1. Introduction
Ornamental plants for garden decoration have been commercialized in spring for quite a number of years. The plants are sold at the beginning of vegetation, or even at the beginning of flowering, and are necessarily grown in pots. One of the essential criteria for their visual quality is their overall shape, inherent to their architectural construction, i.e., the establishment across space of the different aerial organs according to the organization rules specific to each species [
1].
Plant architecture depends on genetic and environmental factors, as well as on their interaction [
2,
3,
4]. Plant shape can thus be genetically controlled by varietal improvement and/or cultivation by environmental control, such as modified light quality [
5], water restriction [
6,
7], or mechanical stimulation [
8]. However, these methods are applied more or less empirically by breeders and horticulturists. More extensive knowledge about the genotype × environment interaction would lead to a more effective control of the plant architecture, and consequently of its shape.
Plant architecture is complex to analyze, particularly in ornamental woody bushes, characterized by different types of axes and several orders of branching [
9,
10]. Its analysis has often been limited to a few architectural characteristics that can be easily measured manually, such as stem length and diameter [
7,
11,
12].
Taking into account the entire plant architecture would make it possible to identify new variables potentially more explanatory of the observed variability. To meet this objective, we developed a method for analyzing the entire plant architecture by 3D digitalization of rose [
9,
13]. This species was chosen for: (i) its architectural complexity, characterized by several orders of branching, (ii) its architectural variability, ranging from a spreading to an upright growth habit, and (iii) its ornamental economic importance [
14].
In this method, the plant architecture is broken down into axes and internods, which can be characterized morphologically (diameter and length), topologically (by determining succession and branching relationships), and geometrically (by characterizing their organization in space) [
9,
10,
13]. An almost exhaustive description of the plant architecture is obtained and generates a large number of variables, many of which are inaccessible manually. For example, two types of axes—short and long—have been identified in rose [
9].
The effect of genetic and environmental factors on rose architecture was demonstrated using this method [
2,
3,
4]. Most architectural variables, such as the number of internodes, are moderately to strongly heritable [
2,
4]. Nevertheless, some of them (e.g., the number of branches, axis length) are highly influenced by the environment, more particularly by light intensity and water supply [
3,
7]. A strong genotype × environment interaction has been demonstrated and can be explained by contrasting or even opposing responses of genotypes to the environment, such as water supply [
2,
3,
4].
Other environmental factors such as light quality strongly influence plant architecture [
15,
16]. Several families of photoreceptors allow the plant to perceive changes in the light quality and thus adapt its development. Red light—red (R; 600–700 nm) and far-red (FR; 700–800 nm)—is perceived by phytochromes, while blue light (B; 400–500 nm) is perceived by several families of photoreceptors, especially cryptochromes [
15,
16].
Elongation, branching, and flowering are particularly affected by red and blue lights [
15,
16]. A decreased R/FR ratio led to increased elongation, reduced branching, and accelerated flowering in chrysanthemum [
5,
17,
18,
19]. The same has been found true for snapdragon [
20]. In contrast, the addition B light inhibited stem elongation in tomato, cucumber, and pepper [
21] and increased the number of flower buds in impatiens [
22].
The effect of red and blue lights on rose architecture has been poorly studied [
11,
12,
23,
24,
25]. Few changes in plant architecture have been observed to date. However, the light spectrum modification devices used in those studies only tested a few wavelength ratios, i.e., the R/FR ratio and B light intensity [
11,
12,
23,
24,
25]. In addition, the effect of these ratios was only evaluated on a few architectural characteristics such as axis length and diameter, without taking the different types of axes and the orders of branching into account, and on one or two varieties, without taking the variability of the genotypic response into account, as often observed in rose [
2,
3,
4].
The light-emitting diode (LED) technology recently made it possible to manipulate the light spectrum, and made it easy to modify B, R and FR wavelength ratios and test a wider range of combinations in many horticultural species [
15,
16]. We conducted a series of tests in rose using a lighting device composed of white (W), R, and FR LEDs to select three light conditions characterized by different B/R and R/FR ratios and test whether they modified the plant architecture.
The objective of this study is to evaluate the effect of these three light conditions on the entire architecture of three varieties of bush rose chosen for their genotype-specific response to the environment [
2,
3,
12].
2. Materials and Methods
2.1. Plant Material
The plant material was composed of three rose varieties with contrasting shapes: Baipome and ‘The Fairy’ (ground cover), chosen for their architectural plasticity, and Knock Out
® Radrazz (upright bush), chosen for its insensitivity to light quality [
2,
3,
12].
