

Exclusive Biosynthesis of Pullulan Using Taguchi’s Approach and Decision Tree Learning Algorithm by a Novel Endophytic Aureobasidium pullulans Strain


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Endophytic Yeast-like Fungus
2.2. Identification
2.2.1. Morphological Characteristics
2.2.2. Molecular Identification
2.3. Fermentation Medium and Cultural Conditions
2.4. Determination of Pullulan
2.5. Taguchi Experimental Design
2.6. Decision Tree Learning Algorithm
2.7. Validation of Taguchi and DT Models
2.8. Spectroscopic Characterization
2.9. Software and Statistical protocol
3. Results and Discussion
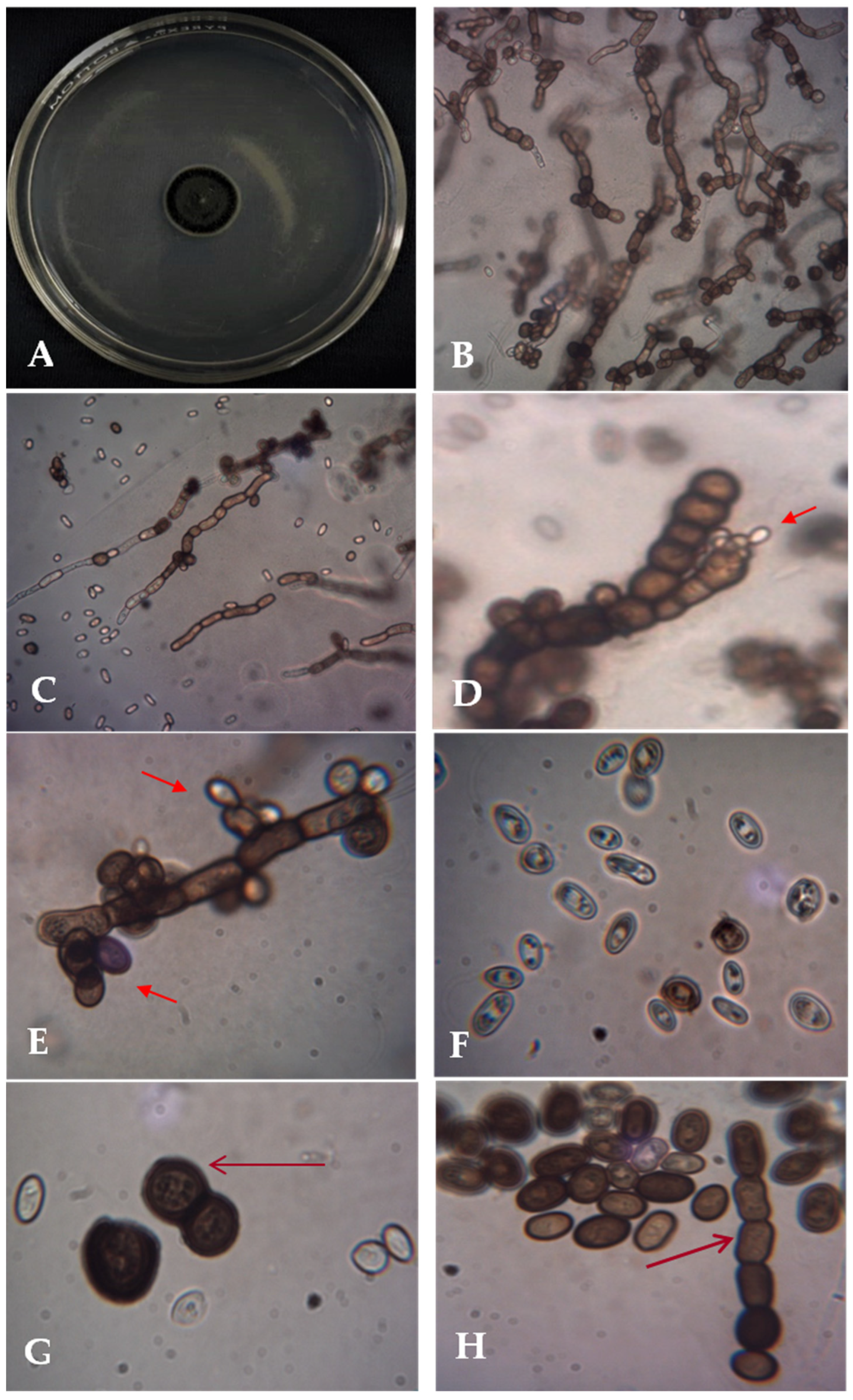
3.1. Isolation and Morphological Identification of A. pullulans
3.2. Molecular Identification of A. pullulans
3.3. Pullulan Biosynthesis Using Taguchi Approach
3.3.1. Determination of Significant Factors
3.3.2. Assignment of the Optimum Factors’ Levels
3.3.3. Residual Analysis
3.4. Decision Tree Learning Algorithm
3.4.1. Decision Tree Selection
3.4.2. The Relative Variable Importance
3.5. Validation of Taguchi and DT Models
3.6. The Structure Characterization of Pullulan
3.6.1. The FT-IR Spectroscopy
3.6.2. The 1H-NMR Spectroscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrews, J.H.; Spear, R.N.; Nordheim, E.V. Population biology of Aureobasidium pullulans on apple leaf surfaces. Can. J. Microbiol. 2002, 48, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Olstorpe, M.; Schnurer, J.; Passoth, V. Microbial changes during storage of moist crimped cereal barley grain under Swedish farm conditions. Anim. Feed Sci. Technol. 2010, 156, 37–46. [Google Scholar] [CrossRef]
- Grube, M.; Schmid, F.; Berg, G. Black fungi and associated bacterial communities in the phyllosphere of grapevine. Fungal Biol. 2011, 115, 978–986. [Google Scholar] [CrossRef]
- Maciel, N.O.P.; Pilo, F.B.; Freitas, L.F.D.; Gomes, F.C.O.; Johann, S.; Nardi, R.M.D.; Lachance, M.A.; Rosa, C.A. The diversity and antifungal susceptibility of the yeasts isolated from coconut water and reconstituted fruit juices in Brazil. Int. J. Food Microbiol. 2013, 160, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Göksungur, Y.; Uҫan, A.; Güvenҫ, U. Production of pullulan from beet molasses and synthetic medium by Aureobasidium pullulans. Yurk. J. Biol. 2004, 28, 23–30. [Google Scholar]
- Gostinčar, C.; Ohm, R.A.; Kogej, T.; Sonjak, S.; Turk, M.; Zajc, J.; Zalar, P.; Grube, M.; Sun, H.; Han, J.; et al. Genome sequencing of four Aureobasidium pullulans varieties: Biotechnological potential, stress tolerance, and description of new species. BMC Genomics 2014, 15, 549. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Wang, F.; Chi, Z.; Yue, L.; Liu, G.; Zhang, T. Bioproducts from Aureobasidium pullulans, a biotechnologically important yeast. Appl. Microbiol. Biotechnol. 2009, 82, 793–804. [Google Scholar] [CrossRef]
