
Semiochemical-Based Attractant for the Ambrosia Pinhole Borer Euplatypus parallelus




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Source of Chemicals
2.2. Source of Beetles
2.3. Collection of Beetle-Produced Volatile Compounds
2.4. Identification of Beetle-Produced Volatile Compounds
2.5. Emission of the Major Male-Specific Volatile Compound Over Time
2.6. Field Bioassay of Synthetic Male-Specific Volatile Compounds
2.7. Statistical Analysis
3. Results
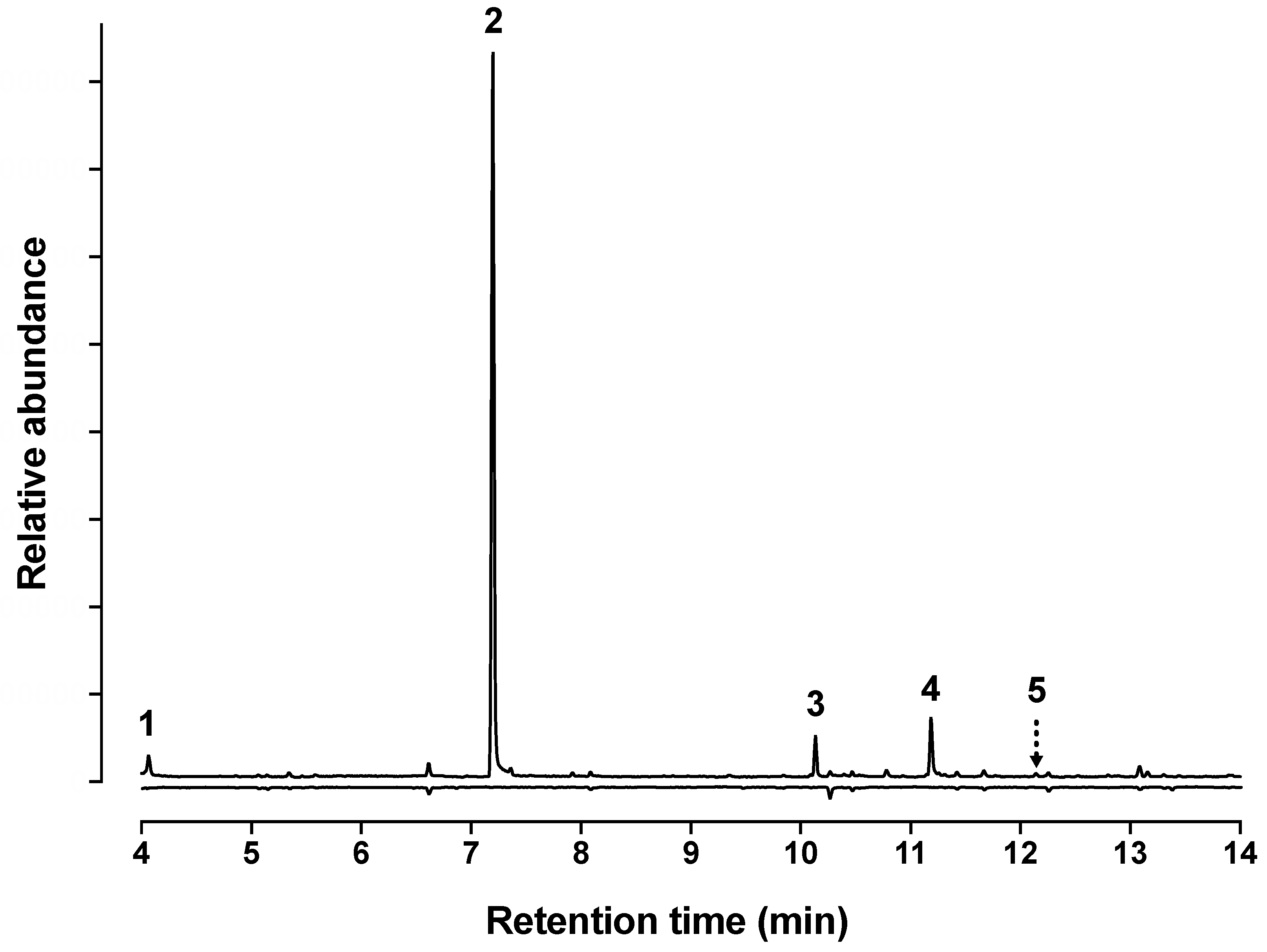
