Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils


 , ,
, ,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Statistical Analysis
2.2. Bayesian Statistics and Decision Trees
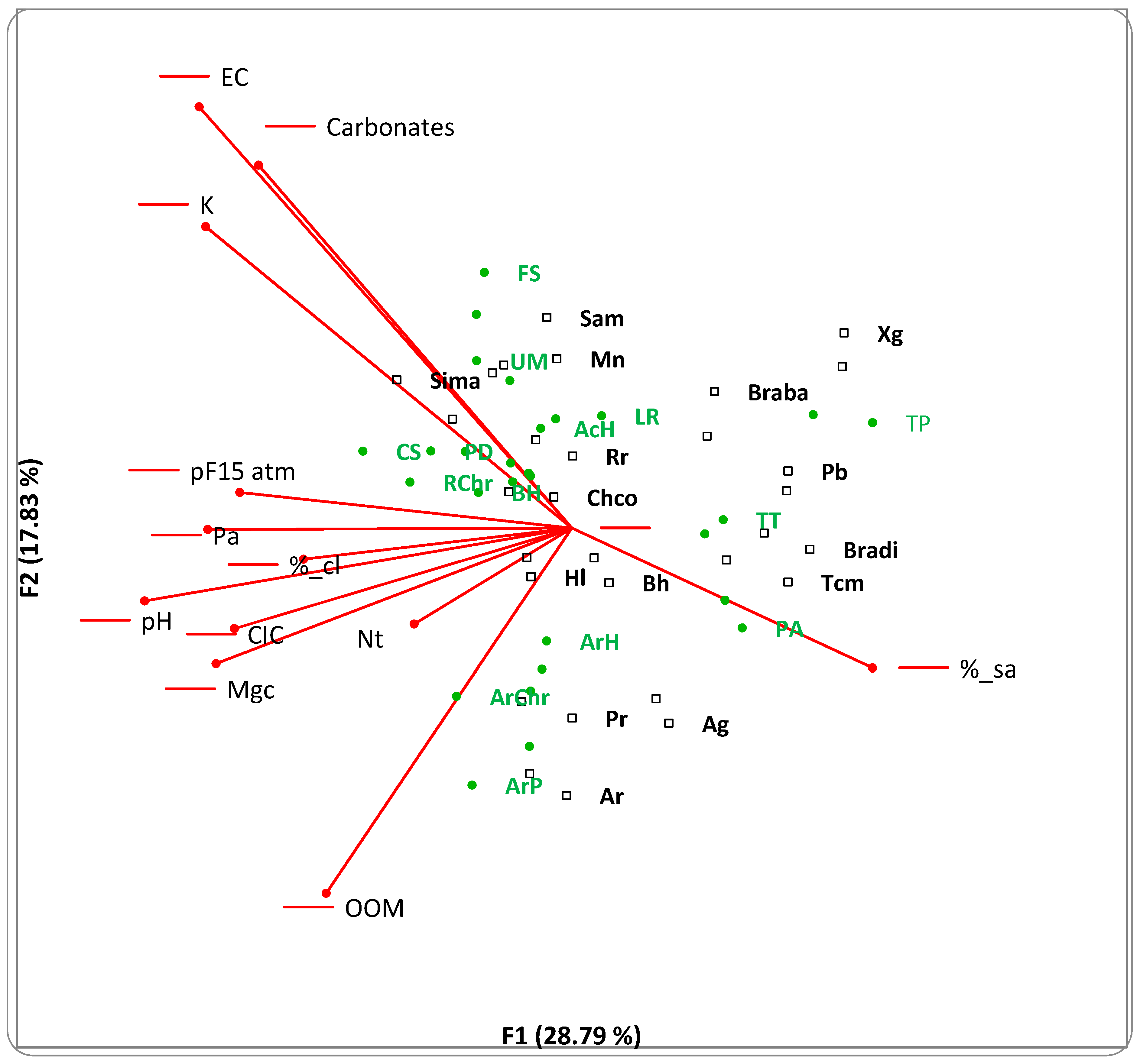
3. Results
3.1. Bayesian Analysis
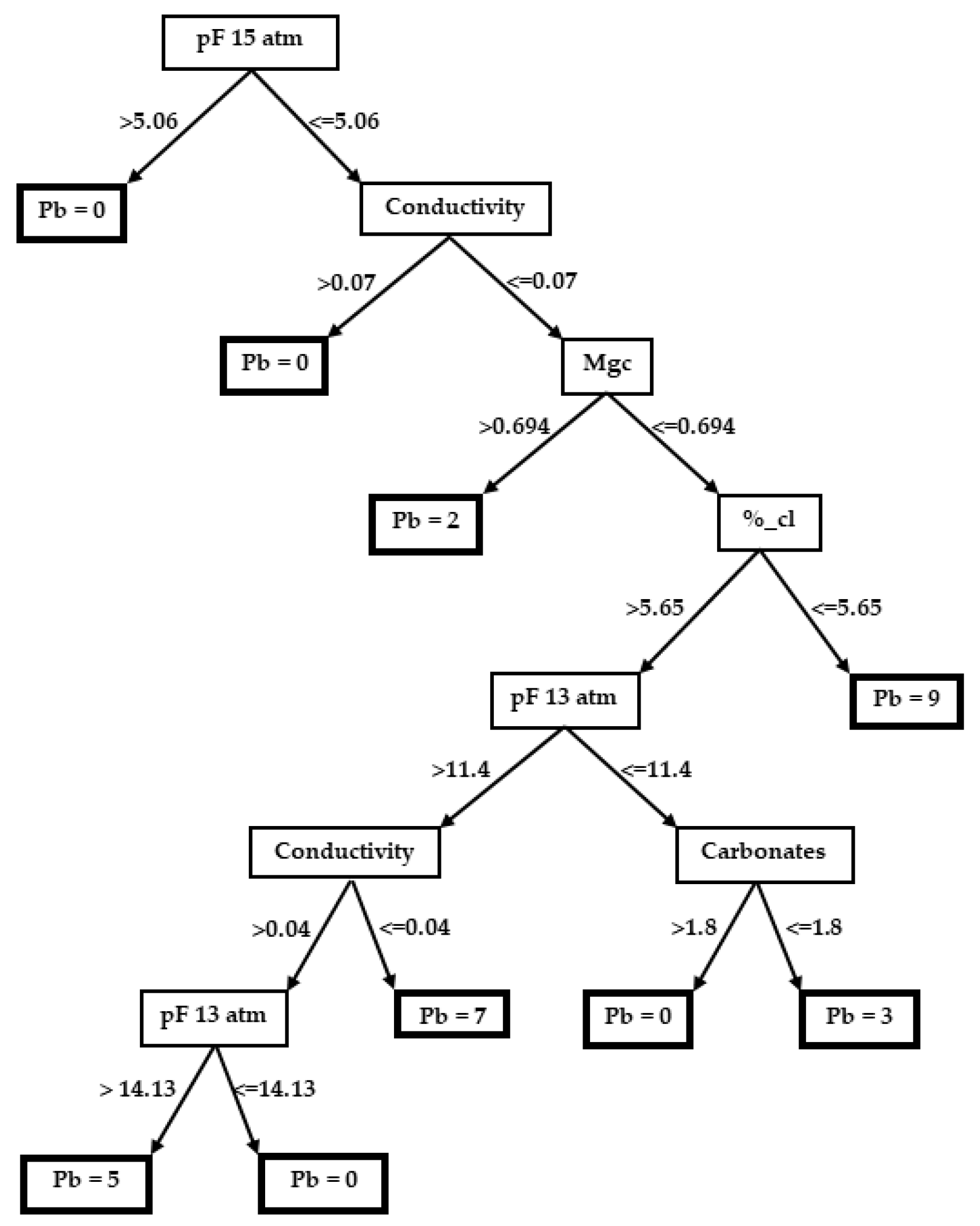
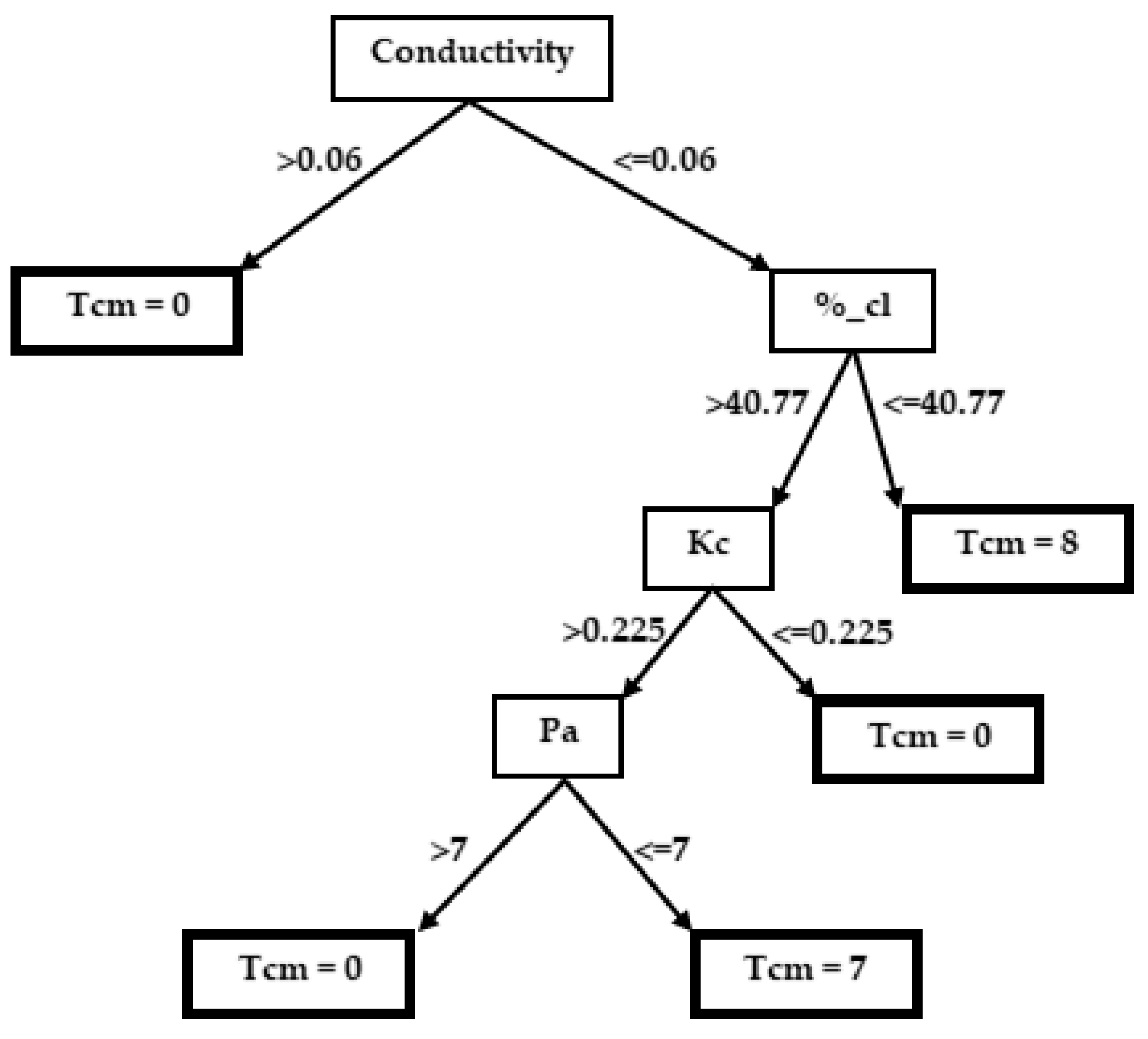
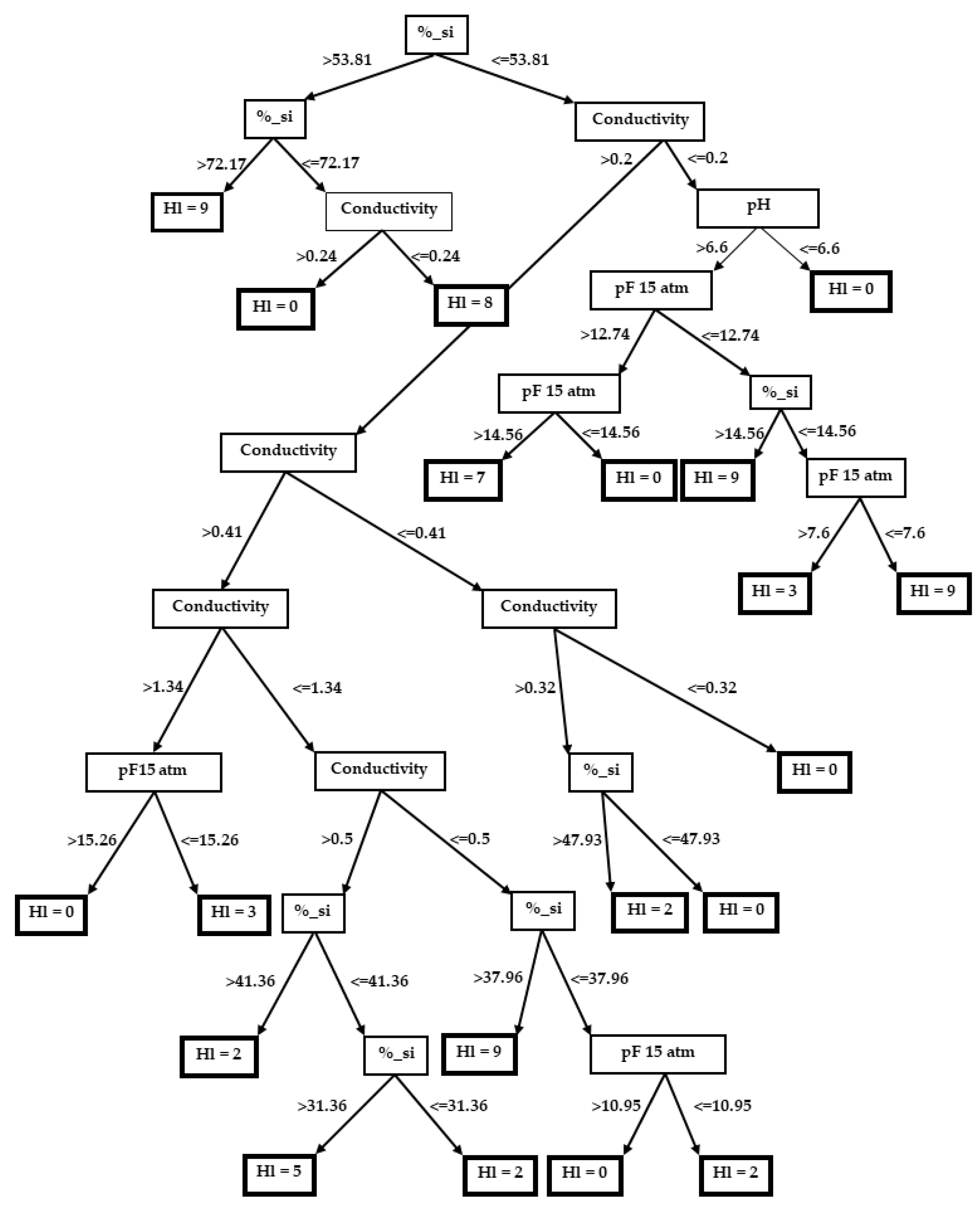
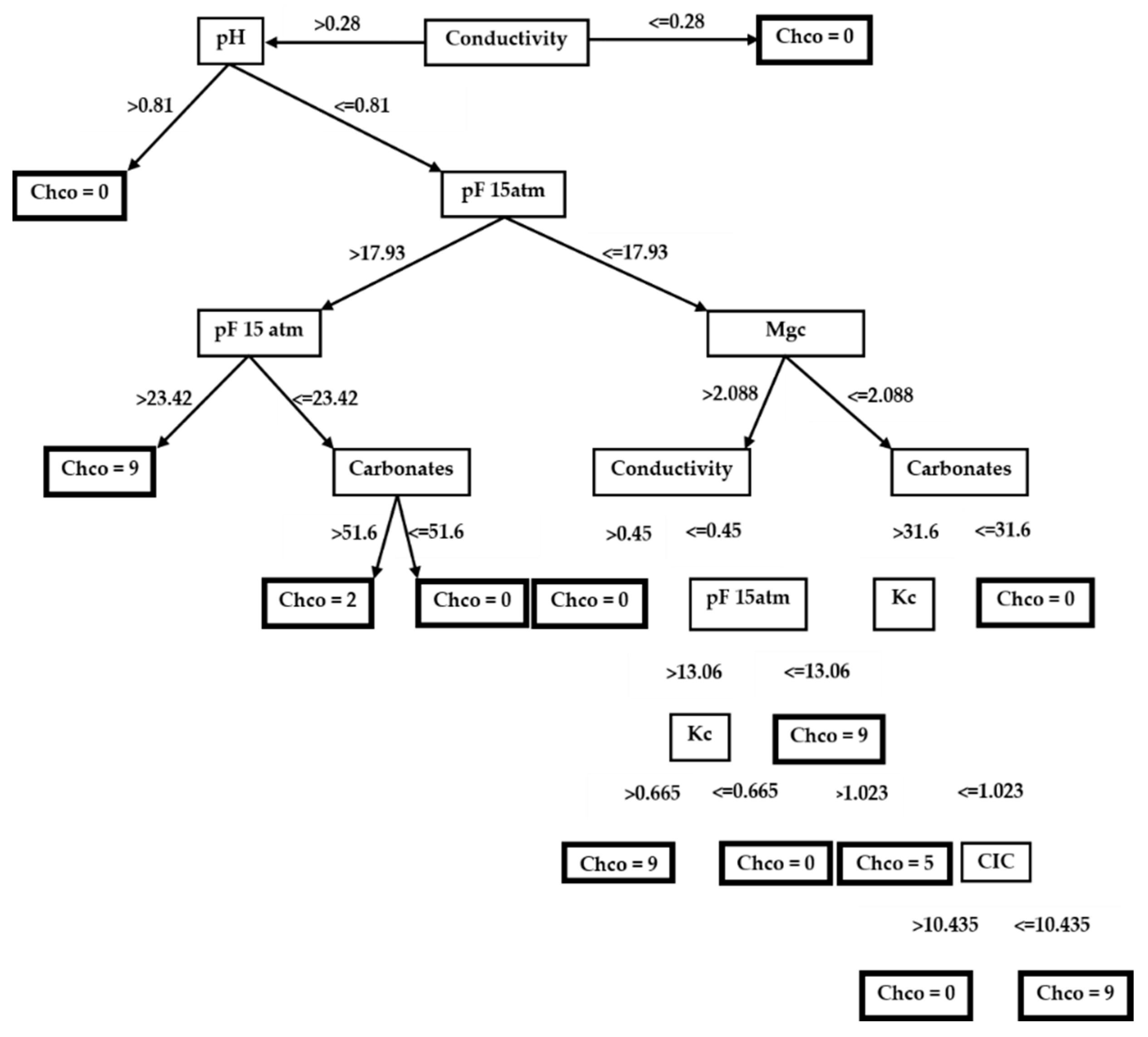
3.2. Decision Trees
3.2.1. Plantago bellardii
3.2.2. Taeniatherum caput-medusae
3.2.3. Hordeum leporinum
3.2.4. Glebionis discolor (=Chrysanthemum coronarium var. discolor)
3.3. Soil Attributes of the Other Species Studied
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cano-Ortiz, A.; Pinto Gomes, C.J.; Cano, E. Current situation of the Gaudinio fragilis—Hordeion bulbosi alliance in the Iberian Peninsula. Acta Bot. Gall. 2009, 156, 19–31. [Google Scholar] [CrossRef][Green Version]
- Perrino, E.V.; Ladisa, G.; Calabrese, G. Flora and plant genetic resources of ancient olive groves of Apulia (southern Italy). Genet. Resour. Crop Evol. 2014, 61, 23–53. [Google Scholar] [CrossRef]
- Perrino, E.V.; Magazzini, P.; Musarella, C.M. Management of grazing “buffalo” to preserve habitats by Directive 92/43 EEC in a wetland protected area of the Mediterranean coast: Palude Frattarolo, Apulia, Italy. Euro-Mediterr. J. Environ. Integr. 2020, in press. [Google Scholar]
- García Fuentes, A.; Cano, E. Malas Hierbas del Olivar Giennense; Instituto de Estudios Giennenses: Jaén, Spain, 1996; 213p. [Google Scholar]
- Pastor Muñoz-Cobo, M. Estudio de Diversos Métodos de Manejo del Suelo Alternativos al Laboreo en el Cultivo del Olivo; Instituto de Estudios Giennenses: Jaén, Spain, 1991; 302p. [Google Scholar]
- Saavedra Saavedra, M.M.; Pastor Muñoz-Cobo, M. Sistemas de Cultivo en Olivar; Agrícola Española, S.A., Ed.; Manejo de mals hierbas y herbicidas: Madrid, Spain, 2002; 429p. [Google Scholar]
- Giusso del Galdo, G.; Marcenò, C.; Musarella, C.M.; Sciandrello, S. La vegetazione costiera della R.N.O. “Torre Salsa” (Siculiana—AG). Inf. Bot. Ital. 2008, 40, 73–89. [Google Scholar]
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.M.; Pinto-Gomes, C. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands—Portugal case study. Plant Biosyst. 2015, 149, 494–502. [Google Scholar] [CrossRef]
- Signorino, G.; Cannavò, S.; Crisafulli, A.; Musarella, C.M.; Spampinato, G. Fagonia cretica L. In Schede per una Lista Rossa della Flora vascolare e crittogamica Italiana. Inf. Bot. Ital. 2011, 43, 381–458. [Google Scholar]
