
The Effect of pH, Metal Ions, and Insoluble Solids on the Production of Fumarate and Malate by Rhizopus delemar in the Presence of CaCO3



Abstract
1. Introduction
2. Results and Discussion
2.1. Biomass
2.2. Effect of CaCO3 Addition
2.3. Effect of Insoluble Solids: Comparison with MgCO3
2.4. Effect of Insoluble Solids: Plaster Sand
2.5. Effect of Calcium Ions
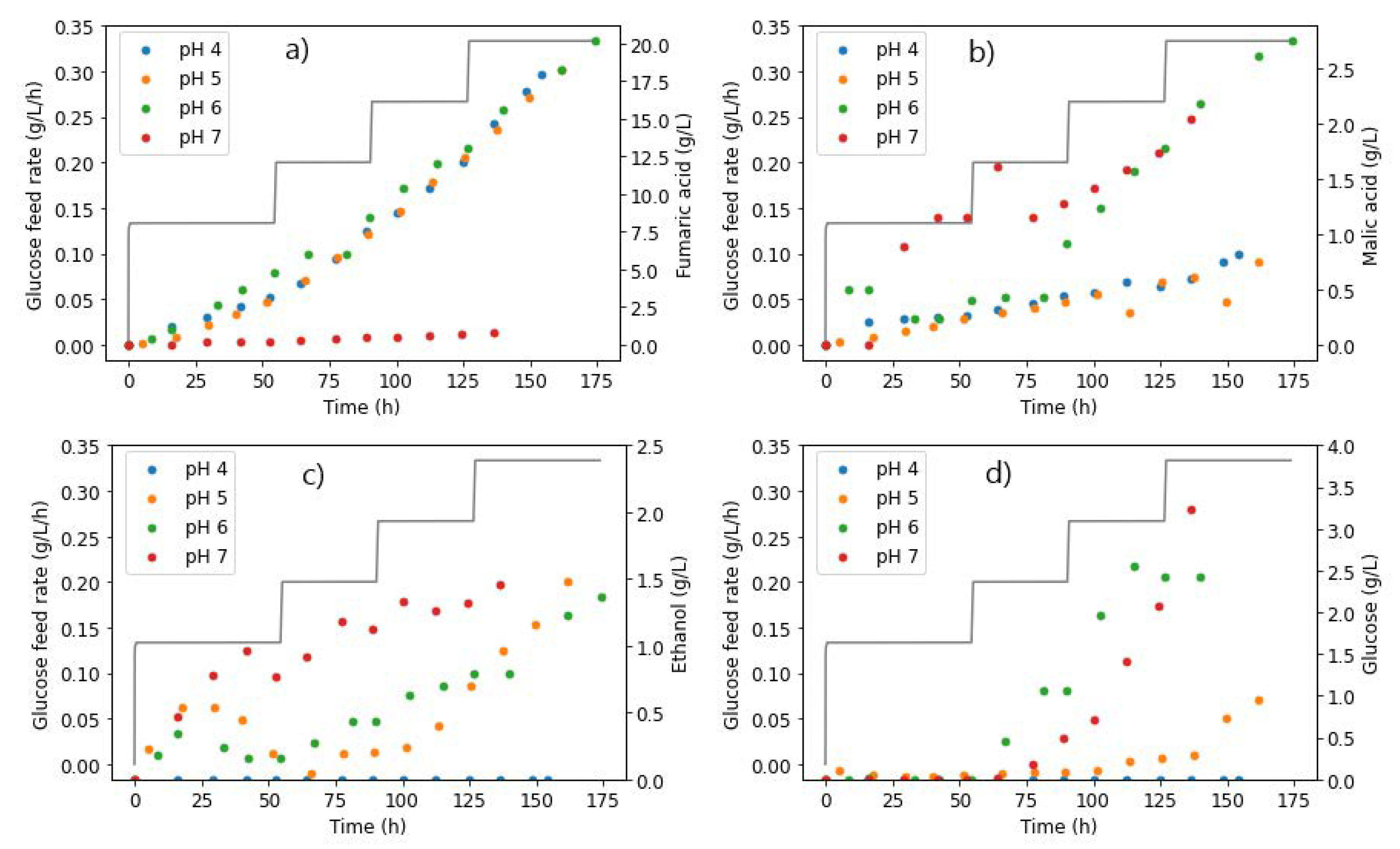
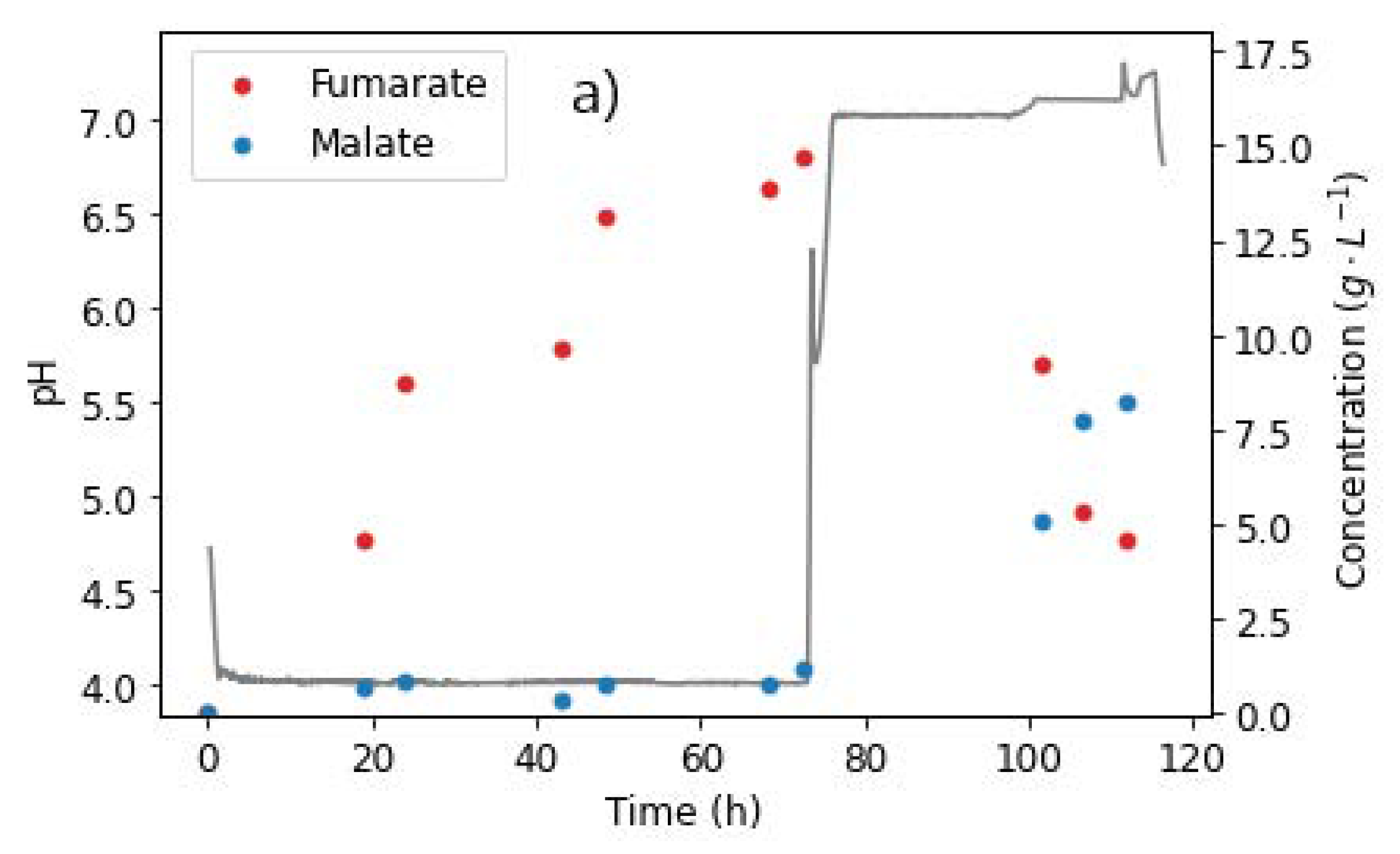
2.6. Effect of pH
2.6.1. Glucose Steps
2.6.2. pH Steps
2.6.3. Hydration of FA to MA
3. Materials and Methods
3.1. Microorganism and Culture Conditions
3.1.1. Inoculum Preparation
3.1.2. Shake Flask Cultivations
Pre-Culture for Biomass Formation
Main Culture for Acid Production
3.1.3. Immobilised Reactor Cultivations
Medium
Reactor Operation and Experimental Design
3.2. Sample Preparation
3.3. Analytical Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| MA | Malic acid |
| FA | Fumaric acid |
| DOE | Department of Energy |
| ICP | Inductively coupled plasma |
| ATCC | American Type Culture Collection |
| OES | Optical emission spectrometry |
| ATP | Adenosine triphosphate |
| RPM | Revolutions per minute |
| Cx | Concentration of components (g L−1) |
| PS | Plaster sand |
| Gibbs free energy (kJ mol−1) | |
| FUM | Fumarase |
| TCA | Tricarboxylic acid |
| MDH | Malate dehydrogenase |
References
- Meussen, B.J.; de Graaff, L.H.; Sanders, J.P.M.; Weusthuis, R.A. Metabolic engineering of Rhizopus oryzae for the production of platform chemicals. Appl. Microbiol. Biotechnol. 2012, 94, 875–886. [Google Scholar] [CrossRef]
- Martin-Dominguez, V.; Bouzas-Santiso, L.; Martinez-Peinado, N.; Santos, V.E.; Ladero, M. Kinetic modelling of the coproduction process of fumaric and malic acids by Rhizopus arrhizus NRRL 1526. Processes 2020, 8, 188. [Google Scholar] [CrossRef]
- Kövilein, A.; Umpfenbach, J.; Ochsenreither, K. Biotechnology for Biofuels Acetate as substrate for L-malic acid production with Aspergillus oryzae DSM 1863. Biotechnol. Biofuels 2021, 48, 14. [Google Scholar] [CrossRef]
