Weed Seed Bank Diversity in Dryland Cereal Fields: Does it Differ Along the Field and Between Fields with Different Landscape Structure?

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
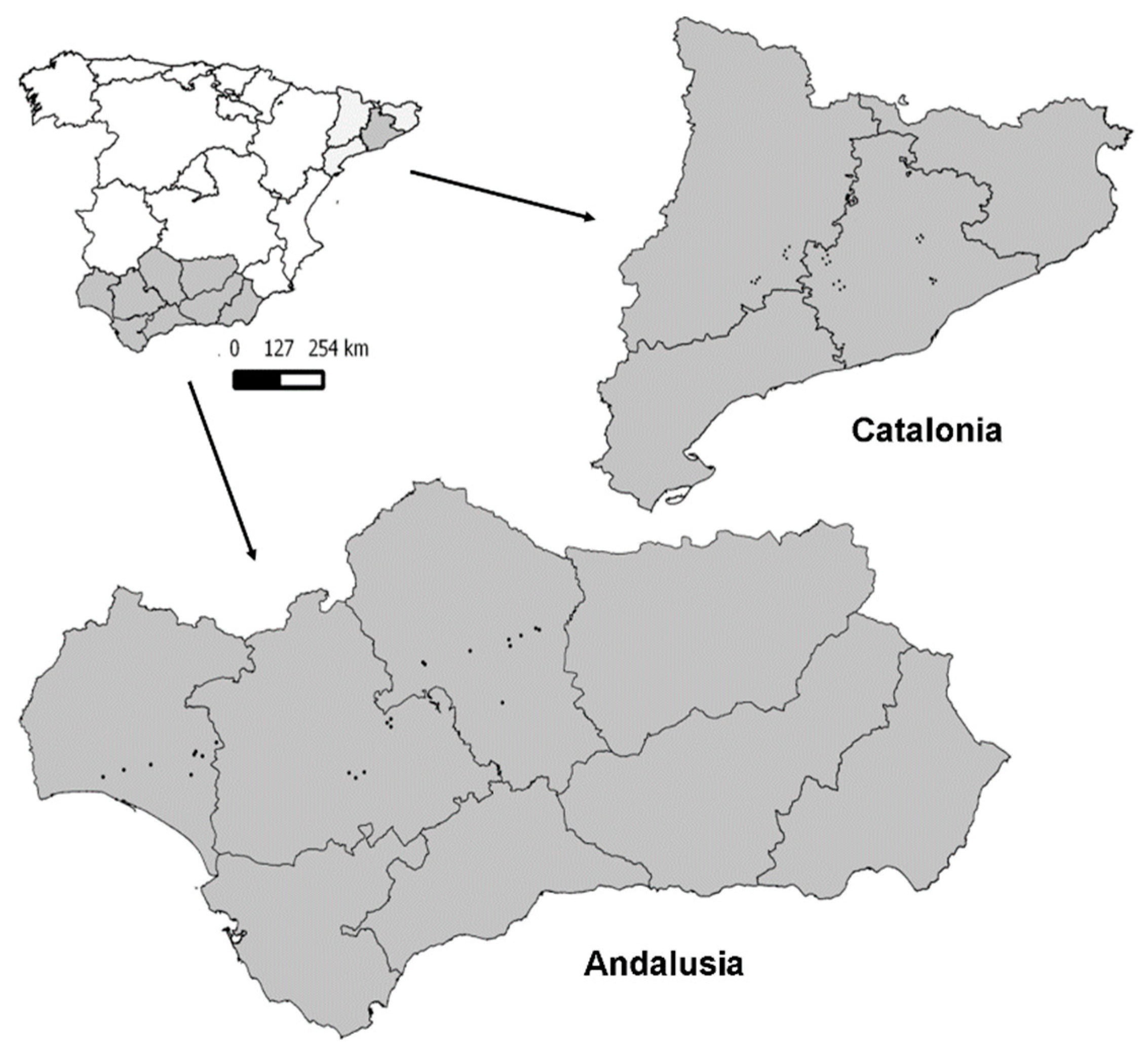
2.1. Study Areas
2.2. Seed Bank Sampling and Seedling Identification
2.3. Weed Functional Traits
2.4. Taxonomic and Functional Diversity Indices
2.5. Statistical Analyses
2.5.1. Taxonomic and Functional Diversity Analyses
2.5.2. Functional Traits Analyses
3. Results
4. Discussion
4.1. Field Position and Soil Properties
4.2. Landscape Configuration
4.3. Higher Species Richness, but Homogeneous Seed Bank Functional Diversity
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Hawes, C.; Squire, G.R.; Hallett, P.D.; Watson, C.A.; Young, M. Arable plant communities as indicators of farming practice. Agric. Ecosyst. Environ. 2010, 138, 17–26. [Google Scholar] [CrossRef]
- Gaba, S.; Perronne, R.; Fried, G.; Gardarin, A.; Bretagnolle, F.; Biju-Duval, L.; Colbach, N.; Cordeau, S.; Fernández-Aparicio, M.; Gauvrit, C.; et al. Response and effect traits of arable weeds in agro-ecosystems: A review of current knowledge. Weed Res. 2017, 57, 123–147. [Google Scholar] [CrossRef]
- Bàrberi, P.; Bocci, G.; Carlesi, S.; Armengot, L.; Blanco-Moreno, J.M.; Sans, F.X. Linking species traits to agroecosystem services: A functional analysis of weed communities. Weed Res. 2018, 58, 76–88. [Google Scholar] [CrossRef]
- José-María, L.; Sans, F.X. Weed seedbanks in arable fields: Effects of management practices and surrounding landscape. Weed Res. 2011, 51, 631–640. [Google Scholar] [CrossRef]