- Puglisi, M.; Sciandrello, S.; Musarella, C.M.; Spampinato, G.; Privitera, M.; Tomaselli, V. Bryosociological remarks on garrigue environments in Apulia Region (Southern Italy). Plant Sociol. 2019, 56, 43–52. [Google Scholar] [CrossRef]
- Quinto-Canas, R.; Mendes, P.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C. Forest fringe communities of the southwestern Iberian Peninsula. Rev. Chapingo Ser. Cienc. 2018, 24, 415–434. [Google Scholar] [CrossRef]
- Sciandrello, S.; Musarella, C.M.; Puglisi, M.; Spampinato, G.; Tomaselli, V.; Minissale, P. Updated and new insights on the coastal halophilous vegetation of southeastern Sicily (Italy). Plant Sociol. 2019, 56, 81–98. [Google Scholar] [CrossRef]
- Spampinato, G.; Musarella, C.M.; Cano-Ortiz, A.; Signorino, G. Habitat, occurrence and conservation status of the Saharo-Macaronesian and Southern-Mediterranean element Fagonia cretica L. (Zygophyllaceae) in Italy. J. Arid Land 2018, 10, 140–151. [Google Scholar] [CrossRef]
- Spampinato, G.; Sciandrello, S.; del Galdo, G.; Puglisi, M.; Tomaselli, V.; Cannavò, S.; Musarella, C.M. Contribution to the knowledge of Mediterranean wetland biodiversity: Plant communities of the Aquila Lake (Calabria, Southern Italy). Plant Sociol. 2019, 56, 53–68. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Latini, M.; Iberite, M.; Bonari, G.; Nicolella, G.; Rosati, L.; Salerno, G.; Abbate, G. The segetal flora of winter cereals and allied crops in Italy: Species inventory with chorological, structural and ecological features. Plant Biosyst. 2020, 154, 935–946. [Google Scholar] [CrossRef]
- Cano-Ortiz, A.; Biondi, E.; Pinto-Gomes, C.J.; Del Río González, S.; Cano, E. Soil and phytosociological characterization of grasslands in the western Mediterranean. Am. J. Plant Sci. 2014, 5, 3213–3240. [Google Scholar] [CrossRef][Green Version]
- Cano-Ortiz, A. Bioindicadores ecológicos y manejo de cubiertas vegetales como herramienta para la implantación de una agricultura sostenible. Ph.D. Thesis, University of Jaén, Jaén, Spain, 2007. [Google Scholar]
- Taffetani, F.; Rismondo, M. Bioindicator system for the evaluation of the environmental quality of agro-ecosystems. Fitosociologia 2009, 46, 3–22. [Google Scholar]
- Baldoni, M.; Biondi, E.; Loiotile, A. La vegetazione infestante i vigneti nelle Marche. Fitosociologia 2001, 38, 63–68. [Google Scholar]
- Biondi, E.; Baldoni, M. La vegetazione di margine stradale dell’ordine Brometalia rubenti-tectori nell’Italia Centrale. Ann. Bot. 1991, 49 (Suppl. 8), 214–217. [Google Scholar]
- Biondi, E.; Bagella, S.; Casavecchia, S.; Pinzi, M.; Vagge, I. La vegetazione a Dasypyrum villosum (L.) P. Candargy lungo le coste dell’Italia Settentrionale. Doc. Phytosociol. 1999, 19, 439–446. [Google Scholar]
