
A Bibliometric Analysis and Review of Pullulan-Degrading Enzymes—Past and Current Trends

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
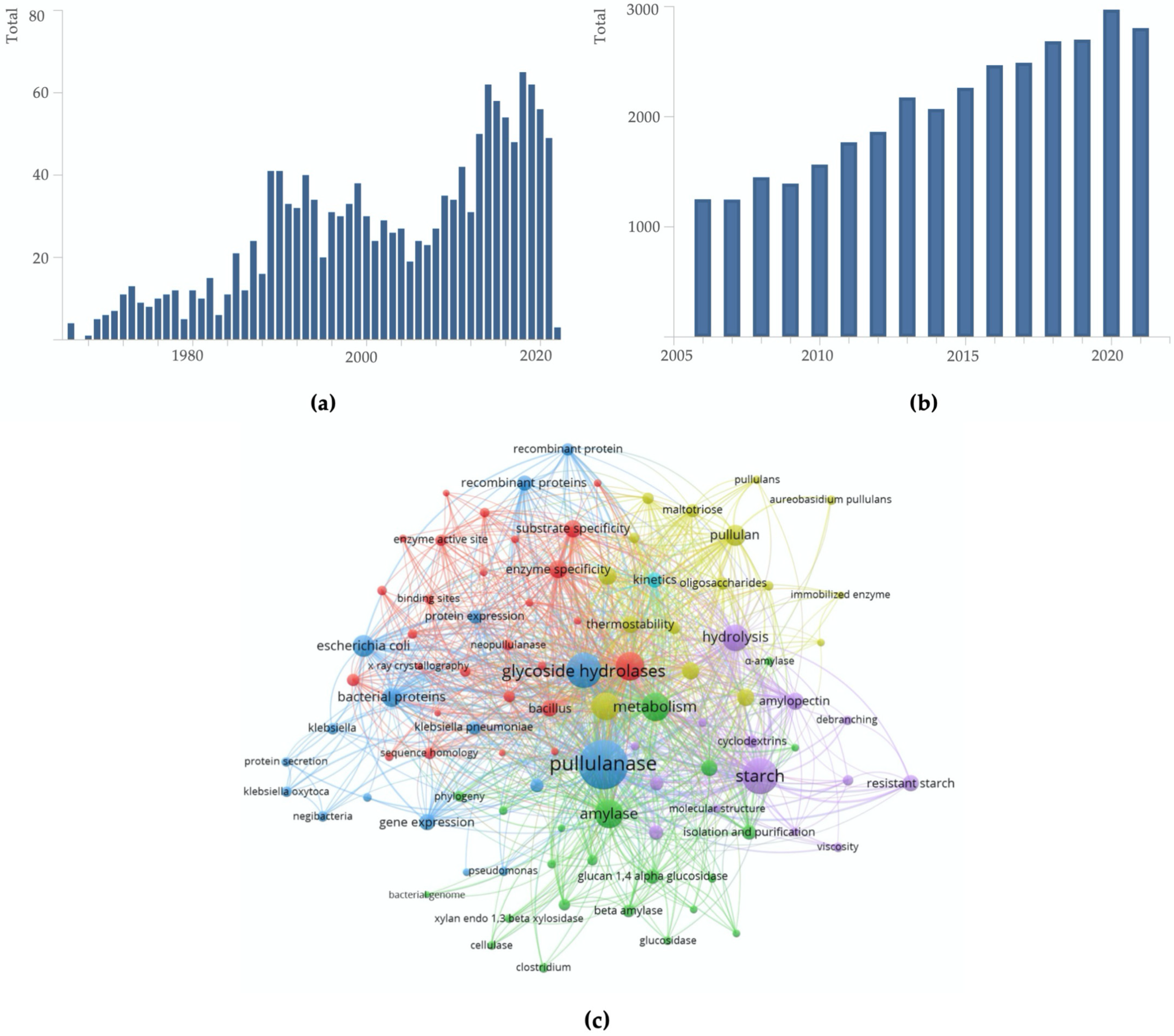
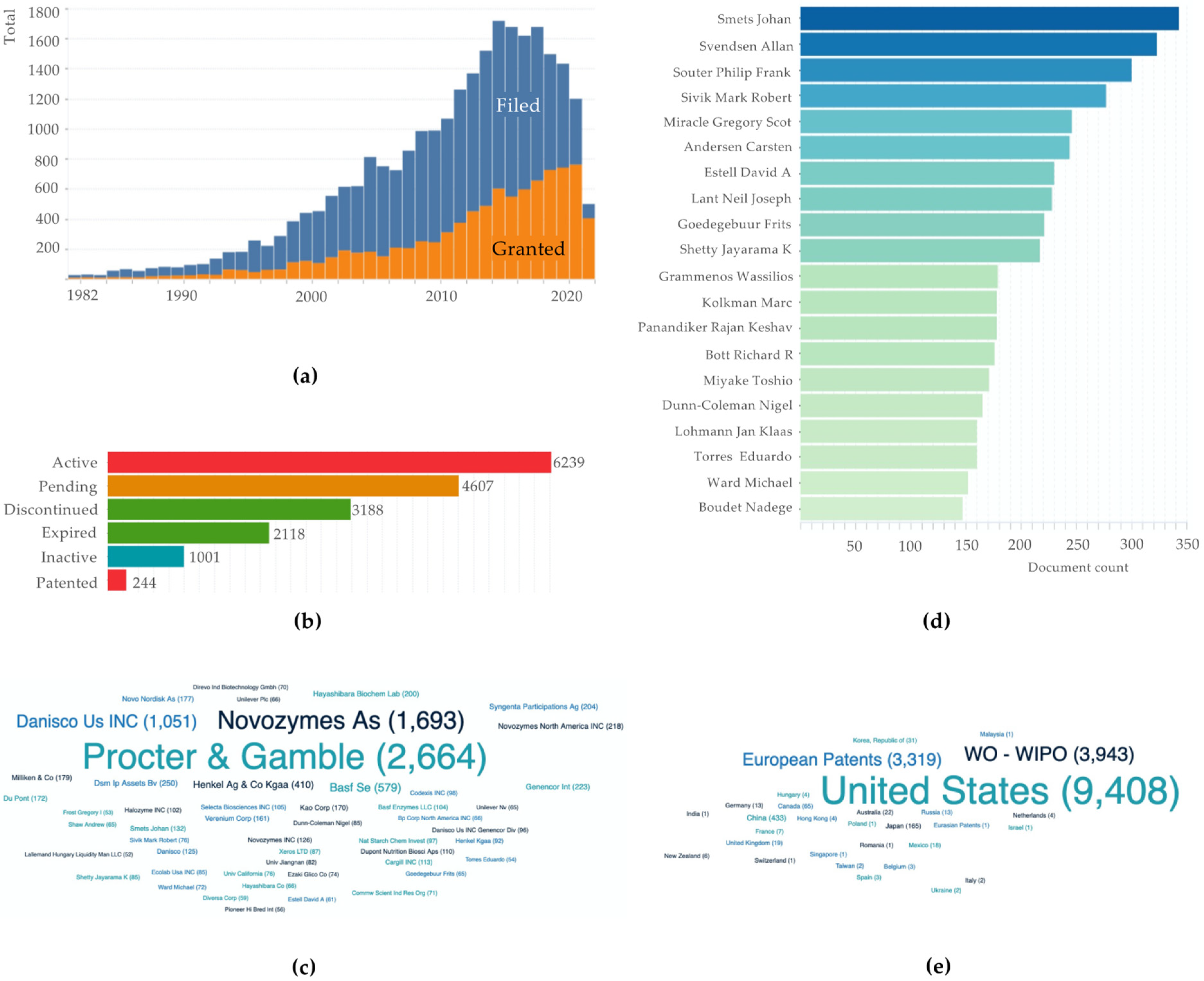
2. Bibliometric Analysis Related to the Publications and Patents of Pullulan-Degrading Enzymes
2.1. Publication Bibliometric Analysis
2.2. Patents Bibliometric Analysis
3. Scientific Review of Pullulan-Degrading Enzymes
3.1. Classification and Action of Pullulan-Degrading Enzymes
3.2. Domains, Structures and Properties of Pullulan-Degrading Enzymes
3.3. Single Immobilisation of Pullulan-Degrading Enzyme
3.4. Co-Immobilisation of Pullulan-Degrading Enzymes
- (i)
- free enzymes mixture (PulASK+TASKA)
- (ii)
- individual immobilised enzymes mixture (HFA403/M–PulASK+ HFA403/M–TASKA)
- (iii)
- co-immobilised enzymes (PulASK–HFA403/M–TASKA).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TE a | Source | Free Enzyme/Support | Topt (°C) | pHopt | Activity Half-Life | Reusability | Reaction Product b | Improvement after Immobilisation | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|
| Pullulan | Starch | |||||||||
| Binding to a carrier approach using covalent attachment | ||||||||||
| I c | Klebsiella pneumoniae | Free enzyme | 30 | 5.0 | 5 min at 50 °C | - | RD | ND | Thermostability | [80] |
| Amylose-aldehyde conjugate | 50 | 5.0 | >30 min at 50 °C | ND | RD | ND | ||||
| I | Bacillus acidopullulyticus | Free enzyme | 60 | 5.0 | 30 min at 60 °C | - | RD | ND | Thermostability | [91] |
| Chitosan-amino beads | 55 | 3.5 | 30 min at 55 °C | ND | RD | ND | ||||
| I | Klebsiella pneumoniae | Free enzyme | 25 | 5.0 | 5 min at 50 °C | - | RD | ND | Thermostability and reusability | [92] |
| Polyacrylate -epoxide beads | 35 | 5.0 | 30 min at 50 °C | 8 cycles at 30 °C | RD | ND | ||||
| I | Klebsiella pneumoniae | Free enzyme | 50 | 6.0 | 30 min at 45 °C | - | RD | ND | Thermostability | [93] |
| Agar-epoxide gel | 50 | 6.0 | 4 h at 45 °C | ND | RD | ND | ||||
| I | Klebsiella pneumoniae | Free enzyme | 50 | 5.5 | 2 h at 70 °C | - | RD | ND | Thermostability and reusability | [94] |
| Magnetic chitosan-amino nanoparticles | 50 | 5.0 | 6 h at 70 °C | 10 cycles at 70 °C | RD | ND | ||||
| I c | Bacillus acidopullulyticus | Free enzyme | 50 | 5.0 | >6 h at 60 °C | - | RD | RD | Reusability | [95] |
| Duolite XAD761 beads | 60 | 5.5 | >6 h at 60 °C | 25 cycles at 60 °C | RD | RD | ||||
| I | Bacilius subtilis | Free enzyme | 50 | 4.4 | 5 h at 60 °C | - | RD | ND | Thermostability and reusability | [96] |
| Magnetic chitosan-amino nanoparticles | 50 | 3.5 | >5 h at 60 °C | 8 cycles at 50 °C | RD | ND | ||||
| I | Klebsiella pneumonia | Free enzyme | 60 | 5.5 | 1 h at 60 °C | - | RD | ND | Thermostability | [81] |
| Calcium alginate-carboxyl | 60 | 5.5 | 50 h at 60 °C | ND | RD | ND | ||||
| I | Klebsiella pneumoniae | Free enzyme | 60 | 4.4 | ND | - | RD | ND | Reusability | [97] |
| Magnetic chitosan-amino nanoparticles | 60 | 4.4 | ND | 10 cycles at 50 °C | RD | ND | ||||
| I | Fontibacillus sp. DSHK 107 | Free enzyme | 35 | 5.0 | 5 h at 35 °C | - | RD | ND | Thermostability and reusability | [98] |
| Silica-amino nanoparticles | 35 | 6.0 | 24 h at 35 °C | 10 cycles at 30 °C | RD | ND | ||||
| I c | Anoxybacillus sp. SK3-4 | Free enzyme | 60 | 6.0 | >4 h at 60 °C | - | G3 | - | Reusability and product specificity on starch | [86] |
| ReliZyme HFA403/M | 60 | 6.0 | 3 h at 60 °C | 12 cycles at 60 °C | G3 | G3 | ||||
| II | Anoxybacillus sp. WB42 | Free enzyme | 65 | 6.0 | 30 min at 70 °C | - | RD | ND | Reusability | [78] |
| Magnetic Ni-IDA nanoparticles | 65 | 5.5 | 30 min at 70 °C | 12 cycles at 60 °C | RD | ND | ||||
| PHI c | Geobacillus stearothermophilus TRS40 | Free enzyme | 60 | 6.0 | >1 h at 60 °C | - | G1, G2, P | ND | Thermostability | [79] |
| Zetaffinity™ 10 beads | 60 | 6.0 | 30 h at 60 °C | ND | G1, G2, P | ND | ||||
| Binding to a carrier approach using ionic bonding | ||||||||||
| I | Streptomyces flavochromogenes | Free enzyme | 55 | 5.5 | 30 min at 55 °C | - | RD | ND | Thermostability | [99] |
| TEAE-cellulose-amino beads | 55 | 6.0 | 30 min at 55 °C | ND | RD | ND | ||||
| II | Anoxybacillus sp. WB42 | Free enzyme | 65 | 6.0 | 30 min at 70 °C | - | RD | ND | Reusability | [82] |
| Magnetic carboxymethyl cellulose (CMC)-amino nanoparticles | 65 | 6.0 | 30 min at 63 °C | 27 cycles at 60 °C | RD | ND | ||||
| Binding to a carrier approach using physical adsorption | ||||||||||
| I | Paenibacillus polymyxa Nws-pp2 | Free enzyme | ND | ND | ND | ND | ND | ND | One-step purification and immobilization | [100] |
| Magnetic nanoparticles | ND | ND | ND | ND | ND | ND | ||||
| Entrapment (encapsulation) approach | ||||||||||
| I | Klebsiella aerogenes | Free enzyme | 25 | ND | ND | - | G3 | ND | None | [101] |
| Polyacrylamide gel | 25 | ND | ND | ND | G3 | ND | ||||
| I | Bacillus acidopullulyticus | Free enzyme | 55 | 5.0 | 1 h at 55 °C | - | RD | RD | Thermostability | [102] |
| Calcium alginate beads | 55 | 5.0 | 2 h at 55 °C | ND | RD | RD | ||||
| I | Klebsiella pneumoniae | Free enzyme | 50 | 4.4 | 1 h at 60 °C | - | RD | ND | Thermostability and reusability | [103] |
| Chitosan-encapsulated magnetic κ-carrageenan nanoparticles | 50 | 4.4 | 3 h at 60 °C | 10 cycles at 50 °C | RD | ND | ||||
| Entrapment (encapsulation) approach | ||||||||||
| I | Klebsiella pneumoniae | Free enzyme | 50 | 4.4 | 20 min at 62 °C | - | ND | ND | Thermostability and reusability | [104] |
| Chitosan (50 kDa)-encapsulated magnetic κ-carrageenan nanoparticles | 50 | 4.4 | 3 h at 62 °C | 6 cycles at 62 °C | ND | ND | ||||
| I | Klebsiella pneumoniae | Free enzyme | 50 | 4.4 | 1 h at 60 °C | - | RD | ND | Thermostability and reusability | [83] |
| Sol-gel-encapsulated magnetic chitosan nanoparticles | 50 | 4.4 | 5 h at 60 °C | 6 cycles at 50 °C | RD | ND | ||||
| I | Geobacillus kaustophilus DSM 7263T | Free enzyme | 65 | 6.0 | 45 h at 65 °C | - | G3 | ND | Thermostability and reusability | [105] |
| Calcium alginate beads | 65 | 6.0 | >70 h at 65 °C | 4 cycles at 65 °C | RD | ND | ||||
| I | Klebsiella pneumoniae | Free enzyme | 50 | 4.4 | ND | - | RD | RD | Thermostability, reusability, and product specificity on resistant starch | [106] |
| Sol-gel-encapsulated magnetic chitosan nanoparticles | 50 | 4.4 | ND | 8 cycles at 50 °C | RD | RD | ||||
| I | Bacillus subtilis | Free enzyme | 50 | 4.4 | 3 h at 50 °C | - | ND | ND | Thermostability and reusability | [107] |
| Streptavidin-encapsulated magnetic nanoparticles | 60 | 4.4 | >6 h at 65 °C | 8 cycles at 60 °C | ND | ND | ||||
| TE a | Source | Support | Topt (°C) | pHopt | Activity Half-Life | Reusability | Starch Degradation Process | Ref. | |
