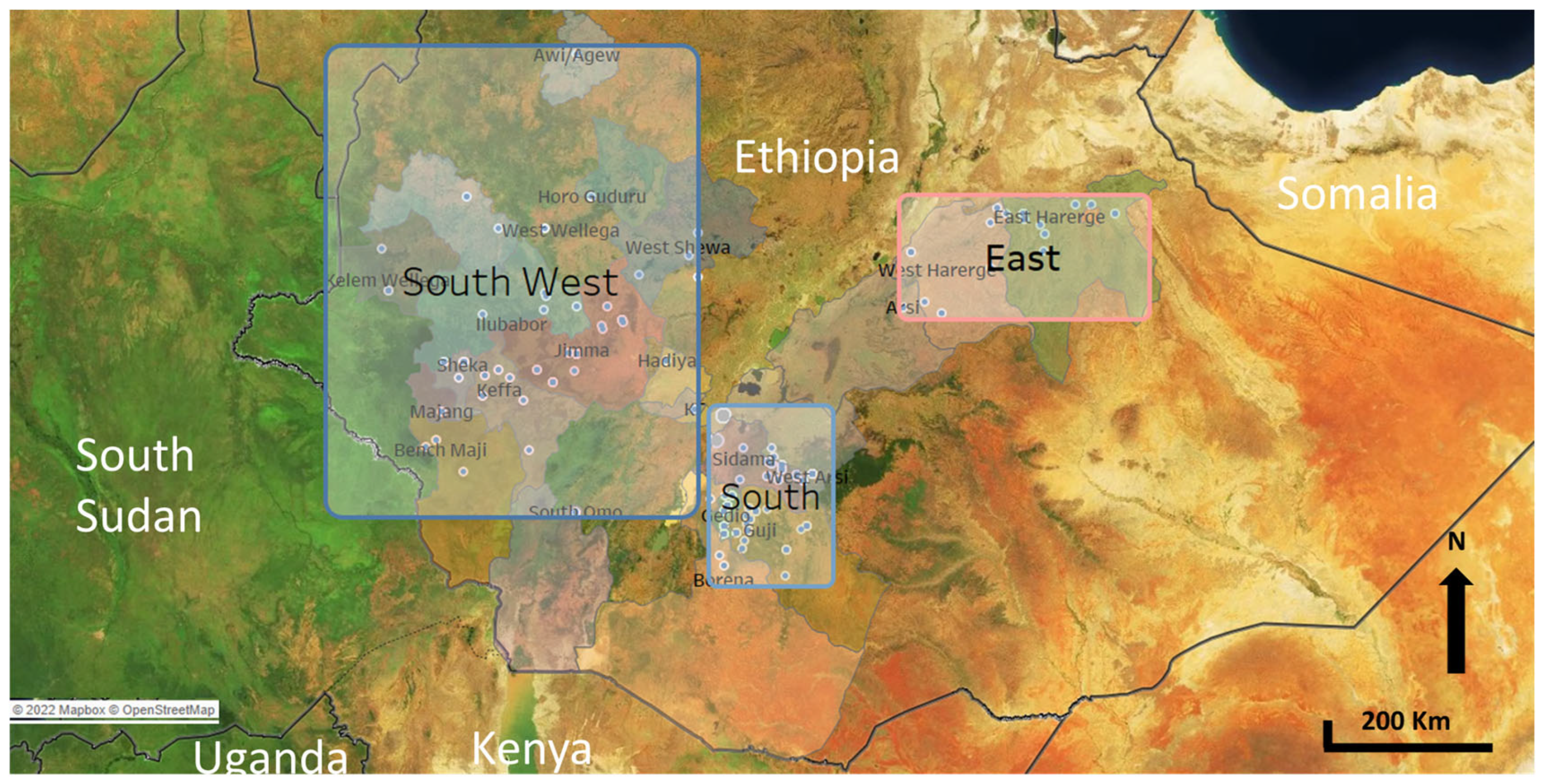
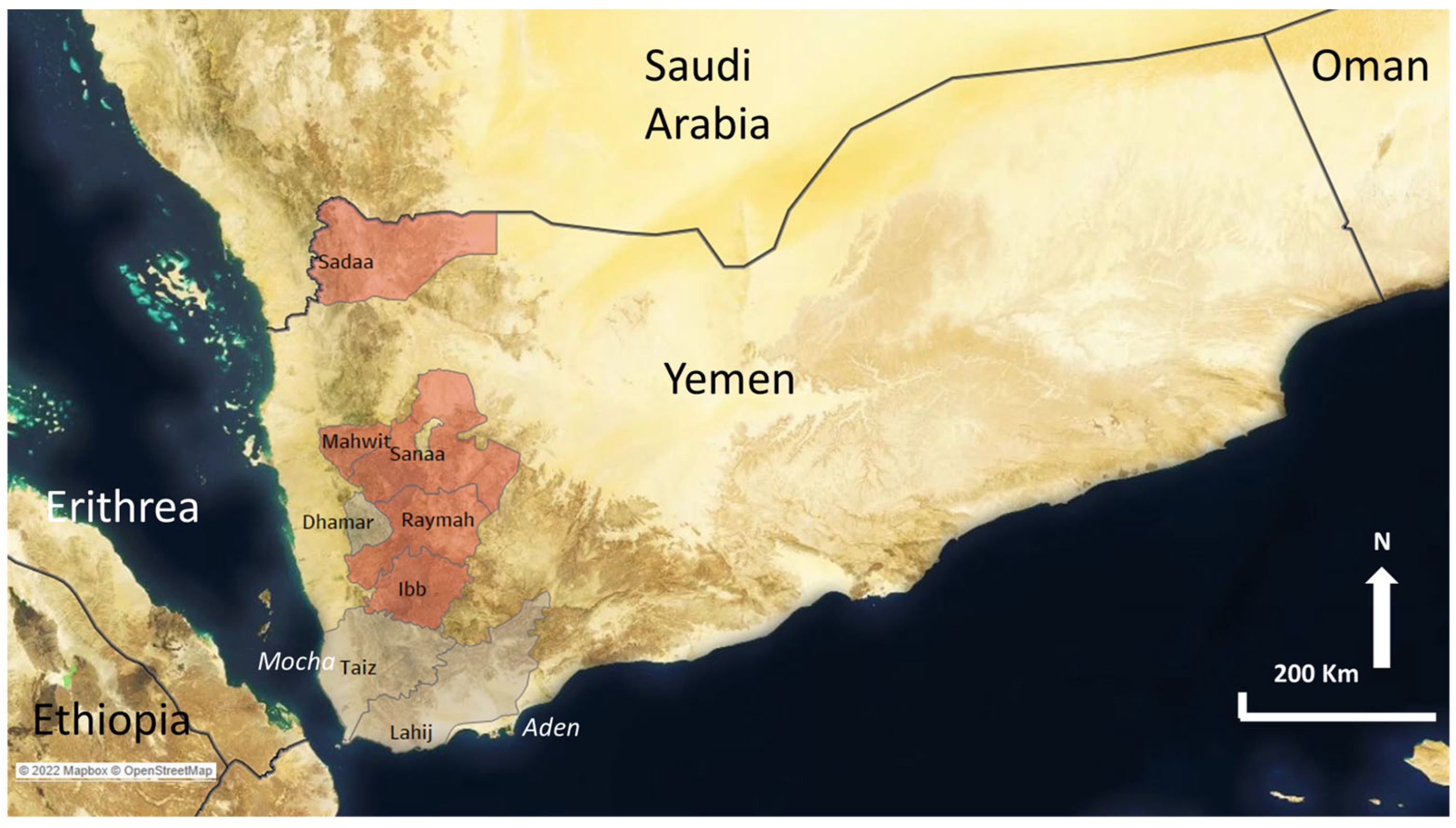
| Ethiopia | Yemen | |||
|---|---|---|---|---|
| High Level Areas | Woreda (District) | # Samples | Governorate | # Samples |
| Southwest | Awi/Agew | 1 | Sadaa | 2 |
| Bench Maji | 4 | Sanaa | 84 | |
| Hadiya | 2 | Mahwit | 25 | |
| Horo guduru | 1 | Dhamar | 31 | |
| Ilubabor | 9 | Ibb | 44 | |
| Jimma | 66 | Total | 186 | |
| Keffa | 9 | |||
| Kelem Wellega | 4 | |||
| KT | 1 | |||
| Majang | 1 | |||
| Sheka | 5 | |||
| South Omo | 1 | |||
| West Shewa | 3 | |||
| West Wellega | 6 | |||
| Southwest | Sub Total | 113 | ||
| South | Borena | 1 | ||
| Gedio | 51 | |||
| Guji | 54 | |||
| Sidama | 41 | |||
| West Arsi | 65 | |||
| South | Sub Total | 212 | ||
| East (Hararghe) | Arsi | 2 | ||
| East Hararghe | 20 | |||
| West Hararghe | 9 | |||
| East (Hararghe) | Sub Total | 31 | ||
| Total | 356 | |||
| Code SSR | Primer Sequence Forward | Primer Sequence Reverse | Size Product (Base Pairs) |
|---|---|---|---|
| Sat-11 | ACCCGAAAGAAAGAACCAA | CCACACAACTCTCCTCATTC | 143–145 |
| Sat-207 | CAATCTCTTTCCGATGCTCT | GAAGCCGTTTCAAGCC | 83–93 |
| Sat-225 | CATGCCATCATCAATTCCAT | TTACTGCTCATCATTCCGCA | 283–317 |