- Seo, H.P.; Son, C.W.; Chung, C.H.; Jung, D.I.; Kim, S.K.; Gross, R.A.; Kaplan, D.L.; Lee, J.W. Production of high molecular weight pullulan by Aureobasidium pullulans HP-2001 with soybean pomace as a nitrogen source. Bioresour. Technol. 2004, 95, 293–299. [Google Scholar] [CrossRef]
- Shingel, K.I. Current knowledge on biosynthesis, biological activity and chemical modification of the exopolysaccharide, pullulan. Carbohydr. Res. 2004, 339, 447–460. [Google Scholar] [CrossRef]
- Rehm, B.H.A. Microbial Production of Biopolymers and Polymers Precursors; Caister Academic Press: Poole, UK, 2009; p. 230. [Google Scholar] [CrossRef]
- Wei, X.; Liu, G.L.; Jia, S.L.; Chi, Z.; Hu, Z.; Chi, Z.M. Pullulan biosynthesis and its regulation in Aureobasidium spp. Carbohydr. Polym. 2021, 251, 117076. [Google Scholar] [CrossRef]
- Ravella, S.R.; Quiñones, T.S.; Retter, A.; Heiermann, M.; Amon, T.; Hobbs, P.J. Extracellular polysaccharide (EPS) production by a novel strain of yeast-like fungus Aureobasidium pullulans. Carbohydr. Polym. 2010, 82, 728–732. [Google Scholar] [CrossRef]
- Srikanth, S.; Swathi, M.; Tejaswini, M.; Sharmila, G.; Muthukumaran, C.; Jaganathan, M.K.; Tamilarasan, K. Statistical optimization of molasses based exopolysaccharide and biomass production by Aureobasidium pullulans MTCC 2195. Biocatal. Agric. Biotechnol. 2014, 3, 7–12. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; Zhao, S.; Zhou, Q.; Xin, X.; Chen, L. Statistical optimization of medium for pullulan production by Aureobasidium pullulans NCPS2016 using fructose and soybean meal hydrolysates. Molecules 2018, 23, 1334. [Google Scholar] [CrossRef] [PubMed]
- Asghar, A.; Abdul Raman, A.A.; Daud, W.M.A.W. A comparison of central composite design and Taguchi method for optimizing Fenton process. Sci. World J. 2014, 2014, 869120. [Google Scholar] [CrossRef]
- El-Moslamy, S.H.; Elkady, M.F.; Rezk, A.H.; Abdel-Fattah, Y.R. Applying Taguchi design and large-scale strategy for mycosynthesis of nano-silver from endophytic Trichoderma harzianum SYA. F4 and its application against phytopathogens. Sci. Rep. 2017, 7, 45297. [Google Scholar] [CrossRef] [PubMed]
- Moussa, Z.; Ghoniem, A.A.; Elsayed, A.; Alotaibi, A.S.; Alenzi, A.M.; Hamed, S.E.; Elattar, K.M.; Saber, W.I. Innovative binary sorption of Cobalt (II) and methylene blue by Sargassum latifolium using Taguchi and hybrid artificial neural network paradigms. Sci. Rep. 2022, 12, 18291. [Google Scholar] [CrossRef]
- Kosev, V.; Vasileva, V.; Kusvuran, A. Orthogonal regressions of pea (Pisum L.) varieties. Turkish J. Field Crop. 2018, 23, 159–166. [Google Scholar] [CrossRef]
- Goodman, K.E.; Lessler, J.; Cosgrove, S.E.; Harris, A.D.; Lautenbach, E.; Han, J.H.; Milstone, A.M.; Massey, C.J.; Tamma, P.D. A clinical decision tree to predict whether a bacteremic patient is infected with an extended-spectrum β-lactamase–producing organism. Clin. Infect. Dis. 2016, 63, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Ashino, K.; Sugano, K.; Amagasa, T.; Ying, B.W. Predicting the decision making chemicals used for bacterial growth. Sci. Rep. 2019, 9, 7251. [Google Scholar] [CrossRef] [PubMed]
