Seed Lipid Composition of New Hybrids of Styrian Oil Pumpkin Grown in Poland

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Lipid Extraction From Seeds
2.3. Fatty Acid Composition of Pumpkin Seed Lipids
2.4. Quality and Composition of Bioactive Compounds in Pumpkin Seed Lipids
2.5. Oxidative Stability of Pumpkin Seed Lipids
2.6. Statistical Analysis
3. Results and Discussion
3.1. Seed Lipid Content and Fatty Acid Composition
3.2. Tocopherol Content and Composition
3.3. Sterols Content and Composition
3.4. Squalene Content
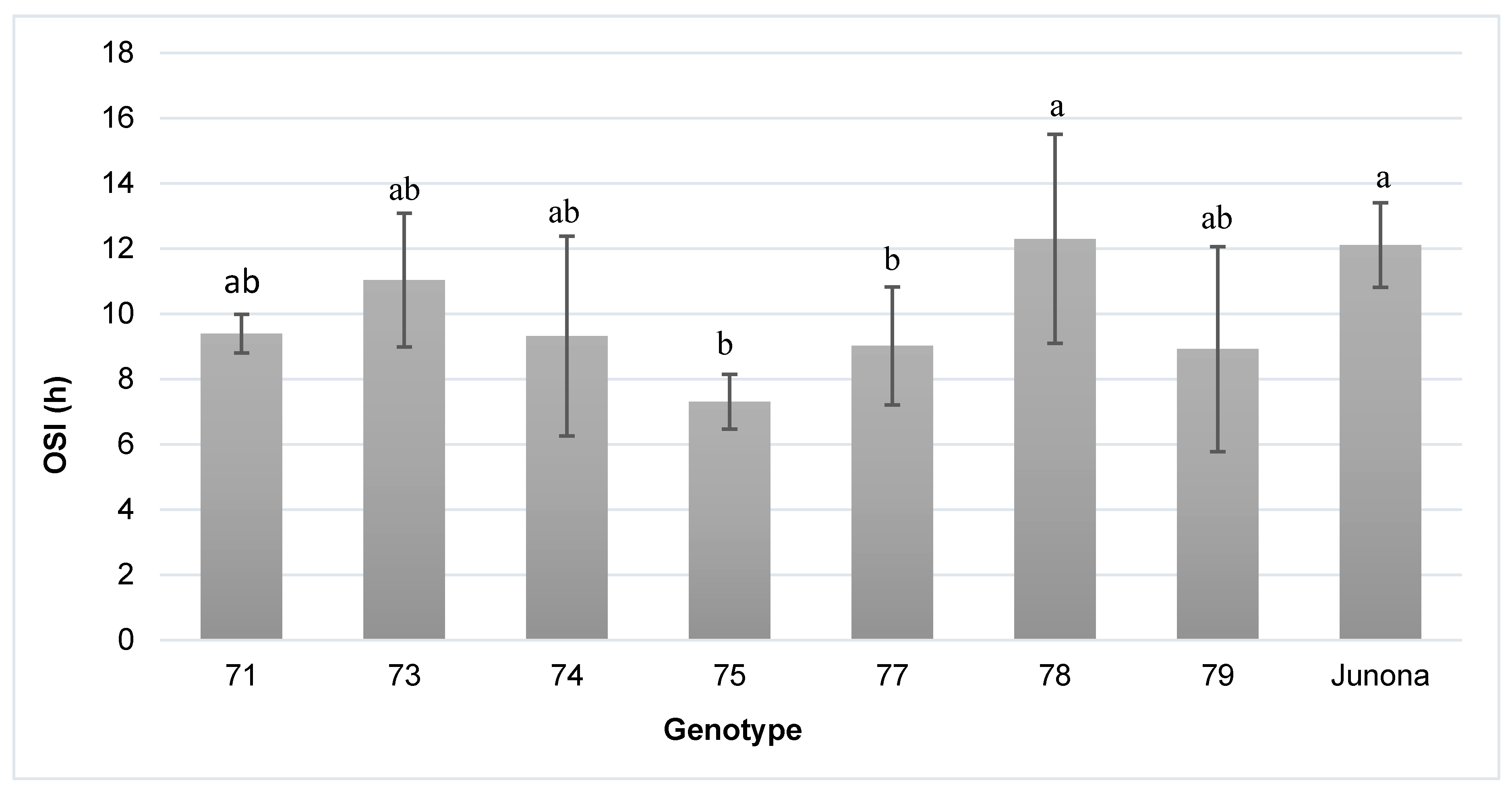
3.5. Oxidative Stability Index (OSI)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Caili, F.U.; Huan, S.; Quanhong, L.I. A review on pharmacological activities and utilization technologies of pumpkin. Plant Foods Hum. Nutr. 2006, 61, 70–77. [Google Scholar] [CrossRef]
- Yadav, M.; Jain, S.; Tomar, R.; Prasad, G.B.K.S.; Yadav, H. Medicinal and biological potential of pumpkin: An updated review. Nutr. Res. Rev. 2010, 23, 184–190. [Google Scholar] [CrossRef]
- Kaur, S.; Panghal, A.; Garg, M.; Mann, S.; Khatkar, S.; Sharma, P.; Chhikara, N. Functional and nutraceutical properties of pumpkin—A review. Nutr. Food Sci. 2019, 50. [Google Scholar] [CrossRef]
- Sharma, R.; Kumar, S.; Kumar, V.; Thakur, A. Comprehensive review on nutraceutical significance of phytochemicals as functional food ingredients for human health management. J. Pharmacogn. Phytochem. 2019, 8, 385–395. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, G.; Kehinde, B.A.; Chhikara, N.; Panghal, A.; Kaur, H. Pharmacological and biomedical uses of extracts of pumpkin and its relatives and applications in the food industry: A review. Int. J. Veg. Sci. 2020, 26, 79–95. [Google Scholar] [CrossRef]
- Alfawaz, M.A. Chemical composition and oil characteristics of pumpkin (Cucurbita maxima) seed kernels. Food Sci. Agric. 2004, 2, 5–18. [Google Scholar]
- Fruhwirth, G.O.; Hermetter, A. Production technology and characteristics of Styrian pumpkin seed oil. Eur. J. Lipid Sci. Technol. 2008, 110, 637–644. [Google Scholar] [CrossRef]
