Sprouts and Microgreens: Trends, Opportunities, and Horizons for Novel Research




Abstract
1. Introduction
2. Plant Species Scarcely Studied for Sprouting: Limits and Opportunities
2.1. Voluntary Species, Wild Relatives, Ancestors, Neglected/Local Accessions of Cultivated Species
2.2. Fruit Tree Species
3. Delving into Elicitors

{kind=link}
{kind=link}
| Elicitor | Application | Plant | Days after Sowing (DAS) | Phytochemicals/Bioactivities | Reference |
|---|---|---|---|---|---|
| Abiotic elicitation–Priming | |||||
| Seeds soaked with 30 mL of 50 mM KCl | Brassica oleracea var. capitata–cabbage | 10 | ↓total and single GLS | [83] | |
| Seeds soaked with 50 mM KCl | Brassica oleracea var. italica–broccoli | 10 | ≈total and single GLS and AA; ↑ flavonoids and TP | [84] | |
| 100% imbibition and aeration of the seeds for 24 h, with MeJA and JA (250 μM) and MET (10 mM) | Brassica oleracea var. italica–broccoli Raphanus sativus–radish | 8 | ↑ total GLS with MeJA and JA priming different effect on single GLS | [85] | |
| Physical factors | |||||
| Wounding | Soaked soybeans were wounded prior to germination stage by cutting the cotyledon individually | Glycine max–soybean | 10 | ↑glyceollin content (i.e., isoflavones) | [92] |
| Ultrasound | Seeds treated with ultrasound at 360 and 180 W for 30, 40, and 60 min in the ultrasonic bath | Phaseolus vulgaris–common bean | 1–4 | ↑ PAs, flavonoids, anthocyanins, and AA | [93] |
| Seeds treated with ultrasound at 100, 200, and 300 W for 30 min in the ultrasonic bath | Glycine max–soybean | 5 | ↑daidzein and genistein (i.e., isoflavones); ↓daidzein and genistein (i.e., isoflavones) | [95] | |
| Seeds subjected to ultrasound at three frequencies (28, 45 and 100 kHz) for 15, 20, and 30 min. | Arachis hypogaea–peanut | 1, 2, 3, 4, 5 | Generally, ↑ resveratrol content (i.e., stilbenes) | [94] | |
| Nanoparticles | Seeds sprinkled every day with NSePs (0, 10, 50 and 100 ppm); NSePs prepared by chemical reduction of NaHSeO3 solution with freshly prepared 0.25% glucose solution (4:1) | Brassica oleracea–broccoli | 9 | ≈ carotenes; ≈ phenolics and AA; different effects on single GLS | [98] |
| Magnetic field | Seeds exposed to 600 mT magnetic field pretreatment (Model: 2G of 755R magnetometer system) | Vigna radiata–mung bean | 6 | ↑anthocyanins | [101] |
| Microwave | Seeds exposed to microwave irradiation at 200, 400, 600, and 800 W for 10 or 30 s | Fagopyrum esculentum–tartary buckwheat | 3, 5, 7 | ↑total flavonoids and AA at 600 W | [102] |
| Clinorotation | Seeds treated with slow-rotating clinostat (CL) with a servo motor and an acrylic chamber. An amplifier controlled the rotation rate, and the chamber was horizontally rotated at 2 rpm | Vigna radiata–mung bean | 2, 3, 4 | ≈ carotenoids, except anthocyanins (↓) | [108] |
| Biotic elicitation | |||||
| Fungi | Rhizopus oryzae spore suspension (0.2 mL/g of beans) added to soybeans during incubation | Glycine max–soybean | 10 | ↑ total pterocarpans, coumestans (i.e., isoflavonoids); ≈ isoflavones | [92] |
| Rhizopus oryzae, Fusarium graminearum CBS 104.09 and Fusarium oxysporum CBS 186.53-fungal spore suspension, applied to 2-day-old seedlings | Sinapis alba–yellow mustard; Brassica napus–rapeseed; Brassica juncea–Chinese mustard | 7 | ≈ total GLS except with Fusarium oxysporum (↓) in yellow and Chinese mustard | [114] | |
| Spore suspension of Rhizopus oryzae (LU581) and Aspergillus oryzae var. effusus (LU009) (0.2 mL/g peanuts) added after soaking or after 2 days of germination | Arachis hypogaea–peanut | 5 or 7 | Effect on stilbenoids depending on the fungus and the time of application | [110] | |
| Rhizopus oryzae (LU 581) and Rhizopus oligosporus (approximately 1.5 × 107 CFU/g seed) added to 2-day-old seedlings | Vigna radiata–mung bean; Phaseolus vulgaris–common bean; Glycine max–soybean; Lupinus albus–white lupine; Lupinus angustifolius–blue lupine; Lupinus luteus–yellow lupine; Arachis hypogaea–peanut; | 8 | ↑ TP except in mung bean and blue lupin; different effects on phenolic subclasses (i.e., flavonoids, phenolic acids, stilbenoids, etc.) | [113] | |
| Spore suspension of Rhizopus oryzae (0.2 mL/g beans) added to 2-day-old seedlings | Phaseolus vulgaris–common bean; Phaseolus coccineus–runner bean; Lablab purpureus–Lablab bean; Vigna angularis–adzuki bean; Vigna unguiculata–cowpea; Vigna radiata–mung bean; Psophocarpus tetragonolobus–Winged bean | 7 | ↑or ≈all isoflavonoids (i.e., coumestans, isoflavans, pterocarpans, isoflavones, isoflavanones) | [111] | |
| Spore suspension of Rhizopus oryzae (0.2 mL/g beans) added to 2-day-old seedlings | Lupinus albus–white lupine; Lupinus angustifolius–blue lupine; Lupinus luteus–yellow lupine | 7 | ≈ or ↑genistein derivatives | [112] | |
| 4-day-old seedlings inoculated by soaking in spore solution of Rhizopus oryzae for 50 min | Triticum aestivum–soft wheat | 7 | ↑ variety of secondary metabolites mainly constituted by benzoxazin-3-one glycosides and benzoxazolinones | [115] | |
| Yeasts | Seeds soaked in distilled water or Lactobacillus plantarum 299v water suspension (1 × 108 CFU per 1 g of seeds (8.00 log10 CFU/g) for 4, 6, and 8 h | Lens culinaris–lentil; Glycine max–soybean; Vigna angularis–adzuki bean; Vigna radiata–mung bean | 4 | ≈ or ↓ phenolics and condensed tannins | [117] |
| Seeds soaked in water or Lactobacillus plantarum 299v water suspension (8.00 log10 CFU/g of seeds) | Lens culinaris–lentil; Glycine max–soybean; Vigna angularis–adzuki bean; Vigna radiata–mung bean | 4 | ≈ ability to quench ABTS•+ and •OH; different effects on the ability to quench O2•_;different effects on total and single phenolics | [119] | |
| Seeds soaked in distilled water or probiotic water suspension (1 × 106 CFU per 1 g of seeds) of Saccharomyces cerevisiae var. boulardii seedlings sprayed daily with 5 mL of Milli-Q water or 5 mL of probiotic water suspension on 1st day of cultivation (1 × 106 CFU per 1 g of seeds) | Lens culinaris–lentil; Vigna angularis–adzuki bean | 4 | ≈ phenolics and antioxidant activities | [118] | |
| Seeds soaked in distilled water or a Lactobacillus plantarum 299 V water suspension (1 × 108 CFU per 1 g of seeds) for 4 h | Glycine max–soybean | 4 | ↓ total isoflavones in fresh sprouts; different effects on single isoflavones; ↑ antioxidant activity | [125] | |
| Sprouts treated with 50, 100, 200, 400 and 800 mg/L of YPS (polysaccharide fraction of YE, Y4250) on days 0, 3, 6, and 9 | Fagopyrum esculentum–tartary buckwheat | 0, 3, 6, 9 | ↑ flavonoids in 3-,6-, and 9-day -old sprouts | [126] | |
| Bacteria | Seeds were surface-sterilized with sodium hypochlorite and rinsed with distilled water. Then, seeds were soaked in liquid suspension of endophytic bacterium, (ST-B2 strain, isolated from stem of buckwheat) at 0, 10% and 20% concentrations for 4 h at room temperature | Fagopyrum esculentum–buckwheat | 10 | Not analyzed | [120] |
| Bacillus megaterium 108 CFU/mL added 10 days before the cultivation of seeds, while Azotobacter chroococcum and Pseudomonas fluorescens 108–109 CFU/mL were added after pea sprouts emergence and after the 1st cut. Eight sub-treatments from different combinations of microbial inoculants were carried out | Pisum sativum–pea | 14 (1st cut) and 12 (2nd cut) | Not analyzed | [100] | |
| Seeds soaked for 30 min in 1:40 solutions of PGPs named Serenade ASO, containing Bacillus subtilis QST 713 at a concentration of 13.96 g/L (1.34%) | Fagopyrum esculentum–buckwheat | 14 | Not analyzed | [123] | |
| Algae | Seeds soaked for 30 min in PGPs: 1:100 solution of Kelpak SL (Ecklonia maxima extract), and 2 g in 500 mL of Polyversum WP (106 Pythium oligandrum oospores per 1 g) | Fagopyrum esculentum–buckwheat | 14 | Not analyzed | [123] |
| Other biostimulants | Seeds soaked for 30 min in 1:500 solution of PGPs named Asahi SL (mixture of sodium p-nitrophenolate, sodium o-nitrophenolate, sodium 5-nitroguaiacolate) | Fagopyrum esculentum–buckwheat | 14 | Not analyzed | [123] |
4. Sanitization and Processing
4.1. Microbiological Safety of Sprouts and Microgreens
4.2. Post-Harvest Shelf Life and Processing
| Disinfection Method | Disinfection Conditions | Plant Species | Pathogen/s | Treatment Effectiveness | Effects on Seed Germination | Effects on Sprouts Yield/Quality | References |
|---|---|---|---|---|---|---|---|
| Chemical | |||||||
| Bleach | 1 mL NaClO (4% v/v), 15 min | Fenugreek | Shiga toxin-producing Escherichia coli O104:H4 | On seeds: no detectable colonies | Not affected | Not affected | [181] |
| Acidified (organic acids) essential oil nanoemulsions | 50 mM acetic acid 50 mM levulinic acid carvacrol nanoemulsion, 4000/8000 ppm, 30 min | Mung bean, broccoli | Salmonella enterica Enteritidis cocktail | On seeds: 2–4 log CFU/g reduction–specie dependent; on sprouts: no detectable pathogens (<1.5 log CFU/g) | Not affected | Not affected | [182] |
| Chlorinated water | Treatment 1: seeds soaking, 100 ppm, 60 min; treatment 2: sprouts washing, 4 times/5 min each, 100 ppm | Mung bean | Shiga toxin-producing Escherichia coli Salmonella spp. Listeria monocytogenes | On seeds: 3.0 log CFU/g reduction; on sprouts: 7.0 log CFU/g reduction | -- | -- | [183] |
| Chlorine dioxide (ClO2) | 3 ppm ClO2, 10 min | Alfalfa | Listeria monocytogenes | On seeds: not effective; on sprouts: not effective | -- | -- | [184] |
| Chlorophyllin (Chl) | 1.5 × 10−5 M Chl, incubation time 1 h, 405 nm, radiant exposure: 18 J cm−2, 1 h | Wheat | Escherichia coli | On seeds: ∼1.5 log CFU/g. | -- | -- | [185] |
| Peracetic acid (PAA)-based solution | PAA solution (5% v/v) diluted in EtOH (20% v/v) 1:9 | Flax, chia | Salmonella spp. Enterococcus faecium | On seeds: 4.0–5.0 log CFU/g reduction–specie dependent | Not affected | -- | [186] |
| Saline organic acid solutions | Tempering solutions: lactic acid (5% v/v) and NaCl (~26% w/v) | Soft wheat, durum wheat | Salmonella enterica Escherichia coli O157:H7 Shiga toxin-producing Escherichia coli | On seeds: 1.6–2.6 log CFU/g – specie and pathogen dependent | -- | -- | [187] |
| Slightly acidic/acidic electrolyzed water (SAEW/AEW) | Available chorine concentrations (ACC) 150 mg/L, 24 h, 1.5 w/v | Brown rice | Bacillus cereus | On sprouts: 2.3 log CFU/g reduction | ↑↓ pH dependent | ↑↓ growth–pH dependent | [188] |
| SAEW | ACC 10.3-91.7 mg/L sprouts washing (1:3 w/v), 5 min | Buckwheat | Escherichia coli O78 Listeria monocytogenes | 1.1–2.7 log CFU/g reduction–pathogen dependent | Not affected | ↓ sprout length–ACC dependent | [189] |
| SAEW | ACC 70.0 mg/L seeds washing (1:5 w/v), 3 times | Pea | Natural population of total bacteria | ~ 2.0 log CFU/g reduction. | -- | Yield not affected; ↑ soluble sugars; ↓ nitrite | [190] |
| SAEW | ACC 25.0-45.0 mg/L seeds soaking (1:5 w/v), 0.5 h | Alfalfa | Natural Enterobacteriaceae | On seeds: ~ 2.5 log CFU/g reduction; on sprouts: 0.7–1.8 log CFU/g reduction | ↑ or not affected – pH dependent | Not affected | [191] |
| Zinc diacetate | Concentration 400 mg/L seeds soaking 0.66 h, 18 °C | Durum wheat | Natural bacteria population | On sprouts: 2.0 log CFU/g reduction | Slightly decreased | ↑ ash, lipids, proteins, zinc, vitamin C, total phenol content | [192] |
| Physical | |||||||
| Atmospheric pressure plasma discharge | Atmospheric pressure volumetric dielectric barrier discharge (DBD), 10 kHz, 8 KV, 500 ns pulses, 5/10 min | Alfalfa, onion, radish, cress | Escherichia coli | On seeds: 1.4–3.4 log CFU/g reduction–specie dependent | ↑↓ depending on temperature, time, pulse frequency or voltage | -- | [193] |
| Electron-beam irradiation | 12 kGy, 140 kV, 87 mm/s (12 kGy) and 250 mm/s (4 kGy) | Mung bean, clover, fenugreek | Escherichia coli | On seeds: not completely inactivated–specie dependent | Not affected | Not affected | [193] |
| Gamma ray irradiation | 7 kGy, 0.073 kGy/min, 4–5 °C | Mung bean, clover, fenugreek | Escherichia coli | On seeds: 5.0 log CFU/g reduction–specie dependent | ↓ species dependent | ↓ yield | [194] |
| Chlorine + drying + dry heat | ClO2-200 mg/mL, 5 min; 45 °C, 23% RH, 24 h; 80 °C, 23% RH, 48 h | Pack choi | Salmonella enterica Escherichia coli O157:H7 | On seeds: >3.8 log CFU/g reduction–pathogen dependent | Not affected | -- | [195] |