- Rashad, E.M.; Shaheen, D.M.; Al-Askar, A.A.; Ghoneem, K.M.; Arishi, A.A.; Hassan, E.S.A.; Saber, W.I.A. Seed endophytic Achromobacter sp. F23KW as a promising growth promoter and biocontrol of Rhizoctonia root rot of fenugreek. Molecules 2022, 27, 5546. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: St. Paul, MN, USA, 1998; p. 218. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T. Compendium of Soil Fungi; Academic Press: London, UK, 1993; ISBN 3980308383. [Google Scholar]
- Hermanides-Nijhof, E.J. Aureobasidium and allied genera. Stud. Mycol. 1977, 15, 141–166. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Singh, R.S.; Singh, H.; Saini, G.K. Response surface optimization of the critical medium components for pullulan production by Aureobasidium pullulans FB-1. App. Biochem. Biotechnol. 2009, 152, 42–53. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Gao, F.P.; Liu, L.R.; Li, X.M.; Zhou, Z.M.; Yang, X.D.; Zhang, Q.Q. Pullulan acetate nanoparticles prepared by solvent diffusion method for epirubicin chemotherapy. Colloids Surf. B Biointerfaces 2009, 71, 19–26. [Google Scholar] [CrossRef]
- Thirumavalavan, K.; Manikkandan, T.R.; Dhanasekar, R. Pullulan production from coconut by-products by Aureobasidium pullulans. Afr. J. Biochem. 2009, 8, 254–258. [Google Scholar]
- Al-Askar, A.A.; Saber, W.I.A.; Ghoneem, K.M.; Hafez, E.E.; Ibrahim, A.A. Crude citric acid of Trichoderma asperellum: Tomato growth promotor and suppressor of Fusarium oxysporum f. sp. lycopersici. Plants 2021, 10, 222. [Google Scholar] [CrossRef]
- Inglis, P.W.; Tigano, M.S. Identification and taxonomy of some entomopathogenic Paecilomyces spp. (Ascomycota) isolates using rDNA-ITS sequences. Genet. Mol. Biol. 2006, 29, 132–136. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Al-Askar, A.A.; Rashad, E.M.; Ghoneem, K.M.; Mostafa, A.A.; Al-Otibi, F.O.; Saber, W.I.A. Discovering Penicillium polinicum with high-lytic capacity on Helianthus tuberosus tubers: Oil-based preservation for mold management. Plants 2021, 10, 413. [Google Scholar] [CrossRef]
- Saber, W.I.; El-Naggar, N.E.; El-Hersh, M.S.; El-Khateeb, A.Y. An innovative synergism between Aspergillus oryzae and Azotobacter chroococcum for bioconversion of cellulosic biomass into organic acids under restricted nutritional conditions using multi-response surface optimization. Biotechnology 2015, 14, 47–57. [Google Scholar] [CrossRef]
- El-Hersh, M.S.; Saber, W.I.; El-Fadaly, H.A. Amino acids associated with optimized alkaline protease production by Bacillus subtilis ATCC 11774 using statistical approach. Biotechnology 2014, 13, 252–262. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; El-Khateeb, A.Y.; Ghoniem, A.A.; El-Hersh, M.S.; Saber, W.I. Innovative low-cost biosorption process of Cr6+ by Pseudomonas alcaliphila NEWG-2. Sci. Rep. 2020, 10, 14043. [Google Scholar] [CrossRef]