- Gohari, A.A.; Farhoosh, R.; Haddad, K.M. Chemical composition and physicochemical properties of pumpkin seeds (Cucurbita pepo subsp. pepo var. Styriaka) grown in Iran. J. Agric. Sci. Technol. 2011, 13, 1053–1063. [Google Scholar]
- Nawirska-Olszańska, A.; Kita, A.; Biesiada, A.; Sokół-Łętowska, A.; Kucharska, A.Z. . Characteristics of antioxidant activity and composition of pumpkin seed oils in 12 cultivars. Food Chem. 2013, 139, 155–161. [Google Scholar] [CrossRef]
- Procida, G.; Stancher, B.; Cateni, F.; Zacchigna, M. Chemical composition and functional characterization of commercial pumpkin seed oil. In Proceedings of the ChimAlSi 2012, IX Italian Congress of Food,” Food, Functional Foods and Nutraceuticals”, Faculty of Food and Agriculture, UAE University, Ischia, Italy, 03–07 June 2012; p. 63. [Google Scholar]
- Neđeral, S.; Petrović, M.; Vincek, D.; Pukec, D.; Škevin, D.; Kraljić, K.; Obranović, M. Variance of quality parameters and fatty acid composition in pumpkin seed oil during three crop seasons. Ind. Crops Prod. 2014, 60, 15–21. [Google Scholar] [CrossRef]
- Potočnik, T.; Ogrinc, N.; Potočnik, D.; Košir, I.J. Fatty acid composition and δ13C isotopic ratio characterisation of pumpkin seed oil. J. Food Compos. Anal. 2016, 53, 85–90. [Google Scholar] [CrossRef]
- Kachel, M.; Matwijczuk, A.; Przywara, A.; Kraszkiewicz, A.; Koszel, M. Profile of fatty acids and spectroscopic characteristics of selected vegetable oils extracted by cold maceration. Agric. Eng. 2018, 22, 61–71. [Google Scholar] [CrossRef]
- Lelley, T.; Loy, B.; Murkovic, M. Hull-less oil seed pumpkin. In Oil Crops; Springer: New York, NY, USA, 2009; pp. 469–492. [Google Scholar]
- Paris, H.S. Overview of the origins and history of the five major cucurbit crops: Issues for ancient DNA analysis of archaeological specimens. Veg. Hist. Archaeobotany 2016, 25, 405–414. [Google Scholar] [CrossRef]
- Chomicki, G.; Schäfer, H.; Renner, S.S. Origin and domestication of Cucurbitaceae crops: Insights from phylogenies, genomics and archaeology. New Phytol. 2020, 226, 1240–1255. [Google Scholar] [CrossRef] [PubMed]
- Ferriol, M.; Picó, B. Pumpkin and Winter Squash; Springer Science and Business Media LLC: Berlin, Germany, 2008; Volume 1, pp. 317–349. [Google Scholar]