| Cold atmospheric pressure plasma | 400 W, 5 min | Lentil | Escherichia coli Salmonella spp. Listeria monocytogenes | On seeds: ~5.0 log CFU/g reduction–pathogen dependent | ↓ after 180 s | -- | [196] |
| High-intensity light pulses | 19.35 J/cm2, 15s | Chia | Salmonella Typhimurium | On seeds: 4.0 log CFU/g reduction. | -- | -- | [197] |
| Intense pulsed light | 24.50 and 20.81 J/cm2 – depending on plant species | Radish, pak choi | Natural total aerobic mesophilic bacteria | On seeds: 1.4–1.8 log CFU/g reduction–specie dependent | Not affected or slightly decreased | Sprout length not affected | [198] |
| Nonthermal plasma | Corona discharge plasma jet, 3 min | Radish | Aerobic bacteria Bacillus cereus Escherichia coli Salmonella spp. | On seeds: 1.2–2.1 log CFU/g reduction–pathogen dependent;on sprouts: >2.0 log CFU/g reduction–pathogen dependent | ↑↓ treatment time dependent | ↓↑ yield–treatment time (min)dependent; ↓ slightly brightness and redness; not affected reducing sugars, total phenolics, radical scavenging activity | [199] |
| Nonthermal plasma | Corona discharge plasma jet, 3 min | Rapeseed | Aerobic bacteria Bacillus cereus Escherichia coli Salmonella spp. | On seeds: 1.2–2.2 log CFU/g reduction–pathogen dependent; on sprouts: ~2.0 log CFU/g reduction–pathogen dependent | ↑↓ treatment time dependent | ↓↑ yield – treatment time dependent; not affected reducing sugars, total phenolics, radical scavenging activity; ↓ slightly “appearance”, “flavor”, “taste”, “overall acceptance”–treatment time dependent | [200] |
| Nonthermal plasma | Corona discharge plasma jet, 3 min | Broccoli | Aerobic bacteria Bacillus cereus Escherichia coli Salmonella spp. | On seeds: 1.2–2.3 log CFU/g reduction–pathogen dependent; on sprouts: ~2.0 log CFU/g reduction–pathogen dependent | ↑↓ treatment time dependent | ↓↑ yield – treatment time dependent; not affected reducing sugars, total phenolics, radical scavenging activity; ↑ slightly “flavor”; ↓ slightly “appearance”, “color”, “taste”, “overall acceptance”– treatment time dependent | [201] |
| Nonthermal plasma | Corona discharge plasma jet, 3 min | Pak choi | Aerobic bacteria Bacillus cereus Escherichia coli Salmonella spp. | On seeds: 1.3–2.1 log CFU/g reduction–pathogen dependent; on sprouts: ~2.0 log CFU/g reduction–pathogen dependent | ↑↓ treatment time dependent | ↓↑ yield – treatment time dependent; not affected reducing sugars, total phenolics, radical scavenging activity; ↑ slightly “texture”; ↓ slightly “appearance”, “taste”, “overall acceptance”–treatment time dependent | [202] |
| Ozone water | 2 ppm, 5 min | Alfalfa | Listeria monocytogenes | On seeds: not effective; on sprouts: not effective | -- | -- | [184] |
| Plasma-treated water (PTW) | 10 mL air-PTW | Mung bean | Natural population of total bacteria | On sprouts: 5.2 log CFU/g reduction | ↑ | ↑ superoxide dismutase (SOD) activity, indole acetic acid (IAA) content; ↓ malondialdehyde (MDA) and abscisic acid (ABA) contents. | [203] |
| Ozone | 2000 ppm, 4 h; 4000 ppm, 2 h | Wheat | Coliforms | On seeds: >1.5 log CFU/g reduction | Not tested | Minimum effect on antioxidant properties, lipid oxidation, moisture content | [204] |
| Ozone | 5 ppm, 20 min | Alfalfa | Salmonella spp. Shiga toxin-producing Escherichia coli | On seeds: 2.1 log CFU/g reduction; on sprouts: up to 3.6 log CFU/g reduction–pathogen dependent | Not affected | Not affected yield and color | [205] |
| Ultraviolet irradiation, krypton-chloride excilamp | 222-nm, 261 mJ/cm2 | Alfalfa | Salmonella Typhimurium Escherichia coli O157:H7 | On seeds: 2.8–3.0 log CFU/g reduction–pathogen dependent | ↓ slightly or not affected | -- | [206] |
| Chemical + physical | |||||||
| Dry heat + hydrogen peroxide | 80 °C, 24 h; H2O2 (30%) solution (2% v/v), 10 min | Alfalfa | Salmonella Typhimurium | On seeds: 1.7–3.6 log CFU/g reduction | ↑ | -- | [207] |
| Mild heat + hydrogen peroxide + acetic acid | 60 °C, 20 min; 4% (30% w/w in H2O), 15 min; 0.2% (5% distilled white vinegar), 15 min | Mung bean | Escherichia coli O157:H7 Listeria monocytogenes Salmonella enterica | On seeds: >4.0 log CFU/g reduction–pathogen dependent; on sprouts: not effective | Not affected | -- | [208] |
| Mild heat + hydrogen peroxide + AcOH sanitizing solution | 50 °C, 10 min; 2.0% H2O2 + 0.1% AcOH (Pure White Vinegar), 10 min | Alfalfa, radish | Escherichia coli O157:H7 Listeria monocytogenes Salmonella enterica | On seeds: 4.6–5.0 log CFU/g reduction – pathogen dependent | Not affected | Yield not affected | [209] |
| Biological | |||||||
| Bacteriocins from Enterococcus hirae (Eh9) | Eh9 - Lactic acid bacteria (LAB) cell-free supernatant (CFS), 0.5 g in 30 mL, 10 min | Butterhead lettuce | Escherichia coli O157:H7 | On seeds: 2.0 log CFU/g reduction | Not affected | ↑ slightly weight and sprouts length | [210] |
| Bacteriophages | Salmonella phage, seed soaking, 2 h, room temperature | Alfalfa | Salmonella enterica | On seeds: 2.5 log CFU/g reduction; on sprouting seeds: reduced without significance | Not affected | Not affected | [211] |
| Bacteriophages | Escherichia coli O104:H4 phage cocktail, (i) free phage application: 2 mL (2.5 × 1010 plaque-forming units (PFU)/mL), 1 h; (ii) impregnated phage application: filter paper in 7 cm diameter blotting with 6 mL (2.5 × 1010 PFU/mL); (iii) encapsulated phage application: seeds placed onto the paper coated with encapsulated phage in alginate beads. | Alfalfa | Escherichia coli O104:H4 | On seeds: below the limits of detection (< 1.0 log CFU/g); on sprouts: 1-log cycle reduction | -- | -- | [212] |
| Non-antagonistic bacterium | Erwinia persicina EUS78 strain, 108 colony-forming unit (CFU)/mL seed, dwelling | Alfalfa | Salmonella enterica | Not affected | Not affected | -- | [213] |
| Lactobacillus plantarum, Pediococcus acidilactici and Pediococcus pentosaceus mixture | 7 log CFU/g, 20 °C, 5 d | Alfalfa | Listeria monocytogenes Salmonella spp. | 1.0–4.5 log CFU/g reduction–depending on pathogen and initial concentration | Not affected | Not affected | [214] |
| Plant extracts | |||||||
| Benzyl isothiocyanate (BIT) | 1.5–2.0% (v/v), 15 min, 22 ± 2 °C | Alfalfa | Salmonella enterica serotypes | On seeds: >1.5 and 0.7 log CFU/g reduction–serovar dependent; on sprouts: 1.2–2.3 log CFU/g reduction–serovar dependent | Not affected | Not tested | [215] |
| Grapefruit seed extract | 0.05% aqueous solution | Lettuce | Natural population of total bacteria | Strong seed sterilization effects | Not affected | No effects on texture and color | [216] |
| Allspice, thyme and rosemary essential oils | Concentrations variable among species and pathogens | Alfalfa | Salmonella Typhimurium Listeria monocytogenes | Strong seed sterilization effects | Not affected | No effects on sensory properties | [217] |
| Savory, bay leaf and thyme hydrosols | 40 min | Wheat, lentil, mung bean | Salmonella Typhimurium Staphylococcus aureus | Strong seed sterilization effects–pathogen dependent | Not affected | -- | [218] |
5. Sprouts and Human Health: Rewards and Riddles
6. Antinutrients and Allergens in Sprouts
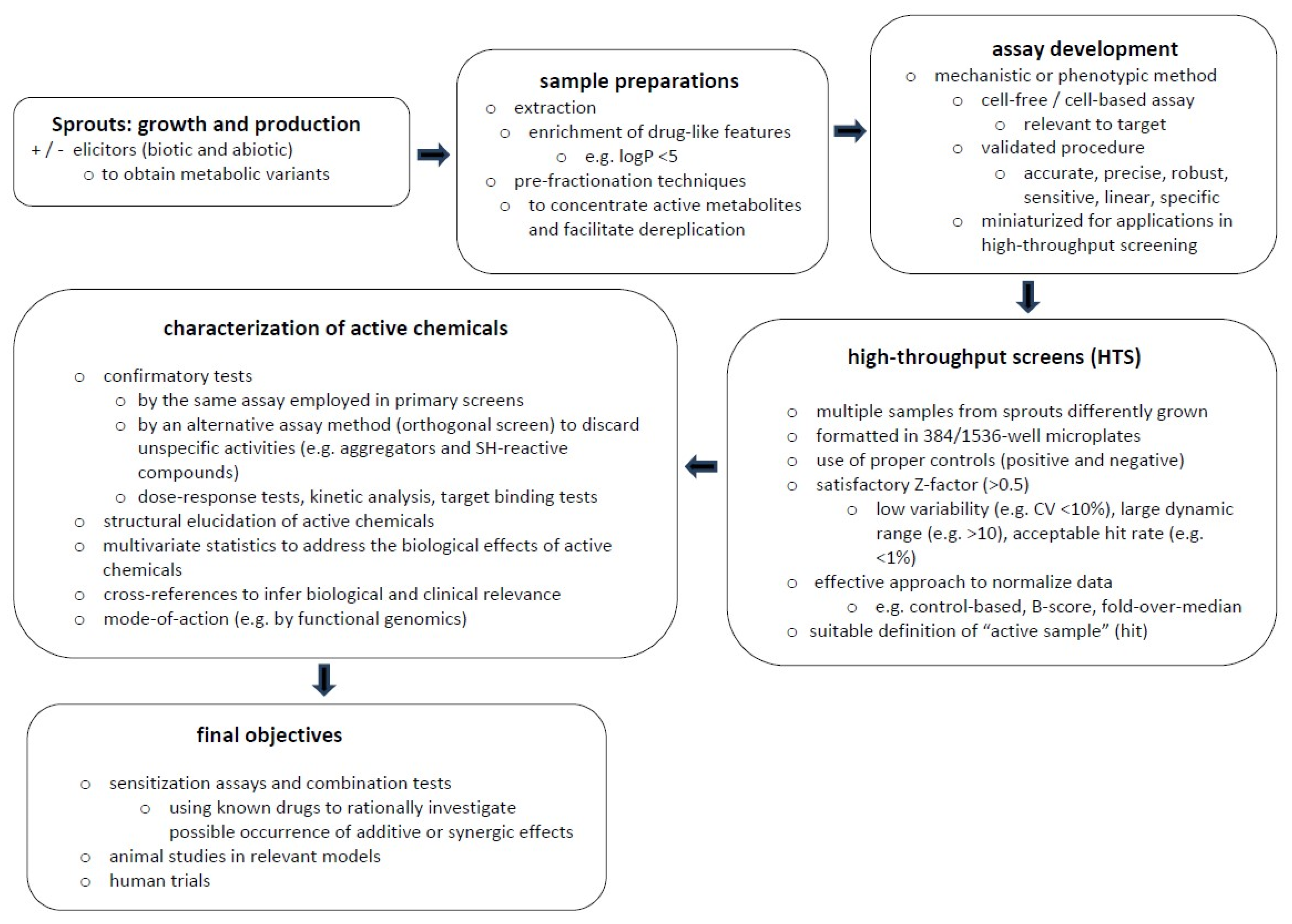
7. Sprouts: A Compelling Case for Drug Discovery?
8. Sprouts in Animal Feeding
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ebert, A.W. Sprouts, microgreens, and edible flowers: The potential for high value specialty produce in Asia. In Proceedings of the SEAVEG 2012 High Value Vegetables Southeast Asia Prod. Supply Demand, Chiang Mai, Thailand, 16 September 2013; pp. 216–227. [Google Scholar]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Di Gioia, F.; Renna, M.; Santamaria, P. Sprouts, Microgreens and “Baby Leaf” Vegetables BT-Minimally Processed Refrigerated Fruits and Vegetables; Yildiz, F., Wiley, R.C., Eds.; Springer US: Boston, MA, USA, 2017; pp. 403–432. ISBN 978-1-4939-7018-6. [Google Scholar]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted Grains: A Comprehensive Review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Gioia, F.D.; Leoni, B.; Mininni, C.; Santamaria, P. Culinary Assessment of Self-Produced Microgreens as Basic Ingredients in Sweet and Savory Dishes. J. Culin. Sci. Technol. 2017, 15, 126–142. [Google Scholar] [CrossRef]
- Di Gioia, F.; Mininni, C.; Santamaria, P. How to grow microgreens. In Microgreens: Microgreens: Novel Fresh and Functional Food to Explore All the Value of Biodiversity; Di Gioia, F., Santamaria, P., Eds.; ECO-logica: Bari, Italy, 2015; pp. 51–79. [Google Scholar]
- Mir, S.A.; Shah, M.A.; Mir, M.M. Microgreens: Production, shelf life, and bioactive components. Crit. Rev. Food Sci. Nutr. 2017, 57, 2730–2736. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Castellino, M.; Leoni, B.; Paradiso, V.M.; Santamaria, P. Microgreens Production with Low Potassium Content for Patients with Impaired Kidney Function. Nutrients 2018, 10, 675. [Google Scholar] [CrossRef]
- Renna, M.; Stellacci, A.M.; Corbo, F.; Santamaria, P. The Use of a Nutrient Quality Score is Effective to Assess the Overall Nutritional Value of Three Brassica Microgreens. Foods 2020, 9, 1226. [Google Scholar] [CrossRef]
- Ghoora, M.D.; Babu, D.R.; Srividya, N. Nutrient composition, oxalate content and nutritional ranking of ten culinary microgreens. J. Food Compos. Anal. 2020, 91, 103495. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Muzolf-Panek, M.; Goliński, P. Phenolic Content Changes in Plants Under Salt Stress BT–Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer New York: New York, NY, USA, 2013; pp. 283–314. ISBN 978-1-4614-4747-4. [Google Scholar]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of divergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense Against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Secondary metabolites and plant defense. Plant Physiol. 2006, 4, 315–344. [Google Scholar]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. CRC Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Peterson, J.; Dwyer, J.; Adlercreutz, H.; Scalbert, A.; Jacques, P.; McCullough, M.L. Dietary lignans: Physiology and potential for cardiovascular disease risk reduction. Nutr. Rev. 2010, 68, 571–603. [Google Scholar] [CrossRef] [PubMed]
- Beran, F.; Köllner, T.G.; Gershenzon, J.; Tholl, D. Chemical convergence between plants and insects: Biosynthetic origins and functions of common secondary metabolites. New Phytol. 2019, 223, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Tocopherols and tocotrienols in plants and their products: A review on methods of extraction, chromatographic separation, and detection. Food Res. Int. 2016, 82, 59–70. [Google Scholar] [CrossRef]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects–A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Harrath, A.H. Phytoestrogens and their effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Ogbe, R.J.; Ochalefu, D.O.; Mafulul, S.G.; Olaniru, O.B. A review on dietary phytosterols: Their occurrence, metabolism and health benefits. Asian J. Plant Sci. Res. 2015, 5, 10–21. [Google Scholar]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Paradiso, V.M. Ongoing Research on Microgreens: Nutritional Properties, Shelf-Life, Sustainable Production, Innovative Growing and Processing Approaches. Foods 2020, 9, 826. [Google Scholar] [CrossRef] [PubMed]
- Abellán, Á.; Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Sorting out the Value of Cruciferous Sprouts as Sources of Bioactive Compounds for Nutrition and Health. Nutrients 2019, 11, 429. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Falcinelli, B.; Tosti, G.; Bocci, L.; Benincasa, P. Taste quality traits and volatile profiles of sprouts and wheatgrass from hulled and non-hulled Triticum species. J. Food Biochem. 2019, 43, e12869. [Google Scholar] [CrossRef]
- Caracciolo, F.; El-Nakhel, C.; Raimondo, M.; Kyriacou, M.C.; Cembalo, L.; De Pascale, S.; Rouphael, Y. Sensory Attributes and Consumer Acceptability of 12 Microgreens Species. Agronomy 2020, 10, 1043. [Google Scholar] [CrossRef]
- Cooper, R. Re-discovering ancient wheat varieties as functional foods. J. Tradit. Complement. Med. 2015, 5, 138–143. [Google Scholar] [CrossRef]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef]