- Chiou, A.H.; Wu, W.F.; Chen, D.Y.; Hsu, C.Y. The use of the grey-Taguchi method for the optimization of a silicon nanowires array synthesized using electroless Ag-assisted etching. J. Nanopart. Res. 2013, 15, 1866. [Google Scholar] [CrossRef]
- Zainal, N.A.; Shukor, S.R.A.; Razak, K.A. Applying the Taguchi method to optimise the size of silica nanoparticles entrapped with rifampicin for a drug delivery system. J. Eng. Sci. 2015, 11, 9–16. [Google Scholar]
- Machado, M.A.; de Oliveira, R.B.; Oliveira, H. Magnesium and sulfate uptake by the ectomycorrhizal fungus Hebeloma cylindrosporum. Mycorrhiza 2010, 20, 327–334. [Google Scholar]
- Kavanagh, K. Fungi: Biology and Applications; John Wiley & Sons: Chichester, UK, 2011; p. 267. [Google Scholar]
- Singh, R.S.; Saini, G.K.; Kennedy, J.F. Pullulan: Microbial sources, production and applications. Carbohydr. Polym. 2008, 73, 515–531. [Google Scholar] [CrossRef]
- Wang, D.H.; Ni, T.F.; Ju, X.M.; Wei, G.Y. Sodium chloride improves pullulan production by Aureobasidium pullulans but reduces the molecular weight of pullulan. Appl. Microbiol. Biotechnol. 2018, 102, 8921–8930. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Sirohi, R.; Gaurand, V.K.; Pandey, A. Production and applications of pullulan. In Biomass, Biofuels, Biochemicals; Binod, P., Raveendran, S., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 165–221. [Google Scholar] [CrossRef]
- Griffin, D.H. Fungal Physiology, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 1996; p. 480. [Google Scholar] [CrossRef]
- Walker, G.M.; White, N.A. Introduction to fungal physiology. In Fungi Biology and Applications; Wiley: Hoboken, NJ, USA, 2017; pp. 1–35. [Google Scholar]
- Arslan, U. Evaluation of antifungal activity of mono and dipotassium phosphates against phytopathogenic fungi. Fresenius Environ. Bull. 2015, 24, 810–816. Available online: http://www.psp-parlar.de (accessed on 11 March 2022).
- Singh, R.S.; Saini, G.K.; Kennedy, J.F. Downstream processing and characterization of pullulan from a novel colour variant strain of Aureobasidium pullulans FB-1. Carbohydr. Polym. 2009, 78, 89–94. [Google Scholar] [CrossRef]
- Saber-Samandari, S.; Gulcan, H.O.; Saber-Samandari, S.; Gazi, M. Efficient removal of anionic and cationic dyes from an aqueous solution using pullulan-graft-polyacrylamide porous hydrogel. Water Air Soil Pollut. 2014, 225, 2177. [Google Scholar] [CrossRef]
- Cheng, K.C.; Demirci, A.; Catchmark, J.M. Effects of plastic composite support and pH profiles on pullulan production in a biofilm reactor. Appl. Microbiol. Biotechnol. 2010, 86, 853–861. [Google Scholar] [CrossRef]
- Tao, X.; Xie, Y.; Zhang, Q.; Qiu, X.; Yuan, L.; Wen, Y.; Li, M.; Yang, X.; Tao, T.; Xie, M.; et al. Cholesterol-modified amino-pullulan nanoparticles as a drug carrier: Comparative study of cholesterol-modified carboxyethyl pullulan and pullulan nanoparticles. Nanomaterials 2016, 6, 165. [Google Scholar] [CrossRef]
