
In Vitro Polyploidization of Thymus vulgaris L. and Its Effect on Composition of Essential Oils

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Seed Germination and Shoot Multiplication
2.2. In Vitro Polyploidization
- (1)
- Cultivation of nodal segments on oryzalin medium for 2 weeks (CMNS)
- (2)
- Overflow of nodal segments with oryzalin solution for 24 h (ONS)
2.3. Multiplication of Polyploid Plants, Rooting and Acclimatization
2.4. Flow Cytometry (FCM)
2.5. Gas Chromatography/Mass Spectrometry (GC/MS)
2.6. Insects
2.7. Acute Toxicity on C. quinquefasciatus and S. littoralis
3. Results
3.1. Multiplication
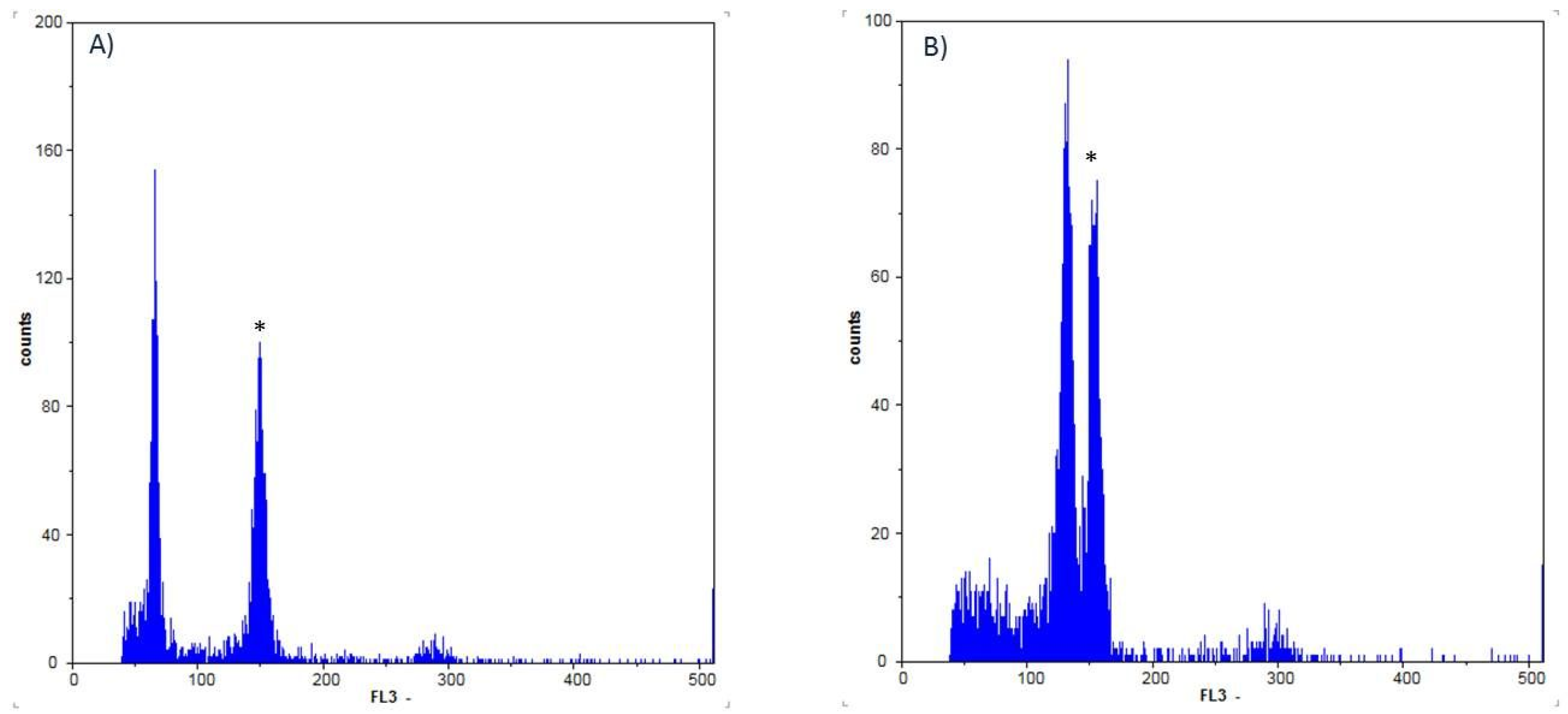
3.2. In Vitro Polyploidization, Ploidy Determination
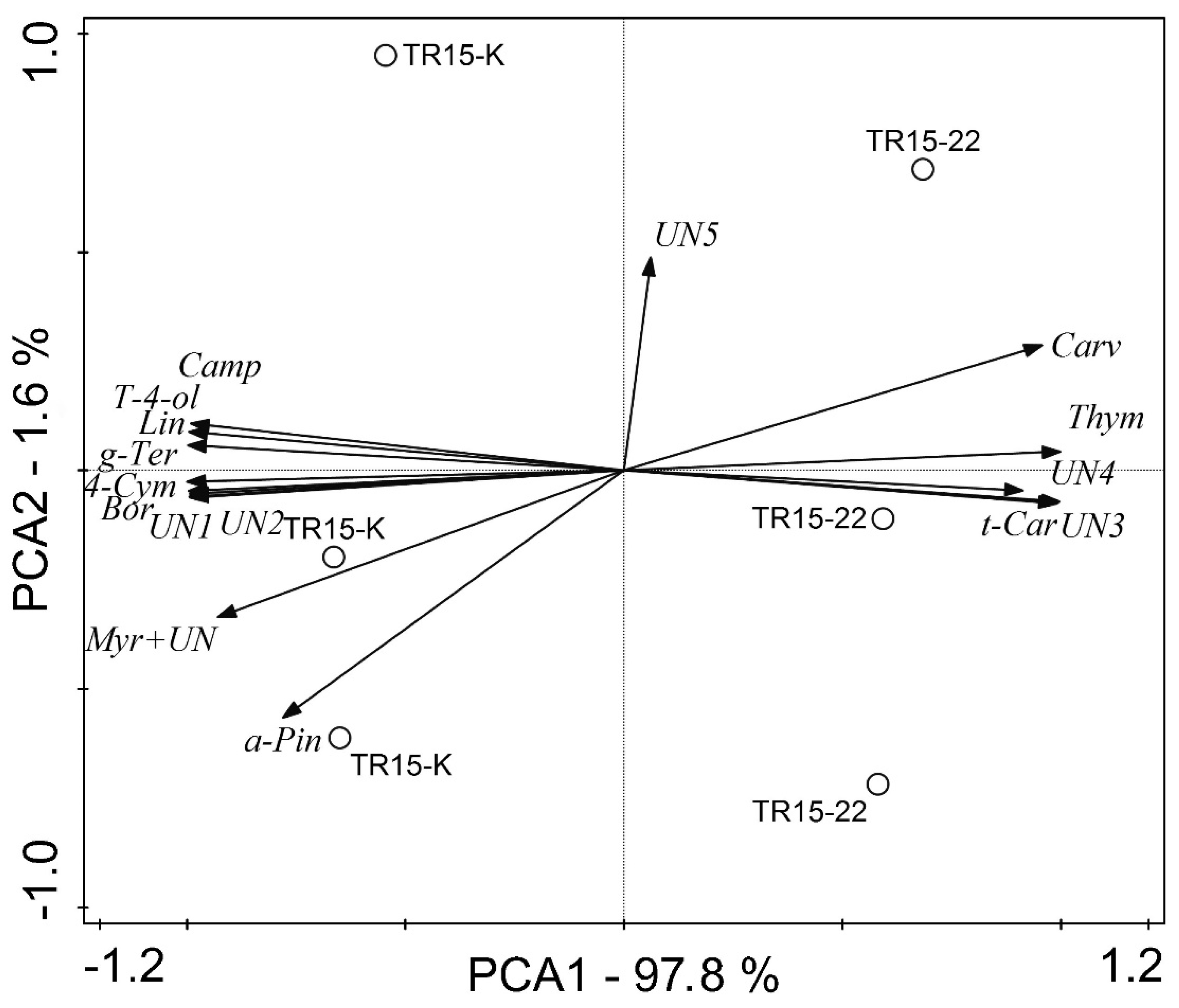
3.3. EO Chemical Composition
3.4. Acute Toxicity on C. quinquefasciatus and S. littoralis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nickavar, B.; Mojab, F.; Dolat-Abadi, R. Analysis of the essential oils of two Thymus species from Iran. Food Chem. 2005, 90, 609–611. [Google Scholar] [CrossRef]
- Rota, M.C.; Herrera, A.; Martínez, R.M.; Sotomayor, J.A.; Jordán, M.J. Antimicrobial activity and chemical composition of Thymus vulgaris, Thymus zygis and Thymus hyemalis essential oils. Food Control. 2008, 19, 681–687. [Google Scholar] [CrossRef]
- Satyal, P.; Murray, B.L.; McFeeters, R.L.; Setzer, W.N. Essential Oil Characterization of Thymus vulgaris from Various Geographical Locations. Foods 2016, 5, 70. [Google Scholar] [CrossRef]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.-N.; Tang, G.-Y.; Li, H.-B. Antibacterial and Antifungal Activities of Spices. Int. J. Mol. Sci. 2017, 18, 1283. [Google Scholar] [CrossRef]
- Mancini, E.; Senatore, F.; Del Monte, D.; De Martino, L.; Grulova, D.; Scognamiglio, M.; Snoussi, M.; De Feo, V. Studies on Chemical Composition, Antimicrobial and Antioxidant Activities of Five Thymus vulgaris L. Essential Oils. Molecules 2015, 20, 12016–12028. [Google Scholar] [CrossRef]