- Kumar, J.; Kaur, A.; Narang, P. Phytochemical screening and metal binding studies on floral extract of Solanum nigrum. Mater. Today Proc. 2020, 26, 3332–3336. [Google Scholar] [CrossRef]
- Nemzer, B.; Al-Taher, F.; Abshiru, N. Phytochemical composition and nutritional value of different plant parts in two cultivated and wild purslane (Portulaca oleracea L.) genotypes. Food Chem. 2020, 320, 126621. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Dias, M.I.; Pereira, C.; Calhelha, R.; Di Gioia, F.; Tzortzakis, N.; Ivanov, M.; Sokovic, M.; Barros, L.; et al. Wild and Cultivated Centaurea raphanina subsp. mixta: A Valuable Source of Bioactive Compounds. Antioxidants 2020, 9, 314. [Google Scholar] [CrossRef]
- Harouna, D.V.; Venkataramana, P.B.; Ndakidemi, P.A.; Matemu, A.O. Under-exploited wild Vigna species potentials in human and animal nutrition: A review. Glob. Food Sec. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Montevecchi, G.; Setti, L.; Olmi, L.; Buti, M.; Laviano, L.; Antonelli, A.; Sgarbi, E. Determination of Free Soluble Phenolic Compounds in Grains of Ancient Wheat Varieties (Triticum sp. pl.) by Liquid Chromatography–Tandem Mass Spectrometry. J. Agric. Food Chem. 2019, 67, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Gaba, S.; Perronne, R.; Fried, G.; Gardarin, A.; Bretagnolle, F.; Biju-Duval, L.; Colbach, N.; Cordeau, S.; Fernández-Aparicio, M.; Gauvrit, C.; et al. Response and effect traits of arable weeds in agro-ecosystems: A review of current knowledge. Weed Res. 2017, 57, 123–147. [Google Scholar] [CrossRef]
- Butnariu, M.; Samfira, I. Vegetal Metabolomics to Seeds of Galium Aparine. J. Bioequiv. Availab. 2013, 5, e45. [Google Scholar] [CrossRef]
- Abbas, M.N.; Rana, S.A.; Shahid, M.; Rana, N.; Mahmood-ul-Hassan, M.; Hussain, M. Chemical evaluation of weed seeds mixed with wheat grains at harvest. J. Anim. Plant Sci. 2012, 22, 283–288. [Google Scholar]
- Bano, Y.; Ahmad, S.; Alam, S.P. Study of seed germination behaviour of Solanum nigrum L. J. Indian Bot. Soc. 2019, 98, 85–88. [Google Scholar] [CrossRef]
- Senila, L.; Neag, E.; Cadar, O.; Kovacs, M.H.; Becze, A.; Senila, M. Chemical, Nutritional and Antioxidant Characteristics of Different Food Seeds. Appl. Sci. 2020, 10, 1589. [Google Scholar] [CrossRef]
- Benincasa, P.; Galieni, A.; Manetta, A.C.; Pace, R.; Guiducci, M.; Pisante, M.; Stagnari, F. Phenolic compounds in grains, sprouts and wheatgrass of hulled and non-hulled wheat species. J. Sci. Food Agric. 2015, 95, 1795–1803. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Liu, R.; Zhu, H.; Draves, J.; Marcone, M.; Sun, Y.; Tsao, R. Characterization of phenolics, betacyanins and antioxidant activities of the seed, leaf, sprout, flower and stalk extracts of three Amaranthus species. J. Food Compos. Anal. 2015, 37, 75–81. [Google Scholar] [CrossRef]
- Paśko, P.; Bartoń, H.; Zagrodzki, P.; Gorinstein, S.; Fołta, M.; Zachwieja, Z. Anthocyanins, total polyphenols and antioxidant activity in amaranth and quinoa seeds and sprouts during their growth. Food Chem. 2009, 115, 994–998. [Google Scholar] [CrossRef]
- Aphalo, P.; Martínez, E.N.; Añón, M.C. Amaranth Sprouts: A Potential Health Promoting and Nutritive Natural Food. Int. J. Food Prop. 2015, 18, 2688–2698. [Google Scholar] [CrossRef][Green Version]
- Lau, T.Q.; Tang, V.T.H.; Kansedo, J. Influence of Soil and Light Condition on the Growth and Antioxidants Content of Amaranthus Cruentus (Red Amaranth) Microgreen. IOP Conf. Ser. Mater. Sci. Eng. 2019, 495, 12051. [Google Scholar] [CrossRef]
- Jan, R.; Saxena, D.C.; Singh, S. Physico-chemical, textural, sensory and antioxidant characteristics of gluten–Free cookies made from raw and germinated Chenopodium (Chenopodium album) flour. LWT Food Sci. Technol. 2016, 71, 281–287. [Google Scholar] [CrossRef]
- Jan, R.; Saxena, D.C.; Singh, S. Effect of extrusion variables on antioxidant activity, total phenolic content and dietary fibre content of gluten-free extrudate from germinated Chenopodium (Chenopodium album) flour. Int. J. Food Sci. Technol. 2017, 52, 2623–2630. [Google Scholar] [CrossRef]
- Jan, R.; Saxena, D.C.; Singh, S. Comparative study of raw and germinated Chenopodium (Chenopodium album) flour on the basis of thermal, rheological, minerals, fatty acid profile and phytocomponents. Food Chem. 2018, 269, 173–180. [Google Scholar] [CrossRef]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Laus, M.; Cataldi, M.; Soccio, M.; Alfarano, M.; Amodio, M.; Colelli, G.; Flagella, Z.; Pastore, D. Effect of germination and sprout storage on antioxidant capacity and phenolic content in quinoa (Chenopodium quinoa Willd.). In Proceedings of the SIBV-SIGA Joint Congress Sustainability of agricultural environment: Contributions of Plant Genetics and Physiology, Pisa, Italy, 19–22 September 2017. [Google Scholar]
- Fischer, S.; Wilckens, R.; Jara, J.; Aranda, M.; Valdivia, W.; Bustamante, L.; Graf, F.; Obal, I. Protein and antioxidant composition of quinoa (Chenopodium quinoa Willd.) sprout from seeds submitted to water stress, salinity and light conditions. Ind. Crops Prod. 2017, 107, 558–564. [Google Scholar] [CrossRef]
- Lazo-Vélez, M.A.; Guajardo-Flores, D.; Mata-Ramírez, D.; Gutiérrez-Uribe, J.A.; Serna-Saldivar, S.O. Characterization and Quantitation of Triterpenoid Saponins in Raw and Sprouted Chenopodium berlandieri spp. (Huauzontle) Grains Subjected to Germination with or without Selenium Stress Conditions. J. Food Sci. 2016, 81, C19–C26. [Google Scholar] [CrossRef]
- Harakotr, B.; Srijunteuk, S.; Rithichai, P.; Tabunhan, S. Effects of Light-Emitting Diode Light Irradiance Levels on Yield, Antioxidants and Antioxidant Capacities of Indigenous Vegetable Microgreens. Sci. Technol. Asia 2019, 24, 59–66. [Google Scholar]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Palladino, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Phenolic Constitution, Phytochemical and Macronutrient Content in Three Species of Microgreens as Modulated by Natural Fiber and Synthetic Substrates. Antioxidants 2020, 9, 252. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Enache, I.-M.; Livadariu, O. Preliminary results regarding the testing of treatments with lightemitting diode (LED) on the seed germination of Artemisia dracunculus L. Sci. Bull. Ser. F. Biotechnol. 2016, 20, 51–56. [Google Scholar]
- Lenzi, A.; Orlandini, A.; Bulgari, R.; Ferrante, A.; Bruschi, P. Antioxidant and Mineral Composition of Three Wild Leafy Species: A Comparison Between Microgreens and Baby Greens. Foods 2019, 8, 487. [Google Scholar] [CrossRef] [PubMed]
- Pająk, P.; Socha, R.; Broniek, J.; Królikowska, K.; Fortuna, T. Antioxidant properties, phenolic and mineral composition of germinated chia, golden flax, evening primrose, phacelia and fenugreek. Food Chem. 2019, 275, 69–76. [Google Scholar] [CrossRef]
- Paradiso, V.M.; Castellino, M.; Renna, M.; Gattullo, C.E.; Calasso, M.; Terzano, R.; Allegretta, I.; Leoni, B.; Caponio, F.; Santamaria, P. Nutritional characterization and shelf-life of packaged microgreens. Food Funct. 2018, 9, 5629–5640. [Google Scholar] [CrossRef]
- Bennett, R.N.; Carvalho, R.; Mellon, F.A.; Eagles, J.; Rosa, E.A.S. Identification and Quantification of Glucosinolates in Sprouts Derived from Seeds of Wild Eruca sativa L. (Salad Rocket) and Diplotaxis tenuifolia L. (Wild Rocket) from Diverse Geographical Locations. J. Agric. Food Chem. 2007, 55, 67–74. [Google Scholar] [CrossRef]
- Loedolff, B.; Brooks, J.; Stander, M.; Peters, S.; Kossmann, J. High light bio-fortification stimulates de novo synthesis of resveratrol in Diplotaxis tenuifolia (wild rocket) micro-greens. Funct. Foods Heal. Dis. 2017, 7, 859–872. [Google Scholar] [CrossRef]
- Xu, L.; Wang, P.; Ali, B.; Yang, N.; Chen, Y.; Wu, F.; Xu, X. Changes of the phenolic compounds and antioxidant activities in germinated adlay seeds. J. Sci. Food Agric. 2017, 97, 4227–4234. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, L.; Wang, X.; Gu, Z.; Beta, T. Changes of phenolic profiles and antioxidant activity in canaryseed (Phalaris canariensis L.) during germination. Food Chem. 2016, 194, 608–618. [Google Scholar] [CrossRef]
- Falcinelli, B.; Benincasa, P.; Calzuola, I.; Gigliarelli, L.; Lutts, S.; Marsili, V. Phenolic Content and Antioxidant Activity in Raw and Denatured Aqueous Extracts from Sprouts and Wheatgrass of Einkorn and Emmer Obtained under Salinity. Molecules 2017, 22, 2132. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; D’Egidio, S.; Falcinelli, B.; Pagnani, G.; Pace, R.; Pisante, M.; Benincasa, P. Effects of sprouting and salt stress on polyphenol composition and antiradical activity of einkorn, emmer and durum wheat. Ital. J. Agron. 2017, 12. [Google Scholar] [CrossRef]
- Benincasa, P.; Tosti, G.; Farneselli, M.; Maranghi, S.; Bravi, E.; Marconi, O.; Falcinelli, B.; Guiducci, M. Phenolic content and antioxidant activity of einkorn and emmer sprouts and wheatgrass obtained under different radiation wavelengths. Ann. Agric. Sci. 2020. [Google Scholar] [CrossRef]
- Chu, C.; Yan, N.; Du, Y.; Liu, X.; Chu, M.; Shi, J.; Zhang, H.; Liu, Y.; Zhang, Z. iTRAQ-based proteomic analysis reveals the accumulation of bioactive compounds in Chinese wild rice (Zizania latifolia) during germination. Food Chem. 2019, 289, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Sritongtae, B.; Sangsukiam, T.; Morgan, M.R.A.; Duangmal, K. Effect of acid pretreatment and the germination period on the composition and antioxidant activity of rice bean (Vigna umbellata). Food Chem. 2017, 227, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Peykarestan, B.; Seify, M.; Shoukat-Fadaei, M.; Hatim, M. UV irradiation effects on seed germination and growth, protein content, peroxidase and protease activity in Portulaca grandiflora and Portula caoleracea. World Appl. Sci. J. 2012, 17, 802–808. [Google Scholar]
- Gunenc, A.; Rowland, O.; Xu, H.; Marangoni, A.; Hosseinian, F. Portulaca oleracea seeds as a novel source of alkylresorcinols and its phenolic profiles during germination. LWT 2019, 101, 246–250. [Google Scholar] [CrossRef]
- Villacís-Chiriboga, J.; Elst, K.; Van Camp, J.; Vera, E.; Ruales, J. Valorization of byproducts from tropical fruits: Extraction methodologies, applications, environmental, and economic assessment: A review (Part 1: General overview of the byproducts, traditional biorefinery practices, and possible applications). Compr. Rev. Food Sci. Food Saf. 2020, 19, 405–447. [Google Scholar] [CrossRef]
- Montoya, C.; GonzÃ!`lez, L.; Pulido, S.; AtehortÃ\textordmasculinea, L.; Robledo, S.M. Identification and quantification of limonoid aglycones content of Citrus seeds. Rev. Bras. Farmacogn. 2019, 29, 710–714. [Google Scholar] [CrossRef]
- Silva, S.; Gomes, L.; Leitão, F.; Bronze, M.; Coelho, A.V.; Boas, L.V. Secoiridoids in olive seed: Characterization of nüzhenide and 11-methyl oleosides by liquid chromatography with diode array and mass spectrometry. Grasas y aceites 2010, 61, 157–164. [Google Scholar] [CrossRef]
- González-Hidalgo, I.; Bañón, S.; Ros, J.M. Evaluation of table olive by-product as a source of natural antioxidants. Int. J. Food Sci. Technol. 2012, 47, 674–681. [Google Scholar] [CrossRef]
- Weidner, S.; Brosowska-Arendt, W.; Szczechura, W.; Karamac, M.; Kosinska, A.; Amarowicz, R. Effect of osmotic stress and post-stress recovery on the content of phenolics and properties of antioxidants in germinating seeds of grapevine Vitis californica. Acta Soc. Bot. Pol. 2011, 80, 11–19. [Google Scholar] [CrossRef]
- Weidner, S.; Chrzanowski, S.; Karamać, M.; Król, A.; Badowiec, A.; Mostek, A.; Amarowicz, R. Analysis of Phenolic Compounds and Antioxidant Abilities of Extracts from Germinating Vitis californica Seeds Submitted to Cold Stress Conditions and Recovery after the Stress. Int. J. Mol. Sci. 2014, 15, 16211–16225. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, B.; Marconi, O.; Maranghi, S.; Lutts, S.; Rosati, A.; Famiani, F.; Benincasa, P. Effect of Genotype on the Sprouting of Pomegranate (Punica granatum L.) Seeds as a Source of Phenolic Compounds from Juice Industry by-Products. Plant Foods Hum. Nutr. 2017, 72, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, B.; Maranghi, S.; Paoletti, A.; Marconi, O.; Rosati, A.; Famiani, F.; Benincasa, P. Sprouting olive (Olea europaea L.) seeds as a source of antioxidants from residual whole stones. Sci. Hortic. 2018, 240, 558–560. [Google Scholar] [CrossRef]