- Trovatti, E.; Fernandes, S.C.M.; Rubatat, L.; Freire, C.S.R.; Silvestre, A.J.D.; Neto, C.P. Sustainable nanocomposite films based on bacterial cellulose and pullulan. Cellulose 2012, 19, 729–737. [Google Scholar] [CrossRef]
- McLntyre, D.D.; Calgary, H.J.V. Structural Studies of Pullulan by Nuclear Magnetic Resonance Spectroscopy. Starch/Stärke 1993, 45, 406–410. [Google Scholar] [CrossRef]
- Sugumaran, K.R.; Gowthami, E.; Swathi, B.; Elakkiya, S.; Srivastava, S.N.; Ravikumar, R.; Gowdhaman, D.; Ponnusami, V. Production of pullulan by Aureobasidium pullulans from Asian palm kernel: A novel substrate. Carbohydr. Polym. 2013, 92, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.M.; Mir, S.A.; Khanday, F.A.; Masoodi, F.A. Advances in pullulan production from agro-based wastes by Aureobasidium pullulans and its applications. Innov. Food Sci. Emerg. Technol. 2021, 74, 102846. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Tested Variable | Pullulan Biosynthesis (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nutritional Component (g/L) | pH | Incubation (h) | Taguchi | Decision Tree | ||||||||||
| Sucrose | K2HPO4 | NaCl | MgSO4 | Yeast Extract | Mean | S/N Ratio | Fitted | Type | Fitted | Residual | Terminal Node | |||
| L1 | 40 | 4 | 0.5 | 0.1 | 1 | 5.5 | 48 | 3.81 ± 0.23 | 11.59 | 3.70 | Test | 2.95 | 1.04 | 1 |
| L2 | 40 | 4 | 0.5 | 0.1 | 2 | 6.5 | 72 | 3.12 ± 0.10 | 9.86 | 3.12 | Test | 2.95 | 0.16 | 1 |
| L3 | 40 | 4 | 0.5 | 0.1 | 3 | 7.5 | 96 | 2.32 ± 0.01 | 7.30 | 2.43 | Training | 2.95 | −0.62 | 1 |
| L4 | 40 | 5 | 1.0 | 0.2 | 1 | 5.5 | 48 | 5.37 ± 0.07 | 14.60 | 5.53 | Test | 5.24 | 0.19 | 4 |
| L5 | 40 | 5 | 1.0 | 0.2 | 2 | 6.5 | 72 | 4.94 ± 0.05 | 13.88 | 4.94 | Training | 5.24 | −0.25 | 4 |
| L6 | 40 | 5 | 1.0 | 0.2 | 3 | 7.5 | 96 | 4.41 ± 0.10 | 12.88 | 4.26 | Test | 4.65 | −0.19 | 5 |
| L7 | 40 | 6 | 1.5 | 0.3 | 1 | 5.5 | 48 | 7.23 ± 0.09 | 17.18 | 7.19 | Training | 6.98 | 0.35 | 7 |
| L8 | 40 | 6 | 1.5 | 0.3 | 2 | 6.5 | 72 | 6.60 ± 0.17 | 16.39 | 6.61 | Test | 6.98 | −0.57 | 7 |
| L9 | 40 | 6 | 1.5 | 0.3 | 3 | 7.5 | 96 | 5.88 ± 0.09 | 15.39 | 5.92 | Training | 5.76 | 0.15 | 9 |
| L10 | 50 | 4 | 1.0 | 0.3 | 1 | 6.5 | 96 | 5.43 ± 0.11 | 14.69 | 5.50 | Training | 5.76 | −0.26 | 9 |
| L11 | 50 | 4 | 1.0 | 0.3 | 2 | 7.5 | 48 | 5.84 ± 0.08 | 15.33 | 5.83 | Training | 6.01 | −0.26 | 8 |
| L12 | 50 | 4 | 1.0 | 0.3 | 3 | 5.5 | 72 | 6.19 ± 0.05 | 15.84 | 6.14 | Test | 6.01 | 0.13 | 8 |
| L13 | 50 | 5 | 1.5 | 0.1 | 1 | 6.5 | 96 | 4.74 ± 0.06 | 13.51 | 4.63 | Test | 4.76 | −0.09 | 2 |
| L14 | 50 | 5 | 1.5 | 0.1 | 2 | 7.5 | 48 | 4.91 ± 0.07 | 13.81 | 4.96 | Training | 5.13 | −0.21 | 3 |
| L15 | 50 | 5 | 1.5 | 0.1 | 3 | 5.5 | 72 | 5.22 ± 0.04 | 14.35 | 5.27 | Training | 5.13 | 0.12 | 3 |
| L16 | 50 | 6 | 0.5 | 0.2 | 1 | 6.5 | 96 | 4.83 ± 0.06 | 13.67 | 4.87 | Training | 4.65 | 0.23 | 5 |