3.1. Identification of Beetle-Produced Volatile Compounds
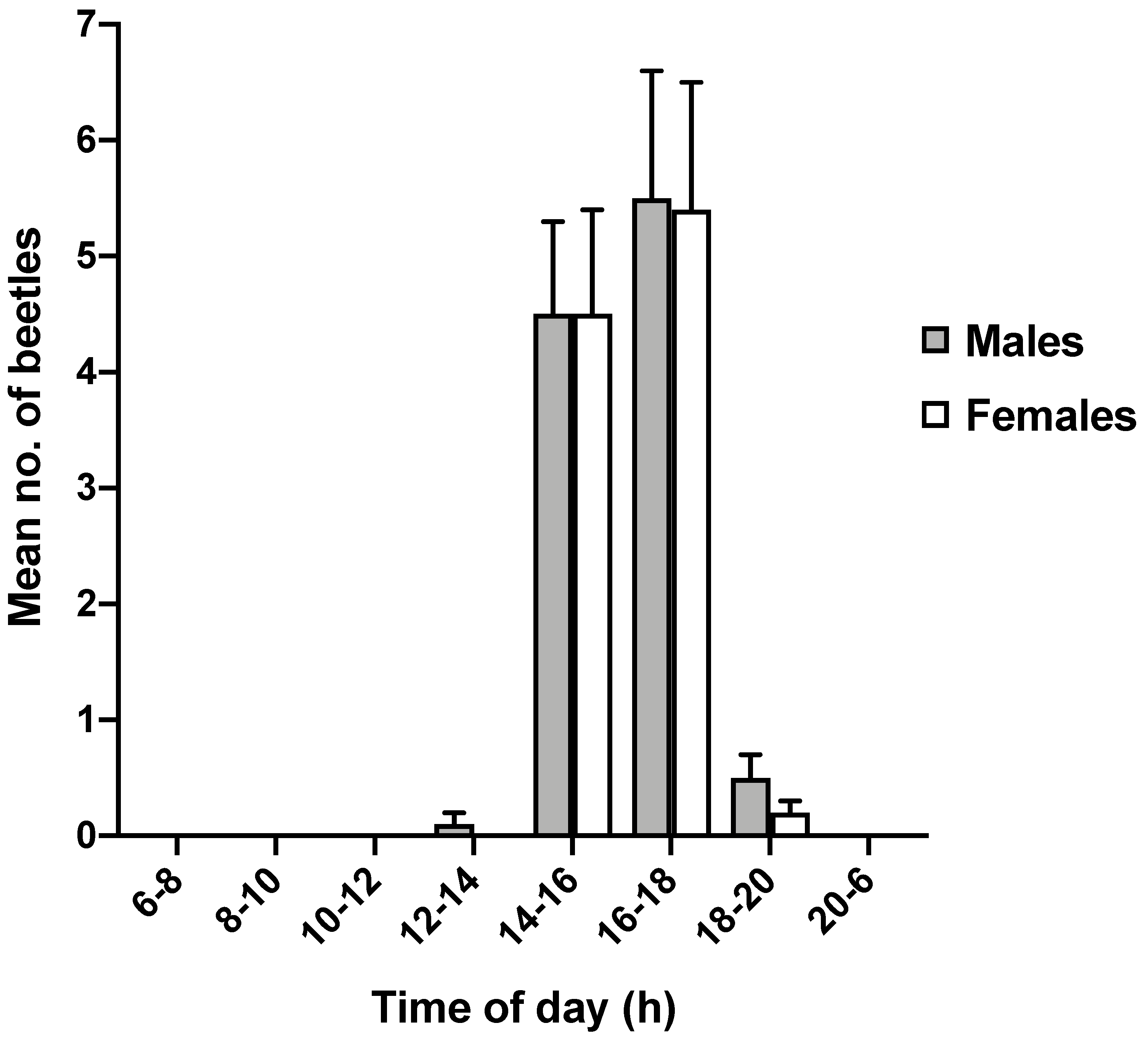
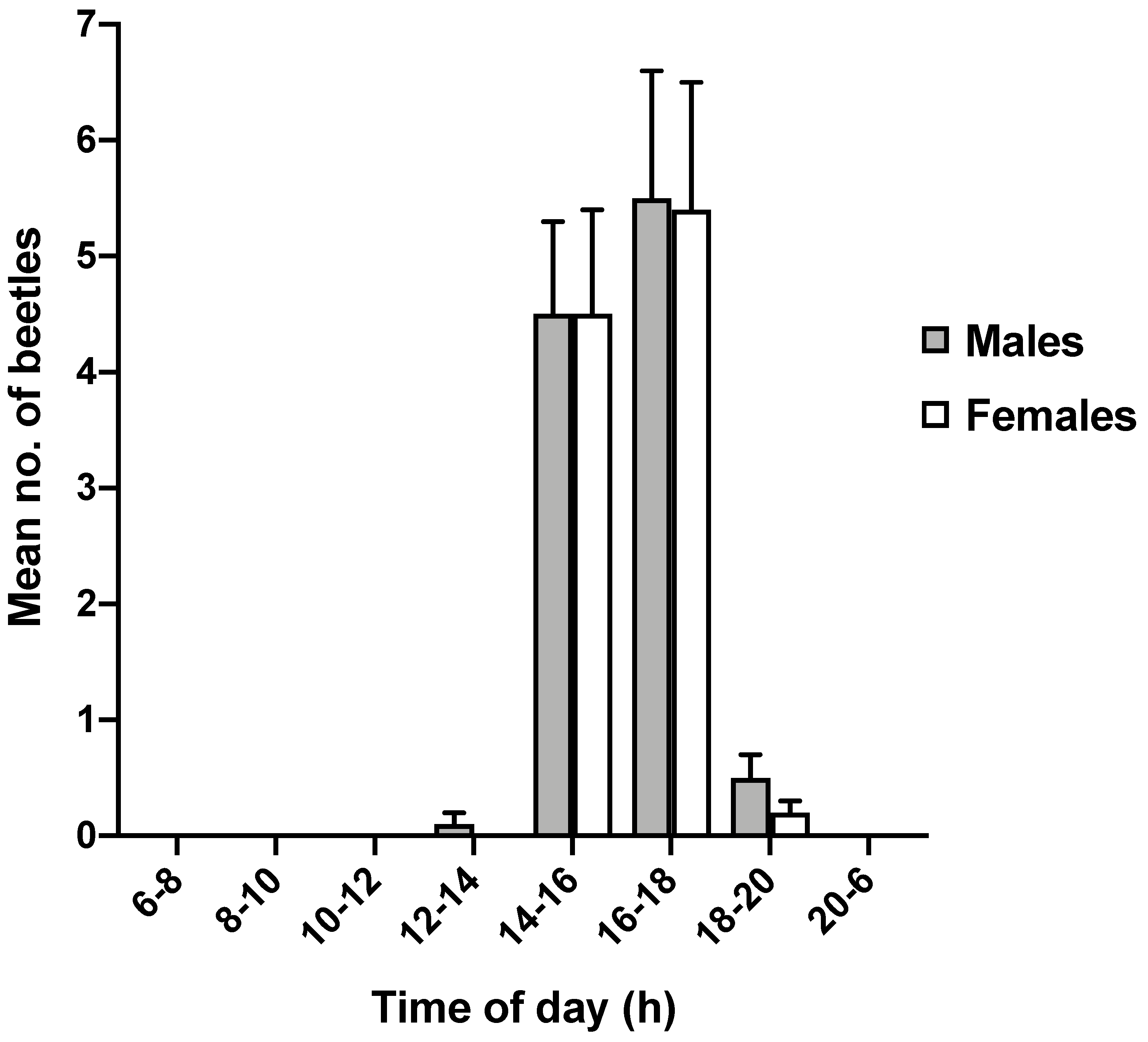
3.2. Circadian Rhythm of Adult Emergence
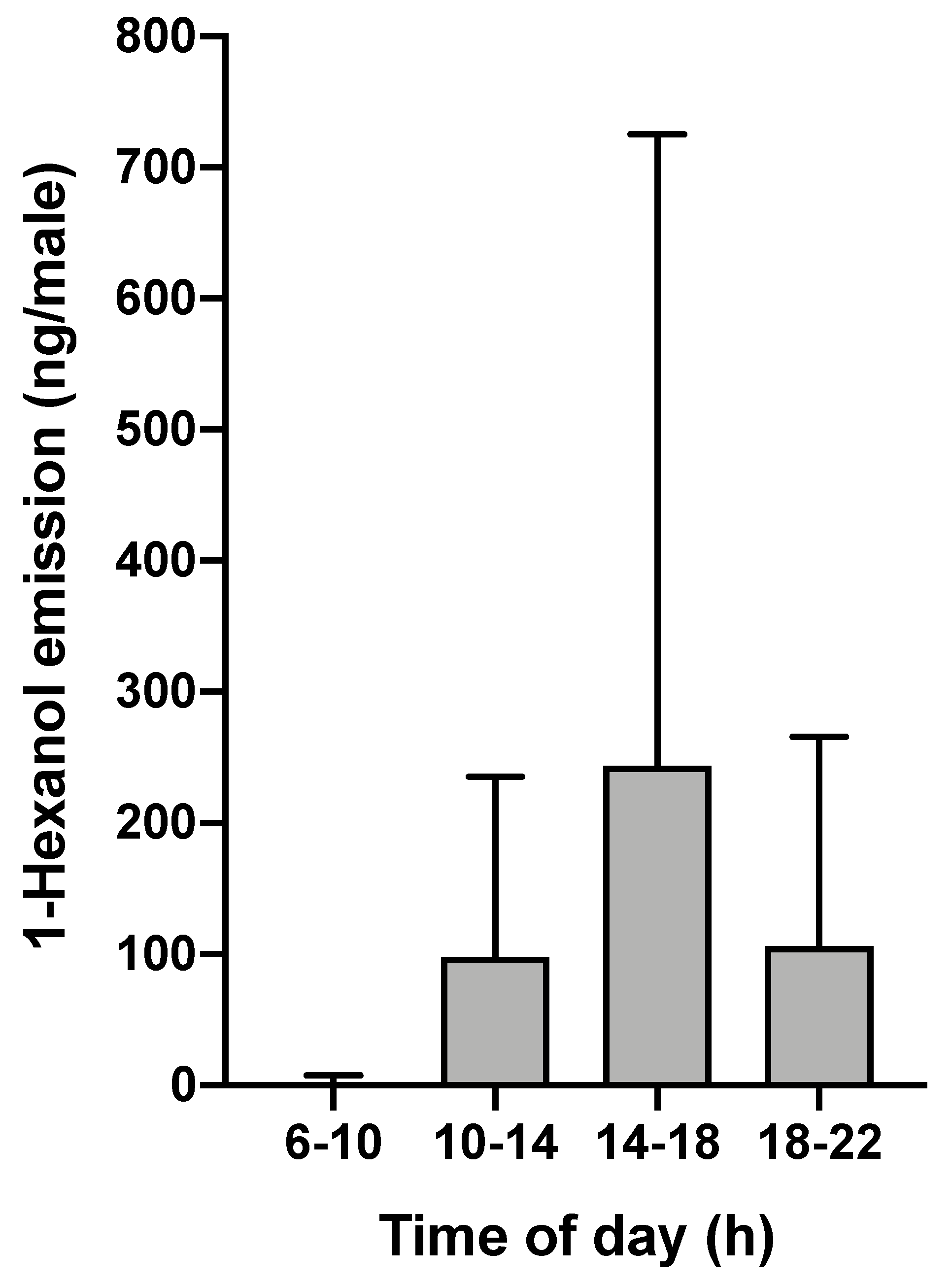
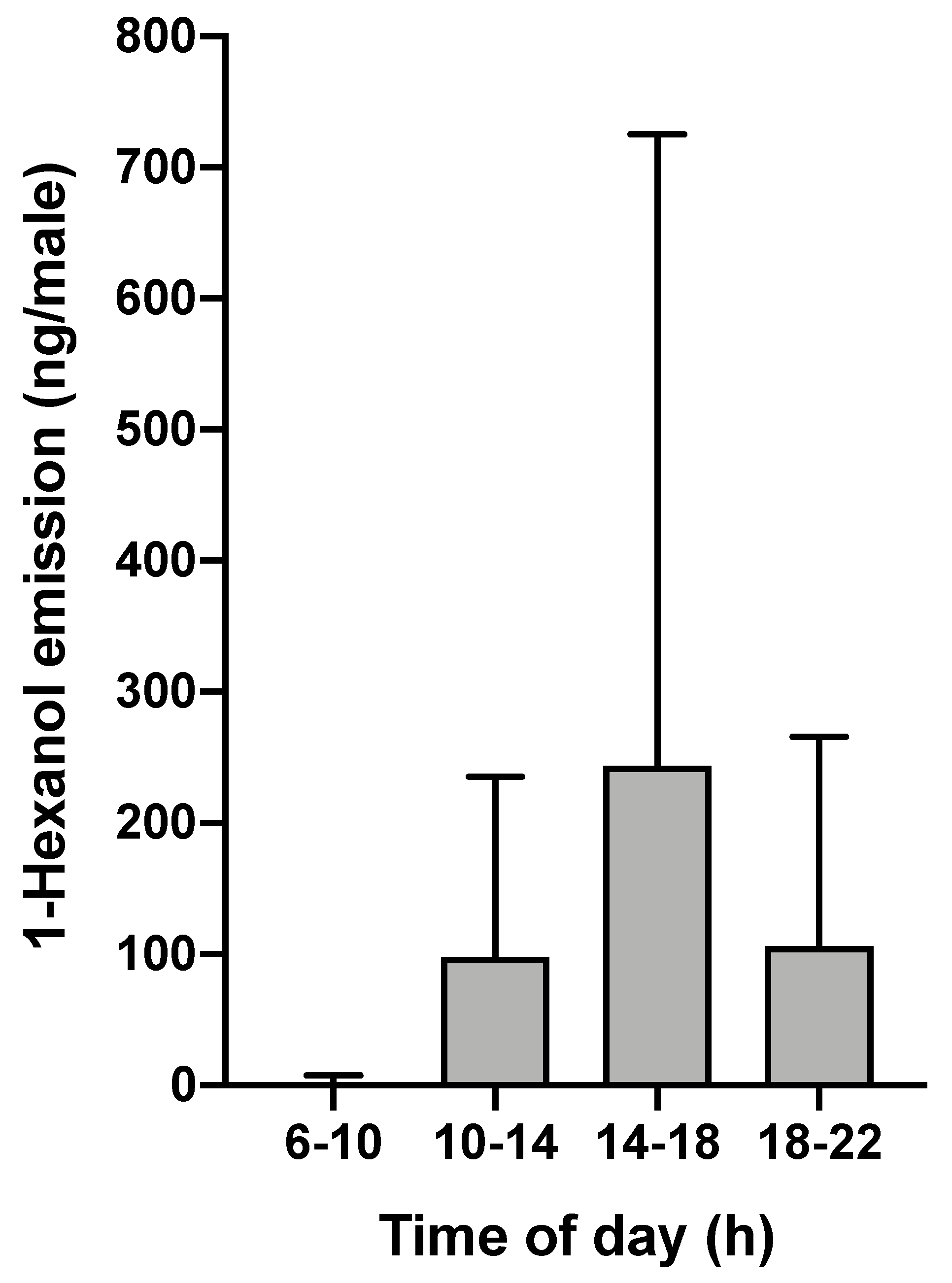
3.3. Emission of the Major Male-Specific Volatile Compound Over Time
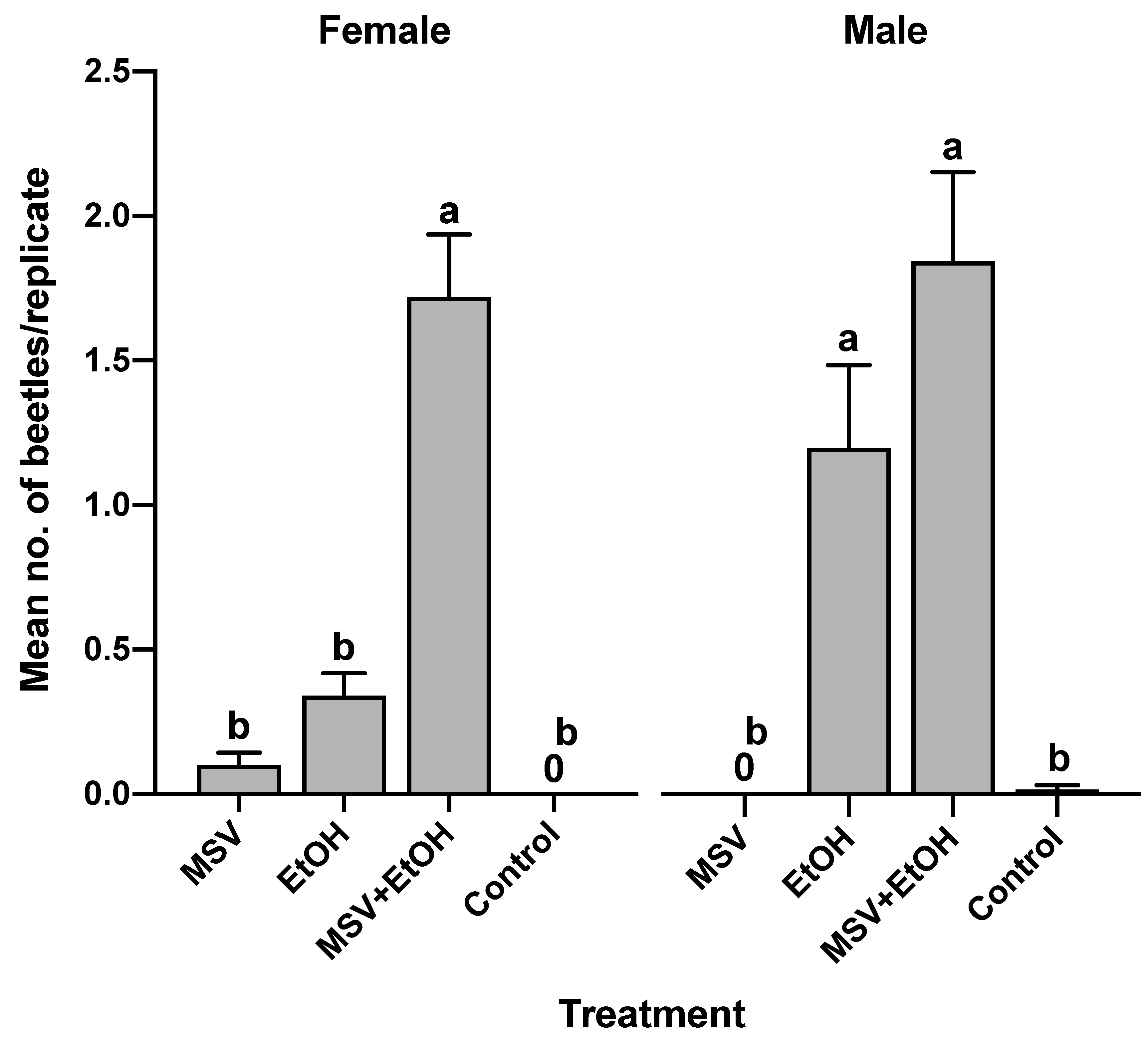
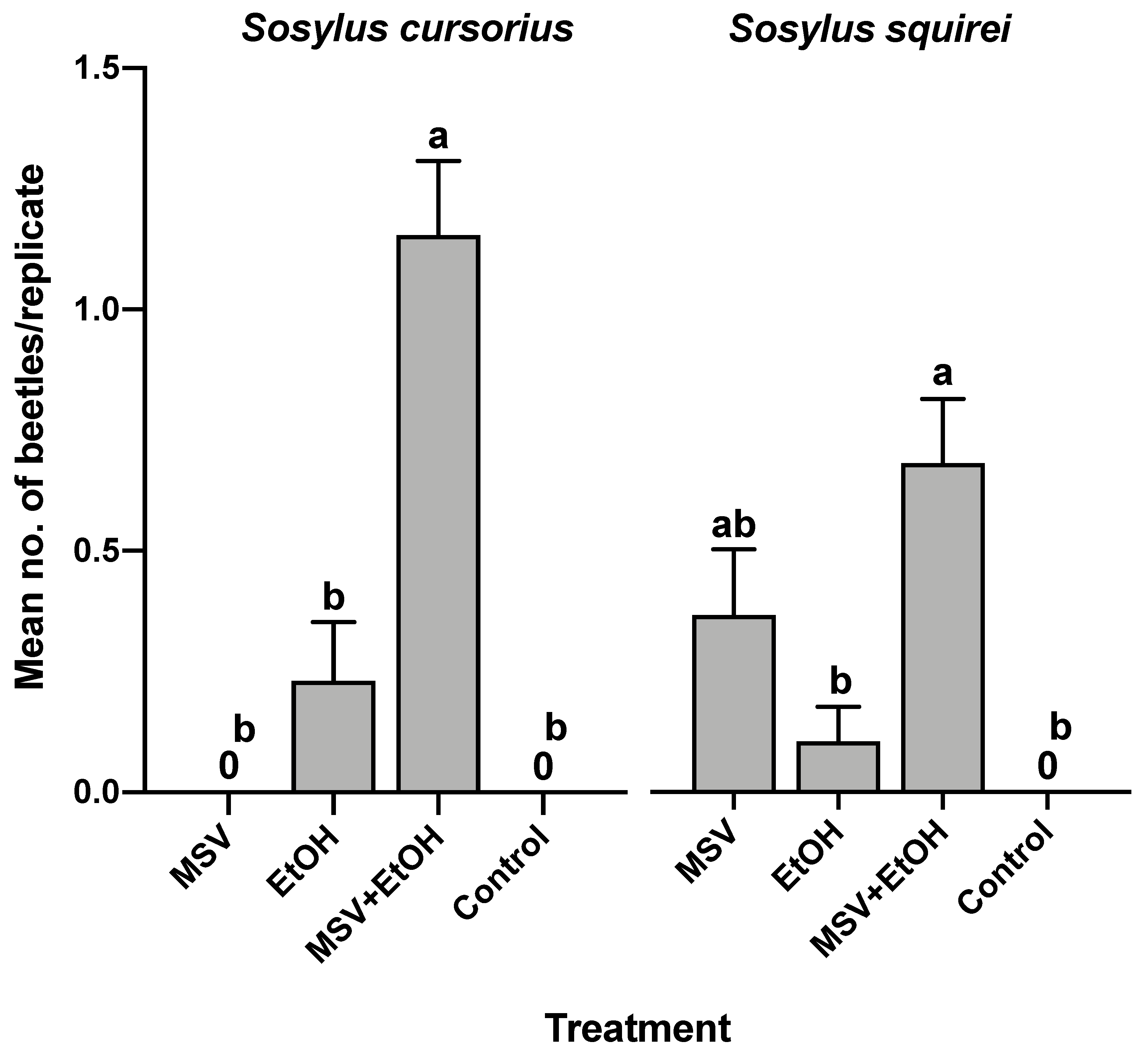
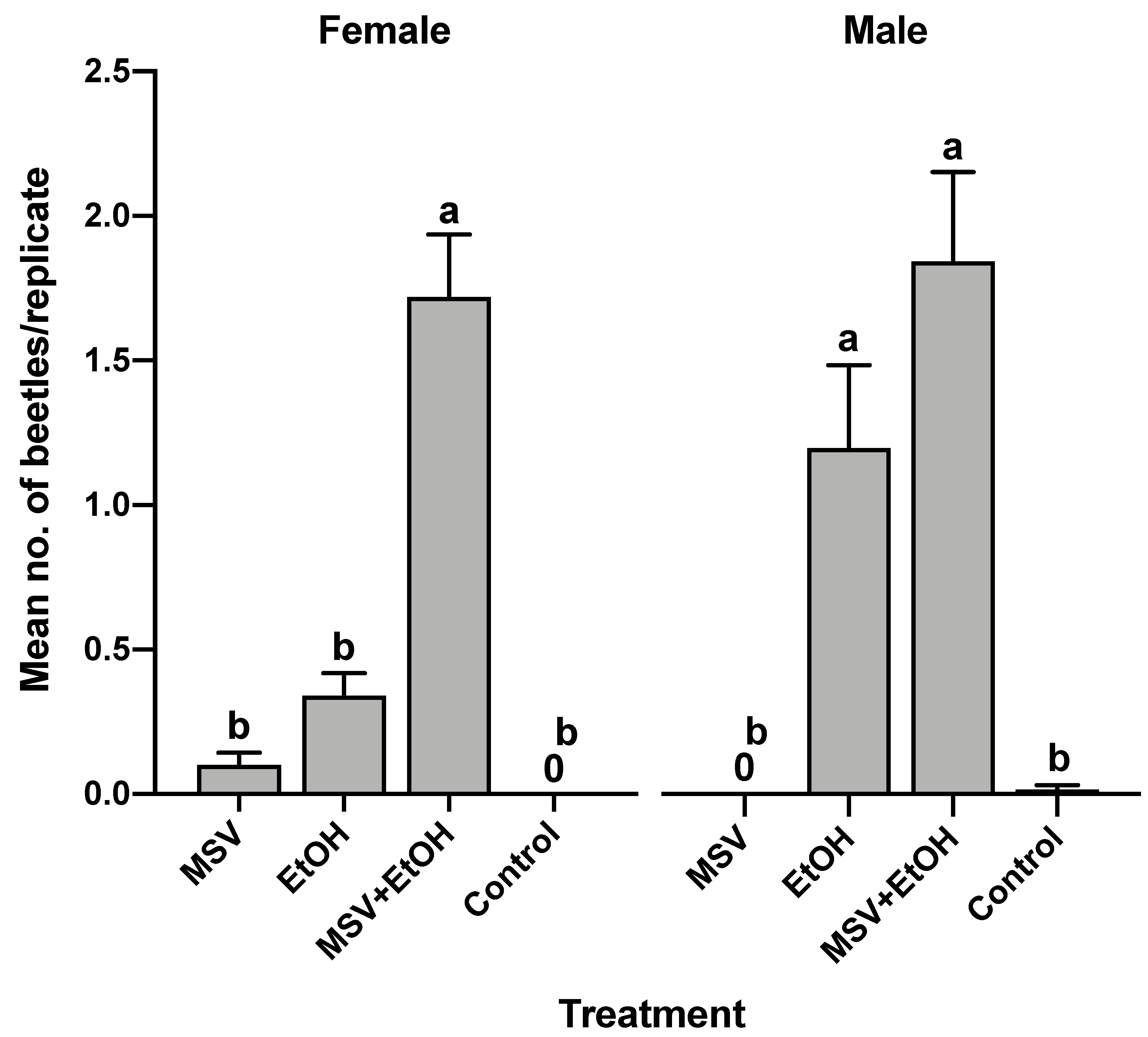
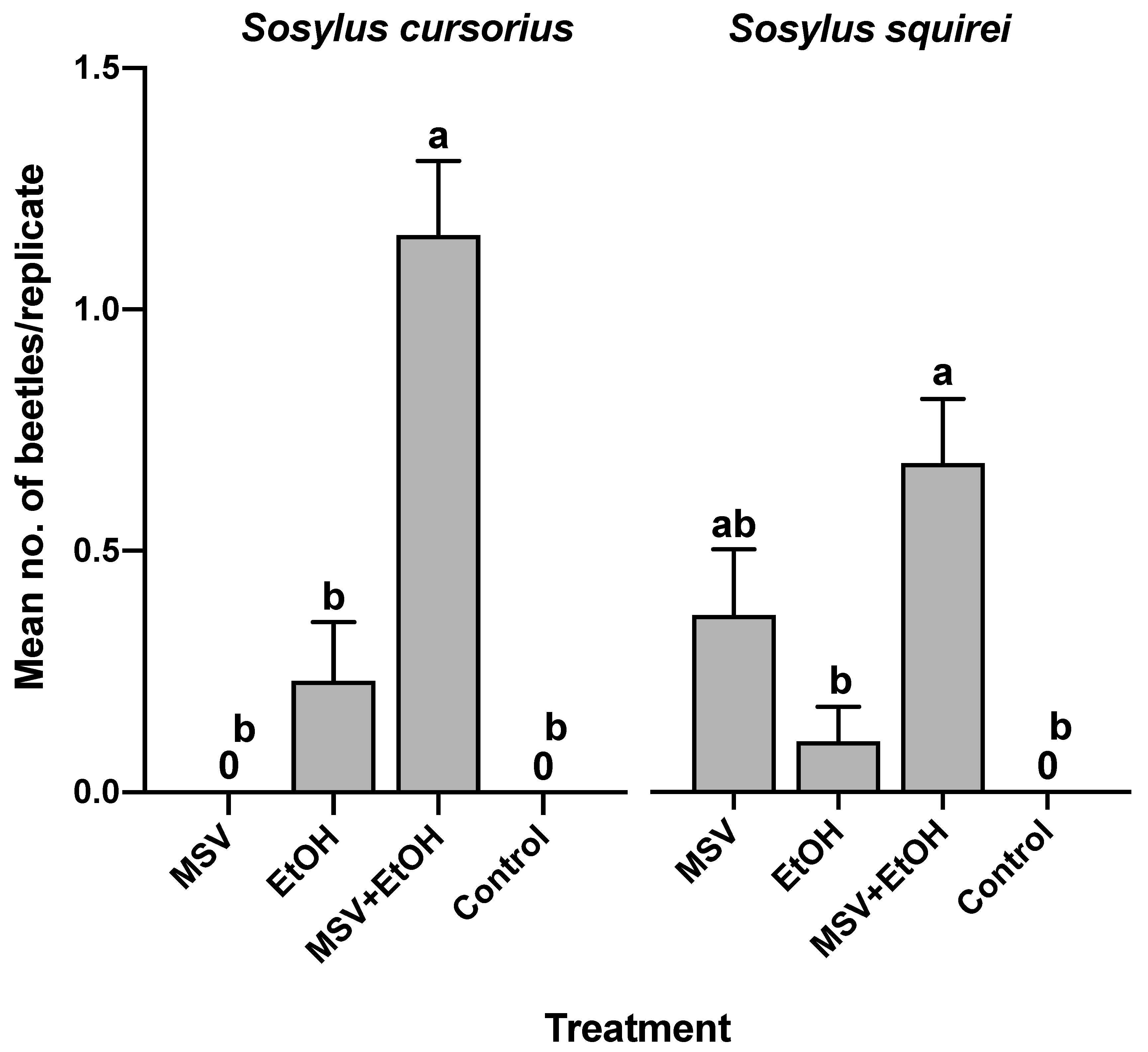