- Poggio, S.L.; Chaneton, E.J.; Ghersa, C.M. Landscape complexity differentially affects alpha, beta, and gamma diversities of plants occurring in fencerows and crop fields. Biol. Conserv. 2010, 143, 2477–2486. [Google Scholar] [CrossRef]
- Cirujeda, A.; Pardo, G.; Mari, A.I.; Aibar, J.; Pallavicini, Y.; Gonzalez-Andujar, J.L.; Recasens, J.; Sole-Senan, X.O. The structural classification of field boundaries in Mediterranean arable cropping systems allows the prediction of weed abundances in the boundary and in the adjacent crop. Weed Res. 2019, 59, 300–311. [Google Scholar] [CrossRef]
- Roschewitz, I.; Gabriel, D.; Tscharntke, T.; Thies, C. The effects of landscape complexity on arable weed species diversity in organic and conventional farming. J. Appl. Ecol. 2005, 42, 873–882. [Google Scholar] [CrossRef]
- José-María, L.; Armengot, L.; Blanco-Moreno, J.M.; Bassa, M.; Sans, F.X. Effects of agricultural intensification on plant diversity in Mediterranean dryland cereal fields. J. Appl. Ecol. 2010, 47, 832–840. [Google Scholar] [CrossRef]
- Gonzalez-Andujar, J.L. Co-operative versus non co-operative farmers’ weed control decisions in an agricultural landscape. Weed Res. 2018, 58, 327–330. [Google Scholar] [CrossRef]
- Gonzalez-Diaz, L.; Van den Berg, F.; Van Den Bosch, F.; González-Andújar, J.L. Controlling annual weeds in cereals by deploying crop rotation at the landscape scale: Avena sterilis as an example. Ecol. Appl. 2012, 22, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Le Coeur, D.; Baudry, J.; Burel, F.; Thenail, C. Why and how we should study field boundary biodiversity in an agrarian landscape context. Agric. Ecosyst. Environ. 2002, 89, 23–40. [Google Scholar] [CrossRef]
- Geographic Information System for Agricultural Parcels. Available online: http://www.alz.org/what-is-dementia.asp (accessed on 24 June 2019).