- Cano-Ortiz, A.; Pinto-Gomes, C.J.; Esteban Ruiz, F.J.; Rodríguez Torres, A.; Goñi, J.; De la Haza, I.; Cano, E. Biodiversity of Hordeion leporini in Portugal: A phytosociological and edaphic análisis. Acta Bot. Gall. 2009, 156, 33–48. [Google Scholar] [CrossRef]
- Cano-Ortiz, A.; Pinto-Gomes, C.J.; Esteban Ruiz, F.J.; Cano, E. Determination of the nutritional state of soils by means of the phytosociological method and different statistical techniques (Bayesian statistics and decision trees) in Spain. Acta Bot. Gall. 2009, 156, 607–624. [Google Scholar] [CrossRef][Green Version]
- Cano-Ortiz, A.; Pinto-Gomes, C.J.; Cano, E. Contribution to the study of the Taeniathero-Aegilopion geniculatae alliance in Portugal. Acta Bot. Gall. 2010, 157, 599–610. [Google Scholar] [CrossRef][Green Version]
- Quinto-Canas, R.; Mendes, P.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C. The Agrostion castellanae Rivas Goday 1957 corr. Rivas Goday & Rivas-Martínez 1963 alliance in the southwestern Iberian Peninsula. Plant Sociol. 2018, 55, 21–29. [Google Scholar] [CrossRef]
- Rivas-Martínez, S. La vegetación de Hordeion leporini en España. Doc. Phytosoc. 1978, 9, 377–392. [Google Scholar]
- Di Pietro, R.; Theurillat, J.P.; Capelo, J.; Fernández-González, F.; Terzi, M.; Carni, A.; Mucina, L. Nomenclature and syntaxonomic notes on some high-rank syntaxa of the European grassland vegetation. Lazaroa 2015, 36, 79–106. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Moretti, M.; Abbate, G. The anthropogenic grasslands of the Securigero securidacae-Dasypyrion villosi in central mediterranean areas: Synecology, distribution and syntaxonomy. Ann. Bot. 2019, 9, 1–38. [Google Scholar] [CrossRef]
- Perrino, E.V.; Tomaselli, V.; Signorile, G.; Angiulli, F.; Silletti, G. Vegetation to Crambe hispanica L. in Apulia Region (Vegetazione a Crambe hispanica L. in Puglia). Fitosociologia 2011, 48, 99–107. [Google Scholar]
- Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Spampinato, G.; Pinto Gomes, C.J. Morphometric analysis bioclimatic distribution of Glebionis coronaria s.l. (Asteraceae) in the Mediterranean area. PhytoKeys 2017, 81, 103–126. [Google Scholar] [CrossRef] [PubMed]
- Perrino, E.V.; Tomaselli, V.; Costa, R.; Pavone, P. Conservation status of habitats (Directive 92/43 EEC) of coastal and low hill belts in a mediterranean biodiversity hot spot (Gargano—Italy). Plant Biosyst. 2013, 147, 1006–1028. [Google Scholar] [CrossRef]