- Falcinelli, B.; Famiani, F.; Paoletti, A.; D’Egidio, S.; Stagnari, F.; Galieni, A.; Benincasa, P. Phenolic Compounds and Antioxidant Activity of Sprouts from Seeds of Citrus Species. Agriculture 2020, 10, 33. [Google Scholar] [CrossRef]
- Liu, H.; Kang, Y.; Zhao, X.; Liu, Y.; Zhang, X.; Zhang, S. Effects of elicitation on bioactive compounds and biological activities of sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current Knowledge on Selenium Biofortification to Improve the Nutraceutical Profile of Food: A Comprehensive Review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef]
- Hassini, I.; Baenas, N.; Moreno, D.A.; Carvajal, M.; Boughanmi, N.; Martinez Ballesta, M.D.C. Effects of seed priming, salinity and methyl jasmonate treatment on bioactive composition of Brassica oleracea var. capitata (white and red varieties) sprouts. J. Sci. Food Agric. 2017, 97, 2291–2299. [Google Scholar] [CrossRef]
- Hassini, I.; Martinez-Ballesta, M.C.; Boughanmi, N.; Moreno, D.A.; Carvajal, M. Improvement of broccoli sprouts (Brassica oleracea L. var. italica) growth and quality by KCl seed priming and methyl jasmonate under salinity stress. Sci. Hortic. 2017, 226, 141–151. [Google Scholar] [CrossRef]
- Baenas, N.; Villaño, D.; García-Viguera, C.; Moreno, D.A. Optimizing elicitation and seed priming to enrich broccoli and radish sprouts in glucosinolates. Food Chem. 2016, 204, 314–319. [Google Scholar] [CrossRef]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed priming: New comprehensive approaches for an old empirical technique. New Chall. Seed Biol. Transl. Res. Driv. Seed Technol. 2016, 1–46. [Google Scholar] [CrossRef]
- Benincasa, P.; D’Amato, R.; Falcinelli, B.; Troni, E.; Fontanella, M.C.; Frusciante, S.; Guiducci, M.; Beone, G.M.; Businelli, D.; Diretto, G. Grain Endogenous Selenium and Moderate Salt Stress Work as Synergic Elicitors in the Enrichment of Bioactive Compounds in Maize Sprouts. Agronomy 2020, 10, 735. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Rosellini, I.; Pezzarossa, B. Production of selenium-biofortified microgreens from selenium-enriched seeds of basil. J. Sci. Food Agric. 2019, 99, 5601–5605. [Google Scholar] [CrossRef] [PubMed]
- Miryeganeh, M.; Saze, H. Epigenetic inheritance and plant evolution. Popul. Ecol. 2020, 62, 17–27. [Google Scholar] [CrossRef]
- Marconi, G.; Pace, R.; Traini, A.; Raggi, L.; Lutts, S.; Chiusano, M.; Guiducci, M.; Falcinelli, M.; Benincasa, P.; Albertini, E. Use of MSAP Markers to Analyse the Effects of Salt Stress on DNA Methylation in Rapeseed (Brassica napus var. oleifera). PLoS ONE 2013, 8, e75597. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.L.; Torres-Pacheco, I. Plant Hormesis Management with Biostimulants of Biotic Origin in Agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef] [PubMed]
- Aisyah, S.; Gruppen, H.; Madzora, B.; Vincken, J.-P. Modulation of Isoflavonoid Composition of Rhizopus oryzae Elicited Soybean (Glycine max) Seedlings by Light and Wounding. J. Agric. Food Chem. 2013, 61, 8657–8667. [Google Scholar] [CrossRef]
- Ampofo, J.O.; Ngadi, M. Ultrasonic assisted phenolic elicitation and antioxidant potential of common bean (Phaseolus vulgaris) sprouts. Ultrason. Sonochem. 2020, 64, 104974. [Google Scholar] [CrossRef]
- Yu, M.; Liu, H.; Shi, A.; Liu, L.; Wang, Q. Preparation of resveratrol-enriched and poor allergic protein peanut sprout from ultrasound treated peanut seeds. Ultrason. Sonochem. 2016, 28, 334–340. [Google Scholar] [CrossRef]
- Yang, H.; Gao, J.; Yang, A.; Chen, H. The ultrasound-treated soybean seeds improve edibility and nutritional quality of soybean sprouts. Food Res. Int. 2015, 77, 704–710. [Google Scholar] [CrossRef]
- Cai, W.; He, H.; Zhu, S.; Wang, N. Biological Effect of Audible Sound Control on Mung Bean (Vigna radiate) Sprout. Biomed Res. Int. 2014, 2014, 931740. [Google Scholar] [CrossRef] [PubMed]
- Zuverza-Mena, N.; Armendariz, R.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Effects of Silver Nanoparticles on Radish Sprouts: Root Growth Reduction and Modifications in the Nutritional Value. Front. Plant Sci. 2016, 7, 90. [Google Scholar] [CrossRef] [PubMed]
- Vicas, S.I.; Cavalu, S.; Laslo, V.; Tocai, M.; Costea, T.O.; Moldovan, L. Growth, Photosynthetic Pigments, Phenolic, Glucosinolates Content and Antioxidant Capacity of Broccoli Sprouts in Response to Nanoselenium Particles Supply. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 821–828. [Google Scholar] [CrossRef]
- Mhamdi, A.; Noctor, G. High CO2 Primes Plant Biotic Stress Defences through Redox-Linked Pathways. Plant Physiol. 2016, 172, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.H.; Zayed, M.S.; Hassanein, M.K.; Abdallah, M.M.F. Green pea sprout response to microbial inoculation and increasing atmospheric CO2 concentration. Arab. Univ. J. Agric. Sci. 2018, 26, 2513–2523. [Google Scholar] [CrossRef]
- Chen, Y.-P.; He, J.-M.; Li, R. Effects of magnetic fields pretreatment of mungbean seeds on sprout yield and quality. Afr. J. Biotechnol. 2012, 11, 8932–8937. [Google Scholar]
- Wang, S.; Wang, J.; Guo, Y. Microwave irradiation enhances the germination rate of tartary buckwheat and content of some compounds in its sprouts. Polish J. food Nutr. Sci. 2018, 68, 195–205. [Google Scholar] [CrossRef]
- Kumar, S.; Gautam, S. A combination process to ensure microbiological safety, extend storage life and reduce anti-nutritional factors in legume sprouts. Food Biosci. 2019, 27, 18–29. [Google Scholar] [CrossRef]
- Alrifai, O.; Hao, X.; Marcone, M.F.; Tsao, R. Current Review of the Modulatory Effects of LED Lights on Photosynthesis of Secondary Metabolites and Future Perspectives of Microgreen Vegetables. J. Agric. Food Chem. 2019, 67, 6075–6090. [Google Scholar] [CrossRef]
- Bartucca, M.L.; Del Buono, D.; Ballerini, E.; Benincasa, P.; Falcinelli, B.G.M. Effect of light spectrum on gas exchange, growth and biochemical characteristics of einkorn seedlings. Agronomy 2020, 10, 1042. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-Specific Modulatory Effects of Select Spectral Bandwidths on the Nutritive and Phytochemical Composition of Microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; De Pascale, S.; Kyratzis, A.; Rouphael, Y. Microgreens as a Component of Space Life Support Systems: A Cornucopia of Functional Food. Front. Plant Sci. 2017, 8, 1587. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Shiraga, K.; Suzuki, T.; Kondo, N.; Ogawa, Y. Chlorophyll, Carotenoid and Anthocyanin Accumulation in Mung Bean Seedling Under Clinorotation. Microgravity Sci. Technol. 2017, 29, 427–432. [Google Scholar] [CrossRef]
- Falcinelli, B.; Sileoni, V.; Marconi, O.; Perretti, G.; Quinet, M.; Lutts, S.; Benincasa, P. Germination under Moderate Salinity Increases Phenolic Content and Antioxidant Activity in Rapeseed (Brassica napus var oleifera Del.) Sprouts. Molecules 2017, 22, 1377. [Google Scholar] [CrossRef]
- Aisyah, S.; Gruppen, H.; Slager, M.; Helmink, B.; Vincken, J.-P. Modification of Prenylated Stilbenoids in Peanut (Arachis hypogaea) Seedlings by the Same Fungi That Elicited Them: The Fungus Strikes Back. J. Agric. Food Chem. 2015, 63, 9260–9268. [Google Scholar] [CrossRef] [PubMed]
- Aisyah, S.; Gruppen, H.; Andini, S.; Bettonvil, M.; Severing, E.; Vincken, J.-P. Variation in accumulation of isoflavonoids in Phaseoleae seedlings elicited by Rhizopus. Food Chem. 2016, 196, 694–701. [Google Scholar] [CrossRef]
- Aisyah, S.; Vincken, J.-P.; Andini, S.; Mardiah, Z.; Gruppen, H. Compositional changes in (iso)flavonoids and estrogenic activity of three edible Lupinus species by germination and Rhizopus-elicitation. Phytochemistry 2016, 122, 65–75. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; den Besten, H.M.W.; Aisyah, S.; Gruppen, H.; Vincken, J.-P. The position of prenylation of isoflavonoids and stilbenoids from legumes (Fabaceae) modulates the antimicrobial activity against Gram positive pathogens. Food Chem. 2017, 226, 193–201. [Google Scholar] [CrossRef]
- Andini, S.; Dekker, P.; Gruppen, H.; Araya-Cloutier, C.; Vincken, J.-P. Modulation of Glucosinolate Composition in Brassicaceae Seeds by Germination and Fungal Elicitation. J. Agric. Food Chem. 2019, 67, 12770–12779. [Google Scholar] [CrossRef]
- de Bruijn, W.J.C.; Vincken, J.-P.; Duran, K.; Gruppen, H. Mass Spectrometric Characterization of Benzoxazinoid Glycosides from Rhizopus-Elicited Wheat (Triticum aestivum) Seedlings. J. Agric. Food Chem. 2016, 64, 6267–6276. [Google Scholar] [CrossRef]
- Shalaby, S.; Horwitz, B.A. Plant phenolic compounds and oxidative stress: Integrated signals in fungal–plant interactions. Curr. Genet. 2015, 61, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Swieca, M.; Gawlik-Dziki, U.; Jakubczyk, A.; Bochnak, J.; Sikora, M.; Suliburska, J. Nutritional quality of fresh and stored legumes sprouts – Effect of Lactobacillus plantarum 299v enrichment. Food Chem. 2019, 288, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Swieca, M.; Kordowska-Wiater, M.; Pytka, M.; Gawlik-Dziki, U.; Seczyk, L.; Złotek, U.; Kapusta, I. Nutritional and pro-health quality of lentil and adzuki bean sprouts enriched with probiotic yeast Saccharomyces cerevisiae var. boulardii. LWT 2019, 100, 220–226. [Google Scholar] [CrossRef]
- Złotek, U.; Świeca, M.; Reguła, J.; Jakubczyk, A.; Sikora, M.; Gawlik-Dziki, U.; Kapusta, I. Effects of probiotic L. plantarum 299v on consumer quality, accumulation of phenolics, antioxidant capacity and biochemical changes in legume sprouts. Int. J. Food Sci. Technol. 2019, 54, 2437–2446. [Google Scholar] [CrossRef]
- Briatia, X.; Jomduang, S.; Park, C.H.; Lumyong, S.; Kanpiengjai, A.; Khanongnuch, C. Enhancing growth of buckwheat sprouts and microgreens by endophytic bacterium inoculation. Int. J. Agric. Biol. 2017, 19, 374–380. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Witkowicz, R.; Biel, W.; Chłopicka, J.; Galanty, A.; Gleń-Karolczyk, K.; Skrzypek, E.; Krupa, M. Biostimulants and Microorganisms Boost the Nutritional Composition of Buckwheat (Fagopyrum esculentum Moench) Sprouts. Agronomy 2019, 9, 469. [Google Scholar] [CrossRef]
- Das, A.; Lee, S.-H.; Hyun, T.K.; Kim, S.-W.; Kim, J.-Y. Plant volatiles as method of communication. Plant Biotechnol. Rep. 2013, 7, 9–26. [Google Scholar] [CrossRef]
- Świeca, M.; Gawlik-Dziki, U.; Złotek, U.; Kapusta, I.; Kordowska-Wiater, M.; Baraniak, B. Effect of cold storage on the potentially bioaccessible isoflavones and antioxidant activities of soybean sprouts enriched with Lactobacillus plantarum 299v. LWT 2020, 118, 108820. [Google Scholar] [CrossRef]
- Zhao, J.L.; Zou, L.; Zhong, L.Y.; Peng, L.X.; Ying, P.L.; Tan, M.L.; Zhao, G. Effects of polysaccharide elicitors from endophytic Bionectria pityrodes Fat6 on the growth and flavonoid production in tartary buckwheat sprout cultures. Cereal Res. Commun. 2015, 43, 661–671. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Dziki, D.; Pietrzak, W.; Nowak, R. Phenolic acids prolife and antioxidant properties of bread enriched with sprouted wheat flour. J. Food Biochem. 2017, 41, e12386. [Google Scholar] [CrossRef]
- Ding, H.; Fu, T.-J.; Smith, M.A. Microbial Contamination in Sprouts: How Effective Is Seed Disinfection Treatment? J. Food Sci. 2013, 78, R495–R501. [Google Scholar] [CrossRef] [PubMed]
- Warriner, K.; Smal, B. Chapter 11–Microbiological Safety of Sprouted Seeds: Interventions and Regulations. In The Produce Contamination Problem, 2nd ed.; Matthews, K.R., Sapers, G.M., Gerba, C.P., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 237–268. ISBN 978-0-12-404611-5. [Google Scholar]
- Riggio, G.M.; Wang, Q.; Kniel, K.E.; Gibson, K.E. Microgreens—A review of food safety considerations along the farm to fork continuum. Int. J. Food Microbiol. 2019, 290, 76–85. [Google Scholar] [CrossRef] [PubMed]
- NSW Food Authority. Products Food Safety Scheme Periodic Review of the Risk; 2019. Available online: https://www.foodauthority.nsw.gov.au/media/4881 (accessed on 27 August 2020).