The plants were obtained from cuttings taken from mother plants, grown in pots in a greenhouse. The cuttings consisted of a single metamer taken from the median zone of the stems. The cuttings were planted in plugs (35 mm in diameter and 40 mm in height), composed of a non-woven cloth containing a mixture of fine peat and perlite. The rooting phase took place in the greenhouse, under a plastic tunnel. The mean temperature was 22 °C during the day and 18 °C at night, and relative humidity was maintained at saturation by a fine mist humidifier.
The young plants were potted four weeks later in 0.5–l pots, in a substrate composed of peat (50%), perlite (40%), and coconut fiber (10%) and then acclimatized in a greenhouse for one week.
2.2. Experimental Conditions
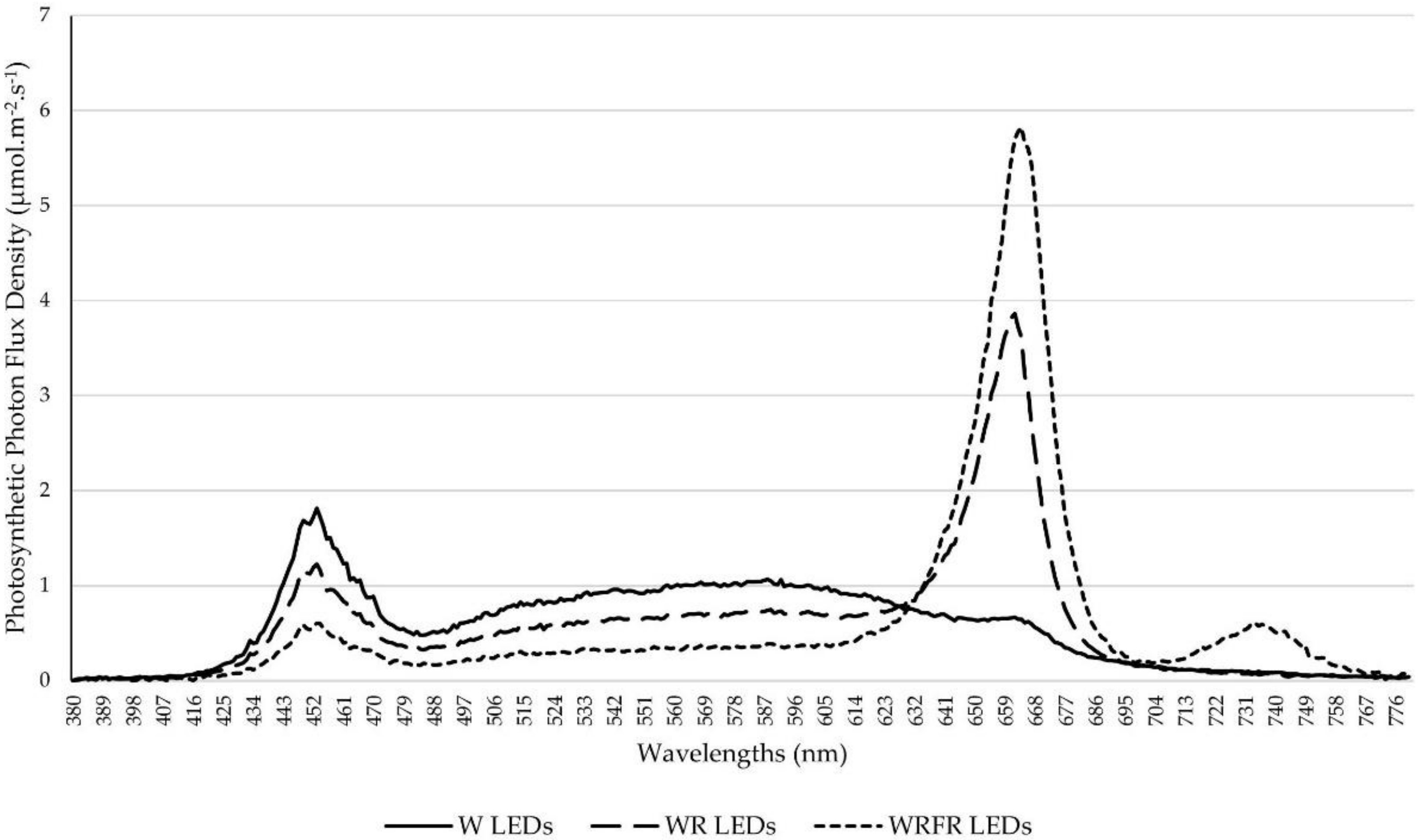
After this acclimatization phase, the plants were grown in a climatic chamber composed of two shelves (3.80 × 0.80 m). Mineral nutrition was provided by fertirrigation, in subirrigation, with a balanced liquid fertilizer (N–P2O5–K2O 3–2–6), a pH of 6.5 and an average electrical conductivity of 1.2 mS cm−1.