|---|---|---|---|---|---|---|---|---|---|
| Time | Product b | ||||||||
| Binding to a carrier approach using covalent attachment | |||||||||
| Type I pullulanase Glucoamylase | Klebsiella aerogenes Aspergillus niger | Free enzymes mixture | 40 | 5.0 | 2 days at 50 °C | - | > 90 h | G2, G3, G4 | [108] |
| Bovine collagen-azide membrane | 60 | 7.0 | 7 days at 50 °C | ND | 80 h | G2, G3, G4 | |||
| Type I pullulanase Glucoamylase | Bacillus acidopullulyticus Aspergillus niger | Free enzymes mixture | 23 | 6.0 | ND | - | 24 h | RD | [109] |
| Hypol® 2002 foam | 23 | 6.0 | ND | ND | 4 h | RD | |||
| Type I pullulanase β-amylase | Klebsiella pneumoniae Barley | Free enzymes mixture | 30 | 4.5 | 10 min at 50 °C | - | 24 h | G2 | [110] |
| Polyacrylate-epoxide beads | 30 | 4.5 | 60 min at 60 °C | 30 cycles at 30 °C | 24 h | G2 | |||
| Type I pullulanase α-amylase Glucoamylase | ND Bacillus licheniforms Aspergillus niger | Free enzymes mixture | 70 | 6.0 | 2 h at 90 °C | - | 150 min | RD | [111] |
| Magnetic-amino nanoparticles | 80 | 7.0 | 4 h at 90 °C | 8 cycles at 70 °C | 90 min | RD | |||
| Type I pullulanase c α-amylase c | Anoxybacillus sp. SK3-4 Anoxybacillus sp. SK3-4 | Free enzymes mixture | 60 | 6.0 | ND | ND | 48 h | G1, G2, G4 | [86] |
| ReliZyme HFA403/M | 60 | 6.0 | ND | ND | 48 h | G1, G2, G3, G4, G5 | |||
| Binding to a carrier approach using ionic bonding | |||||||||
| Type I pullulanase β-amylase | Klebsiella pneumoniae Soybean | Free enzymes mixture | 60 | 6.0 | 6 days at 60 °C | - | 20 days | G2 | [112] |
| Chitopearl® BCW 3505 beads | 65 | 7.0 | 6 days at 60 °C | ND | 20 days | G2 | |||
| Type I pullulanase Maltotetraose- forming amylase | Klebsiella pneumoniae Pseudomonas stutzeri | Free enzymes mixture | 50 | 6.0 | 8 days at 55 °C | - | 25 days | G4 | [113] |
| Chitopearl® BCW 3505 beads | 50 | 6.0 | 8 days at 55 °C | ND | 25 days | G4 | |||
| Type I pullulanase Glucoamylase Xylose isomerase | Bacillus acidopullulyticus Aspergillus niger Streptomyces rubiginosus | Free enzymes mixture | 55 | 7.5 | ND | - | 5 h | F, G1 | [114] |
| BIOBONE™ beads | 55 | 7.0 | ND | ND | 5 h | F, G1 | |||
| Type I pullulanase β-amylase | Bacillus brevis Sweet potato | Free enzymes mixture | 50 | 6.0 | 30 min at 50 °C | - | 14 days | G2, G3 | [115] |
| Chitopearl® BCW 3505 beads | 70 | 6.0 | 30 min at 60 °C | ND | 14 days | G2, G3 | |||
| Cross-linking enzyme aggregates (CLEAs) approach | |||||||||
| Type I pullulanase Glucoamylase | ND ND | Free enzymes mixture | 60 | 5.0 | 1 h at 65 °C | - | 4 h | RD | [116] |
| CLEAs using glutaraldehyde | 70 | 7.0 | >4 h at 65 °C | 8 cycles at 70 °C | 3 h | RD | |||
| Type I pullulanase α-amylase Glucoamylase | ND ND ND | Free enzymes mixture | 65 | 6.0 | 1 h at 75 °C | - | 35 h | RD | [116] |
| CLEAs using glutaraldehyde | 75 | 7.0 | >3 h at 75 °C | 6 cycles at 75 °C | 2 h | RD | |||
3.5. Protein Engineering for Improving the Performance of Pullulan-Degrading Enzymes
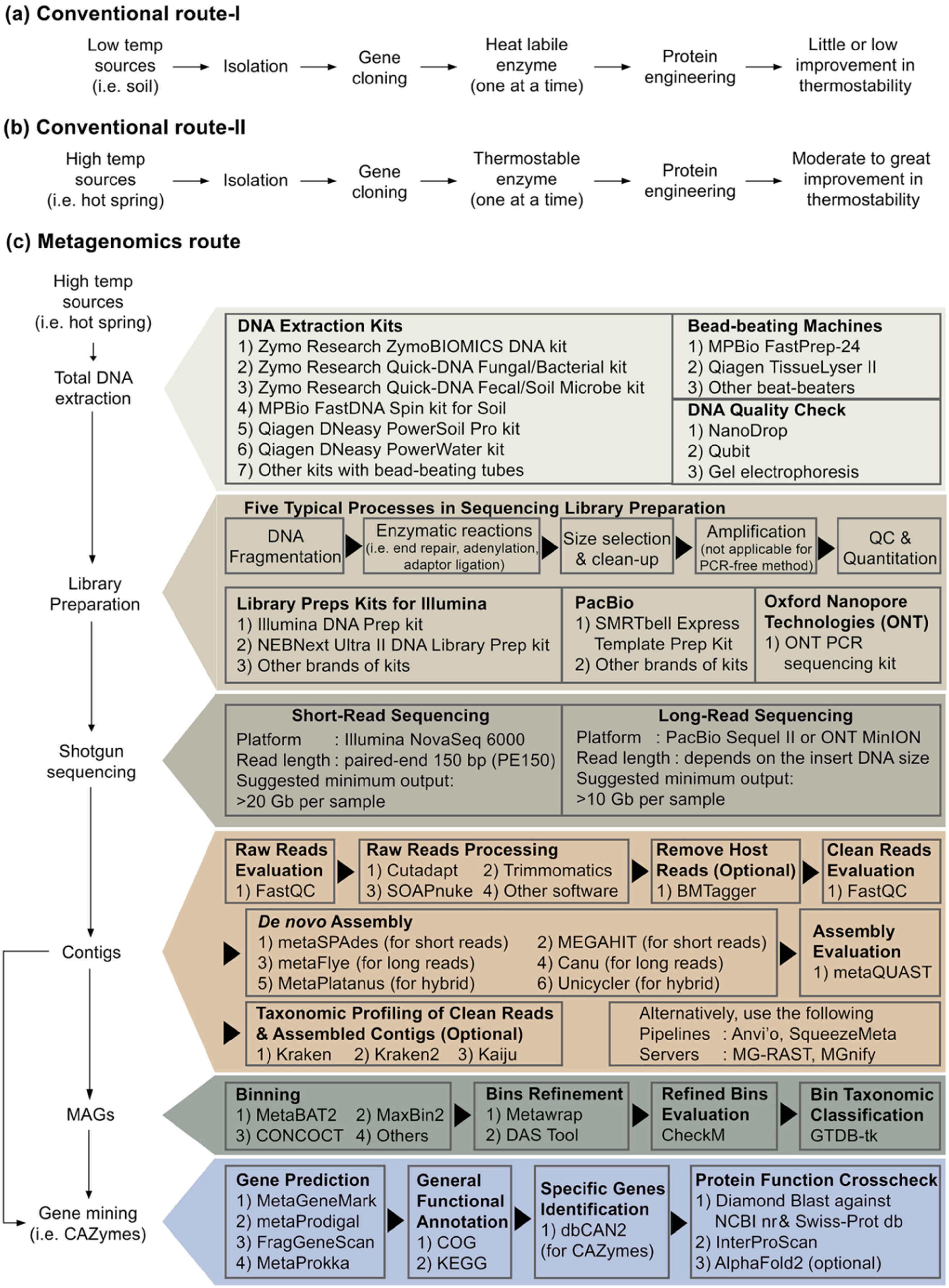
3.6. Influence of Next-Generation Metagenome Sequencing on Novel Pullulan-Degrading Enzymes Discovery
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singh, R.S.; Kaur, N.; Rana, V.; Kennedy, J.F. Pullulan: A novel molecule for biomedical applications. Carbohydr. Polym. 2017, 171, 102–121. [Google Scholar] [CrossRef]
- Singh, R.S.; Saini, G.K.; Kennedy, J.F. Pullulan: Microbial sources, production and applications. Carbohydr. Polym. 2008, 73, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chi, Z.; Wang, G.Y.; Wang, Z.P.; Liu, G.L.; Lee, C.F.; Ma, Z.C.; Chi, Z.M. Taxonomy of Aureobasidium spp. and biosynthesis and regulation of their extracellular polymers. Crit. Rev. Microbiol. 2015, 41, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Sugumaran, K.R.; Ponnusami, V. Review on production, downstream processing and characterization of microbial pullulan. Carbohydr. Polym. 2017, 173, 573–591. [Google Scholar]
- Prajapati, V.D.; Jani, G.K.; Khanda, S.M. Pullulan: An exopolysaccharide and its various applications. Carbohydr. Polym. 2013, 95, 540–549. [Google Scholar] [CrossRef]
- Wu, S.J.; Chen, J. Preparation of maltotriose from fermentation broth by hydrolysis of pullulan using pullulanase. Carbohydr. Polym. 2014, 107, 94–97. [Google Scholar] [CrossRef]
- Klein, M.N.; Kupper, K.C. Biofilm production by Aureobasidium pullulans improves biocontrol against sour rot in citrus. Food Microbiol. 2018, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, T.; Shibuya, T.; Shiobara, S. Safety studies of a novel starch, pullulan: Chronic toxicity in rats and bacterial mutagenicity. Food Chem. Toxicol. 1997, 35, 323–329. [Google Scholar] [CrossRef]
- Cheng, K.C.; Demirci, A.; Catchmark, J.M. Pullulan: Biosynthesis, production, and applications. Appl. Microbiol. Biotechnol. 2011, 92, 29–44. [Google Scholar] [CrossRef]
- Singh, R.S.; Kaur, N.; Rana, V.; Kennedy, J.F. Recent insights on applications of pullulan in tissue engineering. Carbohydr. Polym. 2016, 153, 455–462. [Google Scholar] [CrossRef]
- Nisha, M.; Satyanarayana, T. Characteristics, protein engineering and applications of microbial thermostable pullulanases and pullulan hydrolases. Appl. Microbiol. Biotechnol. 2016, 100, 5661–5679. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.K.; Singhania, R.R.; Pandey, A. Novel enzymatic processes applied to the food industry. Curr. Opin. Food Sci. 2016, 7, 64–72. [Google Scholar] [CrossRef]
- Buchholz, K.; Seibel, J. Industrial carbohydrate biotransformations. Carbohydr. Res. 2008, 343, 1966–1979. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Saini, G.K.; Kennedy, J.F. Maltotriose syrup preparation from pullulan using pullulanase. Carbohydr. Polym. 2010, 80, 401–407. [Google Scholar] [CrossRef]
- Mishra, B.; Manikanta, A.; Zamare, D. Preparation of maltotriose syrup from microbial pullulan by using pullulanase enzyme. Biosci. Biotechnol. Res. Asia 2016, 13, 481–485. [Google Scholar] [CrossRef]
- Bertoldo, C.; Antranikian, G. Starch-hydrolyzing enzymes from thermophilic archaea and bacteria. Curr. Opin. Chem. Biol. 2002, 6, 151–160. [Google Scholar] [CrossRef]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: A biotechnological perspective. Process Biochem. 2003, 38, 1599–1616. [Google Scholar] [CrossRef]
- Ara, K.; Saeki, K.; Igarashi, K.; Takaiwa, M.; Uemura, T.; Hagihara, H.; Kawai, S.; Ito, S. Purification and characterization of an alkaline amylopullulanase with both α-1,4 and α-1,6 hydrolytic activity from alkalophilic Bacillus sp. KSM-1378. BBA-Gen. Subj. 1995, 1243, 315–324. [Google Scholar] [CrossRef]
- Rajaei, S.; Noghabi, K.A.; Sadeghizadeh, M.; Zahiri, H.S. Characterization of a pH and detergent-tolerant, cold-adapted type I pullulanase from Exiguobacterium sp. SH3. Extremophiles 2015, 19, 1145–1155. [Google Scholar] [CrossRef]
