


Comparative Evaluation of Pyrus Species to Identify Possible Resources of Interest in Pear Breeding


 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Biological Material
2.2. Measuring and Assessing Morphological Traits of Trees, Leaves, Flowers, and Fruits
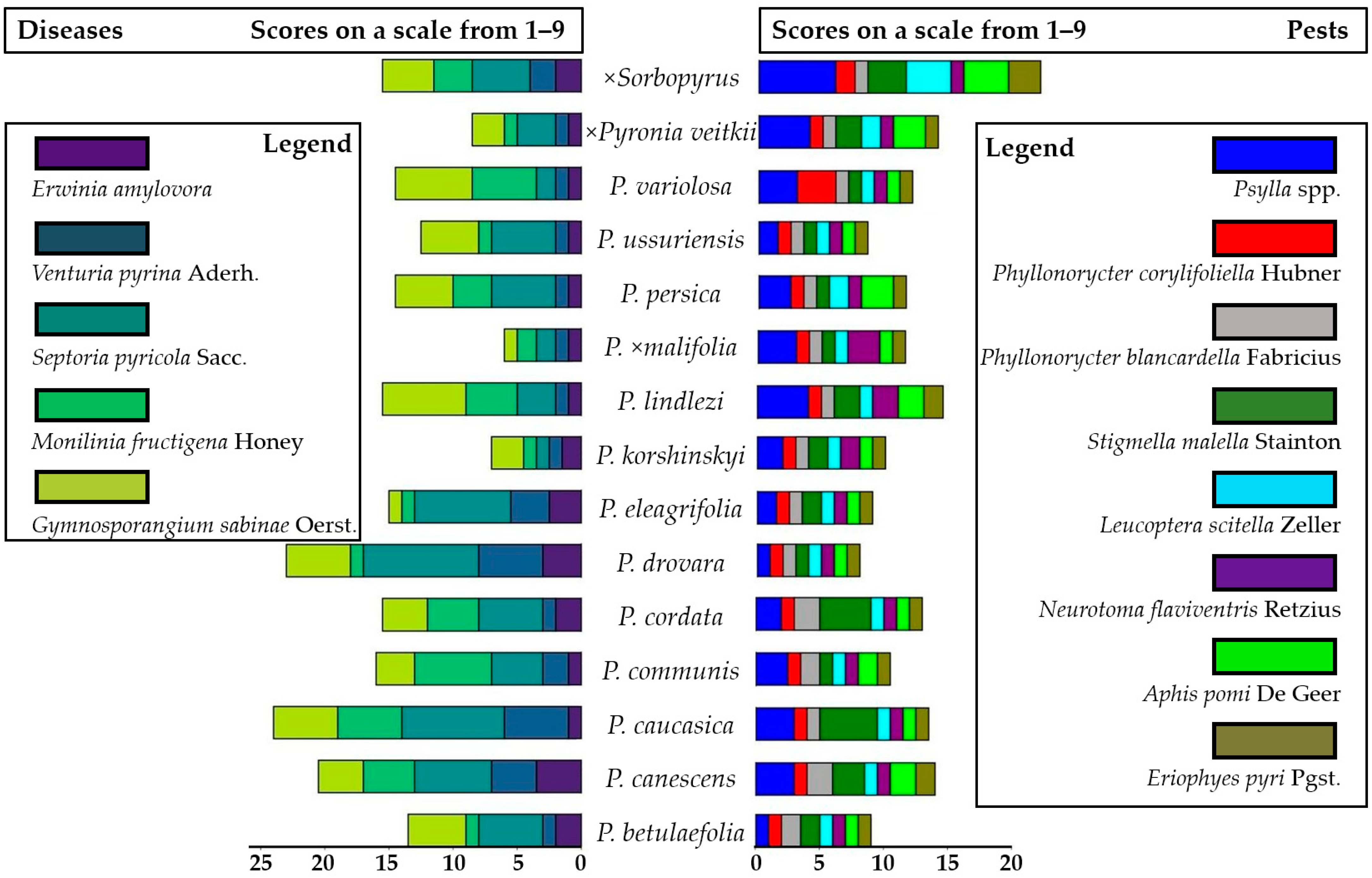
2.3. Assessing the Response to Attacks by Principal Diseases and Pests
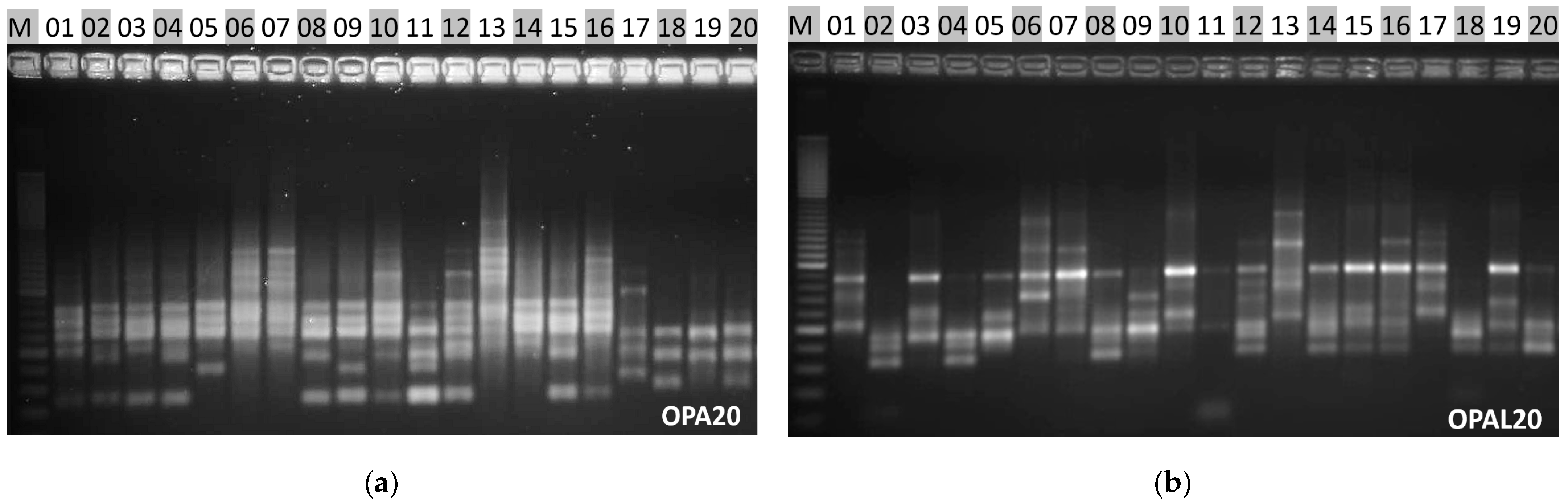
2.4. Genetic Diversity Analysis
2.5. Statistical Analysis
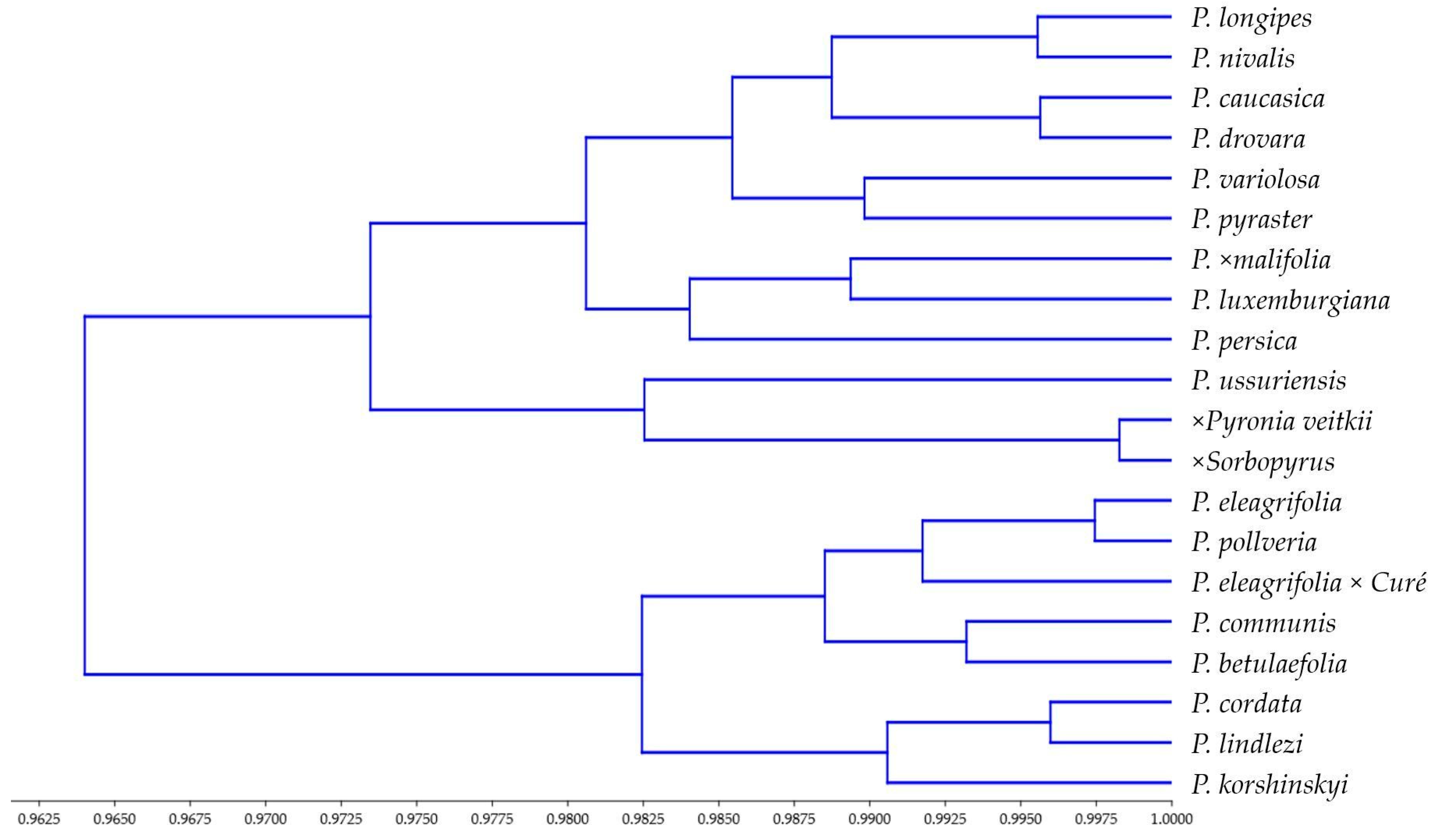
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



References
- Li, J.; Zhang, M.; Li, X.; Khan, A.; Kumar, S.; Allan, A.C.; Lin-Wang, K.; Espley, R.V.; Wang, C.; Wang, R.; et al. Pear genetics: Recent advances, new prospects, and a roadmap for the future. Hort. Res. 2022, 9, uhab040. [Google Scholar] [CrossRef]
- Elzebroek, T.; Wind, K. Edible fruits and nuts. In Guide to Cultivated Plants; CABI: Wallingford, UK, 2008; pp. 25–131. [Google Scholar]
- Khan, A.; Korban, S.S. Breeding and genetics of disease resistance in temperate fruit trees: Challenges and new opportunities. Theor. Appl. Genet. 2022, 135, 3961–3985. [Google Scholar] [CrossRef]
- Mushtaq, M.; Akram, S.; Ishaq, S.; Adnan, A. Pear (Pyrus communis) Seed Oil. In Fruit Oils: Chemistry and Functionality; Ramadan, M.F., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 859–874. [Google Scholar]
- Teixeira, J.D.; Soares Mateus, A.R.; Sanchez, C.; Parpot, P.; Almeida, C.; Sanches Silva, A. Antioxidant capacity and phenolics profile of Portuguese traditional cultivars of apples and pears and their by-products: On the way to newer applications. Foods 2023, 12, 1537. [Google Scholar] [CrossRef]
- Guo, X.; Yang, B.; Tang, J.; Jiang, J.; Li, D. Apple and pear consumption and type 2 diabetes mellitus risk: A meta-analysis of prospective cohort studies. Food Funct. 2017, 8, 927–934. [Google Scholar] [CrossRef]
- Jarić, S.; Mačukanović-Jocić, M.; Djurdjević, L.; Mitrović, M.; Kostić, O.; Karadžić, B.; Pavlović, P. An ethnobotanical survey of traditionally used plants on Suva Planina Mountain (Southeastern Serbia). J. Ethnopharmacol. 2015, 175, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, R.M.; Hammoudi, Z.M. Phytochemistry and pharmacological activity of pear (Pyrus communis Linn): A review. Plant Arch. 2020, 20, 7820–7828. [Google Scholar]
- Postman, J. Pear germplasm needs and conservation. In The Pear Genome; Korban, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 35–50. [Google Scholar]
- Savić, A.; Jarić, S.; Dajić-Stevanović, Z.; Duletić-Laušević, S. Ethnobotanical study and traditional use of autochthonous pear varieties (Pyrus communis L.) in southwest Serbia (Polimlje). Genet. Resour. Crop. Evol. 2019, 66, 589–609. [Google Scholar] [CrossRef]
- Li, X.; Li, X.; Wang, T.; Gao, W. Chapter 24—Nutritional Composition of Pear Cultivars (Pyrus spp.). In Nutritional Composition of Fruit Cultivars; Simmonds, M.S.J., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 573–608. [Google Scholar]
- Oalđe-Pavlović, M.; Alimpić-Aradski, A.; Savić, A.; Janković, S.; Milutinović, M.; Marin, P.D.; Duletić-Laušević, S. Traditional varieties and wild pear from Serbia: A link among antioxidant, antidiabetic and cytotoxic activities of fruit peel and flesh. Bot. Serbica 2021, 45, 203–213. [Google Scholar] [CrossRef]