3.4. Field Bioassay of Synthetic Male-Specific Volatile Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wood, S.L.; Bright, D.R. A Catalog of Scolytidae and Platypodidae (Coleoptera), Part 2: Taxonomic Index Volume B; Brigham Young University: Provo, UT, USA, 1992; Volume 13, pp. 1084–1241. [Google Scholar]
- Beaver, R.A. The invasive neotropical ambrosia beetle Euplatypus parallelus (Fabricius, 1801) in the oriental region and its pest status (Coleoptera: Curculionidae, Platypodinae). Entomol. Mon. Mag. 2013, 149, 143–154. [Google Scholar]
- Gümüs, E.M.; Ergün, A. Report of a pest risk analysis for Platypus parallelus (Fabricius, 1801) for Turkey. EPPO Bull. 2015, 45, 112–118. [Google Scholar] [CrossRef]
- Schedl, K.E. Scolytidae und Platypodidae Afrikas III, Familie Platypodidae. Revista Entomol. Moçamb. 1962, 5, 595–1352. [Google Scholar]
- Kirkendall, L.R.; Biedermann, P.H.W.; Jordal, B.H. Evolution and diversity of bark and ambrosia beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, UK, 2015; pp. 85–156. [Google Scholar]
- Ribeiro, A.; Ferraz Filho, A.C.; Oliveira, E.B. Usos, importância econômica e perspectivas de mercado. In Mogno-Africano (Khaya spp.): Atualidades e Perspectivas do Cultivo no Brasil; Reis, C.A.F., Oliveira, E.B., Santos, A.M., Eds.; Embrapa: Brasília, Brazil, 2019; pp. 50–73. Available online: http://ainfo.cnptia.embrapa.br/digital/bitstream/item/202696/1/Mogno-Africano-08-10-2019.pdf (accessed on 19 January 2021).
- Sanderson, F.R.; Fong, Y.K.; Pheng, Y.C.; Ong, K.H.; Anuar, S. A fusarium wilt (Fusarium oxysporum) of angsana (Pterocarpus indicus) in Singapore. I, Epidemiology and identification of the causal organism. Arboric. J. 1997, 21, 187–204. [Google Scholar] [CrossRef]
- Philip, E. Wilt disease of angsana (Pterocarpus indicus) in Peninsular Malaysia and its possible control. J. Trop. For. Sci. 1999, 11, 519–527. Available online: https://www.jstor.org/stable/43582560 (accessed on 19 January 2021).
