
Study of Forage Quality of Grasslands on the Southern Margin of the Pannonian Basin



Abstract
:1. Introduction
2. Materials and Methods
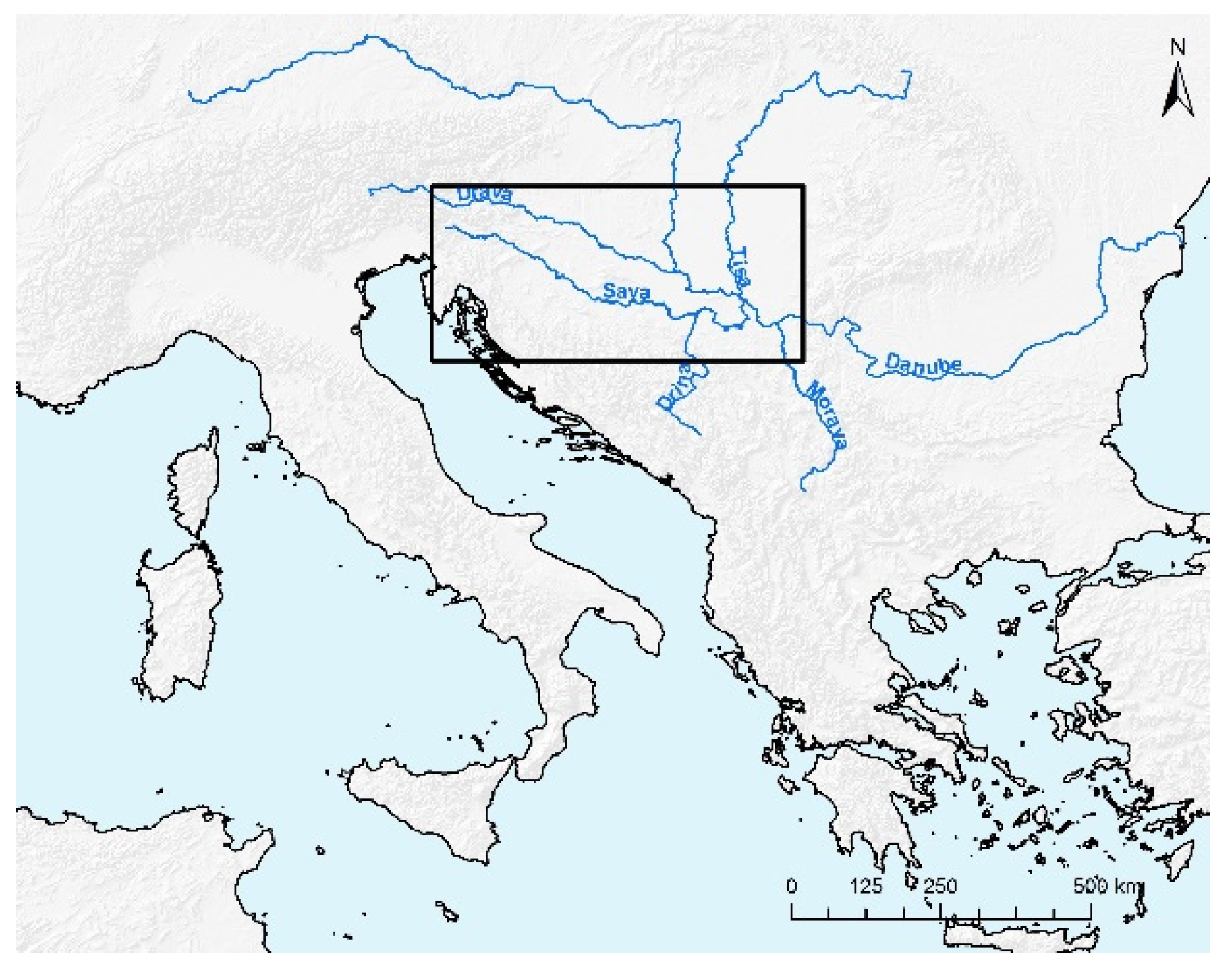
2.1. Study Area
2.2. Data
2.3. Nomenclature
2.4. Numerical Analysis
3. Results
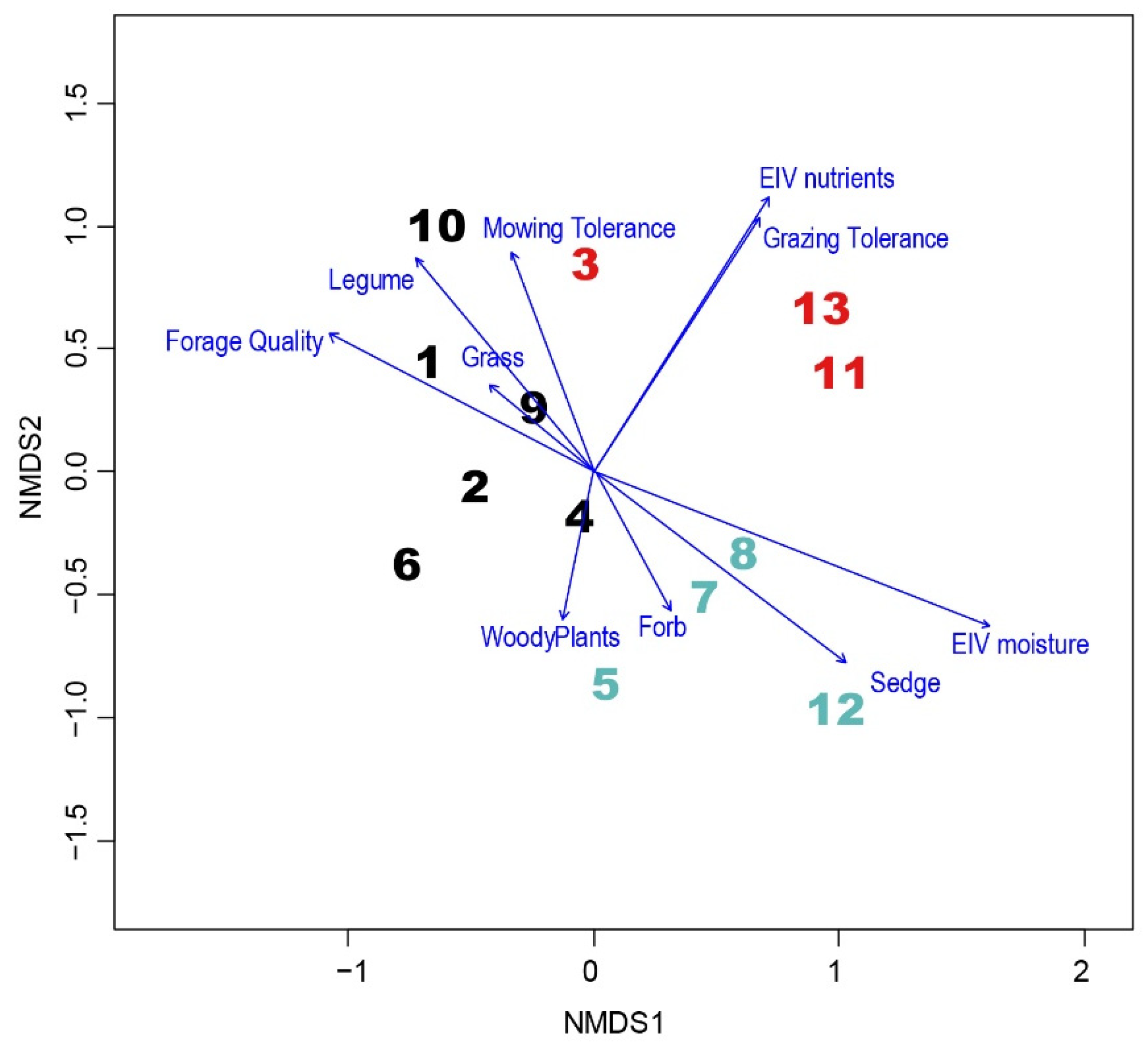
3.1. Ordination with Projected Growth Forms, Tolerance to Mowing, Grazing and EIV Moisture, Nutrients
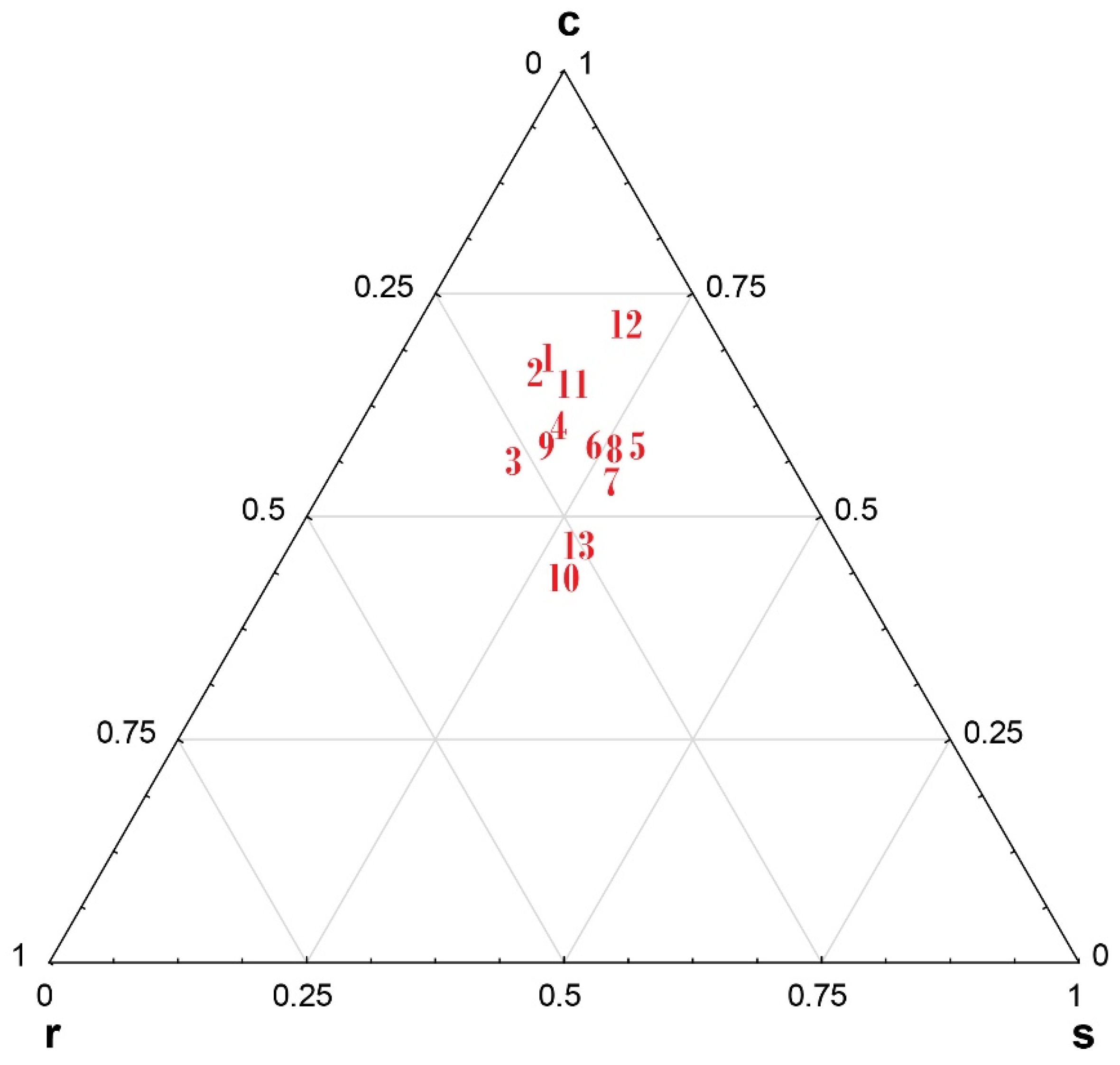
3.2. Ecological Strategies
3.3. Correlations
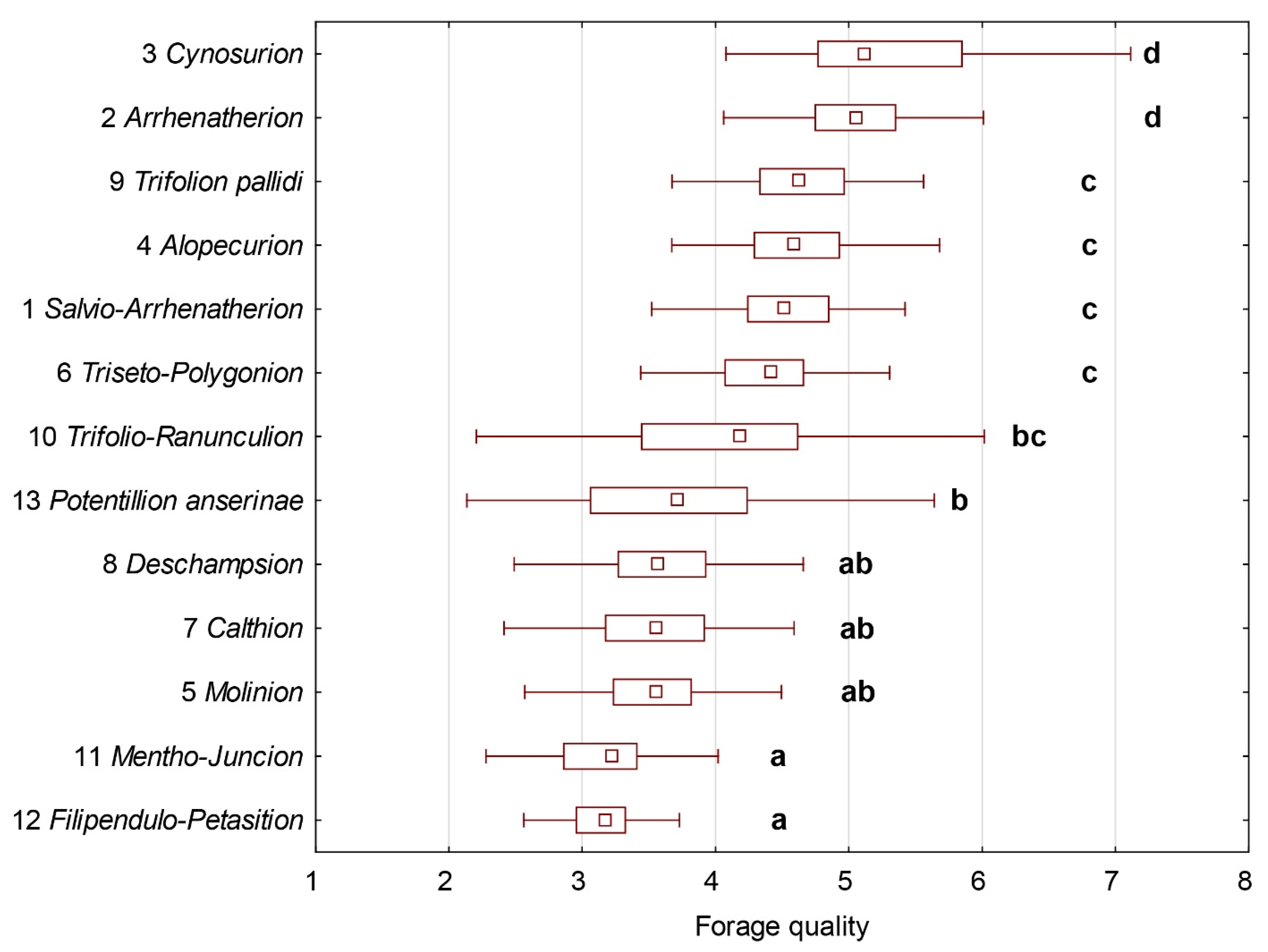
3.4. Forage Quality of Individual Meadow Types
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of vegetation plot | 109 | 332 | 95 | 341 | 179 | 60 | 130 | 216 | 142 | 68 | 54 | 33 | 257 |
| Pastinaca sativa | 85 | 51 | 8 | 6 | 1 | 5 | 1 | . | 17 | 3 | 7 | 3 | 1 |
| Lolium perenne | 8 | 26 | 97 | 20 | . | 10 | 7 | 2 | 7 | 7 | 20 | . | 30 |
| Trifolium repens | 28 | 33 | 89 | 38 | 6 | 13 | 18 | 24 | 29 | 24 | 52 | . | 59 |
| Taraxacum officinale | 34 | 40 | 89 | 40 | 6 | 15 | 10 | 18 | 43 | 22 | 17 | 3 | 38 |
| Plantago major | 11 | 3 | 61 | 5 | 1 | 5 | 6 | 2 | 6 | . | 24 | . | 47 |
| Cynosurus cristatus | . | 34 | 20 | 90 | 18 | 27 | 44 | 36 | 29 | . | 4 | . | 9 |
| Trifolium patens | 6 | 11 | . | 71 | 10 | . | 38 | 37 | 31 | 4 | 4 | . | 4 |
| Galium verum | 46 | 43 | 2 | 65 | 45 | 55 | 8 | 16 | 53 | 43 | 2 | 9 | 1 |
| Succisa pratensis | . | . | . | 17 | 69 | 3 | 20 | 14 | . | . | 2 | 33 | . |
| Carex panicea | . | 2 | . | 17 | 63 | . | 50 | 25 | . | . | 2 | 6 | . |
| Bellis perennis | 11 | 29 | 40 | 18 | 1 | 93 | 8 | . | 15 | 10 | . | . | 13 |
| Stellaria graminea | 5 | 19 | 6 | 23 | 26 | 93 | 8 | 13 | 37 | 9 | 2 | . | 3 |
| Briza media | 3 | 50 | . | 48 | 47 | 92 | 41 | 16 | 18 | 1 | . | . | . |
