
Genotypic and Environmental Effect on the Concentration of Phytochemical Contents of Lentil (Lens culinaris L.)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials
2.3. Sample Preparation
2.3.1. Extraction of Free Phenolics
2.3.2. Extraction of Lipophilic Antioxidants
2.4. Determination of Total Phenolic Content
2.5. Determination of Hydrolyzed Tannin Content
2.6. Determination of Total Flavonoid Content
2.7. Determination of Total Proanthocyanidin Content
2.8. Determination of Antioxidant Capacity
2.8.1. ABTS Radical Scavenging Assay
2.8.2. DPPH (2,2-Diphenyl-1-Picrylhydrazyl) Assay
2.8.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.9. Analysis of Phenolics by HPLC
2.10. Analysis of Tocopherols and Carotenoids
2.11. Statistical Analysis
3. Results and Discussion
3.1. Phytochemical Compounds of Lentil Genotypes
3.1.1. TPC, TFC, TPAC, TNC and Antioxidant Activity
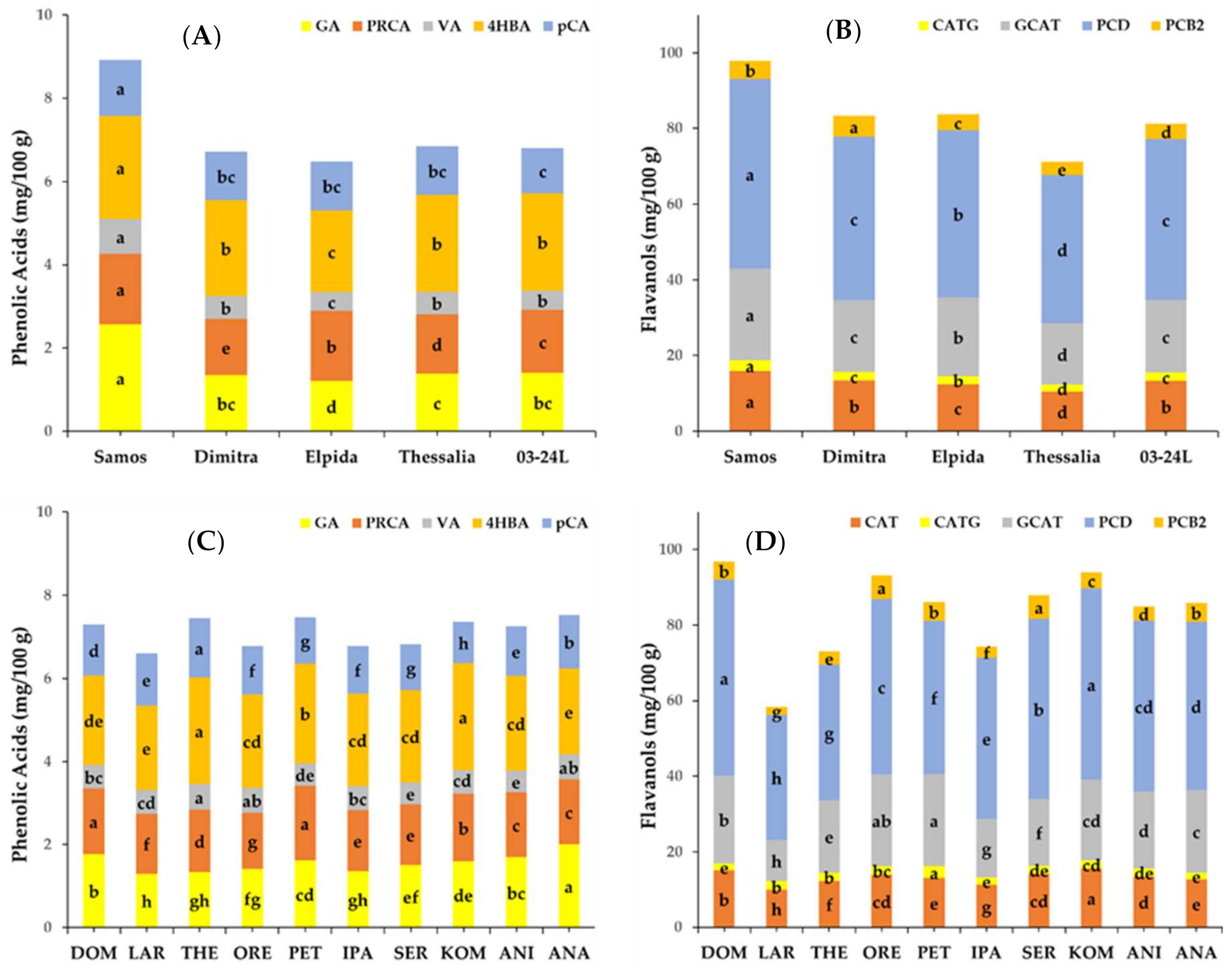
3.1.2. Quantification of Phenolic Compounds
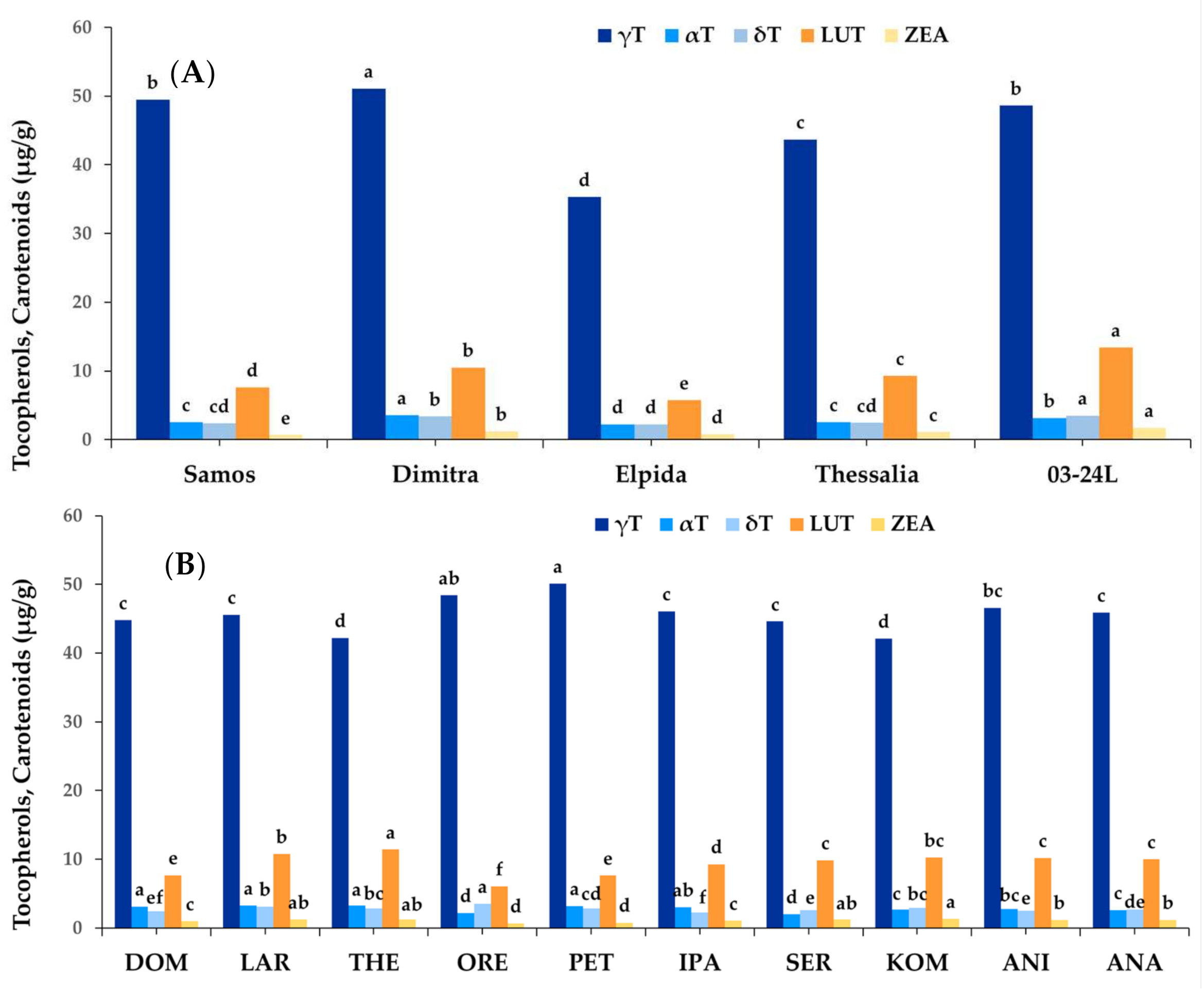
3.1.3. Tocopherols and Carotenoids
3.2. Effect of Genotype and Environment on Phytochemical Components
3.3. Correlation Coefficients among Phytochemicals Compounds
3.4. Multivariate Statistical Analyses
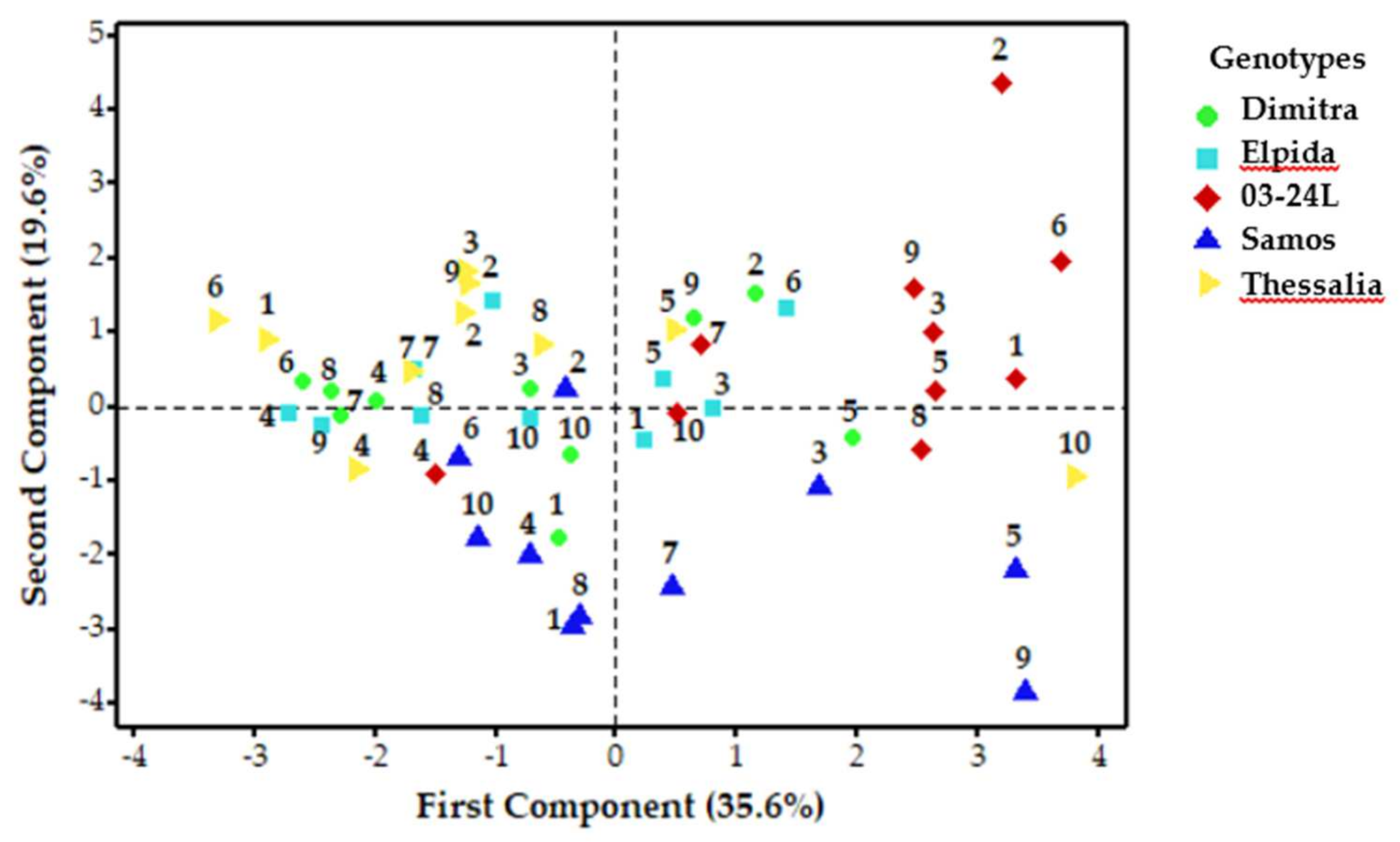
3.4.1. Variation among Genotypes × Locations
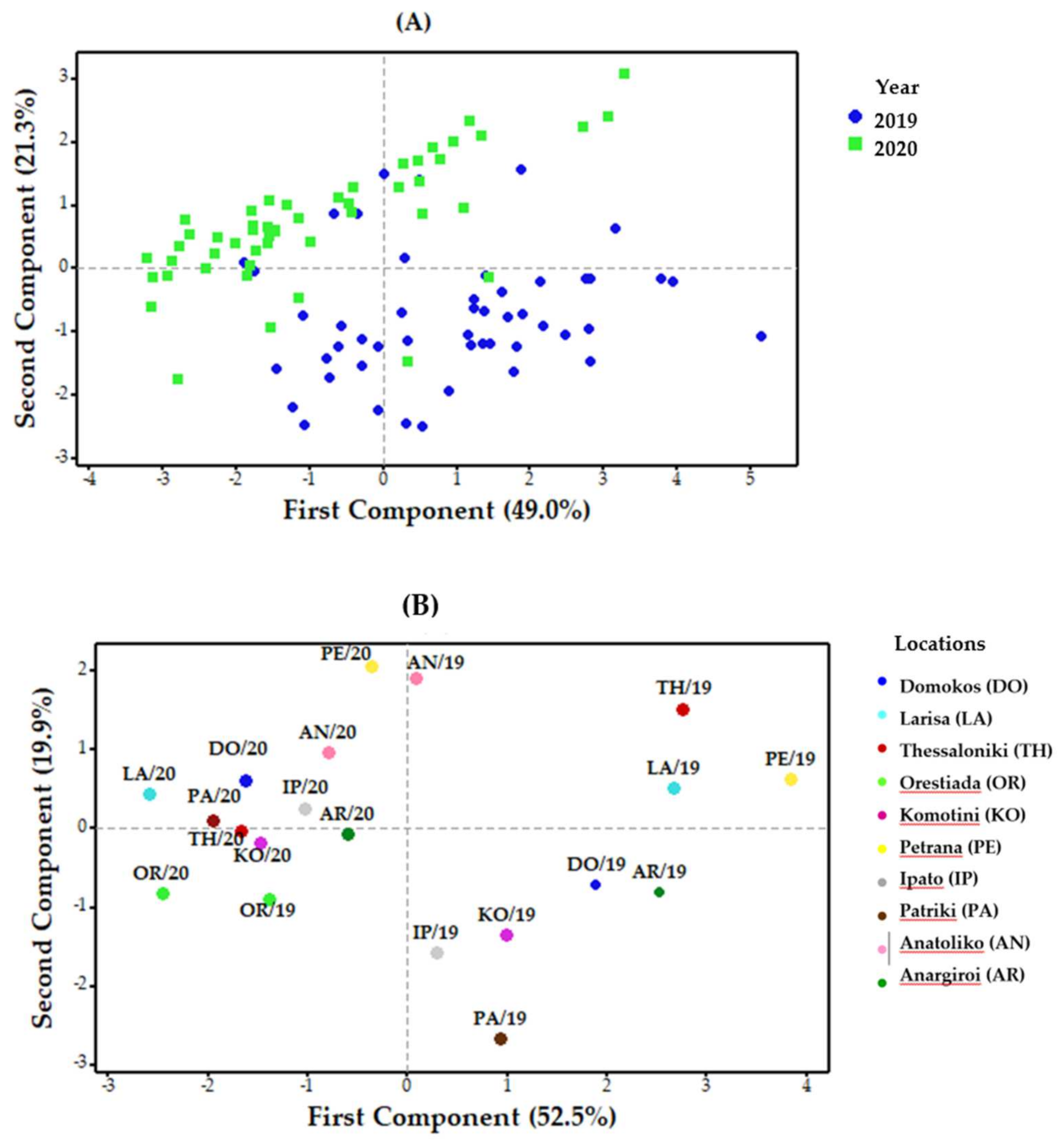
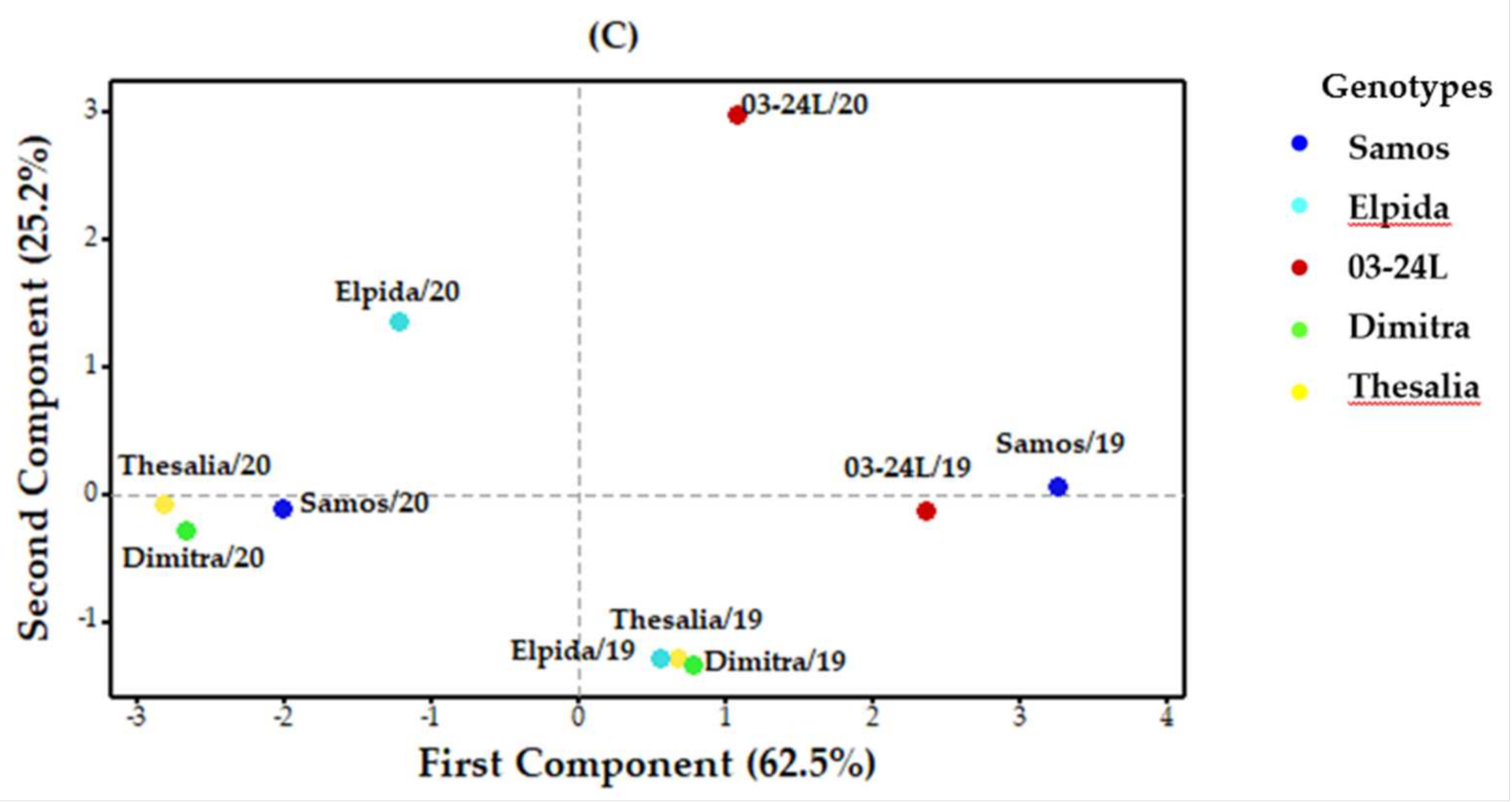
3.4.2. Variation Caused by Year
3.4.3. Variation Caused by Location
3.4.4. Variation Caused by Genotype
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liber, M.; Duarte, I.; Maia, A.T.; Oliveira, H.R. The history of lentil (Lens culinaris subsp. culinaris) domestication and spread as revealed byg enotyping-by-sequencing of wild and landrace accessions. Front. Plant Sci. 2021, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S.; Panozzo, J. Phytochemicals for health, the role of pulses. J. Agric. Food Chem. 2007, 55, 7981–7994. [Google Scholar] [CrossRef] [PubMed]
- Ajila, C.M.; Brar, S.K.; Verma, M.; Rao, U.P. Sustainable Solutions for Agro Processing Waste Management: An Overview. In Environmental Protection Strategies for Sustainable Development; Malik, A., Grohmann, E., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 65–109. [Google Scholar]