- Fried, G.; Kazakou, E.; Gaba, S. Trajectories of weed communities explained by traits associated with species’ response to management practices. Agric. Ecosyst. Environ. 2012, 158, 147–155. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Eastwood, A. Shifts in functional traits and functional diversity between vegetation and seed bank. J. Veg. Sci. 2013, 24, 865–876. [Google Scholar] [CrossRef]
- Bastida, F.; Macias, F.J.; Butler, I.; Gonzalez-Andujar, J.L. Achene dimorphism and protracted release: A trait syndrome allowing continuous reshaping of the seed dispersal kernel in the Mediterranean species Pallenis spinose. Plant. Ecol. Divers. 2018, 11, 429–439. [Google Scholar] [CrossRef]
- Kenkel, N.C.; Derksen, D.A.; Thomas, A.G.; Watson, P.R. Review: Multivariate analysis in weed science research. Weed Sci. 2002, 50, 281–292. [Google Scholar] [CrossRef]
- Bocci, G. TR8: An R package for easily retrieving plant species traits. Methods Ecol. Evol. 2015, 6, 347–350. [Google Scholar] [CrossRef]
- Royal Botanic Gardens Kew, Seed Information Database (SID). Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 18 December 2019).
- Blanca, G.; Cabezudo, B.; Cueto, M.; Morales-Torres, C.; Salazar, C. Flora Vascular de Andalucía Oriental, 2nd ed.; Universidades de Almería: Granada, Spain; Jaén, Spain; Málaga, Spain, 2011. [Google Scholar]
- De Bolòs, O.; Bonada, J.V. Flora dels Països Catalans, 1st ed.; Institut d’Estudis Catalans: Barcino, Spain, 1984. [Google Scholar]
- Botta-Dukát, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Mason, N.W.H.; de Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S.A. Guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veg. Sci. 2012, 24, 794–806. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R Package Version 1.0–12; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M. The vegan package. Community Ecol. Package 2007, 10, 631–637. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer Science & Business Media: New York, NY, USA, 2002. [Google Scholar]
- Dray, S.; Choler, P.; Dolédec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Bates, D.M.; Mächler, M.; Bolker, B.M.; Walker, S. Package Lme4: Linear Mixed-Effects Models Using Eigen and S4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar]
- Rodríguez, C.; Wiegand, K. Evaluating the trade-off between machinery efficiency and loss of biodiversity-friendly habitats in arable landscapes: The role of field size. Agric. Ecosyst. Environ. 2009, 129, 361–366. [Google Scholar] [CrossRef]
- Armengot, L.; José-María, L.; Blanco-Moreno, J.M.; Romero-Puente, A.; Sans, F.X. Landscape and land-use effects on weed flora in Mediterranean cereal fields. Agric. Ecosyst. Environ. 2011, 142, 311–317. [Google Scholar] [CrossRef]
- Marshall, E.J.P. The impact of landscape structure and sown grass margin strips on weed assemblages in arable crops and their boundaries. Weed Res. 2009, 49, 107–115. [Google Scholar] [CrossRef]