The air temperature was maintained at 20 °C during the day and 18 °C at night, with a relative humidity of 70%. The plants were subjected to a photoperiod of 16 h light/8 h dark, with an average photosynthetic photon flux density (PPFD; 400–700 nm) of 224.6 ± 20.1 µmol m
−2 s
−1 (a day light integral of 12.9 mol m
−2 d
−1) provided by a LED lighting device composed of panels of 12 W, R, and FR LED tubes (Cesbron, Saint Sylvain d’Anjou, France) according to the light condition tested. The climatic chamber was divided into three compartments separated by a white opaque panel making it possible to test three light conditions simultaneously, with the following spectral characteristics: the light spectrum of condition 1 (W) was composed of 65.8 µmol m
−2 s
−1 of B (or 29.0% of PPFD; 400–500 nm), 97.0 µmol m
−2 s
−1 of green (G; 42.7%; 500–600 nm), 64.1 µmol m
−2 s
−1 of R (28.3%; 600–700 nm), and 6.6 µmol m
−2 s
−1 of FR (700–800 nm) light on average; condition 2 (WR) was composed of 39.1 µmol m
−2 s
−1 of B (17.8%), 57.8 µmol m
−2 s
−1 of G (26.2%), 123.5 µmol m
−2 s
−1 of R (56.0%), and 5.3 µmol m
−2 s
−1 of FR light; condition 3 (WRFR) was composed of 22.8 µmol m
−2 s
−1 of B (10.1%), 33.3 µmol m
−2 s
−1 of G (14.7%), 170.4 µmol m
−2 s
−1 of R (75.2%), and 22.8 µmol m
−2 s
−1 of FR light (
Figure 1). The PPFD and light spectra were measured using a Rainbow Light MR16 spectrometer (Rainbow Light Technology CO., LTD, Taiwan) placed at the top of pot. The spectral characteristics (PPFD, relative quantum efficiency (RQE), G/B, B/R, and R/FR ratios) of the three light conditions tested are specified in the
Table 1. No significant difference was observed for PPFD and RQE between the light conditions (
Table 1).
The experiment took place from November 2017 to March 2018 and was repeated in spring 2018, from February to May.
For the measurements, nine plants per variety and light condition were randomly selected (in the center of the shelf) at mid-culture (measurement point 1) and at the ‘opened flower of the order 1 axis’ stage, corresponding to the end of cultivation (measurement point 2).
2.3. Description of Plant Shape
At measurement point 2, the plant shape was described from a photograph, using the free image analysis software program ImageJ (National Institutes of Health, USA). A photograph of the widest side of the plant was taken in a standardized way, using a gray background for better color contrast. This photograph was then analyzed using ImageJ software. The convex envelope of the plant was first traced using the ’convex hull selection’ function, and then analyzed using the predefined morphometric variables of the ’set measurements’ function (
Table 2).