- Zhang, B.; Deng, Z.; Tang, Y.; Chen, P.; Liu, R.; Ramdath, D.D.; Liu, Q.; Hernandez, M.; Tsao, R. Fatty acid, carotenoid and tocopherol compositions of 20 Canadian lentil cultivars and synergistic contribution to antioxidant activities. Food Chem. 2014, 161, 296–304. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Amdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Baojun, X.; Chang, S.K.C. Phytochemical profiles and health-promoting effects of cool-season food legumes as influenced by thermal processing. J. Agric. Food Chem. 2009, 57, 10718–10731. [Google Scholar]
- Amarowicz, R.; Estrella, I.; Hernández, T.; Dueñas, M.; Troszyńska, A.; Kosińska, A.; Pegg, R.B. Antioxidant activity of a red lentil extract and its fractions. Int. J. Mol. Sci. 2009, 10, 5513–5527. [Google Scholar] [CrossRef]
- Amarowicz, R.; Estrella, I.; Hernández, T.; Robredo, S.; Troszynska, A.; Kosinska, A.; Pegg, R.B. Free radical-scavenging capacity, antioxidant activity, and phenolic composition of green lentil (Lens culinaris). Food Chem. 2010, 121, 705–711. [Google Scholar] [CrossRef]
- Fratianni, F.; Cardinale, F.; Cozzolino, A.; Granese, T.; Albanese, D.; Matteo, M.D.; Zaccardelli, M.; Coppola, R.; Nazzaro, F. Polyphenol composition and antioxidant activity of different grass pea (Lathyrus sativus), lentils (Lens culinaris), and chickpea (Cicer arietinum) ecotypes of the Campania region (Southern Italy). J. Funct. Foods 2014, 7, 551–557. [Google Scholar] [CrossRef]
- Duenas, M.; Sarmento, T.; Aguilera, Y.; Benitez, Y.V.; Molla, E.; Esteban, R.M.; Martín-Cabrejas, M.A. Impact of cooking and germination on phenolic composition and dietary fibre fractions in dark beans (Phaseolus vulgaris L.) and lentils (Lens culinaris L.). LWT Food Sci. Technol. 2016, 66, 72–78. [Google Scholar] [CrossRef]
- Sun, Y.; Deng, Z.; Liu, R.; Zhang, H.; Zhu, H.; Jiang, L.; Tsao, R. A comprehensive profiling of free, conjugated and bound phenolics and lipophilic antioxidants in red and green lentil processing by-products. Food Chem. 2020, 325, 126925–126937. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Duenas, M.; Estrella, I.; Hernandez, T.; Benitez, V.; Esteban, R.M.; Martin-Cabrejas, M.A. Evaluation of phenolic profile and antioxidant properties of pardina lentil as affected by industrial dehydration. J. Agric. Food Chem. 2010, 58, 10101–10108. [Google Scholar] [CrossRef]
- Yeo, J.; Shahidi, F. Identification and quantification of soluble and insoluble-bound phenolics in lentil hulls using HPLC-ESI-MS/MS and their antioxidant potential. Food Chem. 2020, 315, 126202–126210. [Google Scholar] [CrossRef]
- Mirali, M.; Purves, R.W.; Vandenberg, A. Profiling the phenolic compounds of the four major seed coat types and their relation to color genes in lentil. J. Nat. Prod. 2017, 80, 1310–1317. [Google Scholar] [CrossRef] [PubMed]