- Rajdevi, K.P.; Yogeeswaran, G. Cooperativity and substrate specificity of an alkaline amylase and neopullulanase complex of Micrococcus halobius OR-1. Appl. Biochem. Biotechnol.-Part A Enzym. Eng. Biotechnol. 2001, 90, 233–250. [Google Scholar] [CrossRef]
- Domań-Pytka, M.; Bardowski, J. Pullulan degrading enzymes of bacterial origin. Crit. Rev. Microbiol. 2004, 30, 107–121. [Google Scholar] [CrossRef]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Hii, S.L.; Tan, J.S.; Ling, T.C.; Ariff, A. Bin Pullulanase: Role in starch hydrolysis and potential industrial applications. Enzym. Res. 2012, 2012, 921362. [Google Scholar]
- Xu, P.; Zhang, S.Y.; Luo, Z.G.; Zong, M.H.; Li, X.X.; Lou, W.Y. Biotechnology and bioengineering of pullulanase: State of the art and perspectives. World J. Microbiol. Biotechnol. 2021, 37, 1–10. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, S.; Lv, M. Structural and functional analysis of GH57 family thermostable amylopullulanase—A review. Wei Sheng Wu Xue Bao 2011, 51, 21–28. [Google Scholar] [PubMed]
- Xia, W.; Zhang, K.; Su, L.; Wu, J. Microbial starch debranching enzymes: Developments and applications. Biotechnol. Adv. 2021, 50, 107786. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Nie, Y.; Xu, Y. Industrially produced pullulanases with thermostability: Discovery, engineering, and heterologous expression. Bioresour. Technol. 2019, 278, 360–371. [Google Scholar] [CrossRef]
- Guo, J.; Coker, A.R.; Wood, S.P.; Cooper, J.B.; Keegan, R.M.; Ahmad, N.; Muhamma, M.A.; Rashid, N.; Akhtar, M. Structure and function of the type III pullulan hydrolase from Thermococcus kodakarensis. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 305–314. [Google Scholar] [CrossRef]
- Ryan, S.M.; Fitzgerald, G.F.; Van Sinderen, D. Screening for and identification of starch-, amylopectin-, and pullulan-degrading activities in bifidobacterial strains. Appl. Environ. Microbiol. 2006, 72, 5289–5296. [Google Scholar] [CrossRef]
- Ahmad, N.; Rashid, N.; Haider, M.S.; Akram, M.; Akhtar, M. Novel maltotriose-hydrolyzing thermoacidophilic type III pullulan hydrolase from Thermococcus kodakarensisxs. Appl. Environ. Microbiol. 2014, 80, 1108–1115. [Google Scholar] [CrossRef]
- Zeng, J.; He, C.; Guo, J.; Yuan, L. Effect of truncation of N-Terminal structural modules on enzymatic properties of thermoacidiphilic type III pullulan hydrolase TK-PUL. Shipin Kexue/Food Sci. 2021, 42, 201–208. [Google Scholar]
- Niehaus, F.; Peters, A.; Groudieva, T.; Antranikian, G. Cloning, expression and biochemical characterisation of a unique thermostable pullulan-hydrolysing enzyme from the hyperthermophilic archaeon Thermococcus aggregans. FEMS Microbiol. Lett. 2000, 190, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Rashid, N.; Shehzad, A.; Ahmad, N.; Hussain, Z.; Haider, M.S. Valorization of waste foods using pullulan hydrolase from Thermococcus kodakarensis. Amylase 2019, 2, 39–43. [Google Scholar] [CrossRef]
- Møller, M.S.; Henriksen, A.; Svensson, B. Structure and function of α-glucan debranching enzymes. Cell. Mol. Life Sci. 2016, 73, 2619–2641. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Imai, T. Bacillus stearothermophilus KP 1064 pullulan hydrolase—Its assignment to a unique type of maltogenic α-amylase but to neither pullulanase nor isopullulanase. Appl. Microbiol. Biotechnol. 1985, 21, 20–26. [Google Scholar]
- Toor, K.J.; Ahmad, N.; Muhammad, M.A.; Rashid, N. TK-PUL, a pullulan hydrolase type III from Thermococcus kodakarensis, a potential candidate for simultaneous liquefaction and saccharification of starch. Amylase 2021, 4, 45–55. [Google Scholar] [CrossRef]
- Akeboshi, H.; Tonozuka, T.; Furukawa, T.; Ichikawa, K.; Aoki, H.; Shimonishi, A.; Nishikawa, A.; Sakano, Y. Insights into the reaction mechanism of glycosyl hydrolase family 49: Site-directed mutagenesis and substrate preference of isopullulanase. Eur. J. Biochem. 2004, 271, 4420–4427. [Google Scholar] [CrossRef]
- Aoki, H.; Yopi; Sakano, Y. Molecular cloning and heterologous expression of the isopullulanase gene from Aspergillus niger A.T.C.C. 9642. Biochem. J. 1997, 323, 757–764. [Google Scholar] [CrossRef][Green Version]
- Mizuno, M.; Koide, A.; Yamamura, A.; Akeboshi, H.; Yoshida, H.; Kamitori, S.; Sakano, Y.; Nishikawa, A.; Tonozuka, T. Crystal Structure of Aspergillus niger Isopullulanase, a Member of Glycoside Hydrolase Family 49. J. Mol. Biol. 2008, 376, 210–220. [Google Scholar] [CrossRef]
- Kamasaka, H.; Sugimoto, K.; Takata, H.; Nishimura, T.; Kuriki, T. Bacillus stearothermophilus neopullulanase selective hydrolysis of amylose to maltose in the presence of amylopectin. Appl. Environ. Microbiol. 2002, 68, 1658–1664. [Google Scholar] [CrossRef]
- Jung, J.H.; An, Y.K.; Son, S.Y.; Jeong, S.Y.; Seo, D.H.; Kim, M.K.; Park, C.S. Characterization of a novel extracellular α-amylase from Ruminococcus bromii ATCC 27255 with neopullulanase-like activity. Int. J. Biol. Macromol. 2019, 130, 605–614. [Google Scholar] [CrossRef] [PubMed]
- van Eck, N.J.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Waśko, A.; Polak-Berecka, M.; Targoński, Z. Purification and characterization of pullulanase from Lactococcus lactis. Prep. Biochem. Biotechnol. 2011, 41, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Kunamneni, A.; Singh, S. Improved high thermal stability of pullulanase from a newly isolated thermophilic Bacillus sp. AN-7. Enzym. Microb. Technol. 2006, 39, 1399–1404. [Google Scholar] [CrossRef]
- Xu, J.; Ren, F.; Huang, C.H.; Zheng, Y.; Zhen, J.; Sun, H.; Ko, T.P.; He, M.; Chen, C.C.; Chan, H.C.; et al. Functional and structural studies of pullulanase from Anoxybacillus sp. LM18-11. Proteins Struct. Funct. Bioinform. 2014, 82, 1685–1693. [Google Scholar] [CrossRef]
- Yang, S.; Yan, Q.; Bao, Q.; Liu, J.; Jiang, Z. Expression and biochemical characterization of a novel type I pullulanase from Bacillus megaterium. Biotechnol. Lett. 2017, 39, 397–405. [Google Scholar] [CrossRef]
- Li, X.; Zhao, J.; Fu, J.; Pan, Y.; Li, D. Sequence analysis and biochemical properties of an acidophilic and hyperthermophilic amylopullulanase from Thermofilum pendens. Int. J. Biol. Macromol. 2018, 114, 235–243. [Google Scholar] [CrossRef]
- Kahar, U.M.; Chan, K.-G.; Salleh, M.M.; Hii, S.M.; Goh, K.M. A High Molecular-Mass Anoxybacillus sp. SK3-4 Amylopullulanase: Characterization and Its Relationship in Carbohydrate Utilization. Int. J. Mol. Sci. 2013, 14, 11302–11318. [Google Scholar] [CrossRef]
- Mesbah, N.M.; Wiegel, J. Biochemical characterization of halophilic, alkalithermophilic amylopullulanase PulD7 and truncated amylopullulanases PulD7ΔN and PulD7ΔC. Int. J. Biol. Macromol. 2018, 111, 632–638. [Google Scholar] [CrossRef]