| L17 | 50 | 6 | 0.5 | 0.2 | 2 | 7.5 | 48 | 5.24 ± 0.05 | 14.39 | 5.20 | Training | 5.24 | 0.04 | 4 |
| L18 | 50 | 6 | 0.5 | 0.2 | 3 | 5.5 | 72 | 5.51 ± 0.09 | 14.82 | 5.51 | Training | 5.24 | 0.19 | 4 |
| L19 | 60 | 4 | 1.5 | 0.2 | 1 | 7.5 | 72 | 5.52 ± 0.05 | 14.84 | 5.42 | Training | 5.80 | −0.31 | 6 |
| L20 | 60 | 4 | 1.5 | 0.2 | 2 | 5.5 | 96 | 5.54 ± 0.06 | 14.86 | 5.67 | Test | 5.80 | −0.32 | 6 |
| L21 | 60 | 4 | 1.5 | 0.2 | 3 | 6.5 | 48 | 6.18 ± 0.12 | 15.81 | 6.15 | Training | 5.80 | 0.32 | 6 |
| L22 | 60 | 5 | 0.5 | 0.3 | 1 | 7.5 | 72 | 5.53 ± 0.14 | 14.85 | 5.56 | Test | 6.01 | −0.63 | 8 |
| L23 | 60 | 5 | 0.5 | 0.3 | 2 | 5.5 | 96 | 5.80 ± 0.12 | 15.27 | 5.81 | Test | 5.76 | −0.06 | 9 |
| L24 | 60 | 5 | 0.5 | 0.3 | 3 | 6.5 | 48 | 6.32 ± 0.10 | 16.01 | 6.28 | Test | 6.01 | 0.29 | 8 |
| L25 | 60 | 6 | 1.0 | 0.1 | 1 | 7.5 | 72 | 4.72 ± 0.08 | 13.48 | 4.81 | Training | 4.76 | −0.06 | 2 |
| L26 | 60 | 6 | 1.0 | 0.1 | 2 | 5.5 | 96 | 5.21 ± 0.10 | 14.34 | 5.07 | Test | 5.13 | 0.07 | 3 |
| L27 | 60 | 6 | 1.0 | 0.1 | 3 | 6.5 | 48 | 5.48 ± 0.15 | 14.76 | 5.54 | Training | 5.13 | 0.31 | 3 |
| L1 | 40 | 4 | 0.5 | 0.1 | 1 | 5.5 | 48 | Training | 2.95 | 0.95 | 1 | |||
| L2 | 40 | 4 | 0.5 | 0.1 | 2 | 6.5 | 72 | Training | 2.95 | 0.27 | 1 | |||
| L3 | 40 | 4 | 0.5 | 0.1 | 3 | 7.5 | 96 | Training | 2.95 | −0.64 | 1 | |||
| L4 | 40 | 5 | 1.0 | 0.2 | 1 | 5.5 | 48 | Test | 5.24 | 0.14 | 4 | |||
| L5 | 40 | 5 | 1.0 | 0.2 | 2 | 6.5 | 72 | Test | 5.24 | −0.35 | 4 | |||
| L6 | 40 | 5 | 1.0 | 0.2 | 3 | 7.5 | 96 | Training | 4.65 | −0.18 | 5 | |||
| L7 | 40 | 6 | 1.5 | 0.3 | 1 | 5.5 | 48 | Test | 6.98 | 0.19 | 7 | |||
| L8 | 40 | 6 | 1.5 | 0.3 | 2 | 6.5 | 72 | Training | 6.98 | −0.33 | 7 | |||
| L9 | 40 | 6 | 1.5 | 0.3 | 3 | 7.5 | 96 | Training | 5.76 | 0.02 | 9 | |||
| L10 | 50 | 4 | 1.0 | 0.3 | 1 | 6.5 | 96 | Training | 5.76 | −0.28 | 9 | |||
| L11 | 50 | 4 | 1.0 | 0.3 | 2 | 7.5 | 48 | Test | 6.01 | −0.12 | 8 | |||
| L12 | 50 | 4 | 1.0 | 0.3 | 3 | 5.5 | 72 | Training | 6.01 | 0.19 | 8 | |||
| L13 | 50 | 5 | 1.5 | 0.1 | 1 | 6.5 | 96 | Training | 4.76 | −0.01 | 2 | |||
| L14 | 50 | 5 | 1.5 | 0.1 | 2 | 7.5 | 48 | Training | 5.13 | −0.30 | 3 | |||
| L15 | 50 | 5 | 1.5 | 0.1 | 3 | 5.5 | 72 | Test | 5.13 | 0.11 | 3 | |||
| L16 | 50 | 6 | 0.5 | 0.2 | 1 | 6.5 | 96 | Training | 4.65 | 0.18 | 5 | |||
| L17 | 50 | 6 | 0.5 | 0.2 | 2 | 7.5 | 48 | Training | 5.24 | −0.05 | 4 | |||
| L18 | 50 | 6 | 0.5 | 0.2 | 3 | 5.5 | 72 | Test | 5.24 | 0.26 | 4 | |||
| L19 | 60 | 4 | 1.5 | 0.2 | 1 | 7.5 | 72 | Training | 5.80 | −0.30 | 6 | |||
| L20 | 60 | 4 | 1.5 | 0.2 | 2 | 5.5 | 96 | Training | 5.80 | −0.27 | 6 | |||
| L21 | 60 | 4 | 1.5 | 0.2 | 3 | 6.5 | 48 | Training | 5.80 | 0.30 | 6 | |||
| L22 | 60 | 5 | 0.5 | 0.3 | 1 | 7.5 | 72 | Training | 6.01 | −0.44 | 8 | |||
| L23 | 60 | 5 | 0.5 | 0.3 | 2 | 5.5 | 96 | Training | 5.76 | 0.17 | 9 | |||
| L24 | 60 | 5 | 0.5 | 0.3 | 3 | 6.5 | 48 | Training | 6.01 | 0.22 | 8 | |||