2.4. Description of Plant Architecture
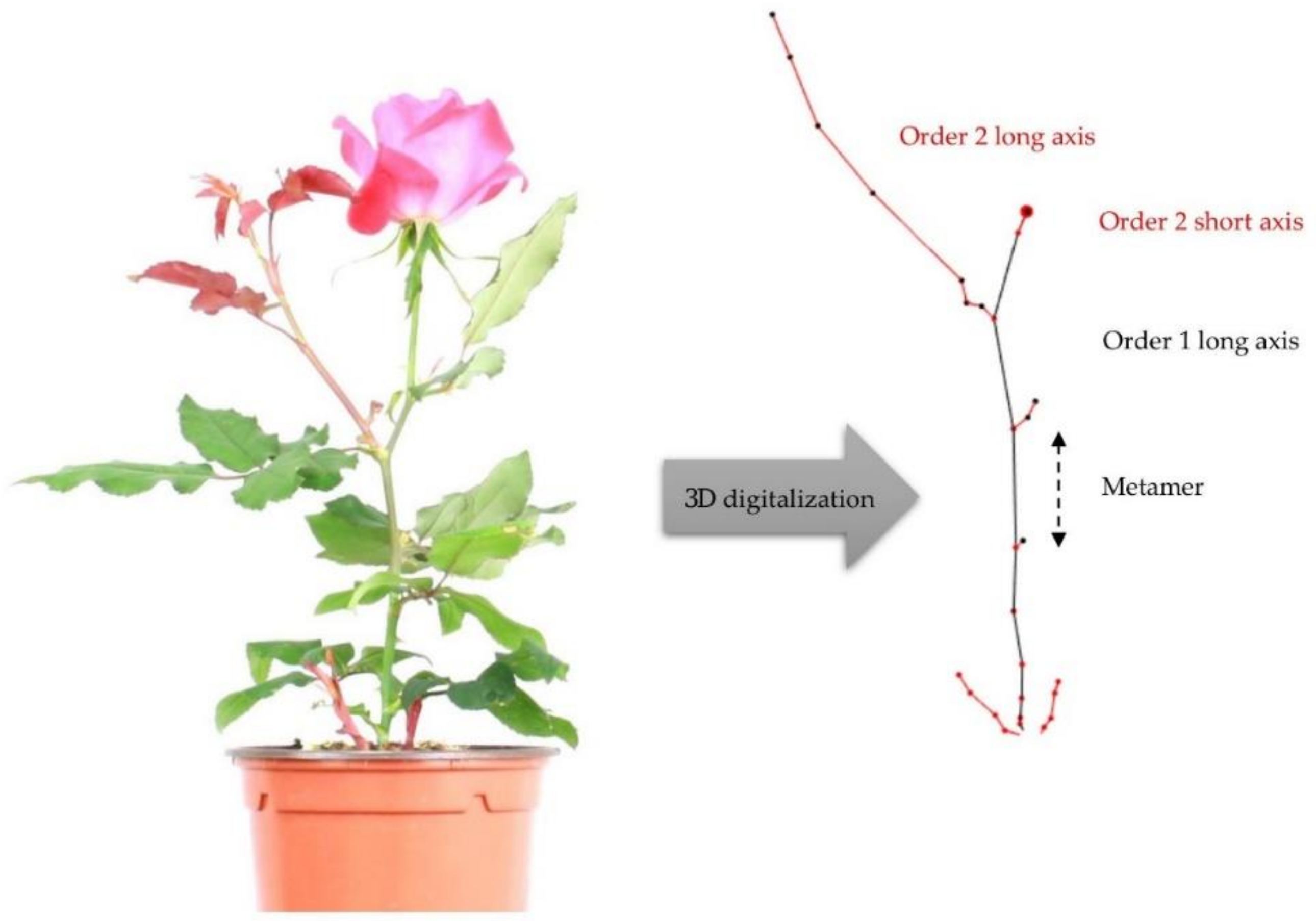
The plant architecture is characterized by two components—the axis and the metamer—and one metamer is composed of one internode, one node, one axillary bud and one leaf (
Figure 2) [
26]. These components have topological (succession or branching) relationships among them. The architectural analysis was carried out at two observation scales—the plant and the axis.
The following variables were measured at measurement point 1: the length (LA1), number of metamers (NbMetA1) and number of branches (NbA2) of the order 1 axis.
The following variables were measured at measurement point 2 (
Table 3):
at the plant scale, the number of axes and determined (i.e., flowering) axes, according to the order of branching, distinguishing the short axes from the long ones as defined by [
9], with short axes comprising one to four metamers and long axes comprising five or more metamers;
at the axis scale, for the two types of axes, their morphology (length and number of metamers), according to the order of branching.
Three categories of variables were thus measured at the plant and axis scale: elongation (length and number of metamers that make up a long and short axis, etc.), branching (number of axes, etc.) and flowering (number of determined axes, etc.).
The architectural measurements were made using a Microscribe 3D digitizer (Solution Technologies, Oella, MD, USA) and the data were saved in an Excel spreadsheet. A database gathering all the measurements made at the two observation scales was built. The architectural variables were extracted from these data using a specially developed R script.