- Kushi, L.H.; Meyer, K.A.; Jacobs, D.R. Cereal, legumes, and chronic disease risk reduction: Evidence from epidemiologic studies. Am. J. Clin. Nutr. 1999, 70, 451s–458s. [Google Scholar] [CrossRef] [PubMed]
- Adebamowo, C.A.; Cho, E.; Sampson, L.; Katan, M.B.; Spiegelman, D.; Willett, W.C.; Holmes, M.D. Dietary flavonols and flavonol-rich foods intake and the risk of breast cancer. Intern. J. Cancer 2005, 114, 628–633. [Google Scholar] [CrossRef]
- Papanikolaou, Y.; Fulgoni, V.L. Bean consumption is associated withgreater nutrient intake, reduced systolic blood pressure, lower body weight, and a smaller waist circumference in adults: Results from the National Health and Nutrition Examination Survey 1999–2002. J. Am. Coll. Nutr. 2008, 27, 569–576. [Google Scholar] [CrossRef]
- Boschin, G.; Arnoldi, A. Legumes are valuable sources of tocopherols. Food Chem. 2011, 127, 1199–1203. [Google Scholar] [CrossRef]
- Knekt, P.; Reunanen, A.; Jarvinen, R.; Seppanen, R.; Heliovaara, M.; Aromaa, A. Antioxidant vitamin intake and coronarymortality in a longitudinal population study. Am. J. Epidemiol. 1994, 139, 1180–1189. [Google Scholar] [CrossRef]
- Tucker, J.M.; Townsend, D.M. Alpha-tocopherol: Roles inprevention and therapy of human disease. Biomed. Pharmacother. 2005, 59, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Padhi, E.M.T.; Liu, R.; Hernandez, M.; Tsao, R.; Ramdath, D.D. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J. Funct. Foods 2017, 38, 602–611. [Google Scholar] [CrossRef]
- Erskine, W.; Williams, P.C.; Nakkoul, H. Genetic and environmental variation in the seed size, protein, yield, and cooking quality of lentils. Field Crop. Res. 1985, 12, 153–161. [Google Scholar] [CrossRef]
- Biçer, N.T.; Sakar, D. Evaluation of some lentil genotypes at different locations in Turkey. Int. J. Agric. Biol. 2004, 6, 317–320. [Google Scholar]
- Shrestha, R.; Rizvi, A.H.; Sarker, A.; Darai, R.; Paneru, R.B.; Vandenberg, A.; Singh, M. Genotypic variability and genotype × environment interaction for iron and zinc content in lentil under Nepalese environments. Crop Sci. 2018, 58, 2503–2510. [Google Scholar] [CrossRef]
- Thavarajah, D.; Thavarajah, P.; Wejesuriya, A.; Rutzke, M.; Glahn, R.P.; Combs, G.F.; Vandenberg, A. The potential of lentil (Lens culinaris L.) as a whole food for increased selenium, iron, and zinc intake: Preliminary results from a 3 year study. Euphytica 2011, 180, 123–128. [Google Scholar] [CrossRef]