- Perrino, E.V.; Calabrese, G. Endangered segetal species in southern Italy: Distribution, conservation status, trends, actions and ethnobotanical notes. Genet. Resour. Crop. Evol. 2018, 65, 2107–2134. [Google Scholar] [CrossRef]
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Sirsat, M.S.; Cernadas, E.; Fernández-Delgado, M.; Khan, R. Classification of agricultural soil parameters in India. Comput. Electron. Agric. 2017, 135, 269–279. [Google Scholar] [CrossRef]
- De Freitas, W.K.; Portz, A.; de Carvalho Peres, A.A.; Martinez Tarré, R.; de Melo Campos, M. Soil nutrient content and plant phytosociology in agroforestry systems of the Rio de Janeiro State highlands, Brazil. Acta Sci. Biol. Sci. 2018, 40, e35368. [Google Scholar] [CrossRef]
- Mansouri, M.; Dumont, B.; Leemans, V.; Destain, M.-F. Bayesian methods for predicting LAI and soil water Content. Precision. Agric. 2014, 15, 184–201. [Google Scholar] [CrossRef]
- Suchithra, M.S.; Pai, M.L. Improving the prediction accuracy of soil nutrient classification by optimizing extreme learning machine parameters. Inf. Process. Agric. 2019, 7, 72–82. [Google Scholar] [CrossRef]
- Braun-Blanquet, J.; Fitosociología, J. Bases para el Estudio de las Comunidades Vegetales; Ediciones Blume: Madrid, Spain, 1979. [Google Scholar]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Cano, E.; Cano-Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Ighbareyeh, J.M.H.; Leiva Gea, F.; del Río, S. Mitigating Climate Change Through Bioclimatic Applications and Cultivation Techniques in Agriculture (Andalusia, Spain). In Sustainable Agriculture, Forest and Environmental; Jhariya, M.K., Banerjee, A., Meena, R.S., Yadav, D.K., Eds.; Springer Nature: Singapore, 2019; pp. 31–69. [Google Scholar]
- Rivas-Martínez, S.; Díez, T.E.; Fernandez-González, F.; Izco, J.; Loidi, J.; Lousã, M.; Penas, A. Vascular Plant Communities of Spain and Portugal. Part II. Itinera Geobotánica 2002, 15, 433–922. [Google Scholar]
- Perrino, E.V.; Brunetti, G.; Farrag, K. Plant communities of multi-metal contaminated soils: A case study in National Park of Alta Murgia (Apulia Region—Southern Italy). Int. J. Phytoremed. 2014, 16, 871–888. [Google Scholar] [CrossRef]
- Bank, R.R. Intraspecific and interspecific pair-wise seedling competition between exotic annual grasses and native perennials: Plant-soil relationship. Plant Soil 2010, 326, 331–343. [Google Scholar] [CrossRef]
- Shiwakoti, S.; Zheljazkov, V.D.; Gollany, H.T.; Kleber, M.; Xing, B. Macronutrients in Soil and Wheat as Affected by a Long-Term Tillage and Nitrogen Fertilization in Winter Wheat–Fallow Rotation. Agronomy 2019, 9, 178. [Google Scholar] [CrossRef]