- Reiland, H.; Slavin, J. Systematic review of pears and health. Food Nutr. Today 2015, 50, 301–305. [Google Scholar] [CrossRef]
- Chandel, R.; Kumar, V.; Kaur, R.; Kumar, S.; Kumar, A.; Kumar, D.; Kapoor, S. Bioactive compounds, health benefits and valorization of (sand pear): A review. Nutr. Food Sci. 2023; ahead-of-print. [Google Scholar] [CrossRef]
- Hussain, S.Z.; Naseer, B.; Qadri, T.; Fatima, T.; Bhat, T.A. Pear (Pyrus communis)—Morphology, Taxonomy, Composition and Health Benefits. In Fruits Grown in Highland Regions of the Himalayas: Nutritional and Health Benefits; Hussain, S.Z., Naseer, B., Qadri, T., Fatima, T., Bhat, T.A., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 35–48. [Google Scholar]
- Parle, M.; Arzoo. Why pear is so dear. Int. J. Res. Ayurveda Pharm. 2016, 7, 108–113. [Google Scholar] [CrossRef]
- Marthy, M. Optimizarea Prelucrabilităţii Prin Frezare şi Prin Şlefuire a Lemnului de păr în Vederea Înglobării în Produse de Mobilă (The Optimization of Processing by Milling and Sanding of Pear Wood, in order to Incorporate it into Furniture Products). Ph.D. Thesis, Transilvania University of Brașov, Braşov, Romania, 2010. [Google Scholar]
- Kimic, K. Pear trees (Pyrus communis L.) as monuments of nature in Warsaw public spaces—State of preservation. In Proceedings of the Public Recreation and Landscape Protection—With Sense Hand in Hand! Brno, Czech Republic, 10–11 May 2021; pp. 349–353. [Google Scholar]
- Bell, R.L.; Itai, A. Pyrus. In Wild Crop Relatives: Genomic and Breeding Resources: Temperate Fruits; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 147–177. [Google Scholar]
- Terpó, A. Pyrus taxa in Hungary, and their practical importance. Thaiszia 1992, 2, 41–57. [Google Scholar]
- Yigit, M.A.; Pinar, H.; Uzun, A.; Demir, O.D.; Uysal, E.; Dilfiruz, T. Using some pome fruit trees in landscape designs. Curr. Trends Nat. Sci. 2018, 7, 297–302. [Google Scholar]
- Irvine, K.N.; Devine-Wright, P.; Payne, S.R.; Fuller, R.A.; Painter, B.; Gaston, K.J. Green space, soundscape and urban sustainability: An interdisciplinary, empirical study. Local Environ. 2009, 14, 155–172. [Google Scholar] [CrossRef]
- Hatipoğlu, İ.H.; Ak, B.E. Chapter 1. The importance of landscape architecture and ornamental plants in sustainable cities. In Ornamental Plants: With Their Features and Usage Principles; Çiğ, A., Ed.; Iksad Publications: Ankara, Turkey, 2020; pp. 3–27. [Google Scholar]
- Şahin, M. Chapter 19. Ornafruit: Fruit species for ornamental purposes. In Ornamental Plants: With Their Features and Usage Principles; Çiğ, A., Ed.; Iksad Publications: Ankara, Turkey, 2020; pp. 397–416. [Google Scholar]
- Challice, J.S.; Westwood, M.N. Numerical taxonomic studies of the genus Pyrus using both chemical and botanical characters. Bot. J. Linn. Soc. 2008, 67, 121–148. [Google Scholar] [CrossRef]
- Hedrick, U.; Howe, G.; Taylor, O.; Francis, E.; Tukey, H. The Pears of New York. 1921. Available online: https://www.gutenberg.org/files/46994/46994-h/46994-h.htm (accessed on 12 March 2023).