- Boudjou, S.; Oomah, B.D.; Zaidi, F.; Hosseinian, F. Phenolics content and antioxidant and anti-inflammatory activities of legume fractions. Food Chem. 2012, 138, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Ninou, E.; Papathanasiou, F.; Vlachostergios, D.N.; Mylonas, I.; Kargiotidou, A.; Pankou, C.; Papadopoulos, I.; Sinapidou, E.; Tokatlidis, I. Intense breeding within lentil landraces for high-yielding pure lines sustained the seed quality characteristics. Agriculture 2019, 9, 175. [Google Scholar] [CrossRef]
- Vlachostergios, D.N.; Tzantarmas, C.; Kargiotidou, A.; Ninou, E.; Pankou, C.; Gaintatzi, C.; Mylonas, I.; Papadopoulos, I.; Foti, C.; Chatzivassiliou, E.K.; et al. Single-plant selection within lentil landraces at ultra-low density: A short-time tool to breed high yielding and stable varieties across divergent environments. Euphytica 2018, 214, 58–73. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K.C. Phenolic substance characterization and chemical and cell-Based antioxidant activities of 11 lentils grown in the Northern United States. J. Agric. Food Chem. 2010, 58, 1509–1517. [Google Scholar] [CrossRef]
- Dave, B.; Caspar, O.F.; Malcolmson, L.J.; Bellido, A.-S. Phenolics and antioxidant activity of lentil and pea hulls. Food Res. Int. 2011, 44, 436–441. [Google Scholar]
- Tsialtas, I.Τ.; Irakli, M.; Lazaridou, A. Traits related to bruchid resistance and its parasitoid in vetch seeds. Euphytica 2018, 214, 238–250. [Google Scholar] [CrossRef]
- Irakli, M.; Chatzopoulou, P.; Kadoglidou, K.; Tsivelika, N. Optimization and development of a high-performance liquid chromatography method for the simultaneous determination of vitamin E and carotenoids in tomato fruits. J. Sep. Sci. 2016, 39, 3348–3356. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventοs, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1998, 299, 152–178. [Google Scholar]
- Makkar, H.P.S.; Bluemmel, M.; Borowy, N.K.; Becker, R.K. Gravimetric determination of tannins and their correlate ions with chemical and protein precipitation methods. J. Sci. Food Agric. 1993, 61, 161–165. [Google Scholar] [CrossRef]
- Bao, J.S.; Cai, Y.; Sun, M.; Wang, G.Y.; Corke, H. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin anddelphinidin. Phytochemistry 1986, 25, 223–230. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C.A. Antioxidant activity applyingan improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Benzie, F.; Strain, J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activityof biological fluids and modified version for simultaneous measurement of total antioxidant power andascorbic acid concentration. Methods Enzymol. 1999, 299, 15–23. [Google Scholar]