- Aicha, N.; Rachida, T.C.; Abdelmalek, E.L.M. Micropropagation of Thymus satureoides Coss. an endangered medicinal plant of Morocco. J. Agric. Tech. 2013, 9, 487–501. [Google Scholar]
- Rout, G.; Samantaray, S.; Das, P. In vitro manipulation and propagation of medicinal plants. Biotechnol. Adv. 2000, 18, 91–120. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; González-Benito, M.E.; Romano, A. Establishment of an in vitro propagation protocol for Thymus lotocephalus, a rare aromatic species of the Algarve (Portugal). Plant Growth Regul. 2011, 66, 69–74. [Google Scholar] [CrossRef]
- Affonso, V.R.; Bizzo, H.R.; Lage, C.L.S.; Sato, A. Influence of Growth Regulators in Biomass Production and Volatile Profile of in Vitro Plantlets of Thymus vulgaris L. J. Agric. Food Chem. 2009, 57, 6392–6395. [Google Scholar] [CrossRef] [PubMed]
- Ozudogru, E.A.; Kaya, E.; Kirdok, E.; Issever-Ozturk, S. In vitro propagation from young and mature explants of thyme (Thymus vulgaris and T. longicaulis) resulting in genetically stable shoots. Vitr. Cell. Dev. Biol. Anim. 2011, 47, 309–320. [Google Scholar] [CrossRef]
- Furmanowa, M.; Furmanowa, M.; Olszowska, O.; Olszowska, O. Micropropagation of Thyme (Thymus vulgaris L.). In Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin, Germany, 1992; Volume 19, pp. 230–243. [Google Scholar]
- Tosca, A.; Pandolfi, R.; Citterio, S.; Fasoli, A.; Sgorbati, S. Determination by flow cytometry of the chromosome doubling capacity of colchicine and oryzalin in gynogenetic haploids of gerbera. Plant Cell Rep. 1995, 14, 455–458. [Google Scholar] [CrossRef]
- Van Duren, M.; Morpurgo, R.; Dolezel, J.; Afza, R. Induction and verification of autotetraploids in diploid banana (Musa acuminata) by in vitro techniques. Euphytica 1996, 88, 25–34. [Google Scholar] [CrossRef]
- De Carvalho, J.F.R.P.; Carvalho, C.R.D.P.D.; Otoni, W.C. In vitro induction of polyploidy in annatto (Bixa orellana). Plant Cell Tissue Organ Cult. (PCTOC) 2005, 80, 69–75. [Google Scholar] [CrossRef]