- Roodi, F.Z.; Aminzadeh, S.; Farrokhi, N.; Karkhane, A.A.; Haghbeen, K. Cohnella amylopullulanases: Biochemical characterization of two recombinant thermophilic enzymes. PLoS ONE 2017, 12, e0175013. [Google Scholar]
- Vishnu, C.; Naveena, B.J.; Altaf, M.; Venkateshwar, M.; Reddy, G. Amylopullulanase—A novel enzyme of L. amylophilus GV6 in direct fermentation of starch to L(+) lactic acid. Enzym. Microb. Technol. 2006, 38, 545–550. [Google Scholar] [CrossRef]
- Motherway, M.O.C.; Fitzgerald, G.F.; Neirynck, S.; Ryan, S.; Steidler, L.; Van Sinderen, D. Characterization of ApuB, an extracellular type II amylopullulanase from Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2008, 74, 6271–6279. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, M.L.; Fuentes, S.; De Greeff, A.; Smith, H.; Wells, J.M. ApuA, a multifunctional α-glucan-degrading enzyme of Streptococcus suis, mediates adhesion to porcine epithelium and mucus. Microbiology 2010, 156, 2818–2828. [Google Scholar] [CrossRef]
- Kim, J.H.; Sunako, M.; Ono, H.; Murooka, Y.; Fukusaki, E.; Yamashita, M. Characterization of gene encoding amylopullulanase from plant-originated lactic acid bacterium, Lactobacillus plantarum L137. J. Biosci. Bioeng. 2008, 106, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Lin, Y.; Wang, G.; Lin, J. Gene cloning, expression and biochemical characterization of a new multi-domain, halotolerant and SDS-resistant alkaline pullulanase from Alkalibacterium sp. SL3. Process Biochem. 2020, 96, 1–10. [Google Scholar] [CrossRef]
- Roy, A.; Messaoud, E.B.; Bejar, S. Isolation and purification of an acidic pullulanase type II from newly isolated Bacillus sp. US149. Enzym. Microb. Technol. 2003, 33, 720–724. [Google Scholar] [CrossRef]
- Igarashi, K.; Ara, K.; Saeki, K.; Ozaki, K.; Kawai, S.; Ito, S. Nucleotide sequence of the gene that encodes a neopullulanase from an alkalophilic Bacillus. Biosci. Biotechnol. Biochem. 1992, 56, 514–516. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Van Der Maarel, M.J.E.C.; Van Der Veen, B.; Uitdehaag, J.C.M.; Leemhuis, H.; Dijkhuizen, L. Properties and applications of starch-converting enzymes of the α-amylase family. J. Biotechnol. 2002, 94, 137–155. [Google Scholar] [CrossRef]
- Christiansen, C.; Abou Hachem, M.; Janeček, Š.; Viksø-Nielsen, A.; Blennow, A.; Svensson, B. The carbohydrate-binding module family 20—Diversity, structure, and function. FEBS J. 2009, 276, 5006–5029. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, H.; Kim, Y.-J.; Jung, D.-H.; Seo, D.-H.; Jung, J.-H.; Park, C.-S. Enzymatic analysis of truncation mutants of a type II pullulanase from Bifidobacterium adolescentis P2P3, a resistant starch-degrading gut bacterium. Int. J. Biol. Macromol. 2021, in press. [Google Scholar] [CrossRef]
- Robert, X.; Haser, R.; Gottschalk, T.E.; Ratajczak, F.; Driguez, H.; Svensson, B.; Aghajari, N. The structure of barley α-amylase isozyme 1 reveals a novel role of domain C in substrate recognition and binding: A pair of sugar tongs. Structure 2003, 11, 973–984. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef]
- Eş, I.; Vieira, J.D.G.; Amaral, A.C. Principles, techniques, and applications of biocatalyst immobilization for industrial application. Appl. Microbiol. Biotechnol. 2015, 99, 2065–2082. [Google Scholar] [CrossRef] [PubMed]
- Vaghari, H.; Jafarizadeh-Malmiri, H.; Mohammadlou, M.; Berenjian, A.; Anarjan, N.; Jafari, N.; Nasiri, S. Application of magnetic nanoparticles in smart enzyme immobilization. Biotechnol. Lett. 2016, 38, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Galan, C.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of different enzyme immobilization strategies to improve enzyme performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEA®s): Stable and recyclable biocatalysts. Biochem. Soc. Trans. 2007, 35, 1583–1587. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Basso, A.; Brady, D. New frontiers in enzyme immobilisation: Robust biocatalysts for a circular bio-based economy. Chem. Soc. Rev. 2021, 50, 5850–5862. [Google Scholar] [CrossRef]
- Bernal, C.; Rodríguez, K.; Martínez, R. Integrating enzyme immobilization and protein engineering: An alternative path for the development of novel and improved industrial biocatalysts. Biotechnol. Adv. 2018, 36, 1470–1480. [Google Scholar] [CrossRef]
- Mylkie, K.; Nowak, P.; Rybczynski, P.; Ziegler-Borowska, M. Polymer-coated magnetite nanoparticles for protein immobilization. Materials 2021, 14, 248. [Google Scholar] [CrossRef] [PubMed]
- Polaina, J.; MacCabe, A.P. Industrial Enzymes: Structure, Function and Applications; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Meryam Sardar, R.A. Enzyme Immobilization: An Overview on Nanoparticles as Immobilization Matrix. Biochem. Anal. Biochem. 2015, 4, 1. [Google Scholar] [CrossRef]
- Hwang, E.T.; Lee, S. Multienzymatic Cascade Reactions via Enzyme Complex by Immobilization. ACS Catal. 2019, 9, 4402–4425. [Google Scholar] [CrossRef]
- Gennari, A.; Führ, A.J.; Volpato, G.; Volken de Souza, C.F. Magnetic cellulose: Versatile support for enzyme immobilization—A review. Carbohydr. Polym. 2020, 246, 116646. [Google Scholar] [CrossRef]
- Mateo, C.; Grazú, V.; Pessela, B.C.C.; Montes, T.; Palomo, J.M.; Torres, R.; López-Gallego, F.; Fernández-Lafuente, R.; Guisán, J.M. Advances in the design of new epoxy supports for enzyme immobilization-stabilization. Biochem. Soc. Trans. 2007, 35, 1593–1601. [Google Scholar] [CrossRef]
- Contesini, F.J.; Figueira, J.d.A.; Kawaguti, H.Y.; Fernandes, P.C.d.B.; Carvalho, P.d.O.; Nascimento, M.d.G.; Sato, H.H. Potential applications of carbohydrases immobilization in the food industry. Int. J. Mol. Sci. 2013, 14, 1335–1369. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Zhou, Z. Regulation of the catalytic behavior of pullulanases chelated onto nickel (II)-modified magnetic nanoparticles. Enzym. Microb. Technol. 2017, 101, 9–16. [Google Scholar] [CrossRef]
- Kuriki, T.; Tsuda, M.; Imanaka, T. Continuous production of panose by immobilized neopullulanase. J. Ferment. Bioeng. 1992, 73, 198–202. [Google Scholar] [CrossRef]
- Lenders, J.-P.; Crichton, R.R. Thermal stabilization of amylolytic enzymes by covalent coupling to soluble polysaccharides. Biotechnol. Bioeng. 1984, 26, 1343–1351. [Google Scholar] [CrossRef]
- Ali, G.; Dulong, V.; Gasmi, S.N.; Rihouey, C.; Picton, L.; Le Cerf, D. Covalent immobilization of pullulanase on alginate and study of its hydrolysis of pullulan. Biotechnol. Prog. 2015, 31, 883–889. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Zhou, Z. Improving pullulanase catalysis via reversible immobilization on modified Fe3O4@polydopamine nanoparticles. Appl. Biochem. Biotechnol. 2017, 182, 1467–1477. [Google Scholar] [CrossRef]