- Ordoñez, J.C.; Van Bodegom, P.M.; Witte, J.P.H.; Wright, I.J.; Reich, P.B.; Aerts, R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob. Ecol. Biogeogr. 2009, 18, 137–149. [Google Scholar] [CrossRef]
- López-Felices, B.; Aznar-Sánchez, J.A.; Velasco-Muñoz, J.F.; Piquer-Rodríguez Mª. Contribution of irrigation ponds to the sustainability of agriculture. A review of worldwide research. Sustainability 2020, 12, 5425. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, W.; Wang, L.; Liu, Y.; Feng, Q.; Fang, X.; Liu, Y. Distribution of Shrubland and Grassland Soil Erodibility on the Loess Plateau. Int. J. Environ. Res. Public Health 2018, 15, 1193. [Google Scholar] [CrossRef]
- Pastor, J.; Lacasta, C.; Hernández, A.J. Evaluación de las cubiertas vegetales en el olivar de una zona semiárida del centro de España. Edafología 2000, 7, 165–175. [Google Scholar]
- Jeong, J.; Zhang, X. Model Application for Sustainable Agricultural Water Use. Agronomy 2020, 10, 396. [Google Scholar] [CrossRef]
- Maffia, A.; Pergola, M.; Palese, A.M.; Celano, G. Environmental Impact Assessment of Organic vs. Integrated Olive-Oil Systems in Mediterranean Context. Agronomy 2020, 10, 416. [Google Scholar] [CrossRef]
- Peter, B.G.; Messina, J.P.; Lin, Z.; Snapp, S.S. Crop climate suitability mapping on the cloud: A geovisualization application for sustainable agriculture. Sci. Rep. 2020, 10, 15487. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and inductionof antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef]
- Sahib, M.R.; Pervaiz, Z.H.; Williams, M.A.; Saleem, M.; DeBolt, S. Rhizobacterial species richness improves sorghum growth and soil nutrient synergism in a nutrient-poor greenhouse soil. Sci. Rep. 2020, 10, 15454. [Google Scholar] [CrossRef]
- Igor, B.; Leon Josip, T.; Paulo, P. Agriculture Management Impacts on Soil Properties and Hydrological Response in Istria (Croatia). Agronomy 2020, 10, 282. [Google Scholar] [CrossRef]
- He, J.; Jin, Y.; Turner, N.C.; Li, F.-M. Irrigation during Flowering Improves Subsoil Water Uptake and Grain Yield in Rainfed Soybean. Agronomy 2020, 10, 120. [Google Scholar] [CrossRef]
- Caser, M.; Demasi, S.; Victorino, Í.M.M.; Donno, D.; Faccio, A.; Lumini, E.; Bianciotto, V.; Scariot, V. Arbuscular Mycorrhizal Fungi Modulate the Crop Performance and Metabolic Profile of Saffron in Soilless Cultivation. Agronomy 2019, 9, 232. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, C.; Teng, W.; Wang, J.; Lyu, X.; Dong, S.; Kang, S.; Gong, Z.; Ma, C. Accumulation and Distribution of Fertilizer Nitrogen and Nodule-Fixed Nitrogen in Soybeans with Dual Root Systems. Agronomy 2020, 10, 397. [Google Scholar] [CrossRef]