- Kirkendall, L.R.; Islam, M.R. Widespread wilting of Dalbergia sissoo in Bangladesh: The role of timber borers. In Mortality of Sissoo (Dalbergia sissoo) and Top Dying of Sundri (Heritiera Fomes) in Bangladesh; Baksha, M.W., Ed.; Bangladesh Forest Research Institute: Chittagong, Bangladesh, 2003; pp. 34–38. [Google Scholar]
- Zanuncio, J.C.; Sossai, M.F.; Couto, L.; Pinto, R. Occurrence of Euplatypus parallelus, Euplatypus sp. (Col.: Euplatypodidae) and Xyleborus affinis (Col.: Scolytidae) in Pinus sp. in Ribas do Rio Pardo, Mato Grosso do Sul, Brazil. Rev. Árvore 2002, 26, 387–389. [Google Scholar] [CrossRef]
- Zanuncio, J.C.; Sossai, M.F.; Flechtmann, C.A.H.; Zanuncio, T.V.; Guimarães, E.M.; Espindula, M.C. Plants of an Eucalyptus clone damaged by Scolytidae and Platypodidae (Coleoptera). Pesqui. Agropecuária Bras. 2005, 40, 513–515. [Google Scholar] [CrossRef]
- Bumrungsri, S.; Beaver, R.A.; Phongpaichit, S.; Sittichaya, W. The infestation by an exotic ambrosia beetle, Euplatypus parallelus (F.) (Coleoptera: Curculionidae: Platypodinae) of Angsana trees (Pterocarpus indicus Willd.) in southern Thailand. Songklanakarin J. Sci. Technol. 2008, 30, 579–582. Available online: https://rdo.psu.ac.th/sjstweb/journal/30-5/0125-3395-30-5-579-582.pdf (accessed on 19 January 2021).
- Maruthadurai, R.; Desai, A.R.; Singh, N.P. First record of ambrosia beetle (Euplatypus parallelus) infestation on cashew from Goa, India. Phytoparasitica 2014, 42, 57–59. [Google Scholar] [CrossRef]
- Hiremath, S.R.; Prathapan, K.D. First report of the invasive South American pinhole borer, Euplatypus parallelus (Fabricius) (Coleoptera: Curculionidae: Platypodinae), on rubber in India. Coleopt. Bull. 2019, 73, 714–717. [Google Scholar] [CrossRef]
- Rainho, H.L.; Flechtmann, C.A.H.; Silva, H.A.S.; Rocha, E.T.A. Ataque de Scolytinae e Platypodinae (Curculionidae) em seringueiras (Hevea brasiliensis) em São Paulo. In Proceedings of the 10th Congresso de Ecologia do Brasil, São Lourenço, Brazil, 16–22 September 2011; Abstract 1101. pp. 1–2. Available online: http://seb-ecologia.org.br/revistas/indexar/anais/xceb/resumos/1101.pdf (accessed on 19 January 2021).
- Elliott, H.J.; Madden, J.L.; Bashford, R. The association of ethanol in the attack behaviour of the mountain pinhole borer Platypus subgranosus Schedl (Coleoptera: Curculionidae: Platypodinae). J. Aus. Entomol. Soc. 1983, 22, 299–302. [Google Scholar] [CrossRef]
- Shore, T.L.; McLean, J.A. Attraction of Platypus wilsoni Swaine (Coleoptera: Platipodidae) to traps baited with sulcatol, ethanol and alpha-pinene. Can. For. Serv. Res. Notes 1983, 3, 24–25. [Google Scholar]
- Kimmerer, T.W.; Kozlowski, T.T. Ethylene, ethane, acetaldehyde, and ethanol production by plants under stress. Plant Physiol. 1982, 69, 840–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranger, C.M.; Reding, M.E.; Schultz, P.B.; Oliver, J.B. Influence of flood-stress on ambrosia beetle host-selection and implications for their management in a changing climate. Agric. For. Entomol. 2013, 15, 56–64. [Google Scholar] [CrossRef]
- Kelsey, R.G.; Gallego, D.; Sánchez-García, F.J.; Pajares, J.A. Ethanol accumulation during severe drought may signal tree vulnerability to detection and attack by bark beetles. Can. J. For. Res. 2014, 44, 554–561. [Google Scholar] [CrossRef]
- Klimetzek, D.; Köhler, J.; Vité, J.P.; Kohnle, U. Dosage response to ethanol mediates host selection by secondary bark beetles. Naturwissenschaften 1986, 73, 270–272. [Google Scholar] [CrossRef]
- Ranger, C.M.; Biedermann, P.H.W.; Phuntumart, V.; Beligala, G.U.; Ghosh, S.; Palmquist, D.E.; Mueller, R.; Barnett, J.; Schultz, P.B.; Reding, M.E.; et al. Symbiont selection via alcohol benefits fungus farming by ambrosia beetles. Proc. Natl. Acad. Sci. USA 2018, 115, 4447–4452. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Audino, P.; Villaverde, R.; Alfaro, R.; Zerba, E. Identification of volatile emissions from Platypus mutatus (=sulcatus) (Coleoptera: Platypodidae) and their behavioral activity. J. Econ. Entomol. 2005, 98, 1506–1509. [Google Scholar] [CrossRef]
- Gatti Liguori, P.; Zerba, E.; Alzogaray, R.A.; Gonzalez-Audino, P. 3-Pentanol: A new attractant present in volatile emissions from the ambrosia beetle, Megaplatypus mutatus. J. Chem. Ecol. 2008, 34, 1446–1451. [Google Scholar] [CrossRef]
- Renwick, J.A.A.; Vité, J.P.; Billings, R.F. Aggregation pheromones in the ambrosia beetle Platypus flavicornis. Naturwissenschaften 1977, 64, 226. [Google Scholar] [CrossRef]
- Algarvio, R.; Teixeira, C.; Barata, E.; Pickett, J.; Casas Novas, P.; Figueiredo, D. Identification of a putative aggregation pheromone from males Platypus cylindrus (Coleoptera: Platypodidae). In Proceedings of the 19th ISCE Annual Meeting, Hamburg, Germany, 3–7 August 2002; p. 152. Available online: https://www.chemecol.org/programs/2002.pdf (accessed on 19 January 2021).
- Kim, J.; Lee, S.G.; Shin, S.C.; Kwon, Y.D.; Park, I.K. Male-produced aggregation pheromone blend in Platypus koryoensis. J. Agric. Food Chem. 2009, 57, 1406–1412. [Google Scholar] [CrossRef] [PubMed]
- Tokoro, M.; Kobayashi, M.; Saito, S.; Kinuura, H.; Nakashima, T.; Shoda-Kagaya, E.; Kashiwagi, T.; Tebayashi, S.; Kim, C.; Mori, K. Novel aggregation pheromone, (1S,4R)-p-menth-2-en-1-ol, of the ambrosia beetle, Platypus quercivorus (Coleoptera: Platypodidae). Bull. FFPRI 2007, 6, 49–57. Available online: https://www.ffpri.affrc.go.jp/labs/kanko/402-5.pdf (accessed on 19 January 2021).