- Quinet, M.; Wesel, J. Botany and taxonomy of pear. In The Pear Genome. Compendium of Plant Genomes; Korban, S., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Zamani, A.; Attar, F.; Maroofi, H. A synopsis of the genus Pyrus (Rosaceae) in Iran. Nord. J. Bot. 2012, 30, 310–332. [Google Scholar] [CrossRef]
- Sestras, R. Ameliorarea Speciilor Horticole; AcademicPres: Cluj-Napoca, Romania, 2004. [Google Scholar]
- Bell, R.L.; Quamme, H.A.; Layne, R.E.C.; Skirvin, R.M. Pears. In Fruit Breeding; Janick, J., Moore, J.N., Eds.; John Wiley and Sons: New York, NY, USA, 1996; pp. 441–514. [Google Scholar]
- Hong, S.-Y.; Lansky, E.; Kang, S.-S.; Yang, M. A review of pears (Pyrus spp.), ancient functional food for modern times. BMC Complement. Med. Ther. 2021, 21, 219. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y. The pear industry and research in China. Acta Hortic. 2011, 909, 161–170. [Google Scholar] [CrossRef]
- Simionca Mărcășan, L.I.; Oltean, I.; Popa, S.; Plazas, M.; Vilanova, S.; Gramazio, P.; Sestras, A.F.; Prohens, J.; Sestras, R.E. Comparative analysis of phenotypic and molecular data on response to main pear diseases and pest attack in a germplasm collection. Int. J. Mol. Sci. 2023, 24, 6239. [Google Scholar] [CrossRef]
- Sardella, D.; Muscat, A.; Brincat, J.-P.; Gatt, R.; Decelis, S.; Valdramidis, V. A comprehensive review of the pear fungal diseases. Int. J. Fruit Sci. 2016, 16, 351–377. [Google Scholar] [CrossRef]
- Sutton, T.B.; Aldwinckle, H.S.; Agnello, A.M.; Walgenbach, J.F. Compendium of Apple and Pear Diseases and Pests; American Phytopath Society: St. Paul, MN, USA, 2014. [Google Scholar]
- Husain, M.; Rathore, J.P.; Sharma, A.; Raja, A.; Qadri, I.; Wani, A. Description and management strategies of important pests of pear: A review. J. Entomol. Zool. Stud. 2018, 6, 677–683. [Google Scholar]
- Vanneste, J.L. Fire Blight: The Disease and Its Causative Agent, Erwinia Amylovora; Cabi Publishing: Wallingford, UK, 2000. [Google Scholar]
- Bokszczanin, K.Ł. Future breeding strategies. In The Pear Genome; Korban, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 301–315. [Google Scholar]
- Brewer, L.R.; Palmer, J.W. Global pear breeding programmes: Goals, trends and progress for new cultivars and new rootstocks. Acta Hortic. 2011, 909, 105–119. [Google Scholar] [CrossRef]
- Sestras, R.E.; Sestras, A.F. Quantitative traits of interest in apple breeding and their implications for selection. Plants 2023, 12, 903. [Google Scholar] [CrossRef] [PubMed]
- Smil, V. Feeding the World: A Challenge for the Twenty-First Century; MIT Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Islam, M.; Ahmad, H.; Khalid, A.N.; Inamullah; Mohammad, K.; Masood, R.; Akhtar, N.; Afradi, S.G.; Ahmad, I. Pear (Pyrus): Genetic diversity and their conservation. Fresenius Environ. Bull. 2021, 30, 2333–2342. [Google Scholar]
- Matei, I.; Pacurar, I.; Rosca, S.; Bilasco, S.; Sestras, P.; Rusu, T.; Jude, E.T.; Tăut, F.D. Land use favourability assessment based on soil characteristics and anthropic pollution. Case study Somesul Mic Valley Corridor, Romania. Agronomy 2020, 10, 1245. [Google Scholar] [CrossRef]
- Hancock, J.F.; Lobos, G.A. Pears. In Temperate Fruit Crop Breeding: Germplasm to Genomics; Hancock, J.F., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 299–336. [Google Scholar]
- Dondini, L.; Sansavini, S. European Pear. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer: Boston, MA, USA, 2012; pp. 369–413. [Google Scholar]
- Rehder, A. Manual of Cultivated Trees and Shrubs; Macmillan Company: New York, NY, USA, 1940; Volume 2. [Google Scholar]
- Rubtsov, G.A. Geographical distribution of the genus Pyrus and trends and factors in its evolution. Am. Nat. 1944, 78, 358–366. [Google Scholar] [CrossRef]
- Schoonhoven, A.V.; Pastor Corrales, M.A. Standard System for the Evaluation of Bean Germplasm; Centro Internacional de Agricultura Tropical (CIAT): Cali, CO, USA, 1987; p. 57. [Google Scholar]
- Cooke, B.M. Disease assessment and yield loss. In The Epidemiology of Plant Diseases; Jones, D.G., Ed.; Springer: Dordrecht, The Netherlands, 1998; pp. 42–72. [Google Scholar]
- Simionca Mărcășan, L.I.; Hulujan, I.B.; Florian, T.; Somsai, P.A.; Militaru, M.; Sestras, A.F.; Oltean, I.; Sestras, R.E. The importance of assessing the population structure and biology of psylla species for pest monitoring and management in pear orchards. Not. Bot. Horti Agrobot. 2022, 50, 13022. [Google Scholar] [CrossRef]