- Aparicio, A. Descriptive analysis of the ‘relictual’ Mediterranean landscape in the Guadalquivir River valley (southern Spain): A baseline for scientific research and the development of conservation action plans. Biodivers. Conserv. 2007, 17, 2219–2232. [Google Scholar] [CrossRef]
- Instituto de Estadística y Cartografía de Andalucía. Anuario estadístico de Andalucía; Junta de Andalucía: Sevilla, Spain, 2018. [Google Scholar]
- Pinke, G.; Gunton, R.M. Refining rare weed trait syndromes along arable intensification gradients. J. Veg. Sci. 2014, 25, 978–989. [Google Scholar] [CrossRef]


{kind=link}
| Andalusia | Catalonia | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Abb. | Position | Mean ± SD | Min. | Max. | Mean ± SD | Min. | Max. |
| Organic Nitrogen (%) | N | Margin | 0.12 ± 0.01 | 0.04 | 0.24 | 0.19 ± 0.02 | 0.06 | 0.40 |
| Edge | 0.10 ± 0.01 | 0.04 | 0.17 | 0.17 ± 0.03 | 0.09 | 0.50 | ||
| Core | 0.09 ± 0.01 | 0.04 | 0.15 | 0.18 ± 0.02 | 0.07 | 0.70 | ||
| Clay (%) | C | Margin | 23.90 ± 1.94 | 10.4 | 44.2 | 15.70 ± 0.82 | 9.00 | 23.00 |
| Edge | 25.99 ± 2.61 | 8.20 | 61.3 | 17.30 ± 0.73 | 10.30 | 22.80 | ||
| Core | 29.60 ± 2.75 | 12.00 | 61.6 | 18.44 ± 0.88 | 11.70 | 27.90 | ||
| Arable land cover (%) | AL | 61.30 ± 34.89 | 6.00 | 100 | 75.60 ± 22.66 | 25.00 | 100 | |
| Andalusia | Catalonia | ||||||
|---|---|---|---|---|---|---|---|
| Functional Traits | Abbreviation | Mean ± SD | Min. | Max. | Mean ± SD | Min. | Max. |
| Plant height (m) | PH | 0.60 ± 0.40 | 0.07 | 2.00 | 0.54 ± 0.30 | 0.12 | 2.00 |
| Seed mass (mg) | SM | 1.99 ± 3.70 | 0.01 | 19.90 | 1.90 ± 4.10 | 0.02 | 19.90 |
| Flowering onset (month, January = 1) | FO | 3.50 ± 1.80 | 1 | 12 | 4.21 ± 1.60 | 1 | 7 |
| Flowering duration (months) | F | 5.40 ± 2.90 | 1 | 12 | 4.90 ± 2.70 | 1 | 12 |
| Raunkiær’s life forms | LF | Geophytes = 3 | Chamaephytes = 2 | ||||
| Hemicryptophytes = 11 | Geophytes = 1 | ||||||
| Therophytes = 63 | Hemicryptophytes = 5 | ||||||
| Therophytes = 54 | |||||||
| Growth form | GF | Forbs = 61 | Forbs = 50 | ||||
| Graminoids = 16 | Graminoids = 12 | ||||||
| Pollen vector | PT | Anemo/entomogamous = 5 | Anemo/entomogamous = 2 | ||||
| Anemogamous = 21 | Anemogamous = 13 | ||||||
| Autogamous = 14 | Autogamous = 15 | ||||||
| Entomogamous = 37 | Entomo/autogamous = 8 | ||||||
| Entomogamous = 24 | |||||||
| Seed dispersal type | DT | Anemochorous = 24 | Anemochorous = 15 | ||||
| Barochorous = 48 | Barochorous = 38 | ||||||
| Zoochorous = 5 | Zoochorous = 9 | ||||||
| Andalusia | Catalonia | ||||||
|---|---|---|---|---|---|---|---|
| Indices | FP | Mean ± SD | Min. | Max. | Mean ± SD | Min. | Max. |
| Richness (S) | Total | 18.5 ± 8.51 | 3 | 41 | 13.9 ± 5.50 | 5 | 31 |
| Margin | 22.4 ± 8.29 | 8 | 41 | 15.5 ± 4.50 | 7 | 31 | |
| Edge | 18.6 ± 6.80 | 6 | 35 | 13.7 ± 4.90 | 5 | 25 | |