- Sebastian, J.; Hegde, K.; Kumar, P.; Rouissi, T.; Brar, S.K. Bioproduction of fumaric acid: An insight into microbial strain improvement strategies. Crit. Rev. Biotechnol. 2019, 39, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass Volume I; National Renewable Energy Laboratory: Golden, CO, USA, 2004; 76p. [Google Scholar] [CrossRef]
- Xu, G.; Chen, X.; Liu, L.; Jiang, L. Fumaric acid production in Saccharomyces cerevisiae by simultaneous use of oxidative and reductive routes. Bioresour. Technol. 2013, 148, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.; Rokem, J.S.; Pines, O. Organic acids: Old metabolites, new themes. J. Chem. Technol. Biotechnol. 2006, 81, 1601–1611. [Google Scholar] [CrossRef]
- Zhou, Z.; Du, G.; Hua, Z.; Zhou, J.; Chen, J. Optimization of fumaric acid production by Rhizopus delemar based on the morphology formation. Bioresour. Technol. 2011, 102, 9345–9349. [Google Scholar] [CrossRef]
- Li, X.; Gu, X.; Lai, C.; Ouyang, J.; Yong, Q. Production of Fumaric Acid by Rhizopus oryzae in Simultaneous Saccharification and Fermentation using Xylo-Oligosaccharides Manufacturing Waste Residue. BioResources 2016, 11, 8831–8843. [Google Scholar] [CrossRef]
- Huang, L.; Wei, P.; Zang, R.; Xu, Z.; Cen, P. High-throughput screening of high-yield colonies of Rhizopus oryzae for enhanced production of fumaric acid. Ann. Microbiol. 2010, 60, 287–292. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Straathof, A.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Oda, Y.; Asano, K.; Sone, T. Rhizopus delemar is the proper name for Rhizopus oryzae fumaric-malic acid producers. Mycologia 2007, 99, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Swart, R.M.; Ronoh, D.K.; Brink, H.; Nicol, W. Continuous Production of Fumaric Acid with Immobilised Rhizopus oryzae: The Role of pH and Urea Addition. Catalysts 2022, 12, 82. [Google Scholar] [CrossRef]
- Cao, N.; Du, J.; Gong, C.S.; Tsao, G.T. Simultaneous Production and Recovery of Fumaric Acid from Immobilized Rhizopus oryzae with a Rotary Biofilm Contactor and an Adsorption Column. Appl. Environ. Microbiol. 1996, 62, 2926–2931. [Google Scholar] [CrossRef] [PubMed]