| Sat-235 | TCGTTCTGTCATTAAATCGTCAA | GCAAATCATGAAAATAGTTGGTG | 245–278 |
| Sat-24 | GGCTCGAGATATCTGTTTAG | TTTAATGGGCATAGGGTCC | 167–181 |
| Sat-244 | GCATACTAAGGAATTATCTGACTGCT | GCATGTGCTTTTTGATGTCGT | 178–306 |
| Sat-254 | ATGTTCTTCGCTTCGCTAAC | AAGTGTGGGAGTGTCTGCAT | 221–237 |
| Sat-29 | GACCATTACATTTCACACAC | GCATTTTGTTGCACACTGTA | 137–154 |
| Sat-32 | AACTCTCCATTCCCGCATTC | CTGGGTTTTCTGTGTTCTCG | 119–125 |
| Sat-47 | TGATGGACAGGAGTTGATGG | TGCCAATCTACCTACCCCTT | 135–169 |
| Core Ethiopia | Domestication Pathway | |||||||
|---|---|---|---|---|---|---|---|---|
| Samples Origin | Areas | Core Eth. 1 | Core Eth. 2 | Ethiopian Legacy | Typica Bourbon | New-Yemen | Harrar | Total |
| Ethiopia COE Survey | Southwest | 62 | 46 | 5 | 113 | |||
| South | 54 | 133 | 17 | 1 | 1 | 6 | 212 | |
| East/Hararghe | 3 | 3 | 2 | 23 | 31 | |||
| Yemen Survey | 29 | 50 | 106 | 1 | 186 | |||
| Varieties cultivated worldwide | 5 | 4 | 9 | |||||
| Ethiopian landraces cultivated worldwide | 4 | 4 | ||||||
| Total | 123 | 179 | 59 | 55 | 109 | 30 | 555 | |