- Jin, H.; Craig, D.; Blenman, P.; Yates, A.; Crerar, S.; Healy, M.; Jin, H.; Craig, D.; Blenman, P.; Yates, A.; et al. A Through-chain Analysis of Food Safety Hazards and Control Measures Associated with the Production and Supply of Seed Sprouts for Human Consumption. Eur. J. Nutr. Food Saf. 2014, 4, 424–428. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ) Scientific Opinion on the risk posed by Shiga toxin-producing Escherichia coli (STEC) and other pathogenic bacteria in seeds and sprouted seeds. EFSA J. 2011, 9, 2424. [CrossRef]
- Jablasone, J.; Warriner, K.; Griffiths, M. Interactions of Escherichia coli O157:H7, Salmonella typhimurium and Listeria monocytogenes plants cultivated in a gnotobiotic system. Int. J. Food Microbiol. 2005, 99, 7–18. [Google Scholar] [CrossRef]
- Symes, S.; Goldsmith, P.; Haines, H. Microbiological Safety and Food Handling Practices of Seed Sprout Products in the Australian State of Victoria. J. Food Prot. 2015, 78, 1387–1391. [Google Scholar] [CrossRef]
- Jin, H.; Yates, A.; Craig, D.; Blenman, P.; Crerar, S.A. Through-chain Analysis of Microbiological Food Safety Hazards and Control Measures Associated with Production and Supply of Seed Sprouts for Human Consumption. In Current Trends in Food Science Vol.1; Khader, V., Ed.; Book Publisher International: London, UK, 2019; pp. 57–71, Print; ISBN1 978-81-940613-5-9. eBook; ISBN2 978-93-89246-32-2. [Google Scholar]
- Liu, D.; Cui, Y.; Walcott, R.; Díaz-Pérez, J.; Tishchenko, V.; Chen, J. Transmission of human enteric pathogens from artificially-inoculated flowers to vegetable sprouts/seedlings developed via contaminated seeds. Food Control 2019, 99, 21–27. [Google Scholar] [CrossRef]
- Praeger, U.; Herppich, W.B.; Hassenberg, K. Aqueous chlorine dioxide treatment of horticultural produce: Effects on microbial safety and produce quality–A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 318–333. [Google Scholar] [CrossRef]
- Santos, C.S.; Silva, B.; Valente, L.M.P.; Gruber, S.; Vasconcelos, M.W. The Effect of Sprouting in Lentil (Lens culinaris) Nutritional and Microbiological Profile. Foods 2020, 9, 400. [Google Scholar] [CrossRef]
- Wright, K.M.; Holden, N.J. Quantification and colonisation dynamics of Escherichia coli O157:H7 inoculation of microgreens species and plant growth substrates. Int. J. Food Microbiol. 2018, 273, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.A.; Beecher, L.; Northcutt, J.K. Effect of irrigation water source and post-harvest washing treatment on the microflora of alfalfa and mung bean sprouts. Food Control 2019, 100, 151–157. [Google Scholar] [CrossRef]
- Turner, E.R.; Luo, Y.; Buchanan, R.L. Microgreen nutrition, food safety, and shelf life: A review. J. Food Sci. 2020, 85, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Aruscavage, D.; Miller, S.A.; Lewis Ivey, M.L.; Lee, K.E.N.; Lejeune, J.T. Survival and Dissemination of Escherichia coli O157:H7 on Physically and Biologically Damaged Lettuce Plants. J. Food Prot. 2008, 71, 2384–2388. [Google Scholar] [CrossRef] [PubMed]
- Işık, H.; Topalcengiz, Z.; Güner, S.; Aksoy, A. Generic and Shiga toxin-producing Escherichia coli (O157:H7) contamination of lettuce and radish microgreens grown in peat moss and perlite. Food Control 2020, 111, 107079. [Google Scholar] [CrossRef]
- Reed, E.; Ferreira, C.M.; Bell, R.; Brown, E.W.; Zheng, J. Plant-Microbe and Abiotic Factors Influencing Salmonella Survival and Growth on Alfalfa Sprouts and Swiss Chard Microgreens. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, F.; De Bellis, P.; Mininni, C.; Santamaria, P.; Serio, F. Physicochemical, agronomical and microbiological evaluation of alternative growing media for the production of rapini (Brassica rapa L.) microgreens. J. Sci. Food Agric. 2017, 97, 1212–1219. [Google Scholar] [CrossRef]
- Sun, J.; Kou, L.; Geng, P.; Huang, H.; Yang, T.; Luo, Y.; Chen, P. Metabolomic Assessment Reveals an Elevated Level of Glucosinolate Content in CaCl2 Treated Broccoli Microgreens. J. Agric. Food Chem. 2015, 63, 1863–1868. [Google Scholar] [CrossRef]
- Lu, Y.; Dong, W.; Alcazar, J.; Yang, T.; Luo, Y.; Wang, Q.; Chen, P. Effect of preharvest CaCl2 spray and postharvest UV-B radiation on storage quality of broccoli microgreens, a richer source of glucosinolates. J. Food Compos. Anal. 2018, 67, 55–62. [Google Scholar] [CrossRef]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Elia, A. Post-harvest performance of ready-to-eat wild rocket salad as affected by growing period, soilless cultivation system and genotype. Postharvest Biol. Technol. 2019, 156, 110909. [Google Scholar] [CrossRef]
- Sikora, M.; Świeca, M.; Franczyk, M.; Jakubczyk, A.; Bochnak, J.; Złotek, U. Biochemical Properties of Polyphenol Oxidases from Ready-to-Eat Lentil (Lens culinaris Medik.) Sprouts and Factors Affecting Their Activities: A Search for Potent Tools Limiting Enzymatic Browning. Foods 2019, 8, 154. [Google Scholar] [CrossRef] [PubMed]
- Dalal, N.; Siddiqui, S. Evaluation of effects of chemical treatments on sensory attributes of sunflower microgreens with storage. J. Pharmacogn. Phytochem. 2020, 9, 439–443. [Google Scholar]
- Vale, A.P.; Santos, J.; Brito, N.V.; Marinho, C.; Amorim, V.; Rosa, E.; Oliveira, M.B.P.P.P. Effect of refrigerated storage on the bioactive compounds and microbial quality of Brassica oleraceae sprouts. Postharvest Biol. Technol. 2015, 109, 120–129. [Google Scholar] [CrossRef]
- Keshri, J.; Krouptiski, Y.; Abu-Fani, L.; Achmon, Y.; Bauer, T.S.; Zarka, O.; Maler, I.; Pinto, R.; Sela Saldinger, S. Dynamics of bacterial communities in alfalfa and mung bean sprouts during refrigerated conditions. Food Microbiol. 2019, 84, 103261. [Google Scholar] [CrossRef]
- Ishikawa, D.; Shigihara, I.; Nakai, R.; Tamate, H.; Tsukada, Y.; Fujii, T. High Pressure Induced Effects on Free Amino Acid Generation in Mung Bean Sprouts during Preservation after Trigger Pressurization. Food Sci. Technol. Res. 2019, 25, 49–55. [Google Scholar] [CrossRef]
- Westphal, A.; Riedl, K.M.; Cooperstone, J.L.; Kamat, S.; Balasubramaniam, V.M.; Schwartz, S.J.; Böhm, V. High-Pressure Processing of Broccoli Sprouts: Influence on Bioactivation of Glucosinolates to Isothiocyanates. J. Agric. Food Chem. 2017, 65, 8578–8585. [Google Scholar] [CrossRef]
- Xiao, Z.; Luo, Y.; Lester, G.E.; Kou, L.; Yang, T.; Wang, Q. Postharvest quality and shelf life of radish microgreens as impacted by storage temperature, packaging film, and chlorine wash treatment. LWT Food Sci. Technol. 2014, 55, 551–558. [Google Scholar] [CrossRef]
- D’ambrosio, T.; Amodio, M.L.; Pastore, D.; De Santis, G.; Colelli, G. Chemical, physical and sensorial characterization of fresh quinoa sprouts (Chenopodium quinoa Willd.) and effects of modified atmosphere packaging on quality during cold storage. Food Packag. Shelf Life 2017, 14, 52–58. [Google Scholar] [CrossRef]
- Wilson, M.D.; Stanley, R.A.; Eyles, A.; Ross, T. Innovative processes and technologies for modified atmosphere packaging of fresh and fresh-cut fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2019, 59, 411–422. [Google Scholar] [CrossRef]
- Bello, C.; Maldini, M.; Baima, S.; Scaccini, C.; Natella, F. Glucoraphanin and sulforaphane evolution during juice preparation from broccoli sprouts. Food Chem. 2018, 268, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Simsek, S.; El, S.N.; Kancabas Kilinc, A.; Karakaya, S. Vegetable and fermented vegetable juices containing germinated seeds and sprouts of lentil and cowpea. Food Chem. 2014, 156, 289–295. [Google Scholar] [CrossRef]
- Wu, F.; Xu, X. Sprouted Grains-Based Fermented Products; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128115251. [Google Scholar]
- Mridula, D.; Sharma, M. Development of non-dairy probiotic drink utilizing sprouted cereals, legume and soymilk. LWT Food Sci. Technol. 2015, 62, 482–487. [Google Scholar] [CrossRef]
- Xiao-mei, L.I. Preparation of Germinated Soybean Yogurt. Food Sci. 2011, 2. [Google Scholar] [CrossRef]
- Aminah, S.; Meikawati, W.; Rosidi, A. Nutrition retention of product based on soybean sprouts flour and corn sprout flour enriched with duck eggshell. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012009. [Google Scholar] [CrossRef]
- Klopsch, R.; Baldermann, S.; Hanschen, F.S.; Voss, A.; Rohn, S.; Schreiner, M.; Neugart, S. Brassica-enriched wheat bread: Unraveling the impact of ontogeny and breadmaking on bioactive secondary plant metabolites of pak choi and kale. Food Chem. 2019, 295, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, B.; Calzuola, I.; Gigliarelli, L.; Torricelli, R.; Polegri, L.; Vizioli, V.; Benincasa, P.; Marsili, V. Phenolic content and antioxidant activity of wholegrain breads from modern and old wheat (Triticum aestivum L.) cultivars and ancestors enriched with wheat sprout powder. Ital. J. Agron. 2018, 13, 297–302. [Google Scholar] [CrossRef]
- Hernandez-Aguilar, C.; Dominguez-Pacheco, A.; Palma Tenango, M.; Valderrama-Bravo, C.; Soto Hernández, M.; Cruz-Orea, A.; Ordonez-Miranda, J. Lentil sprouts: A nutraceutical alternative for the elaboration of bread. J. Food Sci. Technol. 2020, 57, 1817–1829. [Google Scholar] [CrossRef]
- Hidalgo, A.; Tumbas Šaponjac, V.; Ćetković, G.; Šeregelj, V.; Čanadanović-Brunet, J.; Chiosa, D.; Brandolini, A. Antioxidant properties and heat damage of water biscuits enriched with sprouted wheat and barley. LWT 2019, 114, 114. [Google Scholar] [CrossRef]
- Islam, M.Z.; Yu, D.S.; Lee, Y.T. The Effect of Heat Processing on Chemical Composition and Antioxidative Activity of Tea Made from Barley Sprouts and Wheat Sprouts. J. Food Sci. 2019, 84, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- Šaponjac, V.T.; Ćetković, G.; Čanadanović-Brunet, J.; Mandić, A.; Šeregelj, V.; Vulić, J.; Stajčić, S. Bioactive Characteristics and Storage of Salt Mixtures Seasoned with Powdered Cereal Sprouts. J. Chem. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Gubanenko, G.A.; Naimushina, L.V.; Zhukova, K.O.; Rechkina, A.; Mayurnikova, L.A.; Kiseleva, O.V. Rational use of wheat processing products to create a composite flour mixture. IOP Conf. Ser. Earth Environ. Sci. 2020, 421. [Google Scholar] [CrossRef]
- Dziki, D.; Habza-Kowalska, E.; Gawlik-Dziki, U.; Miś, A.; Rózyło, R.; Krzysiak, Z.; Hassoon, W.H. Drying kinetics, grinding characteristics, and physicochemical properties of broccoli sprouts. Processes 2020, 8, 97. [Google Scholar] [CrossRef]
- Mattioli, S.; Dal Bosco, A.; Castellini, C.; Falcinelli, B.; Sileoni, V.; Marconi, O.; Mancinelli, A.C.; Cotozzolo, E.; Benincasa, P. Effect of heat- and freeze-drying treatments on phytochemical content and fatty acid profile of alfalfa and flax sprouts. J. Sci. Food Agric. 2019, 99, 4029–4035. [Google Scholar] [CrossRef]
- Gan, R.Y.; Lui, W.Y.; Chan, C.L.; Corke, H. Hot Air Drying Induces Browning and Enhances Phenolic Content and Antioxidant Capacity in Mung Bean (Vigna radiata L.) Sprouts. J. Food Process. Preserv. 2017, 41, 2–9. [Google Scholar] [CrossRef]
- Złotek, U.; Gawlik-Dziki, U.; Dziki, D.; Awieca, M.; Nowak, R.; Martinez, E. Influence of Drying Temperature on Phenolic Acids Composition and Antioxidant Activity of Sprouts and Leaves of White and Red Quinoa. J. Chem. 2019, 2019, 1–8. [Google Scholar] [CrossRef]
- Klopsch, R.; Baldermann, S.; Voss, A.; Rohn, S.; Schreiner, M.; Neugart, S. Bread enriched with legume microgreens and leaves-ontogenetic and baking-driven changes in the profile of secondary plant metabolites. Front. Chem. 2018, 6, 1–19. [Google Scholar] [CrossRef]
- Peng, J.; Tang, J.; Barrett, D.M.; Sablani, S.S.; Anderson, N.; Powers, J.R. Thermal pasteurization of ready-to-eat foods and vegetables: Critical factors for process design and effects on quality. Crit. Rev. Food Sci. Nutr. 2017, 57, 2970–2995. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Petruzzi, L.; Perricone, M.; Speranza, B.; Campaniello, D.; Sinigaglia, M.; Corbo, M.R. Nonthermal Technologies for Fruit and Vegetable Juices and Beverages: Overview and Advances. Compr. Rev. Food Sci. Food Saf. 2018, 17, 2–62. [Google Scholar] [CrossRef]
- Roobab, U.; Aadil, R.M.; Madni, G.M.; Bekhit, A.E.-D. The Impact of Nonthermal Technologies on the Microbiological Quality of Juices: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 437–457. [Google Scholar] [CrossRef]
- Ali, N.; Popović, V.; Koutchma, T.; Warriner, K.; Zhu, Y. Effect of thermal, high hydrostatic pressure, and ultraviolet-C processing on the microbial inactivation, vitamins, chlorophyll, antioxidants, enzyme activity, and color of wheatgrass juice. J. Food Process Eng. 2020, 43, 1–11. [Google Scholar] [CrossRef]
- Knödler, M.; Berger, M.; Dobrindt, U. Long-term survival of the Shiga toxin-producing Escherichia coli O104:H4 outbreak strain on fenugreek seeds. Food Microbiol. 2016, 59, 190–195. [Google Scholar] [CrossRef]
- Landry, K.S.; Komaiko, J.; Wong, D.E.; Xu, T.; Mcclements, D.J.; Mclandsborough, L. Inactivation of Salmonella on Sprouting Seeds Using a Spontaneous Carvacrol Nanoemulsion Acidified with Organic Acids. J. Food Prot. 2016, 79, 1115–1126. [Google Scholar] [CrossRef]
- Iacumin, L.; Comi, G. Microbial quality of raw and ready-to-eat mung bean sprouts produced in Italy. Food Microbiol. 2019, 82, 371–377. [Google Scholar] [CrossRef]
- Adhikari, A.; Chhetri, V.S.; Bhattacharya, D.; Cason, C.; Luu, P.; Suazo, A. Effectiveness of daily rinsing of alfalfa sprouts with aqueous chlorine dioxide and ozonated water on the growth of Listeria monocytogenes during sprouting. Lett. Appl. Microbiol. 2019, 69, 252–257. [Google Scholar] [CrossRef]
- Žudytė, B.; Lukšienė, Ž. Toward better microbial safety of wheat sprouts: Chlorophyllin-based photosensitization of seeds. Photochem. Photobiol. Sci. 2019, 18, 2521–2530. [Google Scholar] [CrossRef]
- Hylton, R.K.; Sanchez-Maldonado, A.F.; Peyvandi, P.; Rahmany, F.; Dagher, F.; Leon-velarde, C.G.; Warriner, K.; Hamidi, A.M. Decontamination of Chia and Flax Seed Inoculated with Salmonella and Surrogate, Enterococcus faecium NRRL B-2354, Using a Peracetic Acid Sanitizing Solution: Antimicrobial Efficacy and Impact on Seed Functionality. J. Food Prot. 2019, 82, 486–493. [Google Scholar] [CrossRef]
- Sabillón, L.; Stratton, J.; Rose, D.; Bianchini, A. Reduction in pathogenic load of wheat by tempering with saline organic acid solutions at different seasonal temperatures. Int. J. Food Microbiol. 2020, 313, 108381. [Google Scholar] [CrossRef]