- Skory, C.D. Lactic acid production by Rhizopus oryzae transformants with modified lactate dehydrogenase activity. Appl. Microbiol. Biotechnol. 2004, 64, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.A.; Lennartsson, P.R.; Edebo, L.; Taherzadeh, M.J. Zygomycetes-based biorefinery: Present status and future prospects. Bioresour. Technol. 2013, 135, 523–532. [Google Scholar] [CrossRef]
- Braaksma, M.; van den Berg, R.A.; van der Werf, M.J.; Punt, P.J. A Top-Down Systems Biology Approach for the Identification of Targets for Fungal Strain and Process Development. In Cellular and Molecular Biology of Filamentous Fungi; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; Chapter 3; pp. 25–35. [Google Scholar] [CrossRef]
- Gangl, I.C.; Weigand, W.A.; Keller, F.A. Economic comparison of calcium fumarate and sodium fumarate production byRhizopus arrhizus. Appl. Biochem. Biotechnol. Part A 1990, 24–25, 663–677. [Google Scholar] [CrossRef]
- Zhou, Y.; Du, J.; Tsao, G.T. Comparison of fumaric acid production by Rhizopus oryzae using different neutralizing agents. Bioprocess Biosyst. Eng. 2002, 25, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Swart, R.M.; Le Roux, F.; Naude, A.; De Jongh, N.W.; Nicol, W. Fumarate production with Rhizopus oryzae: Utilising the Crabtree effect to minimise ethanol by-product formation. Biotechnol. Biofuels 2020, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lv, C.; Xu, Q.; Li, S.; Huang, H.; Ouyang, P. Enhanced acid tolerance of Rhizopus oryzae during fumaric acid production. Bioprocess Biosyst. Eng. 2015, 38, 323–328. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Van Gulik, W.M.; Marang, L.; Van der Wielen, L.A.; Straathof, A.J. Development of a low pH fermentation strategy for fumaric acid production by Rhizopus oryzae. Enzym. Microb. Technol. 2011, 48, 39–47. [Google Scholar] [CrossRef]