| Core | 14.9 ± 8.91 | 3 | 39 | 12.5 ± 4.50 | 7 | 24 | |
| Exponential Shannon (eH) | Total | 7.66 ± 3.94 | 1.40 | 16.80 | 6.6 ±2.60 | 1.95 | 12.20 |
| Margin | 8.89 ± 3.80 | 2.80 | 16.80 | 7.3 ± 2.45 | 3.00 | 11.30 | |
| Edge | 7.53 ± 3.43 | 1.40 | 18.70 | 6.6 ± 2.60 | 2.25 | 12.20 | |
| Core | 6.55 ± 4.35 | 2.20 | 19.50 | 5.8 ± 2.40 | 1.95 | 10.20 | |
| Evenness (J) | Total | 0.69 ± 0.17 | 0.11 | 0.96 | 0.7 ± 0.10 | 0.48 | 0.92 |
| Margin | 0.69 ± 0.15 | 0.30 | 0.88 | 0.7 ± 0.10 | 0.48 | 0.92 | |
| Edge | 0.68 ± 0.17 | 0.11 | 0.86 | 0.7 ±0.14 | 0.35 | 0.92 | |
| Core | 0.69 ± 0.18 | 0.29 | 0.96 | 0.6 ± 0.16 | 0.30 | 0.91 | |
| Seedling density (D; plants m−2) | Total | 249 ± 393.90 | 2.13 | 2864.00 | 86.2 ± 78.6 | 9.90 | 329 |
| Margin | 278 ± 297.90 | 11.70 | 1026.00 | 85.1 ± 84.95 | 19.10 | 268 | |
| Edge | 294.4 ± 584.1 | 7.40 | 2864.00 | 79.1 ± 76.30 | 9.90 | 329 | |
| Core | 174.5 ± 209.0 | 2.13 | 745.20 | 94.4 ± 76.70 | 19.50 | 268 | |
| Rao’s quadraticentropy index (FDI) | Total | 0.05 ± 0.02 | 0.00 | 0.08 | 0.04 ± 0.020 | 0.01 | 0.08 |
| Margin | 0.05 ± 0.02 | 0.00 | 0.08 | 0.05 ± 0.02 | 0.02 | 0.08 | |
| Edge | 0.05 ± 0.02 | 0.01 | 0.08 | 0.04 ± 0.02 | 0.01 | 0.07 | |
| Core | 0.04 ± 0.02 | 0.02 | 0.08 | 0.04 ± 0.02 | 0.01 | 0.07 | |
| Andalusia | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| null | FP | N | C | AL | N:FP | C:FP | AL:FP | AICc | ∆i | wi | |
| S | x | 188.60 | 0.00 | 0.33 | |||||||
| x | x | 189.65 | 1.04 | 0.20 | |||||||
| x | x | x | 189.97 | 1.37 | 0.17 | ||||||
| x | x | 190.15 | 1.55 | 0.15 | |||||||
| x | x | x | 190.25 | 1.65 | 0.15 | ||||||
| eH’ | x | x | 151.04 | 0.00 | 0.17 | ||||||
| x | 151.73 | 0.69 | 0.12 | ||||||||
| J | x | −89.98 | 0.00 | 0.37 | |||||||
| D | x | 493.90 | 0.69 | 0.34 | |||||||
| FDI | x | −363.80 | 0.00 | 0.37 | |||||||
| Catalonia | |||||||||||
| S | x | x | x | x | x | 123.75 | 0.00 | 0.32 | |||
| x | x | x | 124.71 | 0.96 | 0.20 | ||||||
| x | x | x | 124.79 | 1.04 | 0.19 | ||||||
| x | x | x | 125.22 | 1.47 | 0.15 | ||||||
| x | x | 125.42 | 1.67 | 0.14 | |||||||
| eH’ | x | x | 98.95 | 0.00 | 0.25 | ||||||
| x | x | 99.34 | 0.39 | 0.21 | |||||||
| x | 99.39 | 0.44 | 0.20 | ||||||||
| x | x | x | 100.37 | 1.42 | 0.12 | ||||||
| x | x | x | 100.44 | 1.49 | 0.12 | ||||||
| x | x | 100.85 | 1.90 | 0.10 | |||||||
| J | x | −120.10 | 0.00 | 0.54 | |||||||
| D | x | 332.17 | 0.00 | 0.31 | |||||||
| FDI | x | −444.50 | 0.00 | 0.65 |
| Andalusia | ||||
|---|---|---|---|---|
| Estimate | UnSe | lower CI | upper CI | |
| Richness | ||||
| Intercept | 4.25 | 0.39 | 3.48 | 5.04 |
| FP (core) | −0.33 | 0.49 | −1.32 | 0.06 |
| FP (margin) | 0.48 | 0.38 | −0.27 | 1.25 |
| C | 0.00 | 0.01 | −0.04 | 0.02 |
| C:FP (core) | −0.30 | 0.01 | −0.07 | 0.00 |
| C:FP (margin) | −0.02 | 0.00 | −0.06 | 0.02 |
| AL | 0.00 | 0.00 | −0.00 | 0.01 |
| Catalonia | ||||
| Richness | ||||