- Korzeniewska, A.; Witek, M.; Gałecka, T.; Niemirowicz-Szczytt, K. Evaluation of selected traits in hull-less seed squash (Cucurbita pepo subsp. pepo var. styriaca Greb.). Pol. J. Agron. 2013, 12, 32–37. (In Polish) [Google Scholar]
- Winkler, J. The origin and breeding of the hull-less seeded Styrian oil-pumpkin varieties in Austria. Rep. Cucurbit Genet. Coop. 2000, 23, 101–104. [Google Scholar]
- Cui, H.; Loy, J.B. Heterosis for seed yield exhibited in hull-less seeded pumpkin. In Cucurbitaceae; ASHS Press: Alexandria, VA, USA, 2002; pp. 323–329. [Google Scholar]
- Darrudi, R. , Nazeri, V.; Soltani, F.; Shokrpour, M.; Ercolano, M.R. Evaluation of combining ability in Cucurbita pepo L. and Cucurbita moschata Duchesne accessions for fruit and seed quantitative traits. J. Appl. Res. Med. Aromat. Plants 2018, 9, 70–77. [Google Scholar]
- Council Regulation (EEC). No. 2081/92. Application for Registration: Art. 17, PGI (National application No: 1215-GR/95). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A31992R2081 (accessed on 7 June 2020).
- Adam, E.; Bernhart, M.; Müller, H.; Winkler, J.; Berg, G. The Cucurbita pepo seed microbiome: Genotype-specific composition and implications for breeding. Plant Soil 2018, 422, 35–49. [Google Scholar] [CrossRef]
- Baxter, G.G.; Murphy, A.; Paech, A. The potential to produce pumpkin seed for processing in North East Victoria. Rural Ind. Dev. Corp. 2012, 11/145, 5–36. [Google Scholar]
- Polish National List of Vegetable Plant Varieties. Available online: http://www.coboru.pl/English/index_eng.aspx (accessed on 10 May 2020).
- Korzeniewska, A.; Niemirowicz-Szczytt, K. Possibilities of using presently underrated oil yielding plants exemplified by oil seed pumpkin. In Biological Diversity of Agricultural Ecosystems and Possibilities of Its Protection in Ecological Farms; Tyburski, J., Kostrzewska, M.K., Eds.; Uniwersytet Warmińsko-Mazurski, Olsztyn: Olsztyn, Poland, 2013; pp. 43–51. (In Polish) [Google Scholar]
- Polish Standard PN-73/R-66164. Determination of Fat Content in Seeds, Oil Fruits and Post-Extraction Meal. Polish Committee for Standardization. (In Polish). Available online: https://www.pkn.pl/en/standardization (accessed on 7 June 2020).