- Dhalaria, R.; Kumar, D.; Kumar, H.; Nepovimova, E.; Kuča, K.; Torequl Islam, M.; Verma, R. Arbuscular Mycorrhizal Fungi as Potential Agents in Ameliorating Heavy Metal Stress in Plants. Agronomy 2020, 10, 815. [Google Scholar] [CrossRef]
- Allito, B.B.; Ewusi-Mensah, N.; Logah, V. Legume-Rhizobium Strain Specificity Enhances Nutrition and Nitrogen Fixation in Faba Bean (Vicia faba L.). Agronomy 2020, 10, 826. [Google Scholar] [CrossRef]
- Kalembasa, S.; Szukała, J.; Faligowska, A.; Kalembasa, D.; Symanowicz, B.; Becher, M.; Gebus-Czupyt, B. Quantification of Biologically Fixed Nitrogen by White Lupin (Lupins albus L.) and Its Subsequent Uptake by Winter Wheat Using the 15N Isotope Dilution Method. Agronomy 2020, 10, 1392. [Google Scholar] [CrossRef]
- Nagaraj, K.; Vanishree, S.; Muthukumar, T. Genotypic variation in response and dependency of Cajanus cajan (L.) Millsp., on arbuscular mycorrhizal fungi in a tropical Alfisol. Plant Biosyst. 2020, 1–13. [Google Scholar] [CrossRef]
- Püschel, D.; Janoušková, M.; Voříšková, A.; Gryndlerová, H.; Vosátka, M.; Jansa, J. Arbuscular mycorrhiza stimulates biological nitrogen fixation in two Medicago spp. through improved phosphorus acquisition. Front Plant Sci. 2017, 8, 390. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; de Bruin, S.; Luckerhoff, L.; van Logtestijn, R.S.P.; Schlaeppi, K. A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plant nutrition and seedling recruitment. ISME J. 2016, 10, 389–399. [Google Scholar] [CrossRef]
- Jakubus, M.; Graczyk, M. Microelement Variability in Plants as an Effect of Sewage Sludge Compost Application Assessed by Different Statistical Methods. Agronomy 2020, 10, 642. [Google Scholar] [CrossRef]
- Liu, L.; Li, C.; Zhu, S.; Xu, Y.; Li, H.; Zheng, X.; Shi, R. Combined Application of Organic and Inorganic Nitrogen Fertilizers Affects Soil Prokaryotic Communities Compositions. Agronomy 2020, 10, 132. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Association | Dominant Species | Country | Locality | Date | Soil Type and pH | Pasture or Grassland Management | Reference |
|---|---|---|---|---|---|---|---|
| AcH | Hordeum leporinum | Spain | Jaén | Spring 2005 | Cambisols, basic | Pasturage | [22] |
| ArH | Hordeum leporinum | Portugal | Evora | Spring 2005 | Cambisols, acids and neutral | Tillage and herbicides | [17] |
| BH | Hordeum leporinum | Spain | Jaén | Spring 2005 | Regosols, acids | Pasturage | [22] |