- Yuanwen Teng, Y.; Tanabe, K.; Tamura, F.; Itai, A. Genetic relationships of pear cultivars in Xinjiang, China, as measured by RAPD markers. J. Hortic. Sci. Biotechnol. 2001, 76, 771–779. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Mota, M.; Monte-Corvo, L.; Goulão, L.; Silva, D.M. Molecular typing of Pyrus based on RAPD markers. Sci. Hortic. 1999, 79, 163–174. [Google Scholar] [CrossRef]
- Monte-Corvo, L.; Cabrita, L.; Oliveira, C.; Leitão, J. Assessment of genetic relationships among Pyrus species and cultivars using AFLP and RAPD markers. Genet. Resour. Crop Evol. 2000, 47, 257–265. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- UPOV. Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability. Pear (Pyrus Communis). TG/15/3; International Union for the Protection of New Varieties of Plants: Geneva, Switzerland, 2000; pp. 1–40. [Google Scholar]
- Lateur, M.; Szalatnay, D.; Höfer, M.; Bergamaschi, M.; Guyader, A.; Hjalmarsson, I.; Militaru, M.; Miranda Jiménez, C.; Osterc, G.; Rondia, A.; et al. ECPGR Characterization and Evaluation Descriptors for Pear Genetic Resources; European Cooperative Programme for Plant Genetic Resources: Rome, Italy, 2022; pp. 1–48. [Google Scholar]
- Khadivi, A.; Mirheidari, F.; Moradi, Y.; Paryan, S. Morphological and pomological characterizations of Pyrus syriaca Boiss. germplasm. Sci. Hortic. 2020, 271, 109424. [Google Scholar] [CrossRef]
- Zarei, A.; Erfani-Moghadam, J.; Jalilian, H. Assessment of variability within and among four Pyrus species using multivariate analysis. Flora 2019, 250, 27–36. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, Y.-F.; Huo, H.-L.; Xu, J.-Y.; Tian, L.-M.; Dong, X.-G.; Qi, D.; Liu, C. An assessment of the genetic diversity of pear (Pyrus L.) germplasm resources based on the fruit phenotypic traits. J. Integr. Agric. 2022, 21, 2275–2290. [Google Scholar] [CrossRef]
- Dupraz, C. Prospects for easing land tenure conflicts with agroforestry in Mediterranean France: A research approach for intercropped timber orchards. Agrofor. Syst. 1994, 25, 181–192. [Google Scholar] [CrossRef]
- Rigueiro-Rodríguez, A.; McAdam, J.; Mosquera-Losada, M.R. Agroforestry in Europe: Current Status and Future Prospects; Springer Science + Business Media B.V: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Báder, M.; Németh, R.; Vörös, Á.; Tóth, Z.; Novotni, A. The effect of agroforestry farming on wood quality and timber industry and its supportation by Horizon 2020. Agrofor. Syst. 2023, 97, 587–603. [Google Scholar] [CrossRef]
- Matus, F.; Retamales, J.; Sánchez, P. Effect of particle size and quality of pruning wood residues of Asian Pear (Pyrus pyrifolia and Pyrus communis) on C-and N mineralisation in soils of contrasting textures. Rev. Cienc. Suelo Nutr. Veg. 2006, 6, 1–8. [Google Scholar] [CrossRef]
- Vidaković, A.; Liber, Z.; Šatović, Z.; Idžojtić, M.; Volenec, I.; Zegnal, I.; Pintar, V.; Radunić, M.; Poljak, I. Phenotypic diversity of almond-leaved pear (Pyrus spinosa Forssk.) along Eastern Adriatic Coast. Forests 2021, 12, 1630. [Google Scholar] [CrossRef]
- Filipović, D.; Tasić, N. Vinča-Belo Brdo, a late neolithic site in Serbia consideration of the macro-botanical remains as indicators of dietary habits. Balcanica 2012, 43, 7–27. [Google Scholar] [CrossRef]
- Miladinović-Radmilović, N.; Vitezović, S. Archaeobotany at neolithic sites in Serbia: A critical overview of the methods and results. In Bioarchaeology in the Balkans; Miladinović-Radmilović, N., Vitezović, S., Eds.; Srpsko Arheološko Društvo and Blago Sirmijuma: Beograd, Serbia; Sremska, Serbia; Mitrovica, Serbia, 2013; pp. 25–55. [Google Scholar]
- Filipović, D.; Obradović, Đ.; Tripković, B. Plant storage in Neolithic southeast Europe: Synthesis of the archaeological and archaeobotanical evidence from Serbia. Veg. Hist. Archaeobot. 2018, 27, 31–44. [Google Scholar] [CrossRef]
- Vidaković, A.; Šatović, Z.; Tumpa, K.; Idžojtić, M.; Liber, Z.; Pintar, V.; Radunić, M.; Runjić, T.N.; Runjić, M.; Rošin, J.; et al. Phenotypic variation in European wild pear (Pyrus pyraster (L.) Burgsd.) populations in the North-Western Part of the Balkan Peninsula. Plants 2022, 11, 335. [Google Scholar] [CrossRef] [PubMed]
- Bieniasz, M.; Necas, T.; Dziedzic, E.; Ondrasek, I.; Pawłowska, B. Evaluation of pollen quality and self-fertility in selected cultivars of Asian and European pears. Not. Bot. Horti Agrobot. 2017, 45, 375–382. [Google Scholar] [CrossRef]
- Le Lezec, M.; Belouin, A.; Simard, M.H. A selection from Pyrus betulaefolia as a new pollinator for the main Pyrus communis cultivars. Acta Hortic. 2005, 671, 253–255. [Google Scholar] [CrossRef]
- Kemp, H.; Koskela, E.; van Dieren, M.C.A.; Maas, F.M. Selected Pyrus genotypes as pollinizers for Pyrus communis cultivars. Acta Hortic. 2008, 800, 189–198. [Google Scholar] [CrossRef]