| Rhinanthus alectorolophus | 1 | 11 | . | 5 | . | 85 | 3 | 4 | 1 | . | . | . | . |
| Bromopsis erecta agg. | 1 | 12 | . | 1 | . | 78 | . | . | . | 1 | . | . | . |
| Knautia drymeia | 1 | 10 | 1 | 3 | 13 | 78 | 3 | . | . | . | . | . | . |
| Vicia cracca | 8 | 45 | 5 | 19 | 11 | 77 | 5 | 17 | 12 | 1 | . | 33 | 2 |
| Cruciata glabra | 1 | 6 | . | 4 | 25 | 75 | 5 | 1 | 1 | . | . | 6 | . |
| Trifolium alpestre | . | 1 | . | . | . | 73 | . | . | 1 | . | . | . | . |
| Trifolium medium | . | 8 | . | . | . | 73 | . | . | 7 | 3 | . | . | . |
| Galium album | 5 | 21 | 2 | 9 | . | 68 | 4 | 1 | 1 | . | 9 | 18 | 4 |
| Salvia pratensis | 9 | 34 | 2 | 1 | . | 68 | 2 | . | 13 | 4 | . | . | . |
| Leontodon hispidus | 11 | 45 | 7 | 53 | 32 | 67 | 24 | 11 | 17 | . | 2 | . | 3 |
| Luzula campestris | 1 | 26 | . | 31 | 22 | 63 | 5 | 6 | 4 | 9 | . | . | . |
| Avenula pubescens | 4 | 14 | 1 | 4 | 2 | 62 | 1 | 1 | . | . | . | . | . |
| Lysimachia nummularia | 6 | 18 | 4 | 36 | 16 | . | 65 | 58 | 27 | 3 | 28 | 6 | 35 |
| Equisetum palustre | . | 2 | . | 6 | 20 | . | 63 | 10 | 5 | . | 11 | 45 | 1 |
| Deschampsia cespitosa | 1 | 4 | 2 | 23 | 46 | 17 | 24 | 76 | 1 | . | 2 | 24 | 2 |
| Gratiola officinalis | . | 1 | . | 26 | 20 | . | 45 | 71 | 25 | . | 11 | 6 | 13 |
| Juncus effusus | . | 1 | . | 18 | 30 | . | 48 | 64 | 11 | 1 | 20 | 24 | 28 |
| Carex vulpina | 3 | 1 | . | 11 | 4 | . | 23 | 60 | 17 | 4 | 9 | . | 5 |
| Alopecurus pratensis | 20 | 23 | 16 | 44 | 25 | 20 | 9 | 50 | 92 | 57 | 7 | 12 | 8 |
| Poa pratensis | 57 | 53 | 33 | 48 | 17 | 22 | 20 | 31 | 69 | 28 | . | 3 | 8 |
| Trifolium pallidum | . | 1 | . | 1 | . | . | . | . | 68 | . | . | . | . |
| Ranunculus polyanthemos | 50 | 3 | 8 | 1 | 1 | 27 | . | . | 63 | 16 | . | . | 2 |
| Festuca valesiaca | 28 | 2 | 16 | . | 1 | . | . | . | 30 | 91 | . | . | . |