- Kermani, M.J.; Sarasan, V.; Roberts, A.V.; Yokoya, K.; Wentworth, J.; Sieber, V.K. Oryzalin-induced chromosome doubling in Rosa and its effect on plant morphology and pollen viability. Theor. Appl. Genet. 2003, 107, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Petolino, J.F.; Widholm, J.M. Efficient production of doubled haploid plants through colchicine treatment of anther-derived maize callus. Theor. Appl. Genet. 1989, 77, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.; Gertz, A.; Joersbo, M.; Andersen, S. Antimicrotubule herbicides for in vitro chromosome doubling in Beta vulgaris L. ovule culture. Euphytica 1998, 101, 231–237. [Google Scholar] [CrossRef]
- Tavan, M.; Mirjalili, M.H.; Karimzadeh, G. In vitro polyploidy induction: Changes in morphological, anatomical and phytochemical characteristics of Thymus persicus (Lamiaceae). Plant Cell Tissue Organ Cult. 2015, 122, 573–583. [Google Scholar] [CrossRef]
- Pavela, R.; Sedlák, P. Post-application temperature as a factor influencing the insecticidal activity of essential oil from Thymus vulgaris. Ind. Crop. Prod. 2018, 113, 46–49. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Greplová, M.; Polzerová, H.; Domkářová, J. Intra- and inter-specific crosses of Solanum materials after mitotic polyploidization in vitro. Plant Breeding 2009, 128, 651–657. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef]
- Pavela, R. Acute, synergistic and antagonistic effects of some aromatic compounds on the Spodoptera littoralis Boisd. (Lep., Noctuidae) larvae. Ind. Crop. Prod. 2014, 60, 247–258. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Majdi, M.; Karimzadeh, G.; Malboobi, M.A.; Omidbaigi, R.; Mirzaghaderi, G. Induction of Tetraploidy to Feverfew (Tanacetum parthenium Schulz-Bip.): Morphological, Physiological, Cytological, and Phytochemical Changes. HortScience 2010, 45, 16–21. [Google Scholar] [CrossRef]
- Viehmannová, I.; Trávníčková, M.; Špatenková, E.; Cerna, M.; Trávníček, P. Induced polyploidization and its influence on yield, morphological, and qualitative characteristics of microtubers in Ullucus tuberosus. Plant Cell Tissue Organ Cult. 2011, 109, 83–90. [Google Scholar] [CrossRef]