- Zhang, C.; Xia, X.; Li, B.; Hung, Y.-C. Disinfection efficacy of electrolyzed oxidizing water on brown rice soaking and germination. Food Control 2018, 89, 38–45. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Q.; Zhao, D.; Han, X.; Hao, J. Systematic application of slightly acidic electrolyzed water (SAEW) for natural microbial reduction of buckwheat sprouts. LWT 2019, 108, 14–20. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Y.; Zhao, Z.; Liu, W.; Chen, Y.; Yang, G.; Xia, X.; Cao, Y. The application of slightly acidic electrolyzed water in pea sprout production to ensure food safety, biological and nutritional quality of the sprout. Food Control 2019, 104, 83–90. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, Z.; Yang, G.; Shi, Y.; Zhang, Y.; Shi, C.; Xia, X. Effect of slightly acidic electrolyzed water on natural Enterobacteriaceae reduction and seed germination in the production of alfalfa sprouts. Food Microbiol. 2020, 103414. [Google Scholar] [CrossRef]
- Jribi, S.; Molnàr, H.; Antal, O.T.; Adànyi, N.; Kheriji, O.; Naàr, Z.; Debbabi, H. Zinc fortification as a tool for improving sprout hygienic and nutritional quality: A factorial design approach. J. Sci. Food Agric. 2019, 99, 5187–5194. [Google Scholar] [CrossRef]
- Butscher, D.; Van Loon, H.; Waskow, A.; Rudolf von Rohr, P.; Schuppler, M. Plasma inactivation of microorganisms on sprout seeds in a dielectric barrier discharge. Int. J. Food Microbiol. 2016, 238, 222–232. [Google Scholar] [CrossRef]
- Fan, X.; Sokorai, K.; Weidauer, A.; Gotzmann, G.; Rögner, F.-H.; Koch, E. Comparison of gamma and electron beam irradiation in reducing populations of E. coli artificially inoculated on mung bean, clover and fenugreek seeds, and affecting germination and growth of seeds. Radiat. Phys. Chem. 2017, 130, 306–315. [Google Scholar] [CrossRef]
- Choi, S.; Beuchat, L.R.; Kim, H.; Ryu, J.-H. Viability of sprout seeds as affected by treatment with aqueous chlorine dioxide and dry heat, and reduction of Escherichia coli O157:H7 and Salmonella enterica on pak choi seeds by sequential treatment with chlorine dioxide, drying, and dry heat. Food Microbiol. 2016, 54, 127–132. [Google Scholar] [CrossRef]
- Waskow, A.; Betschart, J.; Butscher, D.; Oberbossel, G.; Klöti, D.; Büttner-Mainik, A.; Adamcik, J.; von Rohr, P.R.; Schuppler, M. Characterization of Efficiency and Mechanisms of Cold Atmospheric Pressure Plasma Decontamination of Seeds for Sprout Production. Front. Microbiol. 2018, 9, 3164. [Google Scholar] [CrossRef]
- Reyes-Jurado, F.; Navarro-Cruz, A.R.; Méndez-Aguilar, J.; Ochoa-Velasco, C.E.; Mani-López, E.; Jiménez-Munguía, M.T.; Palou, E.; López-Malo, A.; Ávila-Sosa, R. High-Intensity Light Pulses To Inactivate Salmonella Typhimurium on Mexican Chia (Salvia hispanica L.) Seeds. J. Food Prot. 2019, 82, 1272–1277. [Google Scholar] [CrossRef]
- Kim, S.-M.; Hwang, H.-J.; Cheigh, C.-I.; Chung, M.-S. Bactericidal effect of intense pulsed light on seeds without loss of viability. Food Sci. Biotechnol. 2019, 28, 281–287. [Google Scholar] [CrossRef]
- Puligundla, P.; Kim, J.-W.; Mok, C. Effects of Nonthermal Plasma Treatment on Decontamination and Sprouting of Radish (Raphanus sativus L.) Seeds. Food Bioprocess Technol. 2017, 10, 1093–1102. [Google Scholar] [CrossRef]
- Puligundla, P.; Kim, J.-W.; Mok, C. Effect of corona discharge plasma jet treatment on decontamination and sprouting of rapeseed (Brassica napus L.) seeds. Food Control 2017, 71, 376–382. [Google Scholar] [CrossRef]
- Kim, J.-W.; Puligundla, P.; Mok, C. Effect of corona discharge plasma jet on surface-borne microorganisms and sprouting of broccoli seeds. J. Sci. Food Agric. 2017, 97, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Puligundla, P.; Kim, J.-W.; Mok, C. Effect of atmospheric pressure plasma treatment on seed decontamination and sprouting of pak choi (Brassica rapa L. subsp. chinensis (L.) Hanelt). Chiang Mai J. Sci. 2018, 45, 2679–2690. [Google Scholar]
- Zhou, R.; Li, J.; Zhou, R.; Zhang, X.; Yang, S. Atmospheric-pressure plasma treated water for seed germination and seedling growth of mung bean and its sterilization effect on mung bean sprouts. Innov. Food Sci. Emerg. Technol. 2019, 53, 36–44. [Google Scholar] [CrossRef]
- Sarooei, S.J.; Abbasi, A.; Shaghaghian, S.; Berizi, E. Effect of Ozone as a Disinfectant on Microbial Load and Chemical Quality of Raw Wheat Germ. Ozone Sci. Eng. 2019, 41, 562–570. [Google Scholar] [CrossRef]
- Mohammad, Z.; Kalbasi-Ashtari, A.; Riskowski, G.; Castillo, A. Reduction of Salmonella and Shiga toxin-producing Escherichia coli on alfalfa seeds and sprouts using an ozone generating system. Int. J. Food Microbiol. 2019, 289, 57–63. [Google Scholar] [CrossRef]
- Kang, J.-W.; Lee, J.-I.; Jeong, S.-Y.; Kim, Y.-M.; Kang, D.-H. Effect of 222-nm krypton-chloride excilamp treatment on inactivation of Escherichia coli O157:H7 and Salmonella Typhimurium on alfalfa seeds and seed germination. Food Microbiol. 2019, 82, 171–176. [Google Scholar] [CrossRef]
- Hong, E.-J.; Kang, D.-H. Effect of sequential dry heat and hydrogen peroxide treatment on inactivation of Salmonella Typhimurium on alfalfa seeds and seeds germination. Food Microbiol. 2016, 53, 9–14. [Google Scholar] [CrossRef]
- Trząskowska, M.; Dai, Y.; Delaquis, P.; Wang, S. Pathogen reduction on mung bean reduction of Escherichia coli O157:H7, Salmonella enterica and Listeria monocytogenes on mung bean using combined thermal and chemical treatments with acetic acid and hydrogen peroxide. Food Microbiol. 2018, 76, 62–68. [Google Scholar] [CrossRef]
- Wang, S.; Chen, J.; Wakeling, C.; Bach, S.; Orban, S.; Delaquis, P. Disinfection of Alfalfa and Radish Sprouting Seed Using Oxidizing Agents and Treatments Compliant with Organic Food Production Principles. J. Food Prot. 2019, 83, 779–787. [Google Scholar] [CrossRef]
- Goñi, M.G.; Tomadoni, B.M.; Audisio, M.C.; Ibarguren, C.; Roura, S.I.; del Moreira, M.R.; Ponce, A.G. Application of bacteriocins from Enterococcus hirae on butterhead lettuce seeds inoculated with Escherichia coli O157: H7. Int. Food Res. J. 2016, 23, 2653–2660. [Google Scholar]
- Fong, K.; LaBossiere, B.; Switt, A.I.M.; Delaquis, P.; Goodridge, L.; Levesque, R.C.; Danyluk, M.D.; Wang, S. Characterization of Four Novel Bacteriophages Isolated from British Columbia for Control of Non-typhoidal Salmonella in Vitro and on Sprouting Alfalfa Seeds. Front. Microbiol. 2017, 8, 2193. [Google Scholar] [CrossRef]
- Lone, A.; Anany, H.; Hakeem, M.; Aguis, L.; Avdjian, A.-C.; Bouget, M.; Atashi, A.; Brovko, L.; Rochefort, D.; Griffiths, M.W. Development of prototypes of bioactive packaging materials based on immobilized bacteriophages for control of growth of bacterial pathogens in foods. Int. J. Food Microbiol. 2016, 217, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-I.; Choi, S.Y.; Han, I.; Cho, S.K.; Lee, Y.; Kim, S.; Kang, B.; Choi, O.; Kim, J. Inhibition of Salmonella enterica growth by competitive exclusion during early alfalfa sprout development using a seed-dwelling Erwinia persicina strain EUS78. Int. J. Food Microbiol. 2020, 312, 108374. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Lathrop, A. Effects of Lactobacillus plantarum, Pediococcus acidilactici, and Pediococcus pentosaceus on the Growth of Listeria monocytogenes and Salmonella on Alfalfa Sprouts. J. Food Prot. 2019, 82, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.-B.; Boomer, A.; Chen, C.-H.; Patel, J. Efficacy of benzyl isothiocyanate for controlling Salmonella on alfalfa seeds and sprouts. Int. J. Food Sci. Technol. 2020, 55, 2669–2676. [Google Scholar] [CrossRef]
- Choi, J.-S. Seed-surface disinfection and germination effects of grapefruit seed extract (GSE) on Lactuca sativa seeds. Toxicol. Environ. Health Sci. 2017, 9, 169–175. [Google Scholar] [CrossRef]
- Lorenzo-Leal, A.C.; Palou, E.; López-Malo, A. Evaluation of the efficiency of allspice, thyme and rosemary essential oils on two foodborne pathogens in in-vitro and on alfalfa seeds, and their effect on sensory characteristics of the sprouts. Int. J. Food Microbiol. 2019, 295, 19–24. [Google Scholar] [CrossRef]
- Sahan, N.; Tornuk, F. Application of plant hydrosols for decontamination of wheat, lentil and mung bean seeds prior to sprouting. Qual. Assur. Saf. Crop. Foods 2016, 8, 575–582. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Lui, W.-Y.; Wu, K.; Chan, C.-L.; Dai, S.-H.; Sui, Z.-Q.; Corke, H. Bioactive compounds and bioactivities of germinated edible seeds and sprouts: An updated review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Williamson, E.M.; Liu, X.; Izzo, A.A. Trends in use, pharmacology, and clinical applications of emerging herbal nutraceuticals. Br. J. Pharmacol. 2020, 177, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Brower, V. Nutraceuticals: Poised for a healthy slice of the healthcare market? Nat. Biotechnol. 1998, 16, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Dong, Y.; Ren, H.; Li, L.; He, C. A review of phytochemistry, metabolite changes, and medicinal uses of the common food mung bean and its sprouts (Vigna radiata). Chem. Cent. J. 2014, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yu, X.; Ma, F.; Li, J. RNA-Seq Analysis of Transcriptome and Glucosinolate Metabolism in Seeds and Sprouts of Broccoli (Brassica oleracea var. italic). PLoS ONE 2014, 9, e88804. [Google Scholar] [CrossRef] [PubMed]
- Shlisky, J.; Bloom, D.E.; Beaudreault, A.R.; Tucker, K.L.; Keller, H.H.; Freund-Levi, Y.; Fielding, R.A.; Cheng, F.W.; Jensen, G.L.; Wu, D.; et al. Nutritional Considerations for Healthy Aging and Reduction in Age-Related Chronic Disease. Adv. Nutr. 2017, 8, 17–26. [Google Scholar] [CrossRef]
- Schäfer, F.; Jeanne, J.-F. Evaluating the effects of food on health in a world of evolving operational challenges. Contemp. Clin. Trials Commun. 2018, 12, 51–54. [Google Scholar] [CrossRef]
- Kroeger, C.M.; Garza, C.; Lynch, C.J.; Myers, E.; Rowe, S.; Schneeman, B.O.; Sharma, A.M.; Allison, D.B. Scientific rigor and credibility in the nutrition research landscape. Am. J. Clin. Nutr. 2018, 107, 484–494. [Google Scholar] [CrossRef]
- Ferruzza, S.; Natella, F.; Ranaldi, G.; Murgia, C.; Rossi, C.; Trošt, K.; Mattivi, F.; Nardini, M.; Maldini, M.; Giusti, A.M.; et al. Nutraceutical Improvement Increases the Protective Activity of Broccoli Sprout Juice in a Human Intestinal Cell Model of Gut Inflammation. Pharmaceuticals 2016, 9, 48. [Google Scholar] [CrossRef]
- Risbridger, G.P. Human Cell Lines as Tools of Our Trade: “Laying It on the (Cell) Line”. Mol. Endocrinol. 2015, 29, 1–2. [Google Scholar] [CrossRef]
- Liu, Y.; Mi, Y.; Mueller, T.; Kreibich, S.; Williams, E.G.; Van Drogen, A.; Borel, C.; Frank, M.; Germain, P.-L.; Bludau, I.; et al. Multi-omic measurements of heterogeneity in HeLa cells across laboratories. Nat. Biotechnol. 2019, 37, 314–322. [Google Scholar] [CrossRef]
- Min, M.; Spencer, S.L. Spontaneously slow-cycling subpopulations of human cells originate from activation of stress-response pathways. PLoS Biol. 2019, 17, 1–25. [Google Scholar] [CrossRef]
- Lee, J.K.; Bloom, J.; Zubeldia-Plazaola, A.; Garbe, J.C.; Stampfer, M.R.; LaBarge, M.A. Different culture media modulate growth, heterogeneity, and senescence in human mammary epithelial cell cultures. PLoS ONE 2018, 13, e0204645. [Google Scholar] [CrossRef]
- Bedard, P.L.; Hansen, A.R.; Ratain, M.J.; Siu, L.L. Tumour heterogeneity in the clinic. Nature 2013, 501, 355–364. [Google Scholar] [CrossRef]
- Cencic, A.; Chingwaru, W. The Role of Functional Foods, Nutraceuticals, and Food Supplements in Intestinal Health. Nutrients 2010, 2, 611–625. [Google Scholar] [CrossRef]
- Narmada, B.C.; Goh, Y.T.; Li, H.; Sinha, S.; Yu, H.; Cheung, C. Human Stem Cell-Derived Endothelial-Hepatic Platform for Efficacy Testing of Vascular-Protective Metabolites from Nutraceuticals. Stem Cells Transl. Med. 2017, 6, 851–863. [Google Scholar] [CrossRef][Green Version]
- Booij, T.H.; Price, L.S.; Danen, E.H.J. 3D Cell-Based Assays for Drug Screens: Challenges in Imaging, Image Analysis, and High-Content Analysis. SLAS Discov. Adv. Sci. Drug Discov. 2019, 24, 615–627. [Google Scholar] [CrossRef]
- Phan, N.; Hong, J.J.; Tofig, B.; Mapua, M.; Elashoff, D.; Moatamed, N.A.; Huang, J.; Memarzadeh, S.; Damoiseaux, R.; Soragni, A. A simple high-throughput approach identifies actionable drug sensitivities in patient-derived tumor organoids. Commun. Biol. 2019, 2, 78. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. A critical review on phytochemical profile and health promoting effects of mung bean (Vigna radiata). Food Sci. Hum. Wellness 2018, 7, 11–33. [Google Scholar] [CrossRef]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.-A.; den Broeck, H.C.; Brouns, F.J.P.H.; De Brier, N.; et al. Impact of Cereal Seed Sprouting on Its Nutritional and Technological Properties: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 305–328. [Google Scholar] [CrossRef]
- Santini, A.; Cammarata, S.M.; Capone, G.; Ianaro, A.; Tenore, G.C.; Pani, L.; Novellino, E. Nutraceuticals: Opening the debate for a regulatory framework. Br. J. Clin. Pharmacol. 2018, 84, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Hasler, C.M. Functional Foods: Benefits, Concerns and Challenges—A Position Paper from the American Council on Science and Health. J. Nutr. 2002, 132, 3772–3781. [Google Scholar] [CrossRef] [PubMed]
- Yagishita, Y.; Fahey, J.W.; Dinkova-Kostova, A.T.; Kensler, T.W. Broccoli or Sulforaphane: Is It the Source or Dose That Matters? Molecules 2019, 24, 3593. [Google Scholar] [CrossRef] [PubMed]
- Quirante-Moya, S.; García-Ibañez, P.; Quirante-Moya, F.; Villaño, D.; Moreno, D.A. The Role of Brassica Bioactives on Human Health: Are We Studying It the Right Way? Molecules 2020, 25, 1591. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Sudini, K.; Diette, G.B.; Breysse, P.N.; McCormack, M.C.; Bull, D.; Biswal, S.; Zhai, S.; Brereton, N.; Peng, R.D.; Matsui, E.C. A Randomized Controlled Trial of the Effect of Broccoli Sprouts on Antioxidant Gene Expression and Airway Inflammation in Asthmatics. J. Allergy Clin. Immunol. Pract. 2016, 4, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Duran, C.G.; Burbank, A.J.; Mills, K.H.; Duckworth, H.R.; Aleman, M.M.; Kesic, M.J.; Peden, D.B.; Pan, Y.; Zhou, H.; Hernandez, M.L. A proof-of-concept clinical study examining the NRF2 activator sulforaphane against neutrophilic airway inflammation. Respir. Res. 2016, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Reynolds, C.; Brooker, A.; Talalay, P.; Fahey, J.W. Sulforaphane improves the bronchoprotective response in asthmatics through Nrf2-mediated gene pathways. Respir. Res. 2015, 16, 106. [Google Scholar] [CrossRef]
- Morand, C.; De Roos, B.; Garcia-Conesa, M.T.; Gibney, E.R.; Landberg, R.; Manach, C.; Milenkovic, D.; Rodriguez-Mateos, A.; Van de Wiele, T.; Tomas-Barberan, F. Why interindividual variation in response to consumption of plant food bioactives matters for future personalised nutrition. Proc. Nutr. Soc. 2020, 79, 225–235. [Google Scholar] [CrossRef]
- Son, T.G.; Camandola, S.; Mattson, M.P. Hormetic Dietary Phytochemicals. NeuroMolecular Med. 2008, 10, 236. [Google Scholar] [CrossRef]