| Podospermum canum | 6 | . | 4 | . | . | . | . | . | . | 65 | . | . | . |
| Bromus hordeaceus | 24 | 14 | 11 | 4 | . | 7 | 4 | 1 | 24 | 62 | . | . | 3 |
| Mentha longifolia | 23 | 2 | 8 | 2 | . | . | 2 | . | 9 | . | 100 | 18 | 6 |
| Juncus inflexus | 1 | . | 2 | 1 | 4 | . | 22 | 4 | 1 | . | 96 | 3 | 12 |
| Filipendula ulmaria | . | 1 | . | 11 | 23 | . | 29 | 17 | . | . | 4 | 100 | 1 |
| Carex acuta | . | 1 | . | 4 | 8 | . | 8 | 23 | . | . | 4 | 67 | . |
| Rorippa sylvestris | 5 | 1 | 9 | 1 | . | . | . | 6 | 13 | 9 | 20 | 3 | 62 |
| Arrhenatherum elatius | 78 | 96 | 11 | 13 | 3 | 58 | 4 | . | 12 | 4 | . | 6 | . |
| Daucus carota | 67 | 54 | 23 | 67 | 11 | 20 | 9 | 6 | 36 | 25 | 19 | . | 11 |
| Trisetum flavescens | 3 | 81 | 3 | 28 | 3 | 97 | 5 | . | 4 | . | . | . | 1 |
| Rumex acetosa | 20 | 64 | 14 | 65 | 30 | 12 | 35 | 28 | 48 | 1 | . | 15 | 3 |
| Anthoxanthum odoratum | 4 | 57 | . | 80 | 34 | 82 | 58 | 40 | 47 | 29 | . | 3 | . |
| Lychnis flos-cuculi | 7 | 21 | 4 | 66 | 39 | 20 | 59 | 63 | 50 | 4 | 4 | 21 | 2 |
| Betonica officinalis | . | 20 | . | 65 | 72 | 28 | 36 | 33 | 31 | 1 | . | 6 | . |
| Potentilla erecta | . | 6 | . | 26 | 83 | 67 | 29 | 16 | . | 3 | . | 6 | . |
| Galium palustre | . | 1 | . | 25 | 31 | . | 65 | 78 | 7 | . | 17 | 33 | 30 |
| Lysimachia vulgaris | . | 2 | 2 | 11 | 50 | 2 | 61 | 30 | 4 | . | 4 | 73 | 1 |
| Agrostis stolonifera | 6 | 5 | 18 | 28 | 17 | . | 12 | 26 | 11 | 3 | 81 | 21 | 94 |
| Dactylis glomerata | 83 | 85 | 41 | 26 | 17 | 95 | 14 | 1 | 24 | 3 | 17 | 12 | 2 |
| Centaurea jacea agg. | 41 | 81 | 17 | 58 | 51 | 85 | 25 | 24 | 66 | 21 | 7 | 12 | 5 |
| Holcus lanatus | 6 | 74 | 12 | 85 | 53 | 53 | 84 | 53 | 13 | . | 13 | 3 | 5 |