- Dhooghe, E.; Van Laere, K.; Eeckhaut, T.; Leus, L.; Van Huylenbroeck, J. Mitotic chromosome doubling of plant tissues in vitro. Plant Cell Tissue Organ Cult. 2011, 104, 359–373. [Google Scholar] [CrossRef]
- Dhooghe, E.; Grunewald, W.; Leus, L.; Van Labeke, M.-C. In vitro polyploidisation of Helleborus species. Euphytica 2009, 165, 89–95. [Google Scholar] [CrossRef]
- Gomes, S.S.L.; Saldanha, C.W.; Neves, C.S.; Trevizani, M.; Raposo, N.R.B.; Notini, M.M.; Santos, M.D.O.; Campos, J.M.S.; Otoni, W.C.; Viccini, L.F. Karyotype, genome size, and in vitro chromosome doubling of Pfaffia glomerata (Spreng.) Pedersen. Plant Cell Tissue Organ Cult. 2014, 118, 45–56. [Google Scholar] [CrossRef]
- Shmeit, Y.H.; Fernandez, E.; Novy, P.; Kloucek, P.; Orosz, M.; Kokoska, L. Autopolyploidy effect on morphological variation and essential oil content in Thymus vulgaris L. Sci. Hortic. 2020, 263, 109095. [Google Scholar] [CrossRef]
- Stanys, V.; Weckman, A.; Staniene, G.; Duchovskis, P. In vitro induction of polyploidy in japanesse quince (Chaenomeles japonica). Plant Cell Tiss. Organ Cult. 2006, 84, 263–268. [Google Scholar] [CrossRef]
- Quesenberry, K.H.; Dampier, J.M.; Lee, Y.Y.; Smith, R.L.; Acuǹa, C.A. Doubling the chromosome number of bahiagrass via tissue culture. Euphytica 2010, 175, 43–50. [Google Scholar] [CrossRef]
- Švécarová, M.; Navrátilová, B.; Ondřej, V. In vitro polyploidization of Ajuga reptans L. using oryzalin. Acta Biol. Crac. Ser. Bot. 2018, 60, 69–73. [Google Scholar]
- Švécarová, M.; Navrátilová, B.; Hašler, P.; Ondřej, V. Artificial induction of tetraploidy in Humulus lupulus L. using oryzalin. Acta Agrobot. 2019, 72, 1764. [Google Scholar] [CrossRef]