2.5. Data Analysis
The variables, measured at measurement point 2, were grouped into three categories as previously defined, i.e., elongation, branching and flowering for architectural description (
Table 3). They were subjected to a principal component analysis (PCA) to represent all the variables in a set of linear combinations uncorrelated with one another and accounting for an increasingly weaker part of the observed variability. The first two principal components were selected for our analysis. For each of them, the variables that best contributed to their formation were classified, and the best qualified of the above-mentioned categories was selected. This approach was also applied to the morphometric variables used to describe plant shape.
A complementary architectural analysis was carried out to specify the position of branching. Three new variables were measured mainly at the order of branching 2, namely the numbers of order 3 short axes carried by order 2 long axes in apical (NbSA3_Top), median (NbSA3_Med) and basal (NbSA3_Bas) position.
An analysis of variance was carried out to evaluate the genotype effect, the light quality effect and their interaction for the variables measured at measurement points 1 and 2 from a mixed linear model, with the repetition of the experiment as a random factor and for a probability
p < 0.05:
where
P is the phenotypic value of genotype i, for light spectrum j and repetition k;
µ is the mean for all genotypes, light spectra and repetitions;
G is the fixed effect of genotype i;
L is the fixed effect of light spectrum j;
G × L is the fixed effect of their interaction;
r is the random effect of the experimental repetition k;
e is the residual error.
The model was estimated using the maximum likelihood (ML) method. Then, a post-hoc comparison of means (Tukey test) was performed based on the adjusted means (least-square means) for a probability p < 0.05. The statistical analyses were carried out using the lme4 and multcomp R packages (R Foundation for Statistical Computing, Vienna, Austria).
4. Discussion
Morphometric analysis using ImageJ free software is simple to implement from plant photographs. The twelve morphometric variables proposed by ImageJ were reduced to two (surface of the convex envelope (Area) and circularity (Circ)) to describe the observed shape variability using PCA. The study of these variables highlighted an effect of genotype and light quality on plant shape, with a higher surface of the convex envelope under the WRFR condition.
This first approach led us to characterize the effect of light quality and its interaction with the genotype on plant architecture, which determines plant shape. To describe the observed variability, nine architectural variables were selected by PCA. Among these variables, the number of determined axes (NbDetA), the number of order 3 short axes (NbSA3) and the length of the short axes (LSA) had been identified in previous works describing the architecture of: (i) eight rose cultivars with contrasting shapes and (ii) two segregated progenies used for the genetic analysis of rose architecture [
4,
13]. These converging results confirm the robustness of these variables to discriminate rose plants of different architectures. In this work, the architectures of the three varieties with contrasting shapes were perfectly differentiated, with a significant genotype effect for the nine selected variables.
A significant effect of light quality was observed for most of the measured elongation, branching, and flowering variables, regardless of the plant developmental stage, with two groups of light conditions: W and WR on the one hand, and WRFR on the other hand, differentiated spectrally by their B/R and R/FR ratios, while the G/B ratio remained the same.
Rose was insensitive to variations in the B/R ratio, more specifically to the reduction of B and G light intensity in favor of R light. No significant difference between the W and WR light conditions was observed for plant architecture. The same was true for basil [
27]. For light conditions close to ours (B
24G
32R
44 and B
16G
10R
74), no significant differences were observed for leaf area and plant width, or for photosynthetic activity. However, in this study, differences in photosynthetic activity were observed under B–R light compared to B–G–R light for different leaf stages. For these photosynthetic wavelengths, B and R lights are mainly absorbed by the upper leaf stages, while G light penetrates deeper into the canopy and is absorbed by the lower leaf stages [
28,
29]. The decrease in photosynthetic activity of the lower stages under WR compared to W may have been offset by an increase in the activity of the upper stages.
In basil, however, plants were taller under the condition richer in G light and characterized by a high G/B ratio (1.3) [
27]. In these conditions, G light might induce stem elongation and antagonize the B light effects [
28,
30]. This antagonistic effect of G light on B light might explain the absence of architectural modifications between the W and WR light conditions recorded in our study, characterized by a high G/B ratio (1.5).
Under the light condition enriched in FR-WRFR-the plants exhibited: (i) longer short axes (+29.3% compared to W) whatever the order of branching and (ii) a higher number of short axes and flowered short axes, more particularly at the order 3 (+61.4% for NbSA3 and +61.1% for NbDetSA3 compared to W).