| Leucanthemum vulgare agg. | 28 | 73 | 7 | 76 | 37 | 90 | 32 | 33 | 38 | 4 | . | 6 | 1 |
| Schedonorus pratensis | 19 | 64 | 20 | 81 | 25 | 45 | 56 | 47 | 81 | . | . | 3 | 12 |
| Lotus corniculatus | 46 | 64 | 42 | 62 | 50 | 80 | 22 | 22 | 49 | 10 | 2 | 3 | 11 |
| Prunella vulgaris | 15 | 36 | 26 | 60 | 51 | 60 | 49 | 60 | 23 | 1 | 43 | 3 | 32 |
| Ranunculus repens | 9 | 11 | 26 | 47 | 25 | . | 85 | 85 | 17 | . | 85 | 27 | 53 |
| Lythrum salicaria | 2 | 2 | . | 14 | 41 | 2 | 68 | 63 | 12 | 1 | 33 | 91 | 14 |
| Plantago lanceolata | 38 | 80 | 55 | 89 | 44 | 62 | 70 | 57 | 50 | 34 | 17 | 3 | 25 |
| Trifolium pratense | 72 | 83 | 43 | 90 | 27 | 60 | 60 | 39 | 55 | 29 | 9 | 3 | 12 |
| Ranunculus acris | 6 | 79 | 26 | 90 | 78 | 38 | 69 | 60 | 9 | . | 22 | 12 | 7 |
| Achillea millefolium agg. | 76 | 77 | 79 | 44 | 31 | 88 | 6 | 4 | 68 | 63 | 19 | 3 | 13 |
References
- Mucina, L.; Bültmann, H.; Dierssen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Škvorc, Ž.; Ćuk, M.; Zelnik, I.; Franjić, J.; Igić, R.; Ilić, M.; Krstonošić, D.; Vukov, D.; Čarni, A. Diversity of wet and mesic grasslands along a climatic gradient on the southern margin of the Pannonian Basin. Appl. Veg. Sci. 2020, 23, 676–697. [Google Scholar] [CrossRef]
- Janssen, A.M.; Rodwell, J.S.; García Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Ssysmak, A.; et al. European Red List of Habitats—Part 2. Terrestrial and Freshwater Habitats; Publication office of EU: Luxembourg, 2016. [Google Scholar]
- Herzon, I.; Raatikainen, K.J.; Wehn, S.; Rūsiņa, S.; Helm, A.; Cousins, S.A.O.; Rašomavičius, V. Semi-natural habitats in boreal europe: A rise of a social-ecological research agenda. Ecol. Soc. 2021, 26, 13. [Google Scholar] [CrossRef]
- Hopkins, A.; Holz, B. Grassland for agriculture and nature conservation: Production, quality and multi-functionality. Agron. Res. 2006, 4, 3–20. [Google Scholar]
