
Structure-Function and Industrial Relevance of Bacterial Aminopeptidase P

 , , ,
, , ,
Abstract
1. Introduction
2. Structure and Function
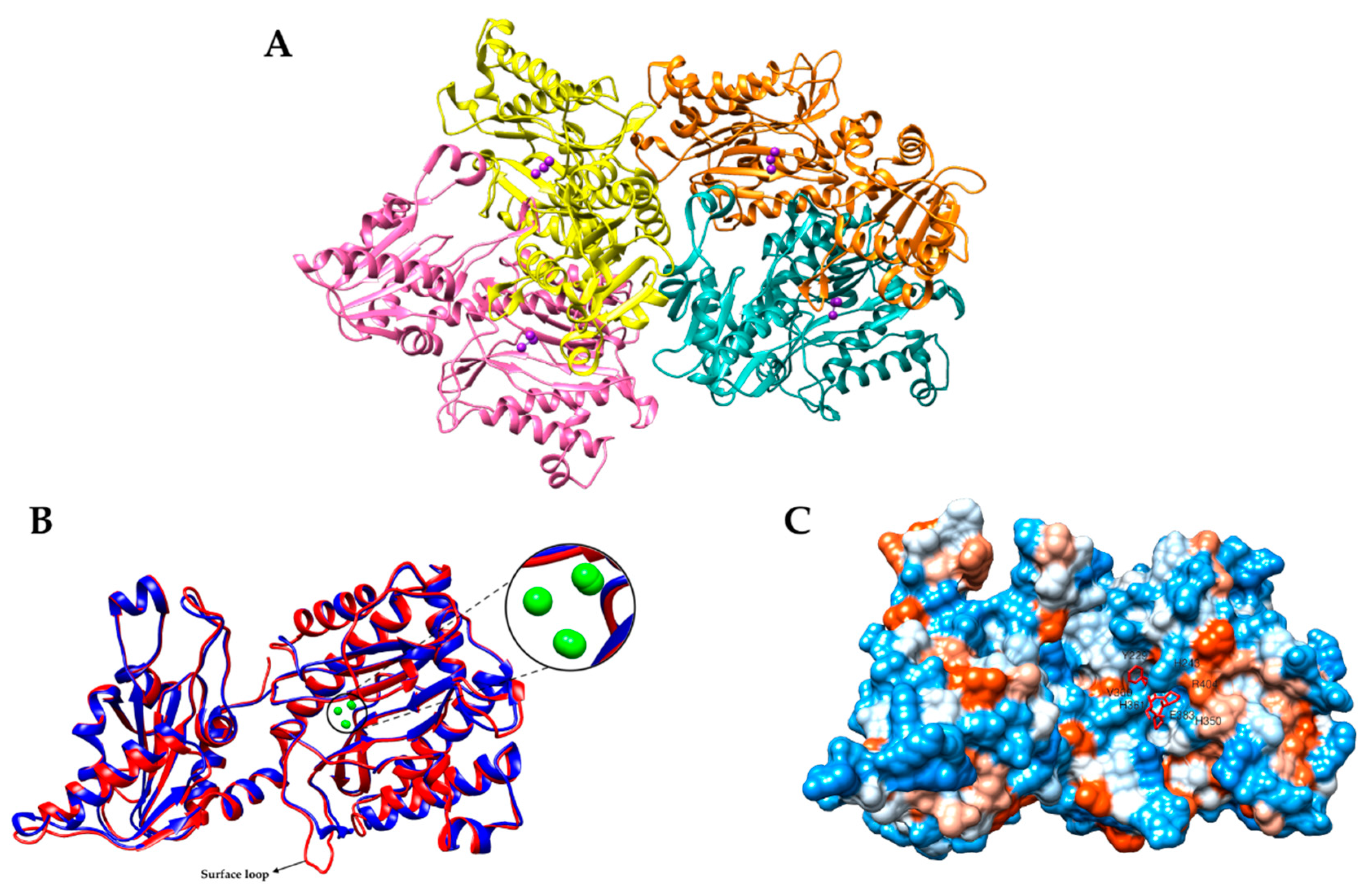