| L25 | 60 | 6 | 1.0 | 0.1 | 1 | 7.5 | 72 | Training | 4.76 | 0.05 | 2 | |||
| L26 | 60 | 6 | 1.0 | 0.1 | 2 | 5.5 | 96 | Training | 5.13 | 0.19 | 3 | |||
| L27 | 60 | 6 | 1.0 | 0.1 | 3 | 6.5 | 48 | Test | 5.13 | 0.51 | 3 | |||
| L1 | 40 | 4 | 0.5 | 0.1 | 1 | 5.5 | 48 | Test | 2.95 | 0.60 | 1 | |||
| L2 | 40 | 4 | 0.5 | 0.1 | 2 | 6.5 | 72 | Training | 2.95 | 0.06 | 1 | |||
| L3 | 40 | 4 | 0.5 | 0.1 | 3 | 7.5 | 96 | Test | 2.95 | −0.64 | 1 | |||
| L4 | 40 | 5 | 1.0 | 0.2 | 1 | 5.5 | 48 | Test | 5.24 | 0.06 | 4 | |||
| L5 | 40 | 5 | 1.0 | 0.2 | 2 | 6.5 | 72 | Training | 5.24 | −0.30 | 4 | |||
| L6 | 40 | 5 | 1.0 | 0.2 | 3 | 7.5 | 96 | Training | 4.65 | −0.35 | 5 | |||
| L7 | 40 | 6 | 1.5 | 0.3 | 1 | 5.5 | 48 | Training | 6.98 | 0.21 | 7 | |||
| L8 | 40 | 6 | 1.5 | 0.3 | 2 | 6.5 | 72 | Training | 6.98 | −0.23 | 7 | |||
| L9 | 40 | 6 | 1.5 | 0.3 | 3 | 7.5 | 96 | Training | 5.76 | 0.19 | 9 | |||
| L10 | 50 | 4 | 1.0 | 0.3 | 1 | 6.5 | 96 | Test | 5.76 | −0.46 | 9 | |||
| L11 | 50 | 4 | 1.0 | 0.3 | 2 | 7.5 | 48 | Training | 6.01 | −0.12 | 8 | |||
| L12 | 50 | 4 | 1.0 | 0.3 | 3 | 5.5 | 72 | Test | 6.01 | 0.23 | 8 | |||
| L13 | 50 | 5 | 1.5 | 0.1 | 1 | 6.5 | 96 | Training | 4.76 | 0.03 | 2 | |||
| L14 | 50 | 5 | 1.5 | 0.1 | 2 | 7.5 | 48 | Training | 5.13 | −0.16 | 3 | |||
| L15 | 50 | 5 | 1.5 | 0.1 | 3 | 5.5 | 72 | Training | 5.13 | 0.04 | 3 | |||
| L16 | 50 | 6 | 0.5 | 0.2 | 1 | 6.5 | 96 | Training | 4.65 | 0.12 | 5 | |||
| L17 | 50 | 6 | 0.5 | 0.2 | 2 | 7.5 | 48 | Training | 5.24 | 0.02 | 4 | |||
| L18 | 50 | 6 | 0.5 | 0.2 | 3 | 5.5 | 72 | Training | 5.24 | 0.36 | 4 | |||
| L19 | 60 | 4 | 1.5 | 0.2 | 1 | 7.5 | 72 | Training | 5.80 | −0.22 | 6 | |||
| L20 | 60 | 4 | 1.5 | 0.2 | 2 | 5.5 | 96 | Test | 5.80 | −0.20 | 6 | |||
| L21 | 60 | 4 | 1.5 | 0.2 | 3 | 6.5 | 48 | Training | 5.80 | 0.51 | 6 | |||
| L22 | 60 | 5 | 0.5 | 0.3 | 1 | 7.5 | 72 | Test | 6.01 | −0.36 | 8 | |||
| L23 | 60 | 5 | 0.5 | 0.3 | 2 | 5.5 | 96 | Training | 5.76 | 0.02 | 9 | |||
| L24 | 60 | 5 | 0.5 | 0.3 | 3 | 6.5 | 48 | Training | 6.01 | 0.41 | 8 | |||
| L25 | 60 | 6 | 1.0 | 0.1 | 1 | 7.5 | 72 | Test | 4.76 | −0.10 | 2 | |||
| L26 | 60 | 6 | 1.0 | 0.1 | 2 | 5.5 | 96 | Training | 5.13 | −0.01 | 3 | |||
| L27 | 60 | 6 | 1.0 | 0.1 | 3 | 6.5 | 48 | Test | 5.13 | 0.22 | 3 | |||
| Source | Freedom Degree | Sum of Square | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Sucrose | 2 | 2.50 | 1.25 | 87.62 | 0.000 |
| K2HPO4 | 2 | 2.53 | 1.27 | 88.94 | 0.000 |
| NaCl | 2 | 4.86 | 2.43 | 170.44 | 0.000 |
| MgSO4 | 2 | 13.04 | 6.52 | 457.60 | 0.000 |
| Yeast extract | 2 | 0.01 | 0.00 | 0.24 | 0.793 |
| pH | 2 | 1.70 | 0.85 | 59.83 | 0.000 |
| Incubation time | 2 | 2.15 | 1.08 | 75.47 | 0.000 |
| Residual Error | 12 | 0.17 | 0.01 | ||
| Total | 26 | 26.96 | |||
| The determination coefficient (R2) | 0.9937 | ||||
| Adjusted-R2 | 0.9863 | ||||
| S/N Ratio Response Analysis | |||||||
|---|---|---|---|---|---|---|---|