- Salek, S.S.; van Turnhout, A.G.; Kleerebezem, R.; van Loosdrecht, M.C. pH control in biological systems using calcium carbonate. Biotechnol. Bioeng. 2015, 112, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Zelle, R.M.; De Hulster, E.; Kloezen, W.; Pronk, J.T.; Van Maris, A.J. Key process conditions for production of C4 dicarboxylic acids in bioreactor Batch cultures of an engineered saccharomyces cerevisiae strain. Appl. Environ. Microbiol. 2010, 76, 744–750. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chi, Z.; Wang, Z.P.; Wang, G.Y.; Khan, I.; Chi, Z.M. Microbial biosynthesis and secretion of L-malic acid and its applications. Crit. Rev. Biotechnol. 2016, 8551, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Geyer, M.; Onyancha, F.M.; Nicol, W.; Brink, H.G. Malic Acid Production by Aspergillus Oryzae: The Role of CaCO3. Chem. Eng. Trans. 2018, 70, 1801. [Google Scholar] [CrossRef]
- Karaffa, L.; Fekete, E.; Kubicek, C.P. The role of metal ions in fungal organic acid accumulation. Microorganisms 2021, 9, 1267. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, N. Organic acid production by immobilized filamentous fungi. Top. Catal. 1991, 5, 188–190. [Google Scholar] [CrossRef]
- Song, P.; Li, S.; Ding, Y.; Xu, Q.; Huang, H. Expression and characterization of fumarase (FUMR) from Rhizopus oryzae. Fungal Biol. 2011, 115, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y. Fumaric acid Fermentation by Rhizopus oryzae in Submerged Systems. Ph.D. Thesis, Purdue University, Ann Arbor, MI, USA, 1999. [Google Scholar]
- Zhang, K.; Yu, C.; Yang, S.T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Fumaric acid fermentation with immobilised Rhizopus oryzae: Quantifying time-dependent variations in catabolic flux. Process Biochem. 2017, 56, 8–20. [Google Scholar] [CrossRef]
- Taymaz-Nikerel, H.; Jamalzadeh, E.; Borujeni, A.E.; Verheijen, P.J.; Van Gulik, W.M.; Heijnen, S.J. A thermodynamic analysis of dicarboxylic acid production in microorganisms. Biothermodynamics 2013, 2013, 547–579. [Google Scholar]
- Goldberg, R.N.; Gajewski, E.; Steckler, D.K.; Tewari, Y.B. Thermodynamics of the conversion of aqueous L-Aspartic acid to fumaric acid and ammonia. Biophys. Chem. 1986, 24, 13–23. [Google Scholar] [CrossRef]