| Intercept | 4.38 | 0.62 | 3.13 | 5.63 |
| AL | −0.01 | 0.00 | −0.02 | 0.00 |
| C | 0.00 | 0.03 | −0.06 | 0.07 |
| N | 1.61 | 0.84 | −0.08 | 3.31 |
| FP (core) | 0.02 | 0.47 | −0.93 | 0.09 |
| FP (margin) | 0.90 | 0.70 | −0.50 | 2.30 |
| C:FP (core) | −0.01 | 0.03 | −0.08 | 0.04 |
| C:FP (margin) | −0.07 | 0.03 | −0.13 | 0.01 |
| Exponential Shannon | ||||
| Intercept | 2.66 | 0.33 | 2.00 | 3.30 |
| N | 1.07 | 0.69 | −0.31 | 2.46 |
| FP (core) | −0.21 | 0.10 | −0.42 | 0.00 |
| FP (margin) | 0.08 | 0.10 | −0.13 | 0.29 |
| AL | 0.00 | 0.00 | −0.01 | 0.00 |
| C | −0.01 | 0.01 | −0.04 | 0.01 |
| PH | SM | MFF | F | LF | GF | PT | DT | |
|---|---|---|---|---|---|---|---|---|
| Andalusia | ||||||||
| FP | F = 2061.90 | F = 309.43 | F = 9.97 | F = 383.70 | χ2 = 1403.70 | χ2 = 1728.50 | χ2 = 892.90 | χ2 = 1802.10 |
| N | r = 0.03 | r = 0.01 | r = −0.06 | r = 0.08 | F = 60.50 | F = 335.12 | F = 360.46 | F = 647.17 |
| C | r = 0.21 | r = −0.01 | r = −0.06 | r = 0.00 | F = 27.90 | F = 785.60 | F = 186.88 | F = 226.60 |
| AL | r = 0.02 | r = 0.10 | r = −0.05 | r = 0.08 | F = 390.03 | F = 330.89 | F = 49.40 | F = 436.63 |
| Catalonia | ||||||||
| FP | F = 5.59 | F = 2.70 | F = 33.88 | F = 3.39 | χ2 = 113.30 | χ2 = 36.01 | χ2 = 109.10 | χ2 = 89.22 |
| N | r = 0.04 | r = 0.09 | r = −0.02 | r = −0.09 | F = 0.27 | F = 130.16 | F = 86.26 | F = 56.28 |
| C | r = −0.10 | r = −0.11 | r = 0.00 | r = 0.06 | F = 2.80 | F = 526.63 | F = 58.01 | F = 138.34 |
| AL | r = 0.08 | r = −0.05 | r = 0.17 | r = 0.12 | F = 47.70 | F = 172.06 | F = 103.40 | F = 140.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallavicini, Y.; Hernandez Plaza, E.; Bastida, F.; Izquierdo, J.; Gallart, M.; Gonzalez-Andujar, J.L. Weed Seed Bank Diversity in Dryland Cereal Fields: Does it Differ Along the Field and Between Fields with Different Landscape Structure? Agronomy 2020, 10, 575. https://doi.org/10.3390/agronomy10040575
Pallavicini Y, Hernandez Plaza E, Bastida F, Izquierdo J, Gallart M, Gonzalez-Andujar JL. Weed Seed Bank Diversity in Dryland Cereal Fields: Does it Differ Along the Field and Between Fields with Different Landscape Structure? Agronomy. 2020; 10(4):575. https://doi.org/10.3390/agronomy10040575
Chicago/Turabian StylePallavicini, Yesica, Eva Hernandez Plaza, Fernando Bastida, Jordi Izquierdo, Montserrat Gallart, and Jose L. Gonzalez-Andujar. 2020. "Weed Seed Bank Diversity in Dryland Cereal Fields: Does it Differ Along the Field and Between Fields with Different Landscape Structure?" Agronomy 10, no. 4: 575. https://doi.org/10.3390/agronomy10040575
APA StylePallavicini, Y., Hernandez Plaza, E., Bastida, F., Izquierdo, J., Gallart, M., & Gonzalez-Andujar, J. L. (2020). Weed Seed Bank Diversity in Dryland Cereal Fields: Does it Differ Along the Field and Between Fields with Different Landscape Structure? Agronomy, 10(4), 575. https://doi.org/10.3390/agronomy10040575