- Long, J.; Li, X.; Zhan, X.; Xu, X.; Tian, Y.; Xie, Z.; Jin, Z. Sol–gel encapsulation of pullulanase in the presence of hybrid magnetic (Fe3O4–chitosan) nanoparticles improves thermal and operational stability. Bioprocess Biosyst. Eng. 2017, 40, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Fernández-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.A.; Fernandez-Lafuente, R. Enhancing the functional properties of thermophilic enzymes by chemical modification and immobilization. Enzym. Microb. Technol. 2011, 49, 326–346. [Google Scholar] [CrossRef] [PubMed]
- Kahar, U.M.; Chan, K.G.; Sani, M.H.; Mohd Noh, N.I.; Goh, K.M. Effects of single and co-immobilization on the product specificity of type I pullulanase from Anoxybacillus sp. SK3-4. Int. J. Biol. Macromol. 2017, 104, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Wang, B.; Tan, J.; Zhu, L.; Li, L. Immobilized multienzymatic systems for catalysis of cascade reactions. Process Biochem. 2016, 51, 1193–1203. [Google Scholar] [CrossRef]
- Schoffelen, S.; Van Hest, J.C.M. Chemical approaches for the construction of multi-enzyme reaction systems. Curr. Opin. Struct. Biol. 2013, 23, 613–621. [Google Scholar] [CrossRef]
- Talekar, S.; Pandharbale, A.; Ladole, M.; Nadar, S.; Mulla, M.; Japhalekar, K.; Pattankude, K.; Arage, D. Carrier free co-immobilization of alpha amylase, glucoamylase and pullulanase as combined cross-linked enzyme aggregates (combi-CLEAS): A tri-enzyme biocatalyst with one pot starch hydrolytic activity. Bioresour. Technol. 2013, 147, 269–275. [Google Scholar] [CrossRef]
- Xue, R.; Woodley, J.M. Process technology for multi-enzymatic reaction systems. Bioresour. Technol. 2012, 115, 183–195. [Google Scholar] [CrossRef]
- Kusano, S.; Shiraishi, T.; Takahashi, S.I.; Fujimoto, D.; Sakano, Y. Immobilization of Bacillus acidopullulyticus pullulanase and properties of the immobilized pullulanases. J. Ferment. Bioeng. 1989, 68, 233–237. [Google Scholar] [CrossRef]
- Dessouki, A.M.; Issa, G.I.; Atia, K.S. Pullulanase immobilization on natural and synthetic polymers. J. Chem. Technol. Biotechnol. 2011, 76, 700–706. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Shoda, H.; Ichikawa, S.; Sato, S.; Mukataka, S. Immobilization and stabilization of pullulanase from Klebsiella pneumoniae by a multipoint attachment method using activated agar gel supports. Process Biochem. 2005, 40, 2637–2642. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, X.; Zheng, S.; Sun, H. Photochemical preparation of magnetic chitosan beads for immobilization of pullulanase. Biochem. Eng. J. 2009, 46, 83–87. [Google Scholar] [CrossRef]
- Singh, R.S.; Saini, G.K.; Kennedy, J.F. Covalent immobilization and thermodynamic characterization of pullulanase for the hydrolysis of pullulan in batch system. Carbohydr. Polym. 2010, 81, 252–259. [Google Scholar] [CrossRef]
- Long, J.; Jiao, A.; Wei, B.; Wu, Z.; Zhang, Y.; Xu, X.; Jin, Z. A novel method for pullulanase immobilized onto magnetic chitosan/Fe3O4 composite nanoparticles by in situ preparation and evaluation of the enzyme stability. J. Mol. Catal. B Enzym. 2014, 109, 53–61. [Google Scholar] [CrossRef]
- Long, J.; Li, X.; Wu, Z.; Xu, E.; Xu, X.; Jin, Z.; Jiao, A. Immobilization of pullulanase onto activated magnetic chitosan/Fe3O4 nanoparticles prepared by in situ mineralization and effect of surface functional groups on the stability. Colloids Surf. A Physicochem. Eng. Asp. 2015, 472, 69–77. [Google Scholar] [CrossRef]
- Alagöz, D.; Yildirim, D.; Güvenmez, H.K.; Sihay, D.; Tükel, S.S. Covalent immobilization and characterization of a novel pullulanase from Fontibacillus sp. strain DSHK 107 onto Florisil® and nano-silica for pullulan hydrolysis. Appl. Biochem. Biotechnol. 2016, 179, 1262–1274. [Google Scholar] [CrossRef]
- Ohba, R.; Chaen, H.; Hayashi, S.; Ueda, S. Immobilization of Streptomyces flavochromogenes pullulanase on tannic acid and TEAE–cellulose. Biotechnol. Bioeng. 1978, 20, 665–676. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, L.; Wang, F.; Zhang, J.; Liu, G.; Gao, B.; Wei, D. Novel application of magnetic protein: Convenient one-step purification and immobilization of proteins. Sci. Rep. 2017, 7, 13329. [Google Scholar] [CrossRef]
- Ohtani, N.; Ishidao, T.; Iwai, Y.; Arai, Y. Hydrolysis of starch or pullulan by glucoamylase or pullulanase immobilized on poly(N-isopropylacrylamide) gel. Colloid Polym. Sci. 1999, 277, 197–202. [Google Scholar] [CrossRef]
- Roy, I.; Gupta, M.N. Hydrolysis of starch by a mixture of glucoamylase and pullulanase entrapped individually in calcium alginate beads. Enzym. Microb. Technol. 2004, 34, 26–32. [Google Scholar] [CrossRef]
- Long, J.; Wu, Z.; Li, X.; Xu, E.; Xu, X.; Jin, Z.; Jiao, A. New method for the immobilization of pullulanase onto hybrid magnetic (Fe3O4-κ-carrageenan) nanoparticles by electrostatic coupling with pullulanase/chitosan complex. J. Agric. Food Chem. 2015, 63, 3534–3542. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Ma, X.; Han, Y.; Wei, Y.; Guo, Q.; Yang, S.; Zhang, Y.; Liao, W.; Gao, Y. Effect of chitosan molecular weight on zein-chitosan nanocomplexes: Formation, characterization, and the delivery of quercetagetin. Int. J. Biol. Macromol. 2020, 164, 2215–2223. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, F.; Lin, L.; He, D.; Chen, J.; Wei, W.; Wei, D. Biochemical characterization of a novel thermostable type I pullulanase produced recombinantly in Bacillus subtilis. Starch/Staerke 2018, 70, 1700179. [Google Scholar] [CrossRef]
- Long, J.; Zhang, B.; Li, X.; Zhan, X.; Xu, X.; Xie, Z.; Jin, Z. Effective production of resistant starch using pullulanase immobilized onto magnetic chitosan/Fe3O4 nanoparticles. Food Chem. 2018, 239, 276–286. [Google Scholar] [CrossRef]
- Long, J.; Li, X.; Liu, X.; Jin, Z.; Xie, Z.; Xu, X.; Lu, C. Preparation of streptavidin-coated magnetic nanoparticles for specific immobilization of enzymes with high activity and enhanced stability. Ind. Eng. Chem. Res. 2021, 60, 1542–1552. [Google Scholar] [CrossRef]
- Ram, K.A.; Venkatasubramanian, K. Enhancement of starch conversion efficiency with free and immobilized pullulanase and α-1,4-glucosidase. Biotechnol. Bioeng. 1982, 24, 355–369. [Google Scholar] [CrossRef]
- Chakrabarti, A.C.; Storey, K.B. Co-immobilization of amyloglucosidase and pullulanase for enhanced starch hydrolysis. Appl. Microbiol. Biotechnol. 1990, 33, 48–50. [Google Scholar] [CrossRef]