- Friedberg, D.; Peleg, Y.; Monsonego, A.; Maissi, S.; Battat, E.; Rokem, J.S.; Goldberg, I. The fumR gene encoding fumarase in the filamentous fungus Rhizopus oryzae: Cloning, structure and expression. Gene 1995, 163, 139–144. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, S.T.; Chalmers, J.J.; Wood, D. Fumaric Acid Fermentation by Rhizopus oryzae with Integrated Separation Technologies; The Ohio State University: Columbus, OH, USA, 2012; pp. 1–210. [Google Scholar]
- Naude, A.; Nicol, W. Malic acid production through the whole-cell hydration of fumaric acid with immobilised Rhizopus oryzae. Biochem. Eng. J. 2018, 137, 152–161. [Google Scholar] [CrossRef]
- Gu, C.; Zhou, Y.; Liu, L.; Tan, T.; Deng, L. Production of fumaric acid by immobilized Rhizopus arrhizus on net. Bioresour. Technol. 2013, 131, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.Q.; Li, S.; Chen, Y.; Xu, Q.; Huang, H.; Sheng, X.Y. Enhancement of fumaric acid production by Rhizopus oryzae using a two-stage dissolved oxygen control strategy. Appl. Biochem. Biotechnol. 2010, 162, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, S.; Huang, H.; Wen, J. Key technologies for the industrial production of fumaric acid by fermentation. Biotechnol. Adv. 2012, 30, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hu, H.; Jin, Y.; Yue, X.; Deng, L.; Wang, F.; Tan, T. Co-fermentation of a mixture of glucose and xylose to fumaric acid by Rhizopus arrhizus RH 7-13-9#. Bioresour. Technol. 2017, 233, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Du, J.; Tsao, G.T. Mycelial pellet formation by Rhizopus oryzae ATCC 20344. Appl. Biochem. Biotechnol.—Part A Enzym. Eng. Biotechnol. 2000, 84–86, 779–789. [Google Scholar] [CrossRef]
- Jongh, N.W.D.; Swart, R.M.; Nicol, W.; Engineering, B. Fed-batch growth of Rhizopus oryzae: Eliminating ethanol formation by controlling glucose addition. Biochem. Eng. J. 2021, 169, 107961. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Cx (g L−1) | Ca2+ (g L−1) | Mg2+ (g L−1) | pH |
|---|---|---|---|---|
| CaCO3 | 0 | 4.79 ± 0.07 | 0.29 ± 0.01 | 4.17 ± 0.21 |
| 60 | 5.63 ± 0.11 | 0.28 ± 0.01 | 7.45 ± 0.07 | |
| 100 | 5.69 ± 0.71 | 0.29 ± 0.02 | 7.37 ± 0.06 | |
| MgCO3 | 0 | 0.74 ± 0.02 | 3.21 ± 0.05 | 4.6 |