- Schaub, S.; Finger, R.; Leiber, F.; Probst, S.; Kreuzer, M.; Weigelt, A.; Buchmann, N.; Scherer-Lorenzen, M. Plant diversity effects on forage quality, yield and revenues of semi-natural grasslands. Nat. Commun. 2020, 11, 768. [Google Scholar] [CrossRef] [Green Version]
- Briemle, G.; Nitsche, S.; Nitsche, L. Nutzungswertzahlen für Gefäßpflanzen des Grünlandes. Schriftr. Für Veg. 2002, 38, 203–225. [Google Scholar]
- Kaiser, T.; Ahlborn, J. Long-term vegetation monitoring in the floodplain grasslands of the lower Havel Valley (northeastern Germany) and conclusions for sustainable management practices. J. Nat. Conserv. 2021, 63, 126053. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An international terminology for grazing lands and grazing animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Chytrý, M.; Hennekens, S.M.; Jiménez-Alfaro, B.; Knollová, I.; Dengler, J.; Jansen, F.; Landucci, F.; Schaminée, J.H.J.; Aćić, S.; Agrillo, E.; et al. European Vegetation Archive (EVA): An integrated database of European vegetation plots. Appl. Veg. Sci. 2016, 19, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liu, Z. Grassland ecosystem services: A systematic review of research advances and future directions. Landsc. Ecol. 2020, 35, 793–814. [Google Scholar] [CrossRef]
- Pittarello, M.; Lonati, M.; Gorlier, A.; Perotti, E.; Probo, M.; Lombardi, G. Plant diversity and pastoral value in alpine pastures are maximized at different nutrient indicator values. Ecol. Indic. 2018, 85, 518–524. [Google Scholar] [CrossRef]
- Kun, R.; Babai, D.; Csathó, A.I.; Vadász, C.; Kálmán, N.; Máté, A.; Malatinszky, Á. Simplicity or complexity? Important aspects of high nature value grassland management in nature conservation. Biodivers. Conserv. 2021, 30, 3563–3583. [Google Scholar] [CrossRef]