- Atia, K.S.; Ismail, S.A.; El-Arnaouty, M.B.; Dessouki, A.M. Use of Co-immobilized β-amylase and pullulanase in reduction of saccharification time of starch and increase in maltose yield. Biotechnol. Prog. 2003, 19, 853–857. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Kambale, S.; Jadhav, S.; Nadar, S.; Ladole, M. A tri-enzyme magnetic nanobiocatalyst with one pot starch hydrolytic activity. Chem. Eng. J. 2017, 325, 80–90. [Google Scholar] [CrossRef]
- Yoshida, M.; Oishi, K.; Kimura, T.; Ogata, M.; Nakakuki, T. Continuous production of maltose using a dual immobilized enzyme system. Agric. Biol. Chem. 1989, 53, 3139–3142. [Google Scholar] [CrossRef]
- Kimura, T.; Ogata, M.; Kobayashi, H.; Yoshida, M.; Oishi, K.; Nakakuki, T. Continuous production of maltotetraose using a dual immobilized enzyme system of maltotetraose-forming amylase and pullulanase. Biotechnol. Bioeng. 1990, 36, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Schafhagser, D.Y.; Storey, K.B. Fructose production: Coimmobilized amyloglucosidase, pullulanase, and glucose isomerase on BIOBONETM. Appl. Biochem. Biotechnol. Part A Enzym. Eng. Biotechnol. 1992, 36, 63–74. [Google Scholar] [CrossRef]
- Noda, T.; Furuta, S.; Suda, I. Sweet potato β-amylase immobilized on chitosan beads and its application in the semi-continuous production of maltose. Carbohydr. Polym. 2001, 44, 189–195. [Google Scholar] [CrossRef]
- Talekar, S.; Desai, S.; Pillai, M.; Nagavekar, N.; Ambarkar, S.; Surnis, S.; Ladole, M.; Nadar, S.; Mulla, M. Carrier free co-immobilization of glucoamylase and pullulanase as combi-cross linked enzyme aggregates (combi-CLEAs). RSC Adv. 2013, 3, 2265–2271. [Google Scholar] [CrossRef]
- Goh, K.M.; Hong, G.P.; Chyi, N.H.; Ng, P.; Piaw, C.K.; Rahman, R.N.Z.R.A. Trends and tips in protein engineering, a review. J. Teknol. 2012, 59, 21–31. [Google Scholar]
- Ece, S.; Evran, S.; Janda, J.O.; Merkl, R.; Sterner, R. Improving thermal and detergent stability of Bacillus stearothermophilus neopullulanase by rational enzyme design. Protein Eng. Des. Sel. 2014, 28, 147–151. [Google Scholar] [CrossRef]
- Miyazaki, T.; Yashiro, H.; Nishikawa, A.; Tonozuka, T. The side chain of a glycosylated asparagine residue is important for the stability of isopullulanase. J. Biochem. 2015, 157, 225–234. [Google Scholar] [CrossRef]
- Zeng, Y.; Zheng, H.; Shen, Y.; Xu, J.; Tan, M.; Liu, F.; Song, H. Identification and analysis of binding residues in the CBM68 of pullulanase PulA from Anoxybacillus sp. LM18-11. J. Biosci. Bioeng. 2019, 127, 8–15. [Google Scholar] [CrossRef]
- Li, S.F.; Xu, J.Y.; Bao, Y.J.; Zheng, H.C.; Song, H. Structure and sequence analysis-based engineering of pullulanase from Anoxybacillus sp. LM18-11 for improved thermostability. J. Biotechnol. 2015, 210, 8–14. [Google Scholar] [CrossRef]
- Duan, X.; Chen, J.; Wu, J. Improving the thermostability and catalytic efficiency of Bacillus deramificans pullulanase by site-directed mutagenesis. Appl. Environ. Microbiol. 2013, 79, 4072–4077. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Chu, X.; Lv, J.; Li, Q.; Tian, J.; Wu, N. Improving the thermostability of acidic pullulanase from bacillus naganoensis by rational design. PLoS ONE 2016, 11, e0165006. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Xie, N.Z.; Du, Q.S.; Qin, Y.; Li, J.X.; Meng, J.Z.; Huang, R.B. Active hydrogen bond network (AHBN) and applications for improvement of thermal stability and pH-sensitivity of pullulanase from Bacillus naganoensis. PLoS ONE 2017, 12, e0169080. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Nie, Y.; Mu, X.; Xu, Y.; Xiao, R. Disorder prediction-based construct optimization improves activity and catalytic efficiency of Bacillus naganoensis pullulanase. Sci. Rep. 2016, 6, 24574. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, F.; Lin, L.; He, D.; Wei, W.; Wei, D. N-Terminal Domain Truncation and Domain Insertion-Based Engineering of a Novel Thermostable Type i Pullulanase from Geobacillus thermocatenulatus. J. Agric. Food Chem. 2018, 66, 10788–10798. [Google Scholar] [CrossRef]
- Bi, J.; Chen, S.; Zhao, X.; Nie, Y.; Xu, Y. Computation-aided engineering of starch-debranching pullulanase from Bacillus thermoleovorans for enhanced thermostability. Appl. Microbiol. Biotechnol. 2020, 104, 7551–7562. [Google Scholar] [CrossRef]
- Mu, G.C.; Nie, Y.; Mu, X.Q.; Xu, Y.; Xiao, R. Single Amino Acid Substitution in the Pullulanase of Klebsiella variicola for Enhancing Thermostability and Catalytic Efficiency. Appl. Biochem. Biotechnol. 2015, 176, 1736–1745. [Google Scholar] [CrossRef]
- Pang, B.; Zhou, L.; Cui, W.; Liu, Z.; Zhou, Z. Improvement of the Thermostability and Activity of Pullulanase from Anoxybacillus sp. WB42. Appl. Biochem. Biotechnol. 2020, 191, 942–954. [Google Scholar] [CrossRef]
- Nisha, M.; Satyanarayana, T. The role of N1 domain on the activity, stability, substrate specificity and raw starch binding of amylopullulanase of the extreme thermophile Geobacillus thermoleovorans. Appl. Microbiol. Biotechnol. 2015, 99, 5461–5474. [Google Scholar] [CrossRef]
- Nisha, M.; Satyanarayana, T. Characterization of recombinant amylopullulanase (gt-apu) and truncated amylopullulanase (gt-apuT) of the extreme thermophile Geobacillus thermoleovorans NP33 and their action in starch saccharification. Appl. Microbiol. Biotechnol. 2013, 97, 6279–6292. [Google Scholar] [CrossRef]
- Lin, F.P.; Ho, Y.H.; Lin, H.Y.; Lin, H.J. Effect of C-terminal truncation on enzyme properties of recombinant amylopullulanase from Thermoanaerobacter pseudoethanolicus. Extremophiles 2012, 16, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Chuang, H.H.; Lin, F.P. Biochemical characterization of engineered amylopullulanase from Thermoanaerobacter ethanolicus 39E-implicating the non-necessity of its 100 C-terminal amino acid residues. Extremophiles 2008, 12, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Sunako, M.; Ono, H.; Murooka, Y.; Fukusaki, E.; Yamashita, M. Characterization of the C-terminal truncated form of amylopullulanase from Lactobacillus plantarum L137. J. Biosci. Bioeng. 2009, 107, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Kahar, U.M.; Ng, C.L.; Chan, K.-G.; Goh, K.M. Characterization of a type I pullulanase from Anoxybacillus sp. SK3-4 reveals an unusual substrate hydrolysis. Appl. Microbiol. Biotechnol. 2016, 100, 6291–6307. [Google Scholar] [CrossRef]
- Chen, A.; Xu, T.; Ge, Y.; Wang, L.; Tang, W.; Li, S. Hydrogen-bond-based protein engineering for the acidic adaptation of Bacillus acidopullulyticus pullulanase. Enzym. Microb. Technol. 2019, 124, 79–83. [Google Scholar] [CrossRef]