- Skendi, A.; Irakli, M.; Chatzopoulou, P. Analysis of phenolic compounds in Greek plants of Lamiaceae family by HPLC. J. Appl. Res. Med. Aromat. Plants 2017, 6, 62–69. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw Hill: New York, NY, USA, 1980; p. 672. [Google Scholar]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- Menga, V.; Codianni, P.; Fares, C. Agronomic management under organic farming may affect the bioactive compounds of lentil (Lens Culinaris L.) and grass pea (Lathyrus Communis L). Sustainability 2014, 6, 1059–1075. [Google Scholar] [CrossRef]
- Lee, Y.; Yeo, Y.S.; Park, S.Y.; Lee, S.G.; Lee, S.M.; Cho, H.S.; Chung, N.J.; Oh, S.W. Compositional analysis of lentil (Lens culinaris) cultivars related to colors and their antioxidative activity. Plant Breed. Biotechnol. 2017, 5, 192–203. [Google Scholar] [CrossRef]
- Shao, Y.; Xu, F.; Chen, Y.; Huang, Y.; Beta, T.; Bao, J. Analysis of genotype, environment, and their interaction effects on the phytochemicals and antioxidant capacities of red rice (Oryza sativa L.). Cereal Chem. 2014, 92, 204–210. [Google Scholar] [CrossRef]
- Manach, C.; Augustin, S.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef]
- Kishore, G.; Ranjan, S.; Pandey, A.; Gupta, S. Influence of altitudinal variation on the antioxidant potential of tartar buckwheat of western Himalaya. Food Sci. Biotechnol. 2010, 19, 1355–1363. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Karjalainen, R.O. High-performance liquid chromatography analysis of black currant (Ribes nigrum L.) fruit phenolics grown either conventionally or organically. J. Agric. Food Chem. 2006, 54, 7530–7538. [Google Scholar] [CrossRef] [PubMed]
- Muzhingi, T.; Palacios-Rojas, N.; Miranda, A.; Cabrera, M.L.; Yeum, K.J.; Tang, G. Genetic variation of carotenoids, vitamin E and phenolic compounds in Provitamin A biofortified maize. J. Sci. Food Agric. 2016, 97, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Baderschneider, B.; Winterhalter, P. Isolation and characterization of novel benzoates, cinnamates, flavonoids and lignans from Riesling wine and screening for antioxidant activity. J. Agric. Food Chem. 2001, 49, 2788–2798. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Isolani, L.; Arfaioli, P.; Ghiselli, L.; Romani, A. Polyphenol content of modern and old varieties of triticum aestivuml. And t. durum. Desf. grains in two years of production. J. Agric. Food Chem. 2010, 58, 7329–7334. [Google Scholar] [CrossRef] [PubMed]