- Yamada, K.; Uematsu, C.; Katayama, H. Pear (Pyrus L.) genetic resources from northern Japan: Organoleptic evaluation of ornamental pear trees. Acta Hortic. 2015, 1094, 117–122. [Google Scholar] [CrossRef]
- Pittenger, D.R. Evaluation of Interspecific Hybrid Pears for Use in Southern California Landscapes; Center for Landscape and Urban Horticulture, University of California Cooperative Extension: Los Angeles, CA, USA, 2011; pp. 1–15. [Google Scholar]
- Culley, T.M.; Hardiman, N.A. The role of intraspecific hybridization in the evolution of invasiveness: A case study of the ornamental pear tree Pyrus calleryana. Biol. Invasions 2009, 11, 1107–1119. [Google Scholar] [CrossRef]
- Hartshorn, J.A.; Palmer, J.F.; Coyle, D.R. Into the wild: Evidence for the enemy release hypothesis in the invasive Callery pear (Pyrus calleryana) (Rosales: Rosaceae). Environ. Entomol. 2021, 51, 216–221. [Google Scholar] [CrossRef]
- Maruşca, T. Însemnări şi Mărturii Agrosilvopastorale; Editura Universităţii Transilvania: Braşov, Romania, 2015. [Google Scholar]
- Coman, M.; Militaru, M.; Butac, M. Fruit varieties breeding in Romania: From the beginning to present. Ann. Acad. Rom. Sci. Ser. Agr. For. Vet. Med. Sci. 2012, 1, 43–52. [Google Scholar]
- Militaru, M.; Braniste, N.; Butac, M.; Sestras, A.; Sotiropoulos, T.; Lukić, M.; Ambrozič Turk, B.; Dzhuvinov, V. Review of pome fruit breeding in Balkan. Acta Hortic. 2013, 981, 177–184. [Google Scholar] [CrossRef]
- Militaru, M.; Braniște, N.; Sestras, A.; Andreieș, N.; Butac, M.; Stanciu, C. Contributions to the improvement of the Romanian pear varieties in the past 10 years. Lucr. Șt. Univ. Șt. Agr. Med. Vet. Iași Ser. Hort. 2010, 53, 341–346. [Google Scholar]
- Sestras, A.F.; Sestras, R.E.; Barbos, A.; Militaru, M. The differences among pear genotypes to fire blight (Erwinia amylovora) attack, based on observations of natural infection. Not. Bot. Horti Agrobot. 2008, 36, 97–103. [Google Scholar] [CrossRef]
- Sestras, A.; Somsai, P.; Militaru, M.; Mitre, V.; Ercişli, S.; Sestras, R. The response of pear cultivars and wild species of Pyrus to Psylla sp. attack, depending on genotype, based on eggs’ and nymphs’ presence on the leaves, before and after the treatment with insecticide. Acta Hort. 2020, 1289, 79–90. [Google Scholar] [CrossRef]
- Sestras, R.E.; Pamfil, D.; Ardelean, M.; Botez, C.; Sestras, A.F.; Mitre, I.; Dan, C.; Mihalte, L. Use of phenotypic and MAS selection based on bulk segregant analysis to reveal the genetic variability induced by artificial hybridization in apple. Not. Bot. Horti Agrobot. 2009, 37, 273–277. [Google Scholar] [CrossRef]
- Teng, Y.; Tanabe, K.; Tamura, F.; Itai, A. Genetic relationships of Pyrus species and cultivars native to East Asia revealed by Randomly Amplified Polymorphic DNA markers. J. Am. Soc. Hortic. Sci. 2002, 127, 262–270. [Google Scholar] [CrossRef]
- Silva, G.J.; Souza, T.M.; Barbieri, R.L.; Costa de Oliveira, A. Origin, domestication, and dispersing of pear (Pyrus spp.). Adv. Agric. 2014, 2014, 541097. [Google Scholar] [CrossRef]
- Puskás, M.; Höfer, M.; Sestraş, R.E.; Peil, A.; Sestraş, A.F.; Hanke, M.-V.; Flachowsky, H. Molecular and flow cytometric evaluation of pear (Pyrus L.) genetic resources of the German and Romanian national fruit collections. Genet. Resour. Crop Evol. 2016, 63, 1023–1033. [Google Scholar] [CrossRef]
- Volk, G.M.; Cornille, A. Genetic diversity and domestication history in Pyrus. In The Pear Genome. Compendium of Plant Genomes; Korban, S., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Liu, Q.; Song, Y.; Liu, L.; Zhang, M.; Sun, J.; Zhang, S.; Wu, J. Genetic diversity and population structure of pear (Pyrus spp.) collections revealed by a set of core genome-wide SSR markers. Tree Genet. Genomes 2015, 11, 128. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Arzani, K.; Yadollahi, A.; Karimzadeh, G.; Abdollahi, H. Genetic diversity and similarity of Asian and European pears (Pyrus spp.) revealed by genome size and morphological traits prediction. Int. J. Fruit Sci. 2021, 21, 619–633. [Google Scholar] [CrossRef]
- Postman, J.D. Intergeneric hybrids in Pyrinae (=Maloideae) subtribe of Pyreae in Family Rosaceae at USDA Genebank. Acta Hortic. 2011, 918, 937–943. [Google Scholar] [CrossRef]
- Robertson, K.R.; Phipps, J.B.; Rohrer, J.R.; Smith, P.G. A synopsis of genera in Maloideae (Rosaceae). Syst. Bot. 1991, 16, 376–394. [Google Scholar] [CrossRef]
- Browicz, K. Concept and chorology of the genus Pyrus L. Arbor. Korn. 1993, 38, 17–33. [Google Scholar]
- Uğurlu Aydın, Z.; Dönmez, A.A. Taxonomic and biogeographic notes on the genus Pyrus L. (Rosaceae): A new record and a new synonym, with data on seed morphology. Plant Fungal Res. 2019, 2, 2–8. [Google Scholar] [CrossRef]