| LR | Raphanus raphanistrum | Spain | Jaén | Spring 2005 | Cambisols, neutral | Tillage and herbicides | [17] |
| PD | Diplotaxis virgata | Spain | Jaén | Spring 2005 | Cambisols, basic and neutral | Tillage and herbicides | [17,26] |
| ArP | Papaver rhoeas | Portugal | Evora | Spring 2005 | Cambisols, neutral | Tillage and herbicides | [17] |
| ArChr | Glebionis coronaria | Portugal | Evora | Spring 2005 | Cambisols, neutral | Tillage and herbicides | [16] |
| RChr | Glebionis coronaria | Spain | Jaén | Spring 2005 | Cambisols, basic | Tillage and herbicides | [17,26] |
| UM | Malva neglecta | Spain | Jaén | Spring 2005 | Cambisols, indifferent pH | Tillage and herbicides | [17] |
| PA | Triticum vagans | Portugal | Evora | Spring 2005 | Regosols, neutral | Pasturage | [24] |
| TT | Taeniatherum caput-medusae | Spain | Jaén | Spring 2005 | Regosols, acids | Pasturage | [24] |
| TP | Plantago bellardii | Spain | Jaén | Spring 2005 | Regosols, acids | Pasturage | [17] |
| FS | Sinapis alba subsp. mairei | Spain | Jaén | Spring 2005 | Cambisols, basic | Tillage and herbicides | [17] |
| CS | Sylibum marianum | Spain | Jaén | Spring 2005 | Cambisols, basic and neutral | Tillage and herbicides | [17] |
| Plant Association | CEC | Nt | Pa | Mgc | Kc | %_Carb | %_Cl | %_ Sa | %_Si | pF 15 Atm | EC | pH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AcH | 0.577 | 0.007 | 1.595 | 0.217 | 0.386 | 2.641 | 3.078 | 2.190 | 3.829 | 2.435 | 0.090 | 0.048 |
| ArH | 0.664 | 0.012 | 2.242 | 0.277 | 0.035 | 2.716 | 1.843 | 2.935 | 1.780 | 0.659 | 0.079 | 0.145 |
| BH | 0.964 | 0.020 | 8.420 | 0.176 | 0.101 | 1.630 | 1.164 | 3.474 | 2.653 | 0.819 | 0.012 | 0.146 |
| LR | 0.688 | 0.004 | 0.713 | 0.123 | 0.021 | 0.572 | 1.941 | 2.633 | 1.549 | 0.664 | 0.148 | 0.218 |
| PD | 0.921 | 0.005 | 4.104 | 0.269 | 0.066 | 3.432 | 1.890 | 1.573 | 1.191 | 0.896 | 0.013 | 0.025 |
| ArP | 0.742 | 0.216 | 2.607 | 0.280 | 0.056 | 4.263 | 2.525 | 2.840 | 2.145 | 0.842 | 0.009 | 0.134 |
| ArChr | 0.989 | 0.034 | 3.295 | 0.201 | 0.149 | 2.223 | 1.954 | 3.381 | 1.733 | 1.104 | 0.012 | 0.054 |
| RChr | 1.023 | 0.031 | 8.558 | 0.306 | 0.445 | 2.869 | 2.967 | 3.303 | 3.048 | 1.325 | 0.102 | 0.036 |
| UM | 1.460 | 0.064 | 11.450 | 0.261 | 0.290 | 4.348 | 2.434 | 4.189 | 2.944 | 1.586 | 0.129 | 0.124 |
| PA | 0.678 | 0.009 | 1.842 | 0.240 | 0.029 | 2.034 | 2.245 | 2.511 | 2.124 | 0.819 | 0.007 | 0.091 |
| TT | 1.127 | 0.007 | 1.989 | 0.199 | 0.015 | 0.156 | 1.409 | 4.170 | 3.326 | 0.749 | 0.001 | 0.074 |
| TP | 0.343 | 0.005 | 2.160 | 0.058 | 0.010 | 0.091 | 1.087 | 1.678 | 1.735 | 0.355 | 0.001 | 0.056 |