- Landberg, R.; Manach, C.; Kerckhof, F.-M.; Minihane, A.-M.; Saleh, R.N.M.; De Roos, B.; Tomas-Barberan, F.; Morand, C.; Van de Wiele, T. Future prospects for dissectinginter-individual variability in the absorption, distribution and elimination of plantbioactives of relevance for cardiometabolic endpoints. Eur. J. Nutr. 2019, 58, 21–36. [Google Scholar] [CrossRef]
- Shahidi, F. Beneficial Health Effects and Drawbacks of Antinutrients and Phytochemicals in Foods. In Antinutrients and Phytochemicals in Food; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1997; Volume 662, p. 1. ISBN 9780841234987. [Google Scholar]
- Peluso, I. Dietary Antioxidants: Micronutrients and Antinutrients in Physiology and Pathology. Antioxidants 2019, 8, 642. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Jain, S. Tannins: An antinutrient with positive effect to manage diabetes. Res. J. Recent Sci. ISSN 2012, 2277, 2502. [Google Scholar]
- Nissar, J.; Ahad, T.; Naik, H.R.; Hussain, S.Z. A review phytic acid: As antinutrient or nutraceutical. J. Pharmacogn. Phytochem. 2017, 6, 1554–1560. [Google Scholar]
- Calder, P.C. Comment on Christiansen et al.: When food met pharma. Br. J. Nutr. 2015, 114, 1109–1110. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akande, K.E.; Doma, U.D.; Agu, H.O.; Adamu, H.M. Major antinutrients found in plant protein sources: Their effect on nutrition. Pakistan J. Nutr. 2010, 9, 827–832. [Google Scholar] [CrossRef]
- Ghavidel, R.A.; Prakash, J. The impact of germination and dehulling on nutrients, antinutrients, in vitro iron and calcium bioavailability and in vitro starch and protein digestibility of some legume seeds. LWT Food Sci. Technol. 2007, 40, 1292–1299. [Google Scholar] [CrossRef]
- Preet, K.; Punia, D. Antinutrients and Digestibility (in vitro) of Soaked, Dehulled and Germinated Cowpeas. Nutr. Health 2000, 14, 109–117. [Google Scholar] [CrossRef]
- Mishra, M.; Jain, S. Effect of Soaking and Germination on Nutritional profile and Antinutrients of Buckwheat Whole (Fagopyrum esculentum). Int. J. Curr. Microbiol. App. Sci. 2019, 8, 3384–3393. [Google Scholar] [CrossRef]
- Padmashree, N.N.; Handu, S.; Khan, M.A.; Semwal, A.D.; Sharma, G.K. Effect of Germination on Nutritional, Antinutritional and Rheological Characteristics of Chenopodium quinoa. Energy 2019, 375, 353.02b. [Google Scholar] [CrossRef]
- Mbithi-Mwikya, S.; Van Camp, J.; Yiru, Y.; Huyghebaert, A. Nutrient and Antinutrient Changes in Finger Millet (Eleusine coracan) During Sprouting. LWT Food Sci. Technol. 2000, 33, 9–14. [Google Scholar] [CrossRef]
- Budhwar, S.; Sethi, K.; Chakraborty, M. Efficacy of germination and probiotic fermentation on underutilized cereal and millet grains. Food Prod. Process. Nutr. 2020, 2, 12. [Google Scholar] [CrossRef]
- Bouajila, A.; Ammar, H.; Chahine, M.; Khouja, M.; Hamdi, Z.; Khechini, J.; Salem, A.-F.Z.M.; Ghorbel, A.; López, S. Changes in phytase activity, phosphorus and phytate contents during grain germination of barley (Hordeum vulgare L.) cultivars. Agrofor. Syst. 2019, 94, 1151–1159. [Google Scholar] [CrossRef]
- Kajla, P.; Sharma, A.; Sood, D.R. Effect of germination on proximate principles, minerals and anti nutrients of flaxseeds. Asian J. Dairy Food Res. 2017, 36, 52–57. [Google Scholar] [CrossRef]
- Pal, R.S.; Bhartiya, A.; Yadav, P.; Kant, L.; Mishra, K.K.; Aditya, J.P.; Pattanayak, A. Effect of dehulling, germination and cooking on nutrients, anti-nutrients, fatty acid composition and antioxidant properties in lentil (Lens culinaris). J. Food Sci. Technol. 2017, 54, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Cardador-Martínez, A.; Martínez-Tequitlalpan, Y.; Gallardo-Velazquez, T.; Sánchez-Chino, X.M.; Martínez-Herrera, J.; Corzo-Ríos, L.J.; Jiménez-Martínez, C. Effect of Instant Controlled Pressure-Drop on the Non-Nutritional Compounds of Seeds and Sprouts of Common Black Bean (Phaseolus vulgaris L.). Molecules 2020, 25, 1464. [Google Scholar] [CrossRef] [PubMed]
- Khattak, A.B.; Zeb, A.; Bibi, N.; Khalil, S.A.; Khattak, M.S. Influence of germination techniques on phytic acid and polyphenols content of chickpea (Cicer arietinum L.) sprouts. Food Chem. 2007, 104, 1074–1079. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Ma, Z.; Cui, Y.; Du, C.; Xia, X.; Qian, H. Beneficial Influence of Short-Term Germination on Decreasing Allergenicity of Peanut Proteins. J. Food Sci. 2016, 81, T255–T261. [Google Scholar] [CrossRef]
- Rao, H.; Chen, C.; Tian, Y.; Li, Y.; Gao, Y.; Tao, S.; Xue, W. Germination results in reduced allergenicity of peanut by degradation of allergens and resveratrol enrichment. Innov. Food Sci. Emerg. Technol. 2018, 50, 188–195. [Google Scholar] [CrossRef]
- Wolejszo, A.; Szymkiewicz, A.; Troszynska, A. Immunoreactive properties and sensory quality of lentil [Lens culinaris] and mung bean [Vigma radiata L.] sprouts. Polish J. Food Nutr. Sci. 2007, 57, 415–420. [Google Scholar]
- Peñas, E.; Gomez, R.; Frias, J.; Baeza, M.L.; Vidal-Valverde, C. High hydrostatic pressure effects on immunoreactivity and nutritional quality of soybean products. Food Chem. 2011, 125, 423–429. [Google Scholar] [CrossRef]
- Guhsl, E.E.; Hofstetter, G.; Hemmer, W.; Ebner, C.; Vieths, S.; Vogel, L.; Breiteneder, H.; Radauer, C. Vig r 6, the cytokinin-specific binding protein from mung bean (Vigna radiata) sprouts, cross-reacts with Bet v 1-related allergens and binds IgE from birch pollen allergic patients’ sera. Mol. Nutr. Food Res. 2014, 58, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Pedersen, M.H.; Skov, P.S.; Poulsen, L.K.; Bindslev-Jensen, C.; Andersen, S.B.; Torp, A.M. Peanut cross-reacting allergens in seeds and sprouts of a range of legumes. Clin. Exp. Allergy 2008, 38, 1969–1977. [Google Scholar] [CrossRef] [PubMed]
- FDA—Food and Drug Administration Compliance with and Implementation of the Standards for the Growing, Harvesting, Packing, and Holding of Produce for Human Consumption for Sprouts Operations: Guidance for Industry; 2017. Available online: https://www.fda.gov/media/102430/download (accessed on 27 August 2020).
- Cragg, G.M.; Katz, F.; Newman, D.J.; Rosenthal, J. The impact of the United Nations Convention on Biological Diversity on natural products research. Nat. Prod. Rep. 2012, 29, 1407–1423. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.-H.; Vederas, J.C. Drug Discovery and Natural Products: End of an Era or an Endless Frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Neugart, S.; Baldermann, S.; Hanschen, F.S.; Klopsch, R.; Wiesner-Reinhold, M.; Schreiner, M. The intrinsic quality of brassicaceous vegetables: How secondary plant metabolites are affected by genetic, environmental, and agronomic factors. Sci. Hortic. 2018, 233, 460–478. [Google Scholar] [CrossRef]
- Guo, R.; Huang, Z.; Deng, Y.; Chen, X.; XuHan, X.; Lai, Z. Comparative Transcriptome Analyses Reveal a Special Glucosinolate Metabolism Mechanism in Brassica alboglabra Sprouts. Front. Plant Sci. 2016, 7, 1497. [Google Scholar] [CrossRef]
- Wang, S.; Alseekh, S.; Fernie, A.R.; Luo, J. The Structure and Function of Major Plant Metabolite Modifications. Mol. Plant 2019, 12, 899–919. [Google Scholar] [CrossRef]
- Koch, M.A.; Schuffenhauer, A.; Scheck, M.; Wetzel, S.; Casaulta, M.; Odermatt, A.; Ertl, P.; Waldmann, H. Charting biologically relevant chemical space: A structural classification of natural products (SCONP). Proc. Natl. Acad. Sci. USA 2005, 102, 17272–17277. [Google Scholar] [CrossRef]
- Barker, A.; Kettle, J.G.; Nowak, T.; Pease, J.E. Expanding medicinal chemistry space. Drug Discov. Today 2013, 18, 298–304. [Google Scholar] [CrossRef]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.I.; et al. A comprehensive map of molecular drug targets. Nat. Rev. Drug Discov. 2017, 16, 19–34. [Google Scholar] [CrossRef]
- Chen, Y.; Garcia de Lomana, M.; Friedrich, N.-O.; Kirchmair, J. Characterization of the Chemical Space of Known and Readily Obtainable Natural Products. J. Chem. Inf. Model. 2018, 58, 1518–1532. [Google Scholar] [CrossRef] [PubMed]
- Karageorgis, G.; Foley, D.J.; Laraia, L.; Waldmann, H. Principle and design of pseudo-natural products. Nat. Chem. 2020, 12, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef]
- Nützmann, H.-W.; Huang, A.; Osbourn, A. Plant metabolic clusters–from genetics to genomics. New Phytol. 2016, 211, 771–789. [Google Scholar] [CrossRef]
- Kautsar, S.A.; Suarez Duran, H.G.; Blin, K.; Osbourn, A.; Medema, M.H. plantiSMASH: Automated identification, annotation and expression analysis of plant biosynthetic gene clusters. Nucleic Acids Res. 2017, 45, W55–W63. [Google Scholar] [CrossRef]
- Töpfer, N.; Fuchs, L.-M.; Aharoni, A. The PhytoClust tool for metabolic gene clusters discovery in plant genomes. Nucleic Acids Res. 2017, 45, 7049–7063. [Google Scholar] [CrossRef]
- Wisecaver, J.H.; Borowsky, A.T.; Tzin, V.; Jander, G.; Kliebenstein, D.J.; Rokas, A. A Global Coexpression Network Approach for Connecting Genes to Specialized Metabolic Pathways in Plants. Plant Cell 2017, 29, 944–959. [Google Scholar] [CrossRef]
- Schläpfer, P.; Zhang, P.; Wang, C.; Kim, T.; Banf, M.; Chae, L.; Dreher, K.; Chavali, A.K.; Nilo-Poyanco, R.; Bernard, T.; et al. Genome-Wide Prediction of Metabolic Enzymes, Pathways, and Gene Clusters in Plants. Plant Physiol. 2017, 173, 2041–2059. [Google Scholar] [CrossRef]
- Moore, B.M.; Wang, P.; Fan, P.; Leong, B.; Schenck, C.A.; Lloyd, J.P.; Lehti-Shiu, M.D.; Last, R.L.; Pichersky, E.; Shiu, S.-H. Robust predictions of specialized metabolism genes through machine learning. Proc. Natl. Acad. Sci. USA 2019, 116, 2344–2353. [Google Scholar] [CrossRef]
- Liu, Z.; Suarez Duran, H.G.; Harnvanichvech, Y.; Stephenson, M.J.; Schranz, M.E.; Nelson, D.; Medema, M.H.; Osbourn, A. Drivers of metabolic diversification: How dynamic genomic neighbourhoods generate new biosynthetic pathways in the Brassicaceae. New Phytol. 2020, 227, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.A.P.; Thornburg, C.C.; Henrich, C.J.; Grkovic, T.; O’Keefe, B.R. Creating and screening natural product libraries. Nat. Prod. Rep. 2020, 37, 893–918. [Google Scholar] [CrossRef] [PubMed]
- Appleton, D.R.; Buss, A.D.; Butler, M.S. A simple method for high-throughput extract prefractionation for biological screening. Chim. Int. J. Chem. 2007, 61, 327–331. [Google Scholar] [CrossRef]
- Tu, Y.; Jeffries, C.; Ruan, H.; Nelson, C.; Smithson, D.; Shelat, A.A.; Brown, K.M.; Li, X.-C.; Hester, J.P.; Smillie, T.; et al. Automated High-Throughput System to Fractionate Plant Natural Products for Drug Discovery. J. Nat. Prod. 2010, 73, 751–754. [Google Scholar] [CrossRef]
- Camp, D.; Davis, R.A.; Campitelli, M.; Ebdon, J.; Quinn, R.J. Drug-like Properties: Guiding Principles for the Design of Natural Product Libraries. J. Nat. Prod. 2012, 75, 72–81. [Google Scholar] [CrossRef]
- Nagana Gowda, G.A.; Raftery, D. Recent Advances in NMR-Based Metabolomics. Anal. Chem. 2017, 89, 490–510. [Google Scholar] [CrossRef]
- Wolfender, J.-L.; Nuzillard, J.-M.; van der Hooft, J.J.J.; Renault, J.-H.; Bertrand, S. Accelerating Metabolite Identification in Natural Product Research: Toward an Ideal Combination of Liquid Chromatography–High-Resolution Tandem Mass Spectrometry and NMR Profiling, in Silico Databases, and Chemometrics. Anal. Chem. 2019, 91, 704–742. [Google Scholar] [CrossRef]
- Grienke, U.; Foster, P.A.; Zwirchmayr, J.; Tahir, A.; Rollinger, J.M.; Mikros, E. 1H NMR-MS-based heterocovariance as a drug discovery tool for fishing bioactive compounds out of a complex mixture of structural analogues. Sci. Rep. 2019, 9, 11113. [Google Scholar] [CrossRef]
- Macarron, R.; Banks, M.N.; Bojanic, D.; Burns, D.J.; Cirovic, D.A.; Garyantes, T.; Green, D.V.S.; Hertzberg, R.P.; Janzen, W.P.; Paslay, J.W.; et al. Impact of high-throughput screening in biomedical research. Nat. Rev. Drug Discov. 2011, 10, 188–195. [Google Scholar] [CrossRef]
- Foito, A.; Stewart, D. Metabolomics: A high-throughput screen for biochemical and bioactivity diversity in plants and crops. Curr. Pharm. Des. 2018, 24, 2043–2054. [Google Scholar] [CrossRef]
- Paananen, J.; Fortino, V. An omics perspective on drug target discovery platforms. Brief. Bioinform. 2019. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, H.; Zheng, S.; Zhang, B.; Deng, Z. Implication of the Significance of Dietary Compatibility: Based on the Antioxidant and Anti-Inflammatory Interactions with Different Ratios of Hydrophilic and Lipophilic Antioxidants among Four Daily Agricultural Crops. J. Agric. Food Chem. 2018, 66, 7461–7474. [Google Scholar] [CrossRef] [PubMed]
- Olszowy, M.; Dawidowicz, A.L.; Jóźwik-Dolęba, M. Are mutual interactions between antioxidants the only factors responsible for antagonistic antioxidant effect of their mixtures? Additive and antagonistic antioxidant effects in mixtures of gallic, ferulic and caffeic acids. Eur. Food Res. Technol. 2019, 245, 1473–1485. [Google Scholar] [CrossRef]
- Bisol, Â.; de Campos, P.S.; Lamers, M.L. Flavonoids as anticancer therapies: A systematic review of clinical trials. Phyther. Res. 2020, 34, 568–582. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic Compounds in Brassica Vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Farnham, M.W.; Wilson, P.E.; Stephenson, K.K.; Fahey, J.W. Genetic and environmental effects on glucosinolate content and chemoprotective potency of broccoli. Plant Breed. 2004, 123, 60–65. [Google Scholar] [CrossRef]
- Li, B.; Xiong, M.; Zhang, H.-Y. Elucidating Polypharmacological Mechanisms of Polyphenols by Gene Module Profile Analysis. Int. J. Mol. Sci. 2014, 15, 11245–11254. [Google Scholar] [CrossRef]
- Grynkiewicz, G. Flavonoids as a library of privileged structures-what are the gains? J. Biochem. Res. 2019, 2, 8–16. [Google Scholar]
- Brown, K.K.; Hampton, M.B. Biological targets of isothiocyanates. Biochim. Biophys. Acta Gen. Subj. 2011, 1810, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; MacMillan, J.B. Targeting and extending the eukaryotic druggable genome with natural products. Nat. Prod. Rep. 2020, 37, 744–746. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Zhang, Y.; Talalay, P. Broccoli sprouts: An exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc. Natl. Acad. Sci. USA 1997, 94, 10367–10372. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A. Sulforaphane: Its “Coming of Age” as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease. Oxid. Med. Cell. Longev. 2019, 2019, 2716870. [Google Scholar] [CrossRef]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective Glucosinolates and Isothiocyanates of Broccoli Sprouts. Cancer Epidemiol. Prev. Biomarkers 2001, 10, 501–508. [Google Scholar]
- Palliyaguru, D.L.; Yuan, J.-M.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the Power of Plants to People. Mol. Nutr. Food Res. 2018, 62, 1700965. [Google Scholar] [CrossRef]