- Zheng, X.; Cai, D.; Potter, D.; Postman, J.; Liu, J.; Teng, Y. Phylogeny and evolutionary histories of Pyrus L. revealed by phylogenetic trees and networks based on data from multiple DNA sequences. Mol. Phylogenet. Evol. 2014, 80, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Zheng, X.; Zong, Y.; Jiang, S.; Hu, C.; Yu, P.; Liu, G.; Cao, Y.; Hu, H.; Teng, Y. Combined analyses of chloroplast DNA haplotypes and microsatellite markers reveal new insights into the origin and dissemination route of cultivated pears native to East Asia. Front. Plant Sci. 2018, 9, 591. [Google Scholar] [CrossRef]
- Kumar, S.; Kirk, C.; Deng, C.; Wiedow, C.; Knaebel, M.; Brewer, L. Genotyping-by-sequencing of pear (Pyrus spp.) accessions unravels novel patterns of genetic diversity and selection footprints. Hort. Res. 2017, 4, 17015. [Google Scholar] [CrossRef]
- Sestras, A.F.; Pamfil, D.; Dan, C.; Bolboaca, S.D.; Jäntschi, L.; Sestras, R.E. Possibilities to improve apple scab (Venturia inaequalis (Cke.) Wint.) and powdery mildew [Podosphaera leucotricha (Ell. et Everh.) Salm.] resistance on apple by increasing genetic diversity using potentials of wild species. Aust. J. Crop Sci. 2011, 5, 748–755. [Google Scholar]
- Dan, C.; Sestras, A.; Bozdog, C.; Sestras, R. Investigation of wild species potential to increase genetic diversity useful for apple breeding. Genetika 2015, 47, 993–1011. [Google Scholar] [CrossRef]
- Nishio, S.; Takada, N.; Saito, T.; Yamamoto, T.; Iketani, H. Estimation of loss of genetic diversity in modern Japanese cultivars by comparison of diverse genetic resources in Asian pear (Pyrus spp.). BMC Genet. 2016, 17, 81. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Barrera, M.E.; Ramos-Cabrer, A.M.; Pereira-Lorenzo, S.; Ríos-Mesa, D.J. Genetic pool of the cultivated pear tree (Pyrus spp.) in the Canary Islands (Spain), studied using SSR molecular markers. Agronomy 2022, 12, 1711. [Google Scholar] [CrossRef]
- Draga, S.; Palumbo, F.; Miracolo Barbagiovanni, I.; Pati, F.; Barcaccia, G. Management of genetic erosion: The (successful) case study of the pear (Pyrus communis L.) germplasm of the Lazio region (Italy). Front. Plant Sci. 2023, 13, 1099420. [Google Scholar] [CrossRef]
- Coe, M.; Evans, K.; Gasic, K.; Main, D. Plant breeding capacity in U.S. public institutions. Crop Sci. 2020, 60, 2373–2385. [Google Scholar] [CrossRef]
- Katayama, H.; Uematsu, C. Pear (Pyrus species) genetic resources in Iwate, Japan. Genet. Resour. Crop. Evol. 2006, 53, 483–498. [Google Scholar] [CrossRef]
- Wang, M.; Hu, Z.; Wang, Y.; Zhao, W. Spatial distribution characteristics of suitable planting areas for Pyrus species under climate change in China. Plants 2023, 12, 1559. [Google Scholar] [CrossRef] [PubMed]
- Höfer, M.; Flachowsky, H. Cryopreservation of Malus and Pyrus wild species in the Fruit Genebank in Dresden-Pillnitz, Germany. Biology 2023, 12, 200. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Origin or Geographical Distribution—References or Sources |
|---|---|---|
| 1 | P. betulaefolia | North and Central China, Laos Southern Manchuria [19,25]. Syn. P. betulifolia, respectively, Sorbus betulaefolia? (https://powo.science.kew.org/, accessed on 10 March 2023). |
| 2 | P. canescens | Interspecific hybrid between P. nivalis × P. salicifolia [9]; https://www.genesys-pgr.org/t/Pyrus, http://www.worldfloraonline.org/ (accessed on 10 March 2023). |
| 3 | P. caucasica | Wild form of P. communis. The common varieties of pear are probably complex hybrids with var. pyraster and var. caucasica, and P. nivalis [25]. |
| 4 | P. communis | Southeast and Western Europe, Turkey. P. communis sensu lato (common pear) is the ancestor of most of the cultivated pears in Europe, America, Australia, and New Zealand [19,25,44,45]. |
| 5 | P. cordata | France, Spain, Turkey, and possibly south-west England [19,25], as well as Portugal [9]. Considered to be merely a form of P. communis [46], a possible relict from a fairly early stage in the evolution of Pyrus species [25]. |
| 6 | P. drovara | Accession of unknown provenance from the HRS and RIFG germplasm collection (https://www.genesys-pgr.org/t/Pyrus, accessed on 10 March 2023). |
| 7 | P. eleagrifolia | Southeast Europe, Turkey, Crimea [19,25]. It often appears under the name P. elaeagrifolia. |
| 8 | P. korshinskyi | Afghanistan, Kyrgyzstan, Tajikistan, Uzbekistan. Critically endangered—threatened by overgrazing and overharvesting [19]. |
| 9 | P. lindlezi | Accession of unknown provenance from the HRS and RIFG germplasm collection (https://www.genesys-pgr.org/t/Pyrus, accessed on 10 March 2023). |