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Zadernowski, R.; Sosulski, F. Composition of total lipids in rapeseed. J. Am. Oil Chem. Soc. 1978, 55, 870–872. [Google Scholar] [CrossRef]
- Czaplicki, S.; Ogrodowska, D.; Derewiaka, D.; Tańska, M.; Zadernowski, R. Bioactive compounds in unsaponifiable fraction of oils from unconventional sources. Eur. J. Lipid Sci. Technol. 2011, 113, 1456–1464. [Google Scholar] [CrossRef]
- Roszkowska, B.; Tańska, M.; Czaplicki, S.; Konopka, I. Variation in the composition and oxidative stability of commercial rapeseed oils during their shelf life. Eur. J. Lipid Sci. Technol. 2015, 117, 673–683. [Google Scholar] [CrossRef]
- Petkova, Z.Y.; Antova, G.A. Changes in the composition of pumpkin seeds (Cucurbita moschata) during development and maturation. Grasas Aceites 2015, 66, 058. [Google Scholar] [CrossRef]
- van der Schoot, C.; Paul, L.K.; Paul, S.B.; Rinne, P.L. Plant lipid bodies and cell-cell signaling: A new role for an old organelle? Plant Signal. Behav. 2011, 6, 1732–1738. [Google Scholar] [CrossRef] [PubMed]
- Nikiforidis, C.V. Structure and functions of oleosomes (oil bodies). Adv. Colloid Interface Sci. 2019, 274, 102039. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yan, T.; Chen, X.; Li, Z.; Wu, D.; Hua, S.; Jiang, L. Effect of high night temperature on storage lipids and transcriptome changes in developing seeds of oilseed rape. J. Exp. Bot. 2018, 69, 1721–1733. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, F.D.; Yarizade, K. Bioinformatics study of delta-12 fatty acid desaturase 2 (FAD2) gene in oilseeds. Mol. Biol. Rep. 2014, 41, 5077–5087. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Semchuk, N.M. Tocopherol biosynthesis: Chemistry, regulation and effects on environmental factors. Acta Physiol. Plant. 2012, 34, 1607–1628. [Google Scholar] [CrossRef]
- Gorjanović, S.Ž.; Rabrenović, B.B.; Novaković, M.M.; Dimić, E.B.; Basić, Z.N.; Sužnjević, D.Ž. Cold-pressed pumpkin seed oil antioxidant activity as determined by a DC polarographic assay based on hydrogen peroxide scavenge. J. Am. Oil Chem. Soc. 2011, 88, 1875–1882. [Google Scholar] [CrossRef]
- Vujasinovic, V.; Djilas, S.; Dimic, E.; Basic, Z.; Radocaj, O. The effect of roasting on the chemical composition and oxidative stability of pumpkin oil. Eur. J. Lipid Sci. Technol. 2012, 114, 568–574. [Google Scholar] [CrossRef]
- Butinar, B.; Bučar-Miklavčič, M.; Mariani, C.; Raspor, P. New vitamin E isomers (gamma-tocomonoenol and alpha-tocomonoenol) in seeds, roasted seeds and roasted seed oil from the Slovenian pumpkin variety ‘Slovenska golica’. Food Chem. 2011, 128, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Naziri, E.; Mitić, M.N.; Tsimidou, M.Z. Contribution of tocopherols and squalene to the oxidative stability of cold-pressed pumpkin seed oil (Cucurbita pepo L.). Eur. J. Lipid Sci. Technol. 2016, 118, 898–905. [Google Scholar] [CrossRef]
- Azzi, A. Tocopherols, tocotrienols and tocomonoenols: Many similar molecules but only one vitamin E. Redox Biol. 2019, 101259. [Google Scholar] [CrossRef]