The lengthening of the axes and the acceleration of flowering have been observed in many species with low light intensity and a low R/FR ratio—the shade avoidance syndrome [
31]. In
Arabidopsis thaliana and sunflower, stem elongation has been found correlated with increased levels of the growth hormones auxins and gibberellins in the internodes [
32,
33]. In tobacco, elongation has also been found accompanied by increased sugar concentrations in the stems providing the energy and carbon required for growth [
34]. The sink strength of the stem might be favored by gibberellins [
35,
36,
37].
In
Arabidopsis thaliana, flowering was accelerated under low light intensity and a low R/FR ratio [
38], probably involving phytochromes B, D and E [
39,
40], as well as PHYTOCHROME AND FLOWERING TIME 1 (PFT1) [
41]. Flowering was also accelerated under high light intensity and a high R/FR ratio [
38], possibly associated with increased sucrose concentrations in the leaves and the stem apex, and involving photosynthesis. In our study, the acceleration of flowering, observed under the light condition enriched in FR-WRFR-might also be associated with photosynthesis. The presence of FR light might indeed increase the photosynthetic activity: (i) of the lower leaf layers by penetrating deeper into the canopy and distributing light more evenly within the canopy thanks to internode elongation [
42,
43] and (ii) by improving photosystem II efficiency [
44,
45,
46]. Other morphological changes induced by FR light, not measured in this study but observed in other species (petiole elongation, increased leaf area, and increased leaf length/width ratio), might also promote light interception and increase overall photosynthesis [
43,
47,
48].
By contrast with the shade avoidance syndrome, a higher number of branches was observed under the FR-enriched (WRFR) condition, particularly in the apical position. The same was true for chrysanthemum, in which bud break and sugar concentrations increased in the basal stem under a low R/FR ratio [
49]. Sugars might control bud break in many species [
50,
51]. They play a trophic role. In rose, a high metabolic activity of sucrose is indeed associated with bud break and might increase the sink strength of the bud [
52,
53,
54]. Sugars might also play a signaling role and cause many hormonal changes, e.g., (i) auxin export from the bud to the stem, (ii) inhibition of the strigolactone signaling pathway, and thereby of the effects of auxin, and (iii) stimulation of cytokinin biosynthesis [
55,
56]. Bud break might therefore depend on sugar availability for the bud, but also on bud competition with the different sink organs [
55,
57]. Sugar distribution between the different organs might be influenced by light quality in rose. Increased sink strength of the stem apex was indeed observed when FR light was added to R light [
23]. Thus, the higher overall photosynthesis and the sink strength of the stem apex under the WRFR condition might have increased sugar availability for apical buds and favored apical bud break.
A strong genotype × light quality interaction was also observed for the elongation, branching and flowering variables in our study, regardless of the plant developmental stage. As expected, two types of genotype-specific responses occurred. Baipome and ‘The Fairy’, chosen for their architectural plasticity, responded to light quality [
2,
3], while Knock Out
® Radrazz remained insensitive, as previously demonstrated by [
12], even though it can respond to light intensity [
7]. A strong genotype × environment interaction was also observed in previous works for the elongation and branching variables [
2,
3]. Year (i.e., amounts of cumulated radiation)—specific QTLs involved in the control of branching, more particularly order 3 short axes (NbSA3), were highlighted in rose by [
4]. They are believed to be at the origin of the genotype × environment interaction observed in this study. They co-locate with an environmentally sensitive branching repressor,
BRANCHED1.
More in-depth eco-physiological and biochemical investigations are needed to better understand the response of rose to light quality, in particular the lack of response of Knock Out® Radrazz, and thus identify the determinants of the genotype × light quality interaction.
Production of compact, branched plants is requested for ornamental plants. This study shows that acting on light quality makes it possible to modify the entire plant architecture and thereby its shape. Of the three light conditions tested, the addition of FR light, usually used to increase photosynthetic activity and therefore biomass, produced the most branched plants, with relative stem elongation. A light spectrum reinforced in B would limit this effect, as demonstrated in petunia by [
58].
This work was carried out in a climatic chamber, where the light effect was strengthened compared to greenhouse conditions which involve solar radiation. This is exactly the objective targeted by this new culture system, i.e., to reach a total control of the culture cycle, independently from the cycle of seasons, with a very high regularity of the climatic conditions, especially for light and temperature.
These interesting results obtained in rose suggest that this culture system in a totally artificialized environment, where the light quality can be modified, could be applied to other horticultural species. The objective would be to “deseasonalize” production cycles and shorten them, while improving plant architecture, more particularly branching, and thereby commercial quality.

 ,
, 




{kind=link}
{kind=link}
{kind=link}