| Total allele number | 65 | 45 | 39 | 30 | 30 | 29 | 77 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montagnon, C.; Sheibani, F.; Benti, T.; Daniel, D.; Bote, A.D. Deciphering Early Movements and Domestication of Coffea arabica through a Comprehensive Genetic Diversity Study Covering Ethiopia and Yemen. Agronomy 2022, 12, 3203. https://doi.org/10.3390/agronomy12123203
Montagnon C, Sheibani F, Benti T, Daniel D, Bote AD. Deciphering Early Movements and Domestication of Coffea arabica through a Comprehensive Genetic Diversity Study Covering Ethiopia and Yemen. Agronomy. 2022; 12(12):3203. https://doi.org/10.3390/agronomy12123203
Chicago/Turabian StyleMontagnon, Christophe, Faris Sheibani, Tadesse Benti, Darrin Daniel, and Adugna Debela Bote. 2022. "Deciphering Early Movements and Domestication of Coffea arabica through a Comprehensive Genetic Diversity Study Covering Ethiopia and Yemen" Agronomy 12, no. 12: 3203. https://doi.org/10.3390/agronomy12123203
APA StyleMontagnon, C., Sheibani, F., Benti, T., Daniel, D., & Bote, A. D. (2022). Deciphering Early Movements and Domestication of Coffea arabica through a Comprehensive Genetic Diversity Study Covering Ethiopia and Yemen. Agronomy, 12(12), 3203. https://doi.org/10.3390/agronomy12123203