- Ranta, M.; Rotar, I.; Vidican, R.; Mălinaș, A.; Ranta, O.; Lefter, N. Influence of the uan fertilizer application on quantitative and qualitative changes in semi-natural grassland in western Carpathians. Agronomy 2021, 11, 267. [Google Scholar] [CrossRef]
- Dullau, S.; Rydgren, K.; Kirmer, A.; Jäger, U.G.; Meyer, M.H.; Tischew, S. The dessau grassland experiment-impact of fertilization on forage quality and species assembly in a species-rich alluvial meadow. Agriculture 2021, 11, 339. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Knollová, I.; Janssen, J.A.M.; Rodwell, J.S.; Peterka, T.; Marcenò, C.; Landucci, F.; Danihelka, J.; et al. EUNIS Habitat Classification: Expert system, characteristic species combinations and distribution maps of European habitats. Appl. Veg. Sci. 2020, 23, 648–675. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L. National vegetation classification of the Czech Republic: A summary of the approach. Phytocoenologia 2018, 48, 121–131. [Google Scholar] [CrossRef]
- van’t Veen, H.; Chalmandrier, L.; Sandau, N.; Nobis, M.P.; Descombes, P.; Psomas, A.; Hautier, Y.; Pellissier, L. A landscape-scale assessment of the relationship between grassland functioning, community diversity, and functional traits. Ecol. Evol. 2020, 10, 9906–9919. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Ewald, J. The sensitivity of Ellenberg indicator values to the completeness of vegetation relevés. Basic Appl. Ecol. 2003, 4, 507–513. [Google Scholar] [CrossRef]
- Euro+Med. Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. 2006. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 1 September 2021).