- Foods High in Alpha-Tocopherol. Available online: https://healthyeating.sfgate.com/foods-high-alphatocopherol-3483.html (accessed on 7 June 2020).
- Ali, M.A.; Nargis, A.; Othman, N.H.; Noor, A.F.; Sadik, G.; Hossen, J. Oxidation stability and compositional characteristics of oils from microwave roasted pumpkin seeds during thermal oxidation. Int. J. Food Prop. 2017, 20, 2569–2580. [Google Scholar] [CrossRef]
- Jiang, Q.; Christen, S.; Shigenaga, M.K.; Ames, B.N. γ-Tocopherol, the major form of vitamin E in the US diet, deserves more attention. Am. J. Clin. Nutr. 2001, 74, 714–722. [Google Scholar] [CrossRef]
- Burbank, A.J.; Duran, C.G.; Pan, Y.; Burns, P.; Jones, S.; Jiang, Q.; Yang, C.; Jenkins, S.; Wells, H.; Alexis, N.; et al. Gamma tocopherol-enriched supplement reduces sputum eosinophilia and endotoxin-induced sputum neutrophilia in volunteers with asthma. J. Allergy Clin. Immunol. 2018, 141, 1231–1238. [Google Scholar] [CrossRef]
- Dulf, F.V.; Bele, C.; Unguresan, M.; Parlog, R.; Socaciu, C. Phytosterols as markers in identification of the adulterated pumpkin seed oil with sunflower oil. Bull. Uasvm Agric. 2009, 66, 301–307. [Google Scholar]
- Ogrodowska, D.; Laaksonen, O.; Tańska, M.; Konopka, I.; Linderborg, K.M. Pumpkin oil addition and encapsulation process as methods to improve oxidative stability of fish oil. Lwt—Food Sci. Technol. 2020, 124, 109142. [Google Scholar] [CrossRef]
- Konopka, I.; Roszkowska, B.; Czaplicki, S.; Tańska, M. Optimization of pumpkin oil recovery by using aqueous enzymatic extraction and comparison of the quality of the obtained oil with the quality of cold-pressed oil. Food Technol. Biotechnol. 2016, 54, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Hrabovski, N.; Sinadinović-Fišer, S.; Nikolovski, B.; Sovilj, M.; Borota, O. Phytosterols in pumpkin seed oil extracted by organic solvents and supercritical CO2. Eur. J. Lipid Sci. Technol. 2012, 114, 1204–1211. [Google Scholar] [CrossRef]
- Yang, R.; Xue, L.; Zhang, L.; Wang, X.; Qi, X.; Jiang, J.; Yu, L.; Wang, X.; Zhang, W.; Zhang, Q.; et al. Phytosterol contents of edible oils and their contributions to estimated phytosterol intake in the Chinese diet. Foods 2019, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, O.N.; Przybylski, R.; Rudzińska, M. Lipid components of flax, perilla, and chia seeds. Eur. J. Lipid Sci. Technol. 2012, 114, 794–800. [Google Scholar] [CrossRef]
- Mandl, A.; Reich, G.; Lindner, W. Detection of adulteration of pumpkin seed oil by analysis of content and composition of specific Δ7-phytosterols. Eur. Food Res. Technol. 1999, 209, 400–406. [Google Scholar] [CrossRef]
- Ramak, P.; Mahboubi, M. The beneficial effects of pumpkin (Cucurbita pepo L.) seed oil for health condition of men. Food Rev. Int. 2019; 35, 166–176. [Google Scholar]
- Tsai, Y.S.; Tong, Y.C.; Cheng, J.T.; Lee, C.H.; Yang, F.S.; Lee, H.Y. Pumpkin seed oil and phytosterol-F can block testosterone/prazosin-induced prostate growth in rats. Urol. Int. 2006, 77, 269–274. [Google Scholar] [CrossRef]