- Martini, D.; Taddei, F.; Nicoletti, I.; Ciccoritti, R.; Corradini, D.; Degidio, M.G. Effects of genotype and environment on phenolic acids content and total antioxidant capacity in durum wheat. Cereal Chem. 2014, 91, 310–317. [Google Scholar] [CrossRef]
- Vagiri, M.; Ekholm, A.; Öberg, E.; Johansson, E.; Andersson, S.C.; Rumpunen, K. Phenols and ascorbic acid in black currants (Ribes nigrum L.): Variation due to genotype, location, and year. J. Agric. Food Chem. 2013, 61, 9298–9306. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, S.S.; Khan, A.M.; Ammar, M.H.; El-Harty, E.H.; Migdadi, H.M.; El-Khalik, S.M.A.; Al-Shameri, A.M.; Javed, M.M.; Al-Faifi, S.A. Phenological, nutritional and molecular diversity assessment among 35 introduced lentil (Lens culinaris Medik.) genotypes grown in Saudi Arabia. Int. J. Mol. Sci. 2014, 15, 277–295. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locations | Latitiude/ Longitude | Altitude | MT 1 (°C) | AP 2 (mm) | Soil Type | pH | OM 3 (%) |
|---|---|---|---|---|---|---|---|
| Domokos | 39°1′13″ N/22°19′74″ E | 500 | 11.4 | 625.4 | C/CL | 7.1 | 1.7 |
| Ipato | 38°22′49″ N/23°22′11″ E | 118 | 14.6 | 704.1 | C/CL | 7.8 | 1.5 |
| Orestiada | 41°30′14″ N/26°32′99″ E | 26 | 12.6 | 399.9 | C/SiCL | 7.5 | 1.7 |
| Patriki | 40°53′90″ N/23°36′57″ E | 50 | 13.7 | 538.1 | C/CL | 8.0 | 1.3 |
| Anatoliko | 39°36′81″ N/22°25′94″ E | 624 | 10.6 | 480.4 | C/CL | 7.2 | 1.3 |
| Larissa | 39°36′81″ N/22°25′94″ E | 77 | 14.3 | 466.2 | C/C | 7.4 | 1.2 |
| Ag. Anargiri | 39°29′89″ N/22°21′20″ E | 121 | 14.1 | 500.5 | C/CL | 7.4 | 1.5 |
| Komotini | 41°5′31″ N/25°20′64″ E | 32 | 13.8 | 558.4 | C/SC | 8.1 | 1.3 |
| Thessaloniki | 40°32′69″ N/22°59′83″ E | 5 | 14.2 | 462.2 | CL/CL | 8.1 | 1.0 |
| Petrana | 40°15′59″ N/21°52′70″ E | 476 | 11.9 | 476.6 | C/CL | 7.9 | 2.2 |
| TPC | TFC | TNC | TPAC | ABTS | DPPH | FRAP | |
|---|---|---|---|---|---|---|---|
| Genotypes | |||||||
| Samos | 5.90 ± 0.50 b | 5.31 ± 1.12 a | 3.07 ± 0.72 b | 8.93 ± 2.30 a | 19.25 ± 2.61 b | 11.63 ± 1.23 bc | 12.10 ± 3.13 c |
| Dimitra | 5.66 ± 0.35 c | 4.77 ± 0.71 b | 2.96 ± 0.87 c | 7.76 ± 1.10 c | 18.47 ± 2.14 d | 11.19 ± 1.33 d | 12.12 ± 2.43 c |
| Elpida | 5.89 ± 0.50 b | 4.80 ± 0.57 b | 3.11 ± 1.02 b | 7.75 ± 0.98 c | 18.93 ± 1.67 c | 11.82 ± 1.40 b | 12.35 ± 1.92 b |
| Thessalia | 5.70 ± 0.62 c | 4.65 ± 0.77 c | 2.86 ± 1.09 d | 7.27 ± 1.69 d | 18.44 ± 2.37 d | 11.50 ± 1.56 c | 12.23 ± 2.22 bc |