- Mládek, J.; Mládková, P.; Hejcmanová, P.; Dvorský, M.; Pavlu, V.; De Bello, F.; Duchoslav, M.; Hejcman, M.; Pakeman, R.J. Plant trait assembly affects superiority of grazer’s foraging strategies in species-rich grasslands. PLoS ONE 2013, 8, e69800. [Google Scholar] [CrossRef] [Green Version]
- Pauler, C.M.; Isselstein, J.; Suter, M.; Berard, J.; Braunbeck, T.; Schneider, M.K. Choosy grazers: Influence of plant traits on forage selection by three cattle breeds. Func. Ecol. 2020, 34, 980–992. [Google Scholar] [CrossRef]
- Busch, V.; Klaus, V.H.; Schäfer, D.; Prati, D.; Boch, S.; Müller, J.; Chisté, M.; Mody, K.; Blüthgen, N.; Fischer, M.; et al. Will I stay or will I go? Plant species-specific response and tolerance to high land-use intensity in temperate grassland ecosystems. J. Veg. Sci. Veg. Sci. 2019, 30, 674–686. [Google Scholar] [CrossRef]
- Bengtsson, J. Which species? What kind of diversity? Which ecosystem function? Some problems in studies of relations between biodiversity and ecosystem function. Appl. Soil Ecol. 1998, 10, 191–199. [Google Scholar] [CrossRef]
- Luo, J.; Liu, X.; Yang, J.; Liu, Y.; Zhou, J. Variation in plant functional groups indicates land degradation on the Tibetan Plateau. Scienfific Rep. 2018, 8, 17606. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties, 2nd ed.; Wiley: New York, NY, USA, 2001. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. BIOLFLOR—Eine Datenbank zu Biologisch-Ökologischen Merkmalen der Gefäßpflanzen in Deutschland; Schriftenreihe für Vegetationskunde 38; Bundesamt für Naturschutz (BfN): Bonn, Germany, 2002. [Google Scholar]
- Hunt, R.; Hodgson, J.; Thompson, K.; Bungener, P.; Dunnett, N.; Askwe, A. A new practical tool for deriving a functional signature for herbaceous vegetation. Appl. Veg. Sci. 2004, 7, 163–170. [Google Scholar] [CrossRef]
- Tichý, L.; Holt, J.; Nejezchlebová, M. Juice. In Program for Management, Analysis and Classification of Ecological Data; Vegetation Science Group, Masaryk University: Brno, Germany, 2006. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Dull, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa; Scripta Geobotanica 18, 1-285; Verlag Erich Goltze: Göttingen, Germany, 1992; ISBN 3884525182. [Google Scholar]
- Troiani, N.; Tradella, F.M.; Malatesta, L.; Corazza, M.; Ferrari, C.; Catorci, A. Long-term cropland abandonment does not lead per se to the recovery of semi-natural herb communities deemed habitats of community interest. Acta Bot. Croat. 2016, 75, 226–235. [Google Scholar] [CrossRef]
- StatSoft, I. Electronic Statistics Textbook; Statsoft: Tulsa, Oklahoma, 2011. [Google Scholar]
- Lengyel, A.; Swacha, G.; Botta-Dukát, Z.; Kącki, Z. Trait-based numerical classification of mesic and wet grasslands in Poland. J. Veg. Sci. 2020, 31, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Čarni, A.; Ćuk, M.; Zelnik, I.; Franjić, J.; Igić, R.; Ilić, M.; Krstonošić, D.; Vukov, D.; Škvorc, Ž. Wet meadow plant communities of the alliance Trifolion pallidi on the southeastern margin of the Pannonian plain. Water 2021, 13, 381. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Horvatić, S. Soziologische Einheiten der Niederungswiesen in Kroatien und Slavonien. Acta Bot. Croat. 1930, 5, 57–118. [Google Scholar]
- Szépligeti, M.; Kőrösi, Á.; Szentirmai, I.; Házi, J.; Bartha, D.; Bartha, S. Evaluating alternative mowing regimes for conservation management of Central European mesic hay meadows: A field experiment. Plant Biosyst. 2018, 152, 90–97. [Google Scholar] [CrossRef]
- Swacha, G.; Botta-Dukát, Z.; Kącki, Z.; Pruchniewicz, D.; Zołnierz, L. The effect of abandonment on vegetation composition and soil properties in Molinion meadows (SW Poland). PLoS ONE 2018, 13, e0197363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, K.R.; Kučera, T. Management effects on plant species composition and ecosystem processes and services in a nutrient-poor wet grassland. Plant Ecol. 2019, 220, 1009–1020. [Google Scholar] [CrossRef]
- Müller, I.B.; Buhk, C.; Alt, M.; Entling, M.H.; Schirmel, J. Plant functional shifts in Central European grassland under traditional flood irrigation. Appl. Veg. Sci. 2016, 19, 122–131. [Google Scholar] [CrossRef]
- Dwire, K.A.; Kauffman, J.B.; Brookshire, E.N.J.; Baham, J.E. Plant biomass and species composition along an environmental gradient in montane riparian meadows. Oecologia 2004, 139, 309–317. [Google Scholar] [CrossRef]
- Hoover, D.L.; Knapp, A.K.; Smith, M.D. Resistance and resilience of a grassland ecosystem to climate extremes. Ecology 2014, 95, 2646–2656. [Google Scholar] [CrossRef] [Green Version]
- Valkó, O.; Török, P.; Matus, G.; Tóthmérész, B. Is regular mowing the most appropriate and cost-effective management maintaining diversity and biomass of target forbs in mountain hay meadows? Flora—Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 303–309. [Google Scholar] [CrossRef]