- Gerónimo-Torres, J.D.C.; Pérez-De La Cruz, M.; De La Cruz-Pérez, A.; Torres-De La Cruz, M. Scolytinae y Platypodinae (Coleoptera: Curculionidae) asociados a manglares de Tabasco, México. Rev. Colomb. Entomol. 2015, 41, 257–261. Available online: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-04882015000200018&lng=en&nrm=iso (accessed on 19 January 2021).
- Poland, T.M.; Rassati, D. Improved biosecurity surveillance of non-native forest insects: A review of current methods. J. Pest Sci. 2019, 92, 37–49. [Google Scholar] [CrossRef]
- Thube, S.H.; Mohan, C.; Pandian, R.T.P.; Saneera, E.K.; Sannagoudra, H.M.; Hedge, V.; Chowdappa, P. First record of the invasive Neotropical ambrosia beetle Euplatypus parallelus (Fabricius, 1801) (Coleoptera: Curculionidae: Platypodinae) infesting arecanut in Karnataka, India. Coleopt. Bull. 2018, 72, 713–716. [Google Scholar] [CrossRef]
- Gonzalez-Audino, P.; Gatti Liguori, P.; Zerba, E. Translucent pheromone traps increase trapping efficiency of ambrosia beetle Megaplatypus mutatus. Crop. Prot. 2011, 30, 745–747. [Google Scholar] [CrossRef]
- Funes, H.; Zerba, E.; Gonzalez-Audino, P. Comparison of three types of traps baited with sexual pheromones for ambrosia beetle Megaplatypus mutatus (Coleoptera: Platypodinae) in poplar plantations. J. Econ. Entomol. 2009, 102, 1546–1550. [Google Scholar] [CrossRef]
- Funes, H.; Zerba, E.; Gonzalez-Audino, P. Effect of release rate and enantiomeric composition response to pheromones of Megaplatypus mutatus (Chapuis) in poplar plantations of Argentina and Italy. Bull. Entomol. Res. 2013, 103, 564–569. [Google Scholar] [CrossRef]
- Gallego, D.; Galián, J.; Diez, J.J.; Pajares, J.A. Kairomonal responses of Tomicus destruens (Col., Scolytidae) to host volatiles α-pinene and ethanol. J. Appl. Entomol. 2008, 132, 654–662. [Google Scholar] [CrossRef]
- Wong, J.C.H.; Meier, L.R.; Zou, Y.; Mongold-Diers, J.A.; Hanks, L.M. Evaluation of methods used in testing attraction of cerambycid beetles to pheromone-baited traps. J. Econ. Entomol. 2017, 110, 2269–2274. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT V.9.3 User’s Guide; SAS Institute: Cary, NC, USA, 2011. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; W.H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar] [CrossRef] [Green Version]
- Newcombe, R.G. Two-sided confidence intervals for the single proportion: Comparison of seven methods. Stat. Med. 1998, 17, 857–872. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.A. Host-tree chemistry affecting colonization in bark beetles. In Chemical Ecology of Insects 2; Cardé, R.T., Bell, W.J., Eds.; Chapman & Hall: New York, NY, USA, 1995; pp. 154–213. [Google Scholar] [CrossRef] [Green Version]
- Graham, K. Anaerobic induction of primary chemical attractancy for ambrosia beetles. Can. J. Zool. 1968, 46, 905–908. [Google Scholar] [CrossRef]
- Moeck, H.A. Ethanol as the primary attractant for the ambrosia beetle Trypodendron lineatum (Coleoptera: Scolytidae). Can. Entomol. 1970, 102, 985–995. [Google Scholar] [CrossRef]
- Madrid, F.; Vité, J.P.; Renwick, J.A.A. Evidence of aggregation pheromones in the ambrosia beetle Platypus flavicornis (F.). Z. Angew. Entomol. 1972, 72, 73–79. [Google Scholar] [CrossRef]
- Nam, Y.; Choi, W.I. Diurnal flight pattern of Platypus koryoensis (Coleoptera: Platypodinae) in relation to abiotic factors in Korea. J. Asia Pac. Entomol. 2014, 17, 417–422. [Google Scholar] [CrossRef]
- Pascoe, F.P. List of the Colydiidae collected in the Amazons valley by H.W. Bates, Esq., and descriptions of new species by Francis P. Pascoe. J. Entomol. 1863, 2, 79–99. [Google Scholar]
- Slipinski, A.; Lord, N.P.; Lawrence, J.F. Bothrideridae Erichson, 1845. In Handbook of Zoology. Arthropoda: Insecta: Coleoptera. Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 411–422. [Google Scholar] [CrossRef]
- Browne, F.G. Sosylus spectabilis Grouvelle (Coleoptera, Colydiidae), a predator and parasite of African ambrosia beetles. In 5th Report West African Timber Borer Research Unit 1961–1962; Crown Agents: London, UK, 1962; pp. 91–96. [Google Scholar]
- Roberts, H. A note on the Nigerian species of the genus Sosylus Erichson (Col. Colydiidae), parasites and predators of ambrosia beetles. J. Nat. Hist. 1969, 3, 85–91. [Google Scholar] [CrossRef]
- Roberts, H. Description of the developmental stages of Sosylus spp. (Coleoptera: Colydiidae) from New Guinea, parasites and predators of ambrosia beetles (Coleoptera: Platypodidae). Bull. Entomol. Res. 1980, 70, 245–252. [Google Scholar] [CrossRef]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R.A. Common pathways by which non-native forest insects move internationally and domestically. J. Pest Sci. 2019, 92, 13–27. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rainho, H.L.; Silva, W.D.; Bento, J.M.S. Semiochemical-Based Attractant for the Ambrosia Pinhole Borer Euplatypus parallelus. Agronomy 2021, 11, 266. https://doi.org/10.3390/agronomy11020266
Rainho HL, Silva WD, Bento JMS. Semiochemical-Based Attractant for the Ambrosia Pinhole Borer Euplatypus parallelus. Agronomy. 2021; 11(2):266. https://doi.org/10.3390/agronomy11020266
Chicago/Turabian StyleRainho, Hugo L., Weliton D. Silva, and José Maurício S. Bento. 2021. "Semiochemical-Based Attractant for the Ambrosia Pinhole Borer Euplatypus parallelus" Agronomy 11, no. 2: 266. https://doi.org/10.3390/agronomy11020266
APA StyleRainho, H. L., Silva, W. D., & Bento, J. M. S. (2021). Semiochemical-Based Attractant for the Ambrosia Pinhole Borer Euplatypus parallelus. Agronomy, 11(2), 266. https://doi.org/10.3390/agronomy11020266