- Heim, S.; Seibt, S.; Stier, H.; Moré, M.I. Uromedic® pumpkin seed derived Δ7-sterols, extract and oil inhibit 5α-reductases and bind to androgen receptor in vitro. Pharm. Pharm. 2018, 9, 193. [Google Scholar]
- Nyam, K.L.; Tan, C.P.; Lai, O.M.; Long, K.; Man, Y.C. Physicochemical properties and bioactive compounds of selected seed oils. Lwt-Food Sci. Technol. 2009, 42, 1396–1403. [Google Scholar] [CrossRef]
- Busquets, A.; Keim, V.; Closa, M.; Del Arco, A.; Boronat, A.; Arró, M.; Ferrer, A. Arabidopsis thaliana contains a single gene encoding squalene synthase. Plant Mol. Biol. 2008, 67, 25–36. [Google Scholar] [CrossRef]
- Ibrahim, N.I.; Fairus, S.; Zulfarina, M.S.; Naina Mohamed, I. The efficacy of squalene in cardiovascular disease risk—A systematic review. Nutrients 2020, 12, 414. [Google Scholar] [CrossRef]
- Lozano-Grande, M.A.; Gorinstein, S.; Espitia-Rangel, E.; Dávila-Ortiz, G.; Martínez-Ayala, A.L. Plant sources, extraction methods, and uses of squalene. Int. J. Agron. 2018. [Google Scholar] [CrossRef]
- Jafari, M.; Goli, S.A.H.; Rahimmalek, M. The chemical composition of the seeds of Iranian pumpkin cultivars and physicochemical characteristics of the oil extract. Eur. J. Lipid Sci. Technol. 2012, 114, 161–167. [Google Scholar] [CrossRef]
- Bardaa, S.; Halima, N.B.; Aloui, F.; Mansour, R.B.; Jabeur, H.; Bouaziz, M.; Sahnoun, Z. Oil from pumpkin (Cucurbita pepo L.) seeds: Evaluation of its functional properties on wound healing in rats. Lipids Health Dis. 2016, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.N.; Breyen, M.D. (Eds.) Joining and Assembly of Medical Materials and Devices; Woodhead Publishing: Sawston, Cambridge, UK, 2013; pp. 386–388. [Google Scholar]
- van Hoed, V.; Sampaio, K.A.; Felkner, B.; Bavec, F.; Scippo, M.L.; Brose, F.; Bavec, M.; Verhé, R. Tocopherols and polyphenols in pumpkin seed oil are moderately affected by industrially relevant roasting conditions. Eur. J. Lipid Sci. Technol. 2017, 119, 1700110. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Hybrid/Variety | Lipids (% of Seed Dry Matter) | Fatty Acids (%) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Free | Bound | Palmitic C16:0 | Stearic C18:0 | Oleic C18:1 | Linoleic C18:2 (n-6) | α-Linolenic C18:3 (n-3) | |||||||||||
| S.D. | S.D. | % in Total | S.D. | % in Total | S.D. | S.D. | S.D. | S.D. | S.D. | |||||||||
| 71 | 49.0 a | 0.7 | 47.5 a | 0.81 | 96.9 | 1.5 bc | 0.4 | 3.1 | 10.5 b | 0.1 | 5.1 b | 0.1 | 29.0 b | 0.3 | 54.6 d | 0.4 | 0.8 a | 0.1 |
| 73 | 47. 7 ab | 1.7 | 45.9 a | 2.17 | 96.0 | 1.8 c | 0.6 | 4.0 | 10.7 bc | 0.1 | 5.4 bcd | 0.1 | 29.8 b | 0.1 | 53.2 c | 0.2 | 0.8 a | 0.2 |
| 74 | 47.7 ab | 0.8 | 47.4 a | 0.77 | 99.3 | 0.3 a | 0.3 | 0.7 | 11.5 a | 0.1 | 5.3 bc | 0.1 | 21.9 a | 0.5 | 60.5 a | 0.6 | 0.7 a | 0.1 |
| 75 | 49.3 a | 2.7 | 45.7 a | 1.91 | 94.1 | 2.8 d | 1.6 | 5.9 | 11.6 a | 0.1 | 4. 5 a | 0.5 | 20.3 c | 0.8 | 62.9 e | 1.2 | 0.6 a | 0.1 |