- Prins, H.H.T.; Ydenberg, R.C.; Drent, R.H. The interaction of Brent Geese Branta bernicla and Sea Plantain Plantago maritima during spring staging: Field observations and experiments. Acta Bot. Neerl. 1980, 29, 585–596. [Google Scholar] [CrossRef]
- Gaberščik, A.; Krek, J.L.; Zelnik, I. Habitat diversity along a hydrological gradient in a complex wetland results in high plant species diversity. Ecol. Eng. 2018, 118, 84–92. [Google Scholar] [CrossRef]
- Bruelheide, H.; Dengler, J.; Purschke, O.; Lenoir, J.; Jiménez-Alfaro, B.; Hennekens, S.M.; Botta-Dukát, Z.; Chytrý, M.; Field, R.; Jansen, F.; et al. Global trait–environment relationships of plant communities. Nat. Ecol. Evol. 2018, 2, 1906–1917. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Hellström, K.; Huhta, A.-P.; Rautio, P.; Tuomi, J.; Oksanen, J.; Laine, K. Use of sheep grazing in the restoration of semi-natural meadows in northern Finland. Appl. Veg. Sci. 2003, 6, 45–52. [Google Scholar] [CrossRef]
- Chelli, S.; Marignani, M.; Barni, E.; Petraglia, A.; Puglielli, G.; Wellstein, C.; Acosta, A.T.R.; Bolpagni, R.; Bragazza, L.; Campetella, G.; et al. Plant—Environment interactions through a functional traits perspective: A review of Italian studies. Plant Biosyst. 2019, 153, 853–869. [Google Scholar] [CrossRef]
- Slavnić, Ž. Slatinska vegetacija Vojvodine. Arh. Za Poljopr. Nauk. I Teh. 1948, 3, 77–143. [Google Scholar]
- Čarni, A. Staudenfluren-und Ufervegetation (Verbände Filipendulion Segal 1966 und Senecion fluviatilis R. Tx. (1947)1950 em. 1967) im Krško Becken. Biološki Vestn. 1995, 40, 71–85. [Google Scholar]
- Dindová, A.; Hakl, J.; Hrevušová, Z.; Nerušil, P. Relationships between long-term fertilization management and forage nutritive value in grasslands. Agric. Ecosyst. Environ. 2019, 279, 139–148. [Google Scholar] [CrossRef]
- Pierik, M.E.; Gusmeroli, F.; Della Marianna, G.; Tamburini, A.; Bocchi, S. Meadows species composition, biodiversity and forage value in an Alpine district: Relationships with environmental and dairy farm management variables. Agric. Ecosyst. Environ. 2017, 244, 14–21. [Google Scholar] [CrossRef]
- Tzialla, C.E.; Veresoglou, D.S.; Papakosta, D.; Mamolos, A.P. Changes in soil characteristics and plant species composition along a moisture gradient in a Mediterranean pasture. J. Environ. Manag. 2006, 80, 90–98. [Google Scholar] [CrossRef]
- Kim, S.-H.; Kim, T.-K. Economic valuation of multi-functionality on an eco-pastoral system in Alpine grassland. J. Korean Soc. Grassl. Forage Sci. 2018, 38, 298–309. [Google Scholar] [CrossRef]
- Kaligarič, M.; Ivanjšič, D. Vanishing landscape of the “classic” Karst: Changed landscape identity and projections for the future. Landsc. Urban Plan. 2014, 132, 148–158. [Google Scholar] [CrossRef]
- Prach, K. Vegetation Changes in a wet meadow complex during the past half-century. Folia Geobot. 2008, 43, 119–130. [Google Scholar] [CrossRef]
- Kaligarič, M.; Čuš, J.; Škornik, S.; Ivajnšič, D. The failure of agri-environment measures to promote and conserve grassland biodiversity in Slovenia. Land Use Policy 2019, 80, 127–134. [Google Scholar] [CrossRef]
- Isbell, F.; Tilman, D.; Reich, P.; Clark, A. Deficits of biodiversity and productivity linger a century after agricultural abandonment. Nat. Ecol. Evol. 2019, 3, 1533–1538. [Google Scholar] [CrossRef] [PubMed]
| Variable | Forage Quality |
|---|---|
| EIV Moisture | −0.67 |
| EIV Nutrients | - |
| Species with the following habitat preferences (behavior) | |
| grasslands | 0.27 |
| forests and their successional series | −0.26 |
| dry grasslands | 0.28 |
| weed and ruderal habitats | 0.55 |
| saline habitats | - |
| humid environment | −0.73 |
| Functional groups | |
| grasses | 0.51 |
| sedges | −0.61 |
| legume | - |
| forbs | −0.36 |
| Ecological strategies | |
| c (competitor) | 0.34 |
| s (stress tolerator) | −0.63 |
| r (ruderal) | - |
| Management | |
| Mowing | 0.75 |
| Grazing | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čarni, A.; Ćuk, M.; Krstonošić, D.; Škvorc, Ž. Study of Forage Quality of Grasslands on the Southern Margin of the Pannonian Basin. Agronomy 2021, 11, 2132. https://doi.org/10.3390/agronomy11112132
Čarni A, Ćuk M, Krstonošić D, Škvorc Ž. Study of Forage Quality of Grasslands on the Southern Margin of the Pannonian Basin. Agronomy. 2021; 11(11):2132. https://doi.org/10.3390/agronomy11112132
Chicago/Turabian StyleČarni, Andraž, Mirjana Ćuk, Daniel Krstonošić, and Željko Škvorc. 2021. "Study of Forage Quality of Grasslands on the Southern Margin of the Pannonian Basin" Agronomy 11, no. 11: 2132. https://doi.org/10.3390/agronomy11112132
APA StyleČarni, A., Ćuk, M., Krstonošić, D., & Škvorc, Ž. (2021). Study of Forage Quality of Grasslands on the Southern Margin of the Pannonian Basin. Agronomy, 11(11), 2132. https://doi.org/10.3390/agronomy11112132