| 03-24L | 6.38 ± 0.44 a | 5.24 ± 0.65 a | 3.20 ± 1.04 a | 7.94 ± 1.42 b | 20.02 ± 1.84 a | 13.05 ± 1.60 a | 13.84 ± 2.54 a |
| Mean | 5.90 ± 0.50 | 4.95 ± 0.83 | 3.04 ± 0.96 | 7.93 ± 1.66 | 19.03 ± 2.22 | 11.82 ± 1.55 | 12.53 ± 2.55 |
| Locations | |||||||
| Domokos | 5.89 ± 0.506 bcd | 5.09 ± 0.74 a | 3.00 ± 0.98 bcd | 8.28 ± 0.94 ab | 18.50 ± 2.54 bc | 11.89 ± 1.87 bc | 12.71 ± 2.68 a–d |
| Larissa | 5.87 ± 0.42 bcd | 4.76 ± 0.90 cd | 3.13 ± 0.74 bc | 6.83 ± 1.30 d | 20.18 ± 3.20 a | 12.30 ± 1.52 ab | 13.12 ± 2.79 abc |
| Thes-niki | 6.19 ± 0.67 a | 5.31 ± 0.92 ab | 3.06 ± 1.00 bcd | 7.95 ± 1.43 abc | 18.61 ± 1.90 bc | 11.99 ± 1.04 bc | 12.91 ± 3.23 abc |
| Orestiada | 5.87 ± 0.52 cd | 4.34 ± 0.34 d | 3.44 ± 0.32 ab | 7.55 ± 0.87 bcd | 17.50 ± 1.41 c | 11.50 ± 1.08 c | 10.93 ± 1.92 d |
| Petrana | 6.12 ± 0.40 ab | 5.28 ± 1.13 ab | 2.99 ± 0.92 bcd | 8.46 ± 1.98 a | 20.65 ± 1.98 a | 13.05 ± 0.84 a | 14.04 ± 2.10 a |
| Ipato | 5.67 ± 0.85 cd | 4.66 ± 0.42 cd | 2.92 ± 0.99 bcd | 7.19 ± 0.72 cd | 19.75 ± 2.06 ab | 11.68 ± 2.33 bc | 12.72 ± 2.17 a–d |
| Patriki | 5.62 ± 0.37 d | 4.75 ± 0.45 cd | 2.67 ± 1.22 cd | 8.31 ± 1.50 ab | 18.44 ± 1.53 bc | 11.36 ± 1.45 cd | 12.02 ± 2.06 bcd |
| Komotini | 5.91 ± 0.40 abc | 4.87 ± 0.59 bc | 2.84 ± 0.99 bcd | 8.24 ± 1.30 ab | 18.56 ± 1.51 bc | 11.12 ± 1.29 cd | 12.15 ± 2.45 bcd |
| Ag. Anargiri | 5.80 ± 0.48 cd | 5.42 ± 1.14 a | 2.42 ± 0.89 d | 7.84 ± 3.18 abc | 19.62 ± 2.23 ab | 11.97 ± 0.20 bc | 13.28 ± 2.75 ab |
| Anatoliko | 6.10 ± 0.44 ab | 5.04 ± 0.64 abc | 3.98 ± 0.27 a | 8.65 ± 1.08 a | 18.39 ± 1.28 bc | 12.41 ± 0.24 ab | 11.36 ± 1.44 cd |
| Mean | 5.90 ± 0.55 | 4.95 ± 0.82 | 3.04 ± 0.96 | 7.93 ± 1.65 | 19.02 ± 2.22 | 11.83 ± 1.56 | 12.53 ± 2.55 |
| Genotype (G) | Environment (E) | G×E | Error | |||||
|---|---|---|---|---|---|---|---|---|
| SS | % | SS | % | SS | % | SS | % | |
| Degree of Freedom | 4 | 19 | 76 | 200 | ||||
| Bioactive Components | ||||||||
| TPC, mg GAE/g | 20.1 | 22 | 18.6 | 20 | 48.6 | 53 | 4.1 | 5 |
| TFC, mg CATE/g | 21.5 | 10 | 101.3 | 49 | 41.55 | 38 | 5.4 | 3 |
| TPAC, mg PCB/g | 89.9 | 11 | 176.1 | 21 | 539.3 | 66 | 14.4 | 2 |
| TNC, mg GAE/g | 4.2 | 2 | 201.8 | 73 | 66.5 | 24 | 3.1 | 1 |
| Phenolic acids, mg/100 g | 128.2 | 56 | 17.1 | 8 | 77.7 | 34 | 5.1 | 2 |
| Flavanols, mg/100 g | 10,929.8 | 24 | 18,391.9 | 41 | 15,013.3 | 33 | 661.3 | 2 |
| Tocopherols, μg/g | 5260.5 | 60 | 873.3 | 10 | 2306.2 | 26 | 385.4 | 4 |
| Carotenoids, μg/g | 1308.3 | 61 | 474.7 | 23 | 314.8 | 15 | 26.3 | 1 |