- Salma, U.; Kundu, S.; Mandal, N. Artificial polyploidy in medicinal plants: Advancement in the last two decades and impending prospects. J. Crop. Sci. Biotechnol. 2017, 20, 9–19. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Canale, A.; Cianfaglione, K.; Ciaschetti, G.; Conti, F.; Nicoletti, M.; Senthil-Nathan, S.; Mehlhorn, H.; Maggi, F. Acute larvicidal toxicity of five essential oils (Pinus nigra, Hyssopus officinalis, Satureja montana, Aloysia citrodora and Pelargonium graveolens) against the filariasis vector Culex quinquefasciatus: Synergistic and antagonistic effects. Parasitol. Int. 2017, 66, 166–171. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivation of Nodal Segments on Oryzalin Medium for 2 Weeks | ||||||
|---|---|---|---|---|---|---|
| Oryzalin (mg L−1) | No. of Explants | Viable Explants (%) | No. of Tetraploids (%) | |||
| Clone 8 | Clone 15 | Clone 8 | Clone 15 | Clone 8 | Clone 15 | |
| 0.346 | 45 | 28 | 19 (42.2) | 15 (53.6) | 3 (15.8) | 6 (40.0) |
| 1.73 | 50 | 29 | 9 (18.0) | 3 (10.3) | 4 (44.4) | 2 (66.7) |
| 3.46 | 48 | 29 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| total | 143 | 86 | 28 (19.6) | 18 (20.2) | 7 (25.0) | 8 (44.4) |
| control | 20 | 20 | 20 (100) | 20 (100) | 0 (0) | 0 (0) |
| Overflow of nodal segments with oryzalin solution for 24 h | ||||||
| 1.73 | 33 | * | 14 (42.2) | * | 1 (7.1) | * |
| 3.46 | 32 | * | 8 (28.1) | * | 2 (25.0) | * |
| 5.19 | 20 | 22 | 0 (0) | 1 (4.5) | 0 (0) | 0 (0) |
| 6.92 | 25 | 20 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| 8.65 | 20 | 21 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| total | 130 | 63 | 22 (16.9) | 1 (1.6) | 3 (13.6) | 0 (0) |
| control | 10 | 10 | 10 (100) | 10 (100) | 0 (0) | 0 (0) |
| Terpenes | TR15-K | TR15-22 | ANOVA | ||||
|---|---|---|---|---|---|---|---|
| Area [%] | RSD [%] | Area [%] | RSD [%] | DF | F | p | |
| UN1 | 1.74 | 5.19 | 1.49 | 5.41 | 1; 4 | 143.0 | 0.0003 |
| α-Pinene | 0.71 | 5.65 | 0.64 | 2.23 | 1; 4 | 4.8 | 0.0936 |
| Myrcene + UN | 1.56 | 4.34 | 1.36 | 3.76 | 1; 4 | 18.6 | 0.0138 |
| UN2 | 2.96 | 4.51 | 2.4 | 3.74 | 1; 4 | 148.6 | 0.0003 |
| 4-Cymene | 14.94 | 4.31 | 12.39 | 4.64 | 1; 4 | 162.8 | 0.0002 |
| γ-Terpinene | 20.66 | 5.17 | 17.78 | 4.96 | 1; 4 | 347.3 | <0.0001 |