| 10 | P. ×malifolia | Accession of unknown provenance from the HRS and RIFG germplasm collection (https://www.genesys-pgr.org/t/Pyrus, accessed on 10 March 2023). |
| 11 | P. persica | Syn. Pyrus spinosa (https://powo.science.kew.org/, accessed on 10 March 2023). |
| 12 | P. ussuriensis | Northern China, Manchuria, Korea, Siberia. One of the edible pears of which various forms were cultivated along with P. pyrifolia (also known as P. serotina) early in Chinese history [19,25]. |
| 13 | P. variolosa | Syn. P. pashia? [19]; https://powo.science.kew.org/, https://www.genesys-pgr.org/t/Pyrus (accessed on 10 March 2023). |
| 14 | ×Pyronia veitkii | Artificial hybrid of Pyrus and Cydonia oblonga [19]. |
| 15 | ×Sorbopyrus | A triploid selection developed in the early 1800s from a cross between Sorbus and Pyrus [9]. |
| 16 | P. eleagrifolia × Curé | Accession from the HRS collection whose provenance is unknown (according to the name, it is an interspecific hybrid, but this is debatable as Curé is a triploid cultivar) |
| 17 | P. longipes | Origin: Algeria [25]. Syn. P. cossonii is one of the three North African pear species [27]. Considered a relict species, close to P. communis [47]. |
| 18 | P. luxemburgiana | A clone of ×Pyronia veitkii (var. luxemburgiana?) [19]. |
| 19 | P. nivalis | Western, central, and southern Europe [25]; south–central Europe, Ukraine, France [19]. P. nivalis Jacq., called snow pear, is considered a subspecies of P. communis [29]. |
| 20 | P. pyraster | A subspecies of P. communis [29]. The trees grow wild throughout Europe and were domesticated as early as 300 BC, being the ancestral form of the European pear [45]. In some European countries, it is threatened; efforts are being made to maintain genetic resources through in situ and ex situ conservation [19]. |
| No | Primer | Sequence (5′–3′) |
|---|---|---|
| 1 | OPA20 | GTTGCGATCC |
| 2 | OPAL20 | AGGAGTCGGA |
| 3 | OPAB11 | GTGCGCAATG |
| 4 | AB11 | GTGCGCAATG |
| 5 | OPA01 | CAGGCCCTTC |
| 6 | OPB10 | CTGCTGGGAC |
| 7 | OPA17 | GACCGCTTGT |
| 8 | OPB08 | GTCCACACGG |
| 9 | OPB18 | CCACAGCAGT |
| 10 | OPC–14 | TGCGTGCTTG |
| 11 | OPC14 | TGCGTGCTTG |
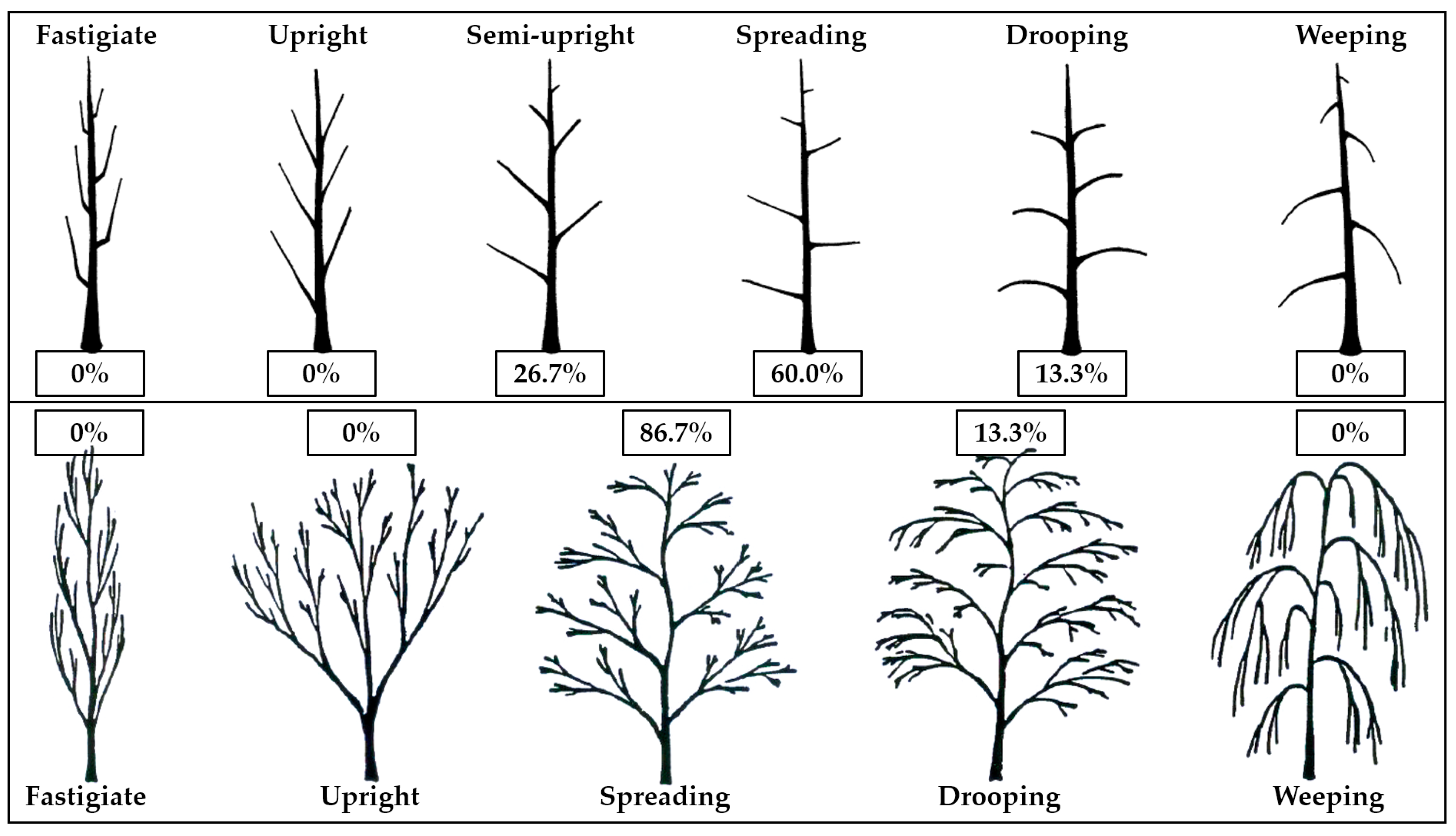
| No. | Species | Tree Habit (1–6 UPOV) 2 | Height of Trees (m) | Trunk Height (m) | Trunk Diameter (cm) | Crown Diameter (m) | No Branches Per Tree |
|---|---|---|---|---|---|---|---|
| 1 | P. betulaefolia | 4 | 3.15 a | 0.72 a,b | 12.50 e | 2.62 b | 6.00 b,c |
| 2 | P. canescens | 4 | 2.72 b | 0.79 a | 11.92 e | 2.00 c | 6.33 b |
| 3 | P. caucasica | 4 | 3.07 a | 0.61 b | 12.47 e | 2.60 b | 6.67 b |
| 4 | P. communis | 4 | 2.53 b | 0.77 a,b | 11.99 e | 1.92 c | 5.67 c |
| 5 | P. cordata | 3 | 2.72 b | 0.61 b | 10.54 f | 1.53 d | 4.67 d,e |
| 6 | P. drovara | 3 | 2.63 b | 0.76 a,b | 9.55 g | 1.83 c | 4.33 e |
| 7 | P. eleagrifolia | 4 | 2.92 a,b | 0.54 c | 15.88 b | 2.92 a | 5.67 c |
| 8 | P. korshinskyi | 4 | 2.97 a,b | 0.47 d | 13.46 d | 2.22 c | 4.33 e |
| 9 | P. lindlezi | 3 | 3.13 a | 0.81 a | 16.42 b | 2.53 b | 5.00 d |
| 10 | P. ×malifolia | 3 | 3.27 a | 0.56 c | 14.63 c | 2.42 b,c | 5.00 d |
| 11 | P. persica | 4 | 3.36 a | 0.56 c | 15.34 b,c | 2.06 c | 5.67 c |
| 12 | P. ussuriensis | 5 | 2.94 a,b | 0.63 d | 10.54 f | 2.22 c | 7.33 a |
| 13 | P. variolosa | 4 | 3.30 a | 0.56 c | 18.77 a | 2.13 c | 6.00 c |
| 14 | ×Pyronia veitkii | 4 | 2.93 a,b | 0.64 b | 11.30 e,f | 2.40 b,c | 6.33 b |
| 15 | ×Sorbopyrus | 5 | 2.38 c | 0.54 c | 9.98 g | 2.28 c | 3.67 f |
| No | Species | Leaf Length (cm) | Leaf Width (cm) | Petiole Length (cm) |
|---|---|---|---|---|
| 1 | P. betulaefolia | 4.57 g | 4.03 d | 4.53 b |
| 2 | P. canescens | 6.10 e | 2.52 f | 3.50 c |
| 3 | P. caucasica | 6.23 e | 3.30 e | 3.58 c |
| 4 | P. communis | 6.23 e | 5.20 b | 4.17 b,c |
| 5 | P. cordata | 5.38 f | 3.50 e | 3.14 d |
| 6 | P. drovara | 7.51 d | 4.17 d | 3.78 c |
| 7 | P. eleagrifolia | 10.67 a | 3.57 e | 3.48 c |
| 8 | P. korshinskyi | 9.78 b | 5.43 b | 2.11 e |
| 9 | P. lindlezi | 10.00 b | 6.67 a | 5.37 a |
| 10 | P. ×malifolia | 11.14 a | 6.20 a | 4.74 b |
| 11 | P. persica | 8.98 c | 4.83 c | 5.43 a |
| 12 | P. ussuriensis | 5.18 f | 3.52 e | 3.41 c |
| 13 | P. variolosa | 7.83 d | 5.21 b | 3.57 c |
| 14 | ×Pyronia veitkii | 6.17 e | 5.83 a | 4.67 b |
| 15 | ×Sorbopyrus | 4.78 g | 1.92 g | 1.73 f |
| No | Species | Corolla Diameter (mm) | Petal Length (mm) | Petal Width (mm) |
|---|---|---|---|---|
| 1 | P. betulaefolia | 39.58 b | 15.34 a,b | 12.68 b,c |
| 2 | P. canescens | 34.34 c | 14.20 c | 11.00 c |
| 3 | P. caucasica | 38.00 b | 16.34 a | 12.34 c |
| 4 | P. communis | 34.68 c | 11.67 d | 11.68 c |