| 77 | 47.5 ab | 1.6 | 46.4 a | 1.4 | 97.8 | 1.1 b | 0.3 | 2.2 | 11.9 e | 1.4 | 4.2 a | 0.1 | 23.0 d | 0.5 | 60.0 a | 0.8 | 0.8 a | 0.1 |
| 78 | 45.0 b | 1.3 | 44.6 a | 1.28 | 99.1 | 0.4 a | 0.7 | 0.9 | 9.9 d | 1.3 | 5.8 d | 0.1 | 32.6 f | 0.1 | 50.9 b | 0.1 | 0.8 a | 0.1 |
| 79 | 47.2 ab | 0.7 | 46.2 a | 0.89 | 98.0 | 0.9 b | 0.3 | 2.0 | 10.9 c | 0.9 | 4.1 a | 0.1 | 24.2 e | 0.3 | 59.8 a | 0.4 | 0.9 a | 0.2 |
| Junona | 48.8 a | 3.1 | 46.2 a | 3.29 | 94.5 | 2.6 d | 2.3 | 5.5 | 11.6 a | 3.3 | 5.6 cd | 0.1 | 21.4 a | 0.4 | 60.6 a | 0.7 | 0.7 a | 0.2 |
| Chemical Compounds/OSI | Genotype | Year | Genotype*Year | Other Factors |
|---|---|---|---|---|
| Total lipids | 37.8 * | 19.59 * | 33.3 * | 9.3 |
| Free lipids | 22.5 * | 24.25 * | 47.1 * | 6.1 |
| Bound lipids | 53.5 * | 0.0 | 7.6 * | 38.9 |
| C16:0 | 96.0 * | 0.2 | 2.0 * | 1.8 |
| C18:0 | 92.5 * | 0.0 | 4.2 * | 3.2 |
| C18:1 | 99.2 * | 0.0 | 0.5 * | 0.3 |
| C18:2 | 97.9 * | 0.1 | 1.2 * | 0.7 |
| C18:3 | 23.6 * | 19.3 * | 15.9 * | 41.2 |
| α-tocopherol | 59.4 * | 9.1 * | 30.3 * | 1.2 |
| γ-tocopherol | 37.4 * | 21.5 * | 40.6 * | 0.5 |
| δ-tocopherol | 63.7 * | 3.6 * | 31.2 * | 1.5 |
| Total tocopherols | 41.0 * | 15.1 * | 43.3 * | 0.6 |
| Campesterol | 83.8 * | 2.1 * | 6.5 * | 7.6 |
| Spinasterol + β-sitosterol | 56.9 * | 25.3 * | 10.7 * | 6.6 |
| Δ7,22,25-stigmastatrienol | 74.9 * | 0.5 | 11.2 * | 13.3 |
| Δ7-stigmastenol | 61.0 * | 6.2 * | 24.1 * | 8.7 |
| 7,25-stigmastadienol | 42.9 * | 36.1 * | 10.8 * | 10.1 |
| Other sterols | 47.2 * | 30.8 * | 6.3 * | 15.7 |
| Total sterols | 41.7 * | 31.6 * | 14.1 * | 12.6 |
| Squalene | 80.8 * | 0.2 | 15.5 * | 3.5 |
| OSI | 38. 8 * | 2.8 * | 55.6 * | 2.8 |
| Hybrid/Variety | Tocopherols (mg/100 g) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| α-tocopherol | γ-tocopherol | δ-tocopherol | Total | ||||||||
| S.D. | % in Total | S.D. | % in Total | S.D. | % in Total | S.D. | |||||
| 71 | 0.11 a | 0.09 | 0.4 | 29.98 ab | 9.86 | 90.9 | 2.72 c | 0.45 | 8.7 | 32.8 ab | 10.1 |
| 73 | 2.55 c | 0.89 | 5.6 | 44.37 b | 10.32 | 89.8 | 2.00 bc | 1.50 | 4.6 | 48.9 b | 8.0 |
| 74 | 0.25 a | 0.23 | 0.6 | 33.44 ab | 18.28 | 96.9 | 0.57 ab | 0.23 | 2.5 | 34.3 ab | 18.3 |
| 75 | 2.28 bc | 2.19 | 7.6 | 23.44 a | 3.71 | 91.8 | 0.13 a | 0.14 | 0.6 | 25.8 a | 5.7 |
| 77 | 0.56 ab | 0.18 | 2.2 | 23.43 a | 1.52 | 91.9 | 1.51 abc | 1.47 | 5.9 | 25.5 a | 0.2 |
| 78 | 2.93 c | 1.55 | 7.5 | 34.02 ab | 4.62 | 91.7 | 0.25 a | 0.28 | 0.8 | 37.2 ab | 5.9 |
| 79 | 0.13 a | 0.14 | 0.3 | 29.32 ab | 11.24 | 99.1 | 0.22 a | 0.26 | 0.5 | 29.7 a | 11.6 |
| Junona | 0.43 ab | 0.54 | 1.1 | 37.74 ab | 1.24 | 98.6 | 0.12 a | 0.14 | 0.3 | 38.3 ab | 1.9 |
| Hybrid/Variety | Sterols (mg/100 g) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Campesterol | Spinasterol + β-sitosterol | Δ7,22,25-stigmastatrienol | Δ7-stigmastenol | Δ7,25-stigmastadienol | Not identified | Total | |||||||||||||
| S.D. | %* | S.D. | %* | S.D. | %* | S.D. | % * | S.D. | % * | S.D. | S.D. | ||||||||