| FS | 0.858 | 0.007 | 5.076 | 0.731 | 0.118 | 2.641 | 2.614 | 2.265 | 1.840 | 1.002 | 0.180 | 0.042 |
| CS | 1.007 | 0.024 | 6.531 | 0.202 | 0.224 | 2.303 | 2.050 | 2.605 | 2.158 | 0.724 | 0.063 | 0.074 |
| Plant Association | CEC | Nt | Pa | Mgc | Kc | %_Carb | %_Cl | %_Sa | %_Si | pF 15 Atm | EC | pH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AcH | 15.3 | 0.1 | 9.7 | 1.6 | 0.7 | 47.53 | 17.7 | 20.4 | 61.7 | 15.2 | 0.35 | 8.2 |
| ArH | 9.1 | 0.1 | 13.9 | 1.8 | 0.2 | 8.10 | 19.7 | 62.4 | 17.8 | 8.6 | 0.20 | 7.4 |
| BH | 10.5 | 0.1 | 15.4 | 1.0 | 0.3 | 4.49 | 14.5 | 54.2 | 31.2 | 8.2 | 0.12 | 7.4 |
| LR | 6.6 | 0.0 | 4.8 | 0.8 | 0.2 | 2.59 | 17.2 | 64.2 | 18.5 | 7.3 | 0.21 | 6.6 |
| PD | 14.3 | 0.0 | 15.3 | 2.3 | 1.0 | 49.28 | 40.0 | 19.9 | 40.0 | 19.1 | 0.28 | 8.0 |
| ArP | 10.8 | 0.1 | 14.7 | 1.8 | 0.4 | 12.38 | 25.3 | 45.6 | 28.9 | 13.0 | 0.16 | 7.6 |
| ArChr | 12.3 | 0.1 | 26.9 | 2.1 | 0.6 | 7.53 | 19.7 | 55.8 | 24.4 | 11.9 | 0.19 | 7.7 |
| RChr | 11.6 | 0.1 | 20.9 | 2.7 | 1.4 | 35.57 | 24.2 | 37.8 | 37.9 | 14.2 | 0.49 | 7.9 |
| UM | 10.8 | 0.1 | 36.1 | 1.6 | 1.2 | 31.96 | 21.2 | 46.0 | 32.7 | 13.1 | 0.56 | 7.7 |
| PA | 10.5 | 0.1 | 5.6 | 1.7 | 0.2 | 3.59 | 25.8 | 44.9 | 29.2 | 11.8 | 0.10 | 7.5 |
| TT | 9.6 | 0.0 | 5.1 | 1.0 | 0.1 | 1.64 | 13.5 | 64.9 | 21.4 | 6.6 | 0.04 | 6.1 |
| TP | 5.2 | 0.0 | 5.2 | 0.5 | 0.1 | 1.44 | 9.6 | 75.4 | 14.8 | 4.7 | 0.04 | 6.0 |
| FS | 12.2 | 0.0 | 13.1 | 2.8 | 0.8 | 52.18 | 35.1 | 24.2 | 40.6 | 18.7 | 0.68 | 8.0 |
| CS | 11.8 | 0.1 | 27.1 | 2.2 | 1.3 | 26.61 | 25.5 | 45.6 | 28.4 | 12.0 | 0.44 | 8.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano-Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Quinto-Canas, R.; del Río, S.; Cano, E. Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils. Agronomy 2021, 11, 1. https://doi.org/10.3390/agronomy11010001
Cano-Ortiz A, Musarella CM, Piñar Fuentes JC, Pinto Gomes CJ, Quinto-Canas R, del Río S, Cano E. Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils. Agronomy. 2021; 11(1):1. https://doi.org/10.3390/agronomy11010001
Chicago/Turabian StyleCano-Ortiz, Ana, Carmelo M. Musarella, José C. Piñar Fuentes, Carlos J. Pinto Gomes, Ricardo Quinto-Canas, Sara del Río, and Eusebio Cano. 2021. "Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils" Agronomy 11, no. 1: 1. https://doi.org/10.3390/agronomy11010001
APA StyleCano-Ortiz, A., Musarella, C. M., Piñar Fuentes, J. C., Pinto Gomes, C. J., Quinto-Canas, R., del Río, S., & Cano, E. (2021). Indicative Value of the Dominant Plant Species for a Rapid Evaluation of the Nutritional Value of Soils. Agronomy, 11(1), 1. https://doi.org/10.3390/agronomy11010001