| 5 | P. cordata | 30.10 d | 13.67 c | 12.20 c |
| 6 | P. drovara | 38.00 b | 16.34 a | 12.10 c |
| 7 | P. eleagrifolia | 34.34 c | 14.68 b | 11.68 c |
| 8 | P. korshinskyi | 28.33 d | 11.66 d | 14.20 b |
| 9 | P. lindlezi | 54.20 a | 17.00 a | 17.00 a |
| 10 | P. ×malifolia | 34.68 c | 15.00 b | 12.10 c |
| 11 | P. persica | 31.34 c,d | 14.68 b | 12.20 c |
| 12 | P. ussuriensis | 34.68 c | 14.66 b | 11.68 c |
| 13 | P. variolosa | 35.34 c | 13.68 c | 12.00 c |
| 14 | ×Pyronia veitkii | 30.34 d | 13.00 c,d | 8.33 d |
| 15 | ×Sorbopyrus | 35.32 c | 11.65 d | 11.00 c |
| No. | Species | Peduncle Length (cm) | Fruit Length (cm) | Fruit Diameter (cm) | Fruit Weight (g) | Fruit Firmness (kgf/cm2) |
|---|---|---|---|---|---|---|
| 1 | P. betulaefolia | 3.20 b | 1.93 g | 2.43 d | 6.23 i | 96.0 a |
| 2 | P. canescens | 1.97 d.e | 2.68 e | 0.75 f | 11.94 h | 90.0 b |
| 3 | P. caucasica | 2.37 d | 5.20 b | 5.53 a | 91.00 a | 81.7 d |
| 4 | P. communis | 1.87 e | 3.33 d | 0.87 f | 11.99 h | 83.7 d |
| 5 | P. cordata | 2.23 d | 2.77 e | 3.57 b | 19.13 f | 94.0 a |
| 6 | P. drovara | 1.43 f | 4.20 c | 3.53 b | 28.13 e | 83.0 d |
| 7 | P. eleagrifolia | 1.53 f | 2.13 g | 2.47 d | 7.30 i | 93.3 a |
| 8 | P. korshinskyi | 4.10 a | 1.73 h | 1.57 e | 2.70 j | 94.7 a |
| 9 | P. lindlezi | 2.30 d | 3.47 d | 3.13 b | 49.23 d | 84.1 d |
| 10 | P. ×malifolia | 2.23 d | 3.37 d | 0.55 f | 16.23 g | 94.0 a |
| 11 | P. persica | 2.23 d | 2.63 f | 3.37 b | 58.23 c | 82.3 d |
| 12 | P. ussuriensis | 2.20 d | 2.88 e | 0.73 f | 9.56 h | 87.7 c |
| 13 | P. variolosa | 2.83 c | 6.33 a | 5.23 a | 78.70 b | 80.2 d |
| 14 | ×Pyronia veitkii | 3.23 b | 2.73 e | 2.63 c | 8.48 i | 95.0 a |
| 15 | ×Sorbopyrus | 2.10 d | 3.10 d,e | 2.77 c | 14.97 g | 86.3 c |
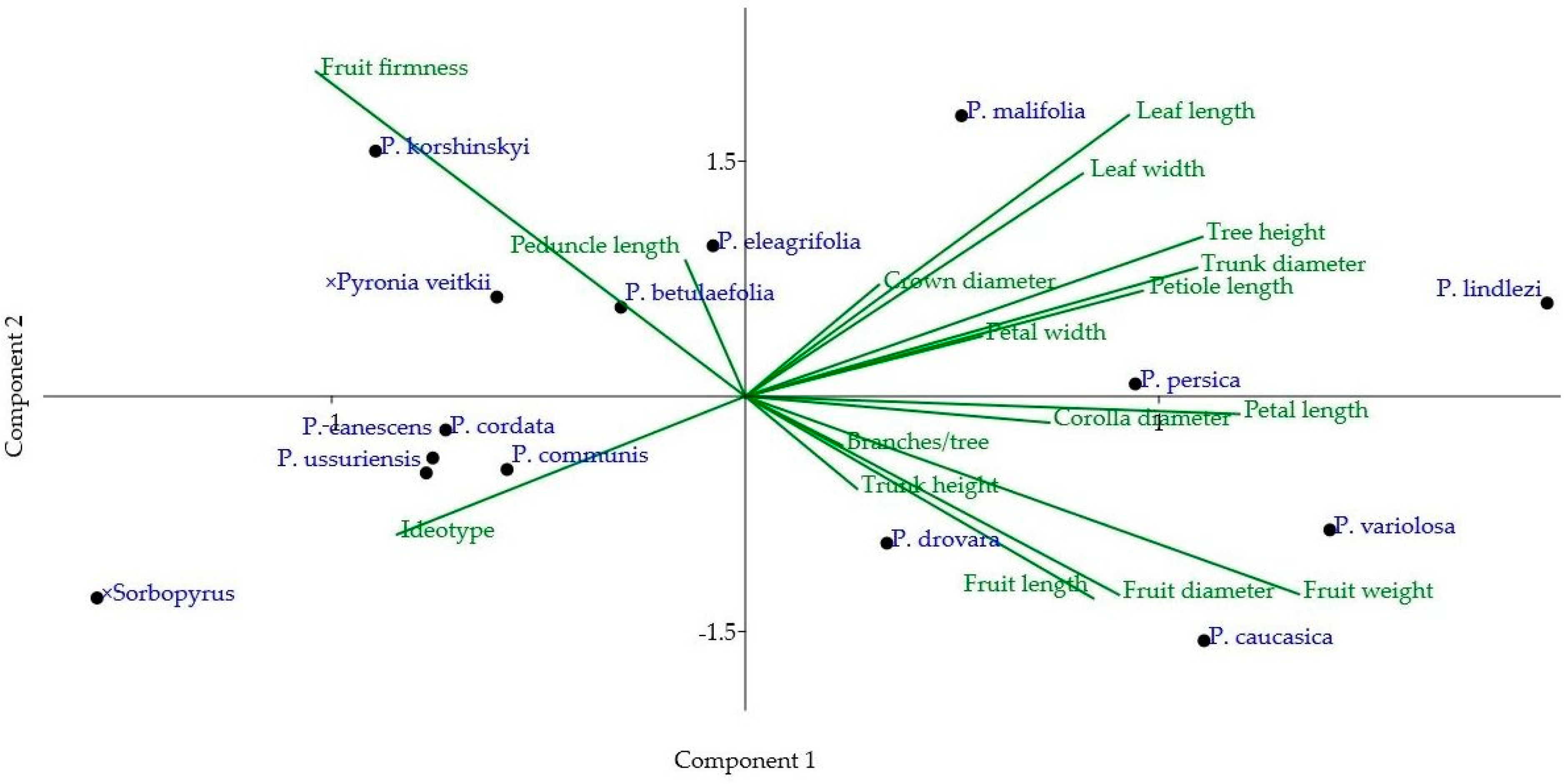
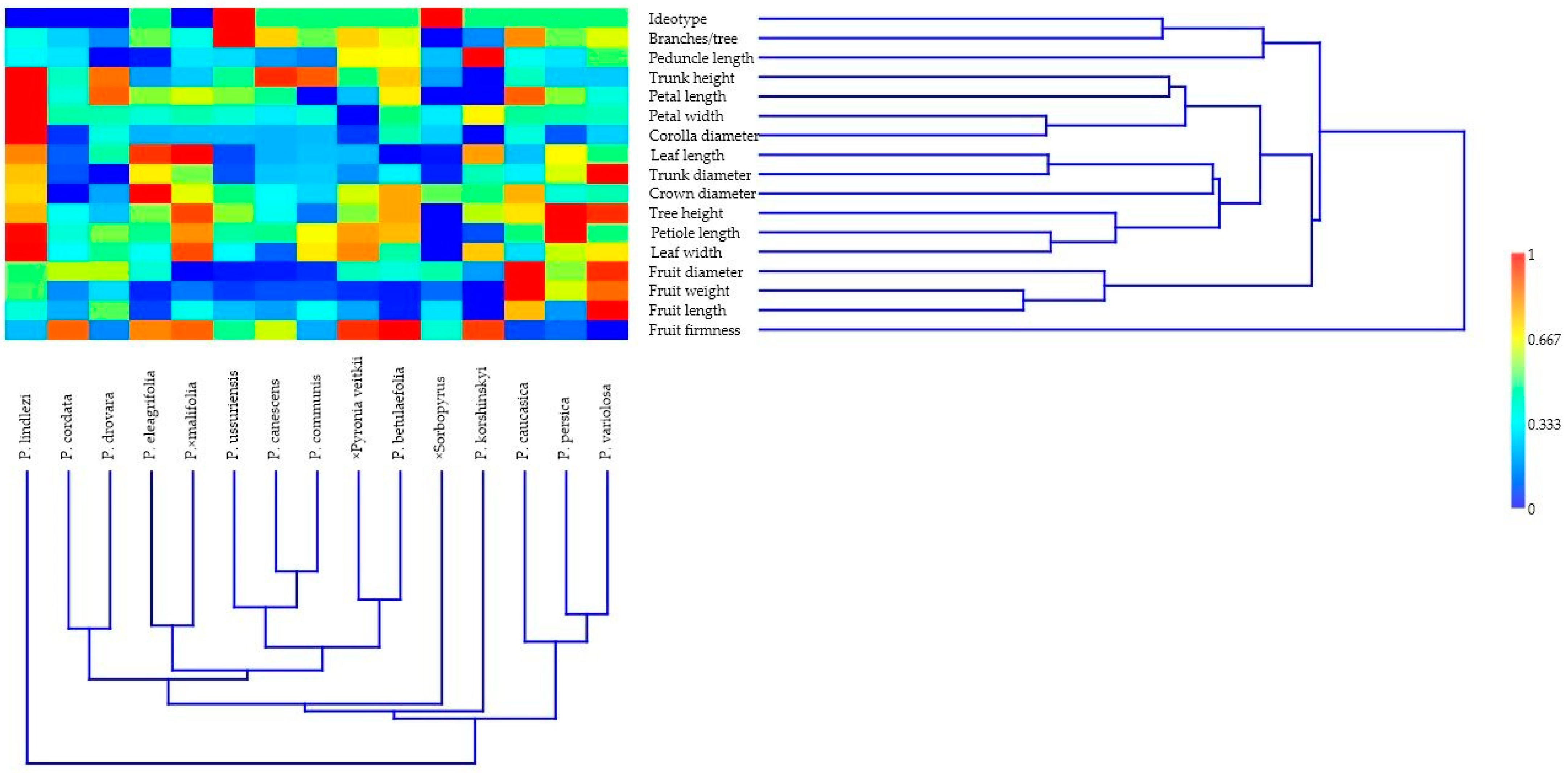
| Trunk Height | Trunk Diameter | Crown Diameter | Branches/Tree | Leaf Length | Leaf Width | Petiole Length | Corolla Diameter | Petal Length | Petal Width | Peduncle Length | Fruit Length | Fruit Diameter | Fruit Weight | Fruit Firmness | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tree height | –0.25 | 0.73 | 0.38 | 0.37 | 0.49 | 0.55 | 0.59 | 0.15 | 0.46 | 0.30 | 0.37 | 0.18 | 0.24 | 0.47 | –0.02 |
| Trunk height | –0.22 | –0.18 | 0.19 | –0.28 | 0.06 | 0.44 | 0.60 | 0.40 | 0.17 | –0.36 | 0.06 | –0.15 | –0.05 | –0.21 | |
| Trunk diameter | 0.39 | 0.12 | 0.67 | 0.52 | 0.38 | 0.27 | 0.20 | 0.41 | 0.16 | 0.30 | 0.26 | 0.47 | –0.21 | ||
| Crown diameter | 0.26 | 0.33 | 0.11 | 0.17 | 0.35 | 0.30 | 0.12 | 0.15 | –0.11 | 0.03 | 0.04 | 0.20 | |||
| Branches/tree | –0.25 | –0.03 | 0.35 | –0.02 | 0.26 | –0.29 | 0.03 | 0.14 | –0.03 | 0.20 | –0.08 | ||||
| Leaf length | 0.60 | 0.30 | 0.14 | 0.23 | 0.44 | –0.02 | –0.02 | –0.07 | 0.11 | 0.02 | |||||
| Leaf width | 0.62 | 0.24 | 0.12 | 0.36 | 0.36 | 0.08 | –0.06 | 0.11 | 0.02 | ||||||
| Petiole length | 0.41 | 0.56 | 0.15 | –0.12 | 0.04 | 0.01 | 0.25 | –0.12 | |||||||
| Corolla diameter | 0.65 | 0.66 | –0.23 | 0.26 | 0.19 | 0.31 | –0.35 | ||||||||
| Petal length | 0.41 | –0.33 | 0.26 | 0.32 | 0.44 | –0.23 | |||||||||
| Petal width | 0.10 | 0.02 | 0.11 | 0.25 | –0.20 | ||||||||||
| Peduncle length | –0.19 | 0.03 | –0.07 | 0.40 | |||||||||||
| Fruit length | 0.65 | 0.80 | –0.74 | ||||||||||||
| Fruit diameter | 0.81 | –0.50 | |||||||||||||
| Fruit weight | –0.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simionca Mărcășan, L.I.; Pop, R.; Somsai, P.A.; Oltean, I.; Popa, S.; Sestras, A.F.; Militaru, M.; Botu, M.; Sestras, R.E. Comparative Evaluation of Pyrus Species to Identify Possible Resources of Interest in Pear Breeding. Agronomy 2023, 13, 1264. https://doi.org/10.3390/agronomy13051264
Simionca Mărcășan LI, Pop R, Somsai PA, Oltean I, Popa S, Sestras AF, Militaru M, Botu M, Sestras RE. Comparative Evaluation of Pyrus Species to Identify Possible Resources of Interest in Pear Breeding. Agronomy. 2023; 13(5):1264. https://doi.org/10.3390/agronomy13051264
Chicago/Turabian StyleSimionca Mărcășan, Leontina I., Rodica Pop, Peter A. Somsai, Ion Oltean, Sergiu Popa, Adriana F. Sestras, Mădălina Militaru, Mihai Botu, and Radu E. Sestras. 2023. "Comparative Evaluation of Pyrus Species to Identify Possible Resources of Interest in Pear Breeding" Agronomy 13, no. 5: 1264. https://doi.org/10.3390/agronomy13051264
APA StyleSimionca Mărcășan, L. I., Pop, R., Somsai, P. A., Oltean, I., Popa, S., Sestras, A. F., Militaru, M., Botu, M., & Sestras, R. E. (2023). Comparative Evaluation of Pyrus Species to Identify Possible Resources of Interest in Pear Breeding. Agronomy, 13(5), 1264. https://doi.org/10.3390/agronomy13051264