| UN3 | 0.48 | 7.45 | 0.78 | 4.17 | 1; 4 | 224.7 | 0.0001 |
| UN4 | 0.08 | 3.29 | 0.09 | 7.29 | 1; 4 | 22.3 | 0.0091 |
| Linalool | 1.52 | 5.88 | 1.1 | 5.41 | 1; 4 | 9621.9 | <0.0001 |
| Camphor | 0.43 | 7.55 | 0.28 | 4.56 | 1; 4 | 619.85 | <0.0001 |
| Borneol | 0.32 | 4.5 | 0.21 | 6.18 | 1; 4 | 124.7 | 0.0004 |
| Terpinen-4-ol | 0.66 | 6.58 | 0.31 | 5.04 | 1; 4 | 2231.4 | <0.0001 |
| UN5 | 0.13 | 6.6 | 0.13 | 15.14 | 1; 4 | 0.0 | 0.9308 |
| Thymol | 52.34 | 6.93 | 58.95 | 6.24 | 1; 4 | 240.3 | 0.0001 |
| Carvacrol | 0.95 | 10.94 | 1.2 | 9.73 | 1; 4 | 25.7 | 0.0071 |
| trans-Caryophyllene | 0.52 | 6.6 | 0.88 | 5.56 | 1; 4 | 22345.4 | <0.0001 |
| TR15-K | TR15-22 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Insect | LC50 | CI95 | LC90 | CI95 | Chi | LD50 | CI95 | LD90 | CI95 | Chi | ||
| Spodoptera littoralis | µg.larvae−1 | 45.2 | 41.1–50.8 | 61.7 | 57.8–72.1 | 3.532 | 36.3 | 32.2–39.1 | 52.5 | 48.5–55.3 | 1.263 | |
| Culex quinquefqsciatus | µg L−1 | 28.9 | 26.5–33.8 | 45.5 | 41.1–47.5 | 2.253 | 25.5 | 22.3–28.7 | 39.2 | 36.8–43.5 | 0.711 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navrátilová, B.; Švécarová, M.; Bednář, J.; Ondřej, V. In Vitro Polyploidization of Thymus vulgaris L. and Its Effect on Composition of Essential Oils. Agronomy 2021, 11, 596. https://doi.org/10.3390/agronomy11030596
Navrátilová B, Švécarová M, Bednář J, Ondřej V. In Vitro Polyploidization of Thymus vulgaris L. and Its Effect on Composition of Essential Oils. Agronomy. 2021; 11(3):596. https://doi.org/10.3390/agronomy11030596
Chicago/Turabian StyleNavrátilová, Božena, Michaela Švécarová, Jan Bednář, and Vladan Ondřej. 2021. "In Vitro Polyploidization of Thymus vulgaris L. and Its Effect on Composition of Essential Oils" Agronomy 11, no. 3: 596. https://doi.org/10.3390/agronomy11030596
APA StyleNavrátilová, B., Švécarová, M., Bednář, J., & Ondřej, V. (2021). In Vitro Polyploidization of Thymus vulgaris L. and Its Effect on Composition of Essential Oils. Agronomy, 11(3), 596. https://doi.org/10.3390/agronomy11030596