| 71 | 0.76 d | 0.10 | 0.2 | 148.93 a | 7.02 | 47.8 | 68.92 c | 0.98 | 22.2 | 12.04 ab | 0.52 | 3.9 | 69.79 ab | 5.73 | 22.4 | 10.81 a | 1.25 | 311 ab | 13 |
| 73 | 0.33 ab | 0.04 | 0.1 | 185.17 b | 16.42 | 56.0 | 60.90 ab | 1.06 | 18.5 | 13.12 a | 0.58 | 4.0 | 57.80 a | 6.01 | 17.4 | 13.15 ab | 1.32 | 330 a | 22 |
| 74 | 0.37 ab | 0.07 | 0.1 | 168.18 ab | 16.11 | 52.4 | 61.80 abc | 2.95 | 19.3 | 13.98 ac | 1.92 | 4.3 | 62.71 ab | 12.52 | 19.3 | 14.55 ab | 2.40 | 322 ab | 0 |
| 75 | 0.55 c | 0.07 | 0.2 | 148.57 a | 14.76 | 47.2 | 64.15 ac | 5.16 | 20.5 | 13.74 a | 2.63 | 4.3 | 73.79 b | 6.09 | 23.5 | 13.66 ab | 3.01 | 314 ab | 24 |
| 77 | 0.25 b | 0.05 | 0.1 | 142.83 a | 2.59 | 51.7 | 54.87 b | 3.64 | 19.9 | 10.05 b | 0.74 | 3.6 | 54.44 a | 4.87 | 19.7 | 10.95 a | 2.17 | 276 b | 9 |
| 78 | 0.39 a | 0.02 | 0.1 | 179.66 b | 5.13 | 53.7 | 62.82 ac | 2.47 | 18.8 | 13.08 a | 0.76 | 3.9 | 68.40 ab | 7.24 | 20.4 | 10.39 a | 3.15 | 335 a | 16 |
| 79 | 0.35 ab | 0.08 | 0.1 | 165.75 ab | 27.01 | 47.1 | 77.12 d | 7.51 | 22.1 | 16.86 c | 2.49 | 4.8 | 75.02 b | 15.20 | 21.2 | 16.41 b | 3.13 | 351 a | 54 |
| Junona | 0.42 a | 0.09 | 0.3 | 150.28 a | 0.14 | 49.3 | 61.68 ab | 2.76 | 20.7 | 12.90 ab | 1.09 | 4.2 | 65.01 ab | 5.30 | 21.3 | 14.25 ab | 0.43 | 304 ab | 11 |
| Hybrid/Variety | Squalene (mg/100 g) | |
|---|---|---|
| S.D. | ||
| 71 | 305 c | 11 |
| 73 | 210 a | 11 |
| 74 | 236 ab | 24 |
| 75 | 260 b | 6 |
| 77 | 215 a | 3 |
| 78 | 228 a | 10 |
| 79 | 233 ab | 13 |
| Junona | 237 ab | 25 |
| Content of Lipid Class | Pearson’s Correlation Coefficient | Share of Selected Fatty Acid | Pearson’s Correlation Coefficient | Content of Antioxidant | Pearson’s Correlation Coefficient |
|---|---|---|---|---|---|
| Free lipids | r = 0.40 | Palmitic | r = −0.48 | α-tocopherol | r = 0.27 |
| Bound lipids | r = −0.14 | Stearic | r = 0.82 * | γ-tocopherol | r = 0.75 * |
| Oleic | r = 0.54 | δ-tocopherol | r = −0.05 | ||
| Linoleic | r = −0.63 | Squalene | r = −0.34 | ||
| α-linolenic | r = 0.42 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tańska, M.; Ogrodowska, D.; Bartoszewski, G.; Korzeniewska, A.; Konopka, I. Seed Lipid Composition of New Hybrids of Styrian Oil Pumpkin Grown in Poland. Agronomy 2020, 10, 1104. https://doi.org/10.3390/agronomy10081104
Tańska M, Ogrodowska D, Bartoszewski G, Korzeniewska A, Konopka I. Seed Lipid Composition of New Hybrids of Styrian Oil Pumpkin Grown in Poland. Agronomy. 2020; 10(8):1104. https://doi.org/10.3390/agronomy10081104
Chicago/Turabian StyleTańska, Małgorzata, Dorota Ogrodowska, Grzegorz Bartoszewski, Aleksandra Korzeniewska, and Iwona Konopka. 2020. "Seed Lipid Composition of New Hybrids of Styrian Oil Pumpkin Grown in Poland" Agronomy 10, no. 8: 1104. https://doi.org/10.3390/agronomy10081104
APA StyleTańska, M., Ogrodowska, D., Bartoszewski, G., Korzeniewska, A., & Konopka, I. (2020). Seed Lipid Composition of New Hybrids of Styrian Oil Pumpkin Grown in Poland. Agronomy, 10(8), 1104. https://doi.org/10.3390/agronomy10081104