| 60 | 0.78 ± 0.02 | 6.54 ± 0.23 | 8.0 | |
| 100 | 0.78 ± 0.01 | 8.05 ± 0.16 | 8.1 | |
| Plaster sand | 40 | 4.70 ± 0.07 | 0.31 ± 0.03 | 3.73 ± 0.06 |
| 80 | 4.69 ± 0.71 | 0.29 ± 0.02 | 3.63 ± 0.06 | |
| 120 | 4.51 ± 0.60 | 0.29 ± 0.01 | 3.87 ± 0.12 |
| Fumaric Acid | Malic Acid | |||
|---|---|---|---|---|
| Process | Titre (g L−1) | Productivity (g L−1 h−1) | Titre (g L−1) | Productivity (g L−1 h−1) |
| Immobilised reactor * | 65.89 | 0.138 | 8.20 | 0.205 |
| Immobilised reactor (Swart et al. [13]) | - | 0.305 | - | - |
| Immobilised reactor (Gu et al. [38]) | 32.03 | 1.33 | - | - |
| Immobilised reactor (Naude et al. [32]) | 40.13 | 0.32 | 2.6 | 0.022 |
| Stirred tank (YQ Fu et al. [39]) | 56.2 | 0.7 | - | - |
| Stirred tank (Xu et al. [40]) | 41.1 | 0.37 | - | - |
| Shake flask † | 34.84 | 0.176 | 33.29 | 0.168 |
| Shake flask (Dominguez et al. [2]) | 30 | - | 25 | - |
| Shake flask (Liu et al. [41]) | 46.7 | - | - | - |
| Shake flask (K. Zhang et al. [31]) | 50.2 | 0.34 | 2.1 | 0.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronoh, D.K.; Swart, R.M.; Nicol, W.; Brink, H. The Effect of pH, Metal Ions, and Insoluble Solids on the Production of Fumarate and Malate by Rhizopus delemar in the Presence of CaCO3. Catalysts 2022, 12, 263. https://doi.org/10.3390/catal12030263
Ronoh DK, Swart RM, Nicol W, Brink H. The Effect of pH, Metal Ions, and Insoluble Solids on the Production of Fumarate and Malate by Rhizopus delemar in the Presence of CaCO3. Catalysts. 2022; 12(3):263. https://doi.org/10.3390/catal12030263
Chicago/Turabian StyleRonoh, Dominic Kibet, Reuben Marc Swart, Willie Nicol, and Hendrik Brink. 2022. "The Effect of pH, Metal Ions, and Insoluble Solids on the Production of Fumarate and Malate by Rhizopus delemar in the Presence of CaCO3" Catalysts 12, no. 3: 263. https://doi.org/10.3390/catal12030263
APA StyleRonoh, D. K., Swart, R. M., Nicol, W., & Brink, H. (2022). The Effect of pH, Metal Ions, and Insoluble Solids on the Production of Fumarate and Malate by Rhizopus delemar in the Presence of CaCO3. Catalysts, 12(3), 263. https://doi.org/10.3390/catal12030263