| *** | *** | *** | ||||||
| Antioxidant Capacity | ||||||||
| ABTS | 102.1 | 7 | 746.7 | 51 | 562.8 | 38 | 58.0 | 4 |
| DPPH | 123.6 | 17 | 162.0 | 22 | 387.4 | 53 | 53.8 | 8 |
| FRAP | 133.1 | 7 | 1122.4 | 58 | 663.7 | 34 | 30.6 | 1 |
| *** | *** | *** | ||||||
| Parameters | TPC | TFC | TPAC | TNC | Pas | Fos | Ts | Cars | ABTS | DPPH | FRAP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TPC | 1 | 0.51 *** | 0.15 | 0.29 *** | 0.23 ** | 0.18 * | 0.26 *** | 0.26 *** | 0.19 * | 0.46 *** | 0.41 *** |
| TFC | 1 | 0.45 *** | −0.14 | 0.35 *** | 0.15 | 0.34 *** | 0.22 ** | 0.51 *** | 0.63 *** | 0.67 *** | |
| TPAC | 1 | 0.17 * | 0.49 *** | 0.27 *** | 0.26 *** | −0.19 * | 0.11 | 0.25 ** | 0.21 ** | ||
| TNC | 1 | 0.19 * | −0.08 | 0.09 | −0.16 * | −0.25 ** | 0.05 | −0.33 ** | |||
| Pas | 1 | 0.59 *** | 0.20 * | −0.21 ** | 0.19 * | 0.28 *** | 0.05 | ||||
| Fos | 1 | 0.16 * | −0.15 | −0.00 | −0.05 | 0.01 | |||||
| Ts | 1 | 0.35 *** | 0.21 * | 0.19 * | 0.23 ** | ||||||
| Cars | 1 | 0.23 ** | 0.33 *** | 0.39 *** | |||||||
| ABTS | 1 | 0.51 *** | 0.60 *** | ||||||||
| DPPH | 1 | 0.50 *** | |||||||||
| FRAP | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irakli, M.; Kargiotidou, A.; Tigka, E.; Beslemes, D.; Fournomiti, M.; Pankou, C.; Stavroula, K.; Tsivelika, N.; Vlachostergios, D.N. Genotypic and Environmental Effect on the Concentration of Phytochemical Contents of Lentil (Lens culinaris L.). Agronomy 2021, 11, 1154. https://doi.org/10.3390/agronomy11061154
Irakli M, Kargiotidou A, Tigka E, Beslemes D, Fournomiti M, Pankou C, Stavroula K, Tsivelika N, Vlachostergios DN. Genotypic and Environmental Effect on the Concentration of Phytochemical Contents of Lentil (Lens culinaris L.). Agronomy. 2021; 11(6):1154. https://doi.org/10.3390/agronomy11061154
Chicago/Turabian StyleIrakli, Maria, Anastasia Kargiotidou, Evangelia Tigka, Dimitrios Beslemes, Maria Fournomiti, Chrysanthi Pankou, Kostoula Stavroula, Nektaria Tsivelika, and Dimitrios N. Vlachostergios. 2021. "Genotypic and Environmental Effect on the Concentration of Phytochemical Contents of Lentil (Lens culinaris L.)" Agronomy 11, no. 6: 1154. https://doi.org/10.3390/agronomy11061154
APA StyleIrakli, M., Kargiotidou, A., Tigka, E., Beslemes, D., Fournomiti, M., Pankou, C., Stavroula, K., Tsivelika, N., & Vlachostergios, D. N. (2021). Genotypic and Environmental Effect on the Concentration of Phytochemical Contents of Lentil (Lens culinaris L.). Agronomy, 11(6), 1154. https://doi.org/10.3390/agronomy11061154