- Wang, X.; Jing, X.; Deng, Y.; Nie, Y.; Xu, F.; Xu, Y.; Zhao, Y.L.; Hunt, J.F.; Montelione, G.T.; Szyperski, T. Evolutionary coupling saturation mutagenesis: Coevolution-guided identification of distant sites influencing Bacillus naganoensis pullulanase activity. FEBS Lett. 2020, 594, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Jing, X.; Wu, L.; Zhou, X.; Gu, J.; Nie, Y.; Xu, Y. Computational design of noncanonical amino acid-based thioether staples at N/C-terminal domains of multi-modular pullulanase for thermostabilization in enzyme catalysis. Comput. Struct. Biotechnol. J. 2021, 19, 577–585. [Google Scholar] [CrossRef]
- Wang, X.; Nie, Y.; Xu, Y. Improvement of the Activity and Stability of Starch-Debranching Pullulanase from Bacillus naganoensis via Tailoring of the Active Sites Lining the Catalytic Pocket. J. Agric. Food Chem. 2018, 66, 13236–13242. [Google Scholar] [CrossRef]
- Bava, K.A.; Gromiha, M.M.; Uedaira, H.; Kitajima, K.; Sarai, A. ProTherm, version 4.0: Thermodynamic database for proteins and mutants. Nucleic Acids Res. 2004, 32, D120–D121. [Google Scholar] [CrossRef]
- Montanucci, L.; Capriotti, E.; Frank, Y.; Ben-Tal, N.; Fariselli, P. DDGun: An untrained method for the prediction of protein stability changes upon single and multiple point variations. BMC Bioinform. 2019, 20, 335. [Google Scholar] [CrossRef]
- Schymkowitz, J.; Borg, J.; Stricher, F.; Nys, R.; Rousseau, F.; Serrano, L. The FoldX web server: An online force field. Nucleic Acids Res. 2005, 33, W382–W388. [Google Scholar] [CrossRef]
- Quan, L.; Lv, Q.; Zhang, Y. STRUM: Structure-based prediction of protein stability changes upon single-point mutation. Bioinformatics 2016, 32, 2936–2946. [Google Scholar] [CrossRef]
- Pandurangan, A.P.; Ochoa-Montaño, B.; Ascher, D.B.; Blundell, T.L. SDM: A server for predicting effects of mutations on protein stability. Nucleic Acids Res. 2017, 45, W229–W235. [Google Scholar] [CrossRef]
- Yang, Y.; Ding, X.; Zhu, G.; Niroula, A.; Lv, Q.; Vihinen, M. ProTstab—Predictor for cellular protein stability. BMC Genom. 2019, 20, 804–809. [Google Scholar] [CrossRef]
- Cao, H.; Wang, J.; He, L.; Qi, Y.; Zhang, J.Z. DeepDDG: Predicting the Stability Change of Protein Point Mutations Using Neural Networks. J. Chem. Inf. Model. 2019, 59, 1508–1514. [Google Scholar] [CrossRef]
- Chen, C.W.; Lin, M.H.; Liao, C.C.; Chang, H.P.; Chu, Y.W. iStable 2.0: Predicting protein thermal stability changes by integrating various characteristic modules. Comput. Struct. Biotechnol. J. 2020, 18, 622–630. [Google Scholar] [CrossRef]
- Yang, Y.; Urolagin, S.; Niroula, A.; Ding, X.; Shen, B.; Vihinen, M. Pon-tstab: Protein variant stability predictor. Importance of training data quality. Int. J. Mol. Sci. 2018, 19, 1009. [Google Scholar] [CrossRef]
- Sanavia, T.; Birolo, G.; Montanucci, L.; Turina, P.; Capriotti, E.; Fariselli, P. Limitations and challenges in protein stability prediction upon genome variations: Towards future applications in precision medicine. Comput. Struct. Biotechnol. J. 2020, 18, 1968–1979. [Google Scholar] [CrossRef]
- Goh, K.M.; Shahar, S.; Chan, K.-G.; Chong, C.S.; Amran, S.I.; Sani, M.H.; Zakaria, I.I.; Kahar, U.M. Current status and potential applications of underexplored prokaryotes. Microorganisms 2019, 7, 468. [Google Scholar] [CrossRef]
- Urbieta, M.S.; Donati, E.R.; Chan, K.-G.; Shahar, S.; Sin, L.L.; Goh, K.M. Thermophiles in the genomic era: Biodiversity, science, and applications. Biotechnol. Adv. 2015, 33, 633–647. [Google Scholar] [CrossRef]
- Kaushal, G.; Kumar, J.; Sangwan, R.S.; Singh, S.P. Metagenomic analysis of geothermal water reservoir sites exploring carbohydrate-related thermozymes. Int. J. Biol. Macromol. 2018, 119, 882–895. [Google Scholar] [CrossRef]
- Reichart, N.J.; Bowers, R.M.; Woyke, T. High potential for biomass-degrading enzymes revealed by hot spring metagenomics. Front. Microbiol. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Liew, K.J.; Lim, C.C.; Chan, C.S.; Wei, K.Y.; Salleh, M.M.; Sani, R.K.; Chan, K.G.; Goh, K.M. Direct Cellulase Gene Amplification from Hot Spring Using the Guidance of 16S rRNA Amplicon Metagenomics; Academic Press: London, UK, 2018; ISBN 9780128134030. [Google Scholar]
- Perez-Mon, C.; Qi, W.; Vikram, S.; Frossard, A.; Makhalanyane, T.; Cowan, D.; Frey, B. Shotgun metagenomics reveals distinct functional diversity and metabolic capabilities between 12,000-year-old permafrost and active layers on Muot da Barba Peider (Swiss Alps). Microb. Genom. 2021, 7, 000558. [Google Scholar]
- Sadeghian Motahar, S.F.; Salami, M.; Ariaeenejad, S.; Emam-Djomeh, Z.; Sheykh Abdollahzadeh Mamaghani, A.; Kavousi, K.; Moghadam, M.; Hosseini Salekdeh, G. Synergistic effect of metagenome-derived starch-degrading enzymes on quality of functional bread with antioxidant activity. Starch/Staerke 2021, 74, 1–2. [Google Scholar] [CrossRef]
- Thakur, M.; Sharma, N.; Rai, A.K.; Singh, S.P. A novel cold-active type I pullulanase from a hot-spring metagenome for effective debranching and production of resistant starch. Bioresour. Technol. 2021, 320, 124288. [Google Scholar] [CrossRef]
- Yarygin, K.; Tyakht, A.; Larin, A.; Kostryukova, E.; Kolchenko, S.; Bitner, V.; Alexeev, D. Abundance profiling of specific gene groups using precomputed gut metagenomes yields novel biological hypotheses. PLoS ONE 2017, 12, e0176154. [Google Scholar] [CrossRef]
- Lee, Y.S.; Seo, S.H.; Yoon, S.H.; Kim, S.Y.; Hahn, B.S.; Sim, J.S.; Koo, B.S.; Lee, C.M. Identification of a novel alkaline amylopullulanase from a gut metagenome of Hermetia illucens. Int. J. Biol. Macromol. 2016, 82, 514–521. [Google Scholar] [CrossRef]
- Sadeghian Motahar, S.F.; Ariaeenejad, S.; Salami, M.; Emam-Djomeh, Z.; Sheykh Abdollahzadeh Mamaghani, A. Improving the quality of gluten-free bread by a novel acidic thermostable α-amylase from metagenomics data. Food Chem. 2021, 352, 129307. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Brady, D. The limits to biocatalysis: Pushing the envelope. Chem. Commun. 2018, 54, 6088–6104. [Google Scholar] [CrossRef]
- Dalapati, R.; Nandi, S.; Gogoi, C.; Shome, A.; Biswas, S. Metal–Organic framework (MOF) derived recyclable, superhydrophobic composite of cotton fabrics for the facile removal of oil spills. ACS Appl. Mater. Interfaces 2021, 13, 8563–8573. [Google Scholar] [CrossRef]





| Name | Publications |
|---|---|
| Biochemistry, Genetics and Molecular Biology | 944 |
| Agricultural and Biological Sciences | 504 |
| Immunology and Microbiology | 501 |
| Chemistry | 343 |
| Chemical Engineering | 237 |
| Environmental Science | 88 |
| Engineering | 61 |
| Name | Organization, Country | Publications a | Citations b | Citations Mean b |
|---|---|---|---|---|
| Anthony P Pugsley | Pasteur Institute, France | 56 | 3801 | 66.68 |
| Garabed Antranikian | Hamburg University of Technology, Germany | 33 | 1552 | 48.50 |
| Yoshiyuki Sakano | Tokyo University of Agriculture and Tech., Japan | 28 | 259 | 10.36 |