- Clulow, J.A.; Storck, E.M.; Lanyon-Hogg, T.; Kalesh, K.A.; Jones, L.H.; Tate, E.W. Competition-based{,} quantitative chemical proteomics in breast cancer cells identifies new target profiles for sulforaphane. Chem. Commun. 2017, 53, 5182–5185. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, W.; Zhou, Z.; Sun, C. Benefits and Risks of the Hormetic Effects of Dietary Isothiocyanates on Cancer Prevention. PLoS ONE 2014, 9, e114764. [Google Scholar] [CrossRef]
- Anighoro, A.; Bajorath, J.; Rastelli, G. Polypharmacology: Challenges and Opportunities in Drug Discovery. J. Med. Chem. 2014, 57, 7874–7887. [Google Scholar] [CrossRef]
- Proschak, E.; Stark, H.; Merk, D. Polypharmacology by Design: A Medicinal Chemist’s Perspective on Multitargeting Compounds. J. Med. Chem. 2019, 62, 420–444. [Google Scholar] [CrossRef]
- Farghaly, M.M.; Abdullah, M.A.M.; Youssef, I.M.I.; Abdel-Rahim, I.R.; Abouelezz, K. Effect of feeding hydroponic barley sprouts to sheep on feed intake, nutrient digestibility, nitrogen retention, rumen fermentation and ruminal enzymes activity. Livest. Sci. 2019, 228, 31–37. [Google Scholar] [CrossRef]
- Chantiratikul, A.; Borisuth, L.; Chinrasri, O.; Saenthaweesuk, N.; Chookhampaeng, S.; Thosaikham, W.; Sriart, N.; Chantiratikul, P. Evaluation of the toxicity of selenium from hydroponically produced selenium-enriched kale sprout in laying hens. J. Trace Elem. Med. Biol. 2016, 35, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Abouelezz, K.F.M.; Sayed, M.A.M.; Abdelnabi, M.A. Evaluation of hydroponic barley sprouts as a feed supplement for laying Japanese quail: Effects on egg production, egg quality, fertility, blood constituents, and internal organs. Anim. Feed Sci. Technol. 2019, 252, 126–135. [Google Scholar] [CrossRef]
- Chantiratikul, A.; Chinrasri, O.; Pakmaruek, P.; Chantiratikul, P.; Thosaikham, W.; Aengwanich, W. Responses of Growing Japanese Quails that Received Selenium from Selenium Enriched Kale Sprout (Brassica oleracea var. alboglabra L.). Biol. Trace Elem. Res. 2011, 144, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Chinrasri, O.; Chantiratikul, P.; Maneetong, S.; Chookhampaeng, S.; Chantiratikul, A. Productivity and Selenium Concentrations in Egg and Tissue of Laying Quails Fed Selenium from Hydroponically Produced Selenium-Enriched Kale Sprout (Brassica oleracea var. alboglabra L.). Biol. Trace Elem. Res. 2013, 155, 381–386. [Google Scholar] [CrossRef]
- Mattioli, S.; Dal Bosco, A.; Martino, M.; Ruggeri, S.; Marconi, O.; Sileoni, V.; Falcinelli, B.; Castellini, C.; Benincasa, P. Alfalfa and flax sprouts supplementation enriches the content of bioactive compounds and lowers the cholesterol in hen egg. J. Funct. Foods 2016, 22, 454–462. [Google Scholar] [CrossRef]
- Chantiratikul, A.; Chinrasri, O.; Chantiratikul, P. Effect of Selenium from Selenium-Enriched Kale Sprout Versus Other Selenium Sources on Productivity and Selenium Concentrations in Egg and Tissue of Laying Hens. Biol. Trace Elem. Res. 2018, 182, 105–110. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Castellini, C.; Martino, M.; Mattioli, S.; Marconi, O.; Sileoni, V.; Ruggeri, S.; Tei, F.; Benincasa, P. The effect of dietary alfalfa and flax sprouts on rabbit meat antioxidant content, lipid oxidation and fatty acid composition. Meat Sci. 2015, 106, 31–37. [Google Scholar] [CrossRef]
- Mohsen, M.K.; Abdel-Raouf, E.M.; Gaafar, H.M.A.; Yousif, A.M. Nutritional evaluation of sprouted barley grains on agricultural by-products on performance of growing New Zealand white rabbits. Nat. Sci. 2015, 13, 35–45. [Google Scholar]
- Al-Saadi, M.J.; Al-Zubiadi, I.A.H. Effects of substitution barley by 10%, 30% of sprouted barley on rumen characters, digestibility and feed efficiency in diet of awassi male lambs. Int. J. Sci. Res. 2016, 5, 2228–2233. [Google Scholar]
- Helal, H.G. Productive and reproductive performance of Barki ewes fed on sprouted barley grains on desert by-products during lactating period. Res. J. Anim. Vet. Sci. 2018, 10, 37–48. [Google Scholar]
- North, M.K.; Dalle Zotte, A.; Hoffman, L.C. The use of dietary flavonoids in meat production: A review. Anim. Feed Sci. Technol. 2019, 257, 114291. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Morales-Medina, J.C.; Palmieri, B.; Pezzuto, F.; Cocco, R.; Flores, G.; Iannitti, T. Functional foods in pet nutrition: Focus on dogs and cats. Res. Vet. Sci. 2017, 112, 161–166. [Google Scholar] [CrossRef]
- Bol, S.; Caspers, J.; Buckingham, L.; Anderson-Shelton, G.D.; Ridgway, C.; Buffington, C.A.T.; Schulz, S.; Bunnik, E.M. Responsiveness of cats (Felidae) to silver vine (Actinidia polygama), Tatarian honeysuckle (Lonicera tatarica), valerian (Valeriana officinalis) and catnip (Nepeta cataria). BMC Vet. Res. 2017, 13, 70. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A. Prospects of insects as food and feed. Org. Agric. 2020, 1–8. [Google Scholar] [CrossRef]

| Class | Function in Plants | References | |
|---|---|---|---|
| Polyphenols | Flavonoids | Pigments (i.e., anthocyanins, flavanols, and flavones); UV protection (i.e., flavones and flavonols); scavenger of H2O2, superoxide, hydroxyl radical, singlet oxygen, peroxyl radical; feeding and wounding deterrents (i.e., hydrolyzable tannins); insecticidal and antimicrobial action (i.e., isoflavones). | [13,14,15] |
| Phenolic acids | Plant-microbe symbiosis; allelopathic effect (caffeic and ferulic acid); feeding deterrents as astringent compounds (i.e., condensed tannins); resistance to pathogen attack; components of cell-wall; attract pollinators and seed dispersers; signaling molecules (i.e., salicylic acid). | [14,15] | |
| Stilbenes | Defensive response to pathogens or stress conditions; protective agents from viral and microbial attack, injuries, diseases, UV exposure, aluminum, etc. | [15,16] | |
| Lignans | Building blocks for the fiber lignin; plant defense diseases and pests. | [15,17] | |
| Terpenoids | Overall | Plant growth regulators; defense against herbivores and pathogens; attracting pollinators influence the growth and the development of neighbor plant. | [13,18] |
| Carotenoids | Pigments; protection against photooxidation; light-harvesting pigments. | [13,14] | |
| Tocotrienols and tocopherols | Singlet oxygen scavengers in photosystem ii; signaling molecules; maintain the integrity of long-chain polyunsaturated fatty acids in the cell membranes. | [19] | |
| N-containing metabolites | Glucosinolates | Herbivore toxins and feeding repellents; insects deterrents and pathogens growth inhibitors (isothiocyanates). | [12,13,14,20] |
| Chemical Class | Subclass | Health Benefits | References |
|---|---|---|---|
| Polyphenols | Flavonoids | Antioxidants: long-lived radical anions scavengers; anti-lipoperoxidant, -tumoral, -platelet, ischemic, -allergic, -inflammatory activities. | [21] |
| Phenolic acids | Radical scavengers; singlet oxygen quenching; block biosynthesis of leukotrienes involved in immunoregulation disease, asthma and allergic reaction (caffeic acid). | [21] | |
| Stilbenes | Anti-oxidative, -carcinogenic, -tumor properties; retard cardiovascular disease and cancer; prevent lipid oxidation and the formation of toxic oxidation products. | [21] | |
| Phytoestrogens (i.e., isoflavones, lignans, coumestans) | Prevent menopausal symptoms (hot-flashes, vasomotor symptoms, vaginal atrophy); anti-aging effect on the skin; enhance bone formation and increase bone mineral density; beneficial effects on endothelial cells, vascular smooth muscle; reduce levels of low-density lipoprotein (LDL) cholesterol; anti-mutagenic, -proliferative, -angiogenic, pro-apoptotic and general anti-cancer effects. | [22] | |
| Terpenoids | Carotenoids | Chemical quenchers of 1O2; reduce the risk of prostate cancer and age-related macular degeneration; stimulate the immune system in inflammatory diseases or human immunodeficiency disease. | [23] |
| Tocotrienols and tocopherols | Retard cancer lesions and tumors; tackle free radical reactions; scavengers of lipid peroxyl radicals; singlet oxygen quenchers. | [23] | |
| Phytosterols | Reduce cholesterol levels; attenuate inflammatory processes; induce apoptosis when added to cultured human prostate, breast, and colon cancer cells. | [24] | |
| N-containing metabolites | Glucosinolates | Protect against Helicobacter pylori infections, light-induced damage of retina brain ischemia/reperfusion, traumatic brain injury, intracerebral hemorrhage, and contusion spinal cord injury (sulforaphane); decrease infarct size, brain (or spinal cord) edema, and cortical apoptosis (sulforaphane); restoration of skin integrity; inhibit growth of human tumor cells. | [25] |
| Family | Species | Days after Sowing (DAS) | Secondary Metabolites | References |
|---|---|---|---|---|
| Amaranthaceae | Amaranthus caudatus–amaranth | 10 | PAs, total PC, FC | [43] |
| Amaranthus cruentus–amaranth | 4, 6, 7 | Total PC, AA | [44] | |
| Amaranthus hypochondriacus–amaranth | 2 | AA | [45] | |
| N.S. | Carotenoids | [46] | ||
| Chenopodium album–pigweed | 48 h | AA, total PC | [47] | |
| 48 h | Total PC | [48] | ||
| 48 h | Tocopherols | [49] | ||
| Chenopodium quinoa–quinoa | 4, 6, 7 | Total PC, AA | [44] | |
| 82 h | Total PC, AA, single phenolics | [50] | ||
| 4 | Total PC, AA | [51] | ||
| 5 | Total PC | [52] | ||
| Chenopodium berlandieri–huauzontle | 12, 24, 36, 48, 72 h | N.R. | [53] | |
| Apiaceae | Anethum graveolens–dill | 8–12 | Total PC, FC, AA | [54] |
| Coriandrum sativum L.–coriander | N.S. | Carotenoids, total PC, single phenolics | [55] | |
| 20 | AA, carotenoids, total PC, single phenolics | [56] | ||
| Asteraceae | Artemisia dracunculus–tarragon | 1, 3, 5, 7 | N/A | [57] |
| Taraxacum officinale–common dandelion | 16 | Anthocyanins and carotenoids | [58] | |
| Boraginaceae | Phacelia tanacetifolia–phacelia | 7 | Total PC, AA, free and bound PAs and flavonoids | [59] |
| Brassicaceae | Cichorium intybus–chicory | 12 | Total PC, tocopherols, anthocyanins, carotenoids | [60] |
| Diplotaxis tenuifolia–wild rocket | 4, 7 | Total and single GLS | [61] | |
| 7, 21 | Anthocyanins, phenolics, AA, resveratrol | [62] | ||
| Convolvulaceae | Ipomea aquatica–water convolvulus | 8–12 | Total PC, FC, AA | [54] |
| Malvaceae | Corchorus olitorius L.–jute | 22 | AA, carotenoids, total PC, single phenolics | [56] |
| Poaceae | Coix lacryma-jobi–adlay seed | 12, 24, 36, 48, 60 h | Free, bound total PC, flavonoid, PAs and AA | [63] |
| Phalaris canariensis–canary seed | 24, 48, 72, 96, 120 h | Free, bound total PC, PAs and AA | [64] | |
| Triticum aestivum spp. spelta–spelt | 5 and 12 | Total PC, free and bound PAs, AA | [42] | |
| Triticum monococcum ssp. monococcum–einkorn | 5 and 12 | Total PC, free and bound PAs, AA | [42] | |
| 5 and 13 | Total PC, FC, AA | [65] | ||
| 5 and 12 | Total PC, free and bound PAs, AA | [66] | ||
| 5 and 9 | Total PC, free and bound PAs, AA | [67] | ||
| Triticum turgidum spp. dicoccum–emmer | 5 and 12 | Total PC, free and bound PAs, AA | [42] | |
| 5 and 13 | Total PC, FC, AA | [45] | ||
| 5 and 12 | Total PC, free and bound PAs, AA | [65] | ||
| Zizania latifolia–wild rice | Free, bound and total phenolics | [68] | ||
| Lamiaceae | Salvia hispanica–chia | 7 | Total PC, AA, free and bound PAs and FC | [59] |
| Leguminosae | Vigna umbellata–rice bean | 6, 12, 18, 24 h | Total PC, single PAs, single flavonoids and AA | [69] |
| Onagraceae | Oenothera biennis–evening primrose | 7 | Total PC, AA, free and bound PAs and flavonoids | [59] |
| Portulacaceae | Portulaca oleracea–common purslane | 1–6 | N/A | [70] |
| 3 | Free, bound soluble conjugated total PC, PAs and flavonoids | [71] | ||
| Portulaca grandiflora–moss-rose purslane | 1–6 | N/A | [70] | |
| Livestock | Sprout Species | Sprout Family | Days after Sowing (DAS) | Effects on Livestock Health and Products | Phytochemicals Transferred to Livestock Products | References |
|---|---|---|---|---|---|---|
| Poultry | ||||||
| Betagro laying hens | Brassica oleracea var. Alboglabra–Chinese kale | Brassicaceae | 15 | ↑Se content in eggs and tissues | N/A | [323] |
| Japanese quails | Hordeum vulgare–barley | Poaceae | 7 | ↑egg laying rate; ↑relative weights of gizzard and testis; ↑ fertility; ↑ number of hatched chicks/female; no effect on egg quality indices | N/A | [324] |
| Brassica oleracea var. alboglabra–Chinese kale | Brassicaceae | 7 | No effects on performances and carcass characteristics; ↑Se content in tissues | N/A | [325] | |
| 15 | Se from sprout does not affect performances and egg quality; ↑Se content in eggs | N/A | [326] | |||
| Leghorn hens | Medicago sativa–alfalfa | Leguminosae | 3 | ↓cholesterol content in eggs | ↑ isoflavones, antioxidants and vitamin content in eggs | [327] |
| Linum usitatissimum–flax | Linaceae | 3 | ↓cholesterol content in eggs | ↑ isoflavones, antioxidants and vitamin content in eggs | [327] | |
| Lohmann Brown hens | Brassica oleracea var. alboglabra–Chinese kale | Brassicaceae | 7 | ↑ Se bioavailability and Se content in eggs, tissues and plasma | N/A | [328] |
| Rabbits | ||||||
| New Zealand white rabbits | Medicago sativa–alfalfa | Leguminosae | 3 | ↑ n-3 fatty acid contents of meat | ↑phytoestrogens in muscles | [329] |
| Linum usitatissimum–flax | Linaceae | 3 | ↑ n-3 fatty acid contents of meat | ↑phytoestrogens in muscles | [329] | |
| Hordeum vulgare–barley | Poaceae | 12 | No adverse effects on performances and meat characteristics | N/A | [330] | |
| Ruminants | ||||||
| Awassi male lambs | Hordeum vulgare – barley | Poaceae | 7 | ↑some rumen characters; ↑most nutrient digestibility; ↑ feeding efficiency | N/A | [331] |
| Barky ewes | Hordeum vulgare–barley | Poaceae | 15 | ↑NDF and ADF digestibility; ↑fat, total solid, ash and energy of milk | N/A | [332] |
| Rams | Hordeum vulgare–barley | Poaceae | 8 | ↓DM intake; ↑enzyme activity and digestive function | N/A | [332] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galieni, A.; Falcinelli, B.; Stagnari, F.; Datti, A.; Benincasa, P. Sprouts and Microgreens: Trends, Opportunities, and Horizons for Novel Research. Agronomy 2020, 10, 1424. https://doi.org/10.3390/agronomy10091424
Galieni A, Falcinelli B, Stagnari F, Datti A, Benincasa P. Sprouts and Microgreens: Trends, Opportunities, and Horizons for Novel Research. Agronomy. 2020; 10(9):1424. https://doi.org/10.3390/agronomy10091424
Chicago/Turabian StyleGalieni, Angelica, Beatrice Falcinelli, Fabio Stagnari, Alessandro Datti, and Paolo Benincasa. 2020. "Sprouts and Microgreens: Trends, Opportunities, and Horizons for Novel Research" Agronomy 10, no. 9: 1424. https://doi.org/10.3390/agronomy10091424
APA StyleGalieni, A., Falcinelli, B., Stagnari, F., Datti, A., & Benincasa, P. (2020). Sprouts and Microgreens: Trends, Opportunities, and Horizons for Novel Research. Agronomy, 10(9), 1424. https://doi.org/10.3390/agronomy10091424