| Level | Sucrose | K2HPO4 | NaCl | MgSO4 | Yeast Extract | Initial pH | Incubation Time |
| 1 | 13.23 | 13.35 | 13.09 | 12.56 | 14.27 | 14.76 | 14.83 |
| 2 | 14.49 | 14.35 | 14.42 | 14.42 | 14.24 | 14.29 | 14.26 |
| 3 | 14.92 | 14.94 | 15.13 | 15.66 | 14.13 | 13.59 | 13.55 |
| Delta | 1.69 | 1.59 | 2.04 | 3.11 | 0.14 | 1.17 | 1.29 |
| Rank | 3 | 4 | 2 | 1 | 7 | 6 | 5 |
| MeanResponse Analysis | |||||||
| 1 | 4.854 | 4.883 | 4.720 | 4.391 | 5.243 | 5.543 | 5.597 |
| 2 | 5.324 | 5.249 | 5.289 | 5.282 | 5.245 | 5.292 | 5.263 |
| 3 | 5.589 | 5.634 | 5.757 | 6.093 | 5.278 | 4.931 | 4.906 |
| Delta | 0.736 | 0.750 | 1.037 | 1.702 | 0.035 | 0.612 | 0.691 |
| Rank | 4 | 3 | 2 | 1 | 7 | 6 | 5 |
| Model | Nutritional Component (g/L) | pH | Incubation (h) | Pullulan, % | S/N Ratio | Terminal Node | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sucrose | K2HPO4 | NaCl | MgSO4 | Yeast Extract | Predicted | Actual | |||||
| Taguchi | 60 | 4 | 1.5 | 0.3 | 1 | 5.5 | 48 | 7.17 | 7.23 ± 0.13 | 17.64 | - |
| Decision tree | 40 | 4 | 1.5 | 0.3 | 1 | 5.5 | 48 | 6.98 | 7.11 ± 0.07 | - | 7 |
| Absorption (cm−1) | Appearance | Functional Group | Compound Class | Relative Intensity |
|---|---|---|---|---|
| 3309 | sharp | O-H stretching | Alcohol | 0.250 |
| 2924 | medium | C-H stretching | Alkane | 0.066 |
| 1642 | weak | O-C-O stretching | Aliphatic ether | 0.073 |
| 1417 | medium | C-H bending | Alkane | 0.001 |
| 1360 | medium | C-O-H | Aliphatic ether | 0.097 |
| 1148 | strong | C-O-C stretching | Aliphatic ether | 0.047 |
| 1077 | strong | C-O stretching | Alcohol | 0.047 |
| 995 | strong | C-O stretching | primary alcohol | 0.034 |
| 930 | strong | C-H bending | trisubstituted | 0.034 |
| 755 | strong | C-H bending | disubstituted | 0.012 |
| 706 | strong | C-H bending | disubstituted | 0.012 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saber, W.I.A.; Al-Askar, A.A.; Ghoneem, K.M. Exclusive Biosynthesis of Pullulan Using Taguchi’s Approach and Decision Tree Learning Algorithm by a Novel Endophytic Aureobasidium pullulans Strain. Polymers 2023, 15, 1419. https://doi.org/10.3390/polym15061419
Saber WIA, Al-Askar AA, Ghoneem KM. Exclusive Biosynthesis of Pullulan Using Taguchi’s Approach and Decision Tree Learning Algorithm by a Novel Endophytic Aureobasidium pullulans Strain. Polymers. 2023; 15(6):1419. https://doi.org/10.3390/polym15061419
Chicago/Turabian StyleSaber, WesamEldin I. A., Abdulaziz A. Al-Askar, and Khalid M. Ghoneem. 2023. "Exclusive Biosynthesis of Pullulan Using Taguchi’s Approach and Decision Tree Learning Algorithm by a Novel Endophytic Aureobasidium pullulans Strain" Polymers 15, no. 6: 1419. https://doi.org/10.3390/polym15061419
APA StyleSaber, W. I. A., Al-Askar, A. A., & Ghoneem, K. M. (2023). Exclusive Biosynthesis of Pullulan Using Taguchi’s Approach and Decision Tree Learning Algorithm by a Novel Endophytic Aureobasidium pullulans Strain. Polymers, 15(6), 1419. https://doi.org/10.3390/polym15061419