| Zheng-Yu Jin | Jiangnan University, China | 23 | 552 | 23.00 |
| Birte Svensson | Technical University of Denmark, Denmark | 22 | 618 | 47.54 |
| Takashi Kuriki | Ezaki Glico, Japan | 19 | 538 | 53.80 |
| Yao Nie | Jiangnan University, China | 18 | 203 | 11.94 |
| J Gregory Zeikus | University of Wisconsin-Madison, USA | 16 | 685 | 48.93 |
| Yasunori Nakamura | Akita Prefectural University, Japan | 15 | 1669 | 128.38 |
| Journal Name | Publications |
|---|---|
| Agricultural and Biological Chemistry | 56 |
| Starch—Stärke | 44 |
| Carbohydrate Research | 43 |
| International Journal of Biological Macromolecules | 32 |
| Carbohydrate Polymers | 40 |
| Journal of Bacteriology | 40 |
| Applied Microbiology and Biotechnology | 38 |
| Applied and Environmental Microbiology | 31 |
| Molecular Microbiology | 29 |
| Enzyme and Microbial Technology | 28 |
| Food Chemistry | 25 |
| Bioscience Biotechnology and Biochemistry | 21 |
| Journal of Biological Chemistry | 18 |
| Biotechnology Letters | 17 |
| Biotechnology and Bioengineering | 16 |
| Applied Biochemistry and Biotechnology | 15 |
| FEMS Microbiology Letters | 15 |
| Food Hydrocolloids | 15 |
| Company | Company Headquarter (Founded Year) | Document Count |
|---|---|---|
| Procter & Gamble | Cincinnati, Ohio, USA (1837) | 980 |
| Novozymes | Bagsværd, Denmark (2000) | 624 |
| Danisco (DuPont) | Copenhagen, Denmark (1989) | 502 |
| Henkel AG & Co. KGaA | Düsseldorf, Germany (1876) | 431 |
| BASF Se and BASF Enzyme | Ludwigshafen, Germany (1865) | 388 |
| Verenium Corporation | California, USA (2007) | 194 |
| DuPont Nutrition & Biosciences | Copenhagen, Denmark (1989) | 169 |
| Genencor | California, USA (1982) | 136 |
| DSM | Sittard, Netherlands (1902) | 126 |
| Milliken & Company | South Carolina, USA (1865) | 122 |
| Source | Purpose | Ref. |
|---|---|---|
| Type I pullulanase | ||
| Anoxybacillus sp. LM18-11 | Point mutations at CBM68 to identify changes of substrate affinity. | [120] |
| Anoxybacillus sp. LM18-11 | Enhancing thermostability by multiple point mutations and deleting flexible region at the C-terminus. | [121] |
| Bacillus deramificans | Point mutations at flexible loops to enhance thermostability. | [122] |
| Bacillus naganoensis | Point mutations enhanced thermostability by introducing additional ion interactions. | [123] |
| Bacillus naganoensis | Single points and double mutagenesis to increase “active hydrogen bond network” for enhancing thermostability. | [124] |
| Bacillus naganoensis | Various levels of N- and C-terminal truncation–affected catalytic efficiency. | [125] |
| Geobacillus thermocatenulatus | N-terminal domain truncation and new domain insertion for functional study. | [126] |
| Bacillus thermoleovorans | Point mutations to enhance thermostability. | [127] |
| Klebsiella variicola | Site-saturated mutagenesis to enhance thermostability. | [128] |
| Type II pullulanase (amylopullulanase) | ||
| Anoxybacillus sp. WB42 | Surface Lys replacement and introduction of S = S bond to enhance thermostability. | [129] |
| Geobacillus thermoleovorans | Deletion of N1 domain to understand its role. | [130] |
| Geobacillus thermoleovorans | 300 amino acids C terminal truncation to understand its role. | [131] |
| Thermoanaerobacter pseudoethanolicus | Various extend of C terminal truncation to understand its role. | [132] |
| Thermoanaerobacter ethanolicus | C terminal truncation to understand its role. | [133] |
| Type II pullulanase (α-amylase-pullulanase) | ||
| Lactobacillusplantarum L137 | Removal of 39 repeats (Asp-Ala/Thr-Ala-Asn-Ser-Thr) to understand its role. | [134] |
| Bifidobacterium adolescentis P2P3 | Four variants (i) Deleted α-amylase catalytic domain, (ii) deleted α-amylase catalytic + CBM, (iii) deleted type I pullulan catalytic domain and (iv) (iii) deleted type I pullulan catalytic domain + CBM. To understand biochemical role. | [61] |
| Pullulan hydrolase type I (neopullulanase) | ||
| Geobacillus stearothermophilus | Point mutations to enhance thermostability. | [118] |
| Pullulan hydrolase type II (isopullulanase) | ||
| Aspergillus niger | Point mutations to enhance thermostability. | [119] |
| Aspergillus niger | Point mutations at highly conserved residues to understand its role. | [37] |
| No. | Name of Software or Programs | Remarks | Ref. |
|---|---|---|---|
| 1 | FoldX | Requires high-resolution 3D protein structure. | [142] |
| 2 | STRUM | Can use low-resolution modelled protein structure. STRUM constructs models using I-TASSER. (STRUM is unavailable at the time of writing) | [143] |
| 3 | SDM (Site Directed Mutator) | Developer claimed that predictor do not rely on the number of features for training due to recalculated environment- specific substitution tables (ESSTs). Web-based and easy to use. http://marid.bioc.cam.ac.uk/sdm2/ | [144] |
| 4 | ProTstab | Developed based on machine learning method using gradient boosting of regression trees. Suitable for large scale protein prediction. | [145] |
| 5 | DDGun & DDGun3D | Untrained algorithms that exhibit anti-symmetric prediction property, i.e., ΔΔG(A → B) = −ΔΔG(B → A), where A and B are amino acids. | [141] |
| 6 | DeepDDG | A neural network-based approach and trained using >5700 curated experimental data (ProTherm v.4 + literature data). | [146] |
| 7 | iStable 2.0 | A integrated platform with 11 software: I-Mutant2.0, MUpro and iPTREE-STAB and the structure-based tools I- Mutant2.0, CUPSAT, PoPMuSiC, AUTO-MUTE2.0, SDM, DUET, mCSM, MAESTRO and SDM2. | [147] |
| 8 | DNASTAR | Commercial software Lasergene Protein suite for structural analysis and stability prediction. Academic licensing available. https://www.dnastar.com | nil. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahar, U.M.; Latif, N.A.; Amran, S.I.; Liew, K.J.; Goh, K.M. A Bibliometric Analysis and Review of Pullulan-Degrading Enzymes—Past and Current Trends. Catalysts 2022, 12, 143. https://doi.org/10.3390/catal12020143
Kahar UM, Latif NA, Amran SI, Liew KJ, Goh KM. A Bibliometric Analysis and Review of Pullulan-Degrading Enzymes—Past and Current Trends. Catalysts. 2022; 12(2):143. https://doi.org/10.3390/catal12020143
Chicago/Turabian StyleKahar, Ummirul Mukminin, Nurriza Ab Latif, Syazwani Itri Amran, Kok Jun Liew, and Kian Mau Goh. 2022. "A Bibliometric Analysis and Review of Pullulan-Degrading Enzymes—Past and Current Trends" Catalysts 12, no. 2: 143. https://doi.org/10.3390/catal12020143
APA StyleKahar, U. M., Latif, N. A., Amran, S. I., Liew, K. J., & Goh, K. M. (2022). A Bibliometric Analysis and Review of Pullulan-Degrading Enzymes—Past and Current Trends. Catalysts, 12(2), 143. https://doi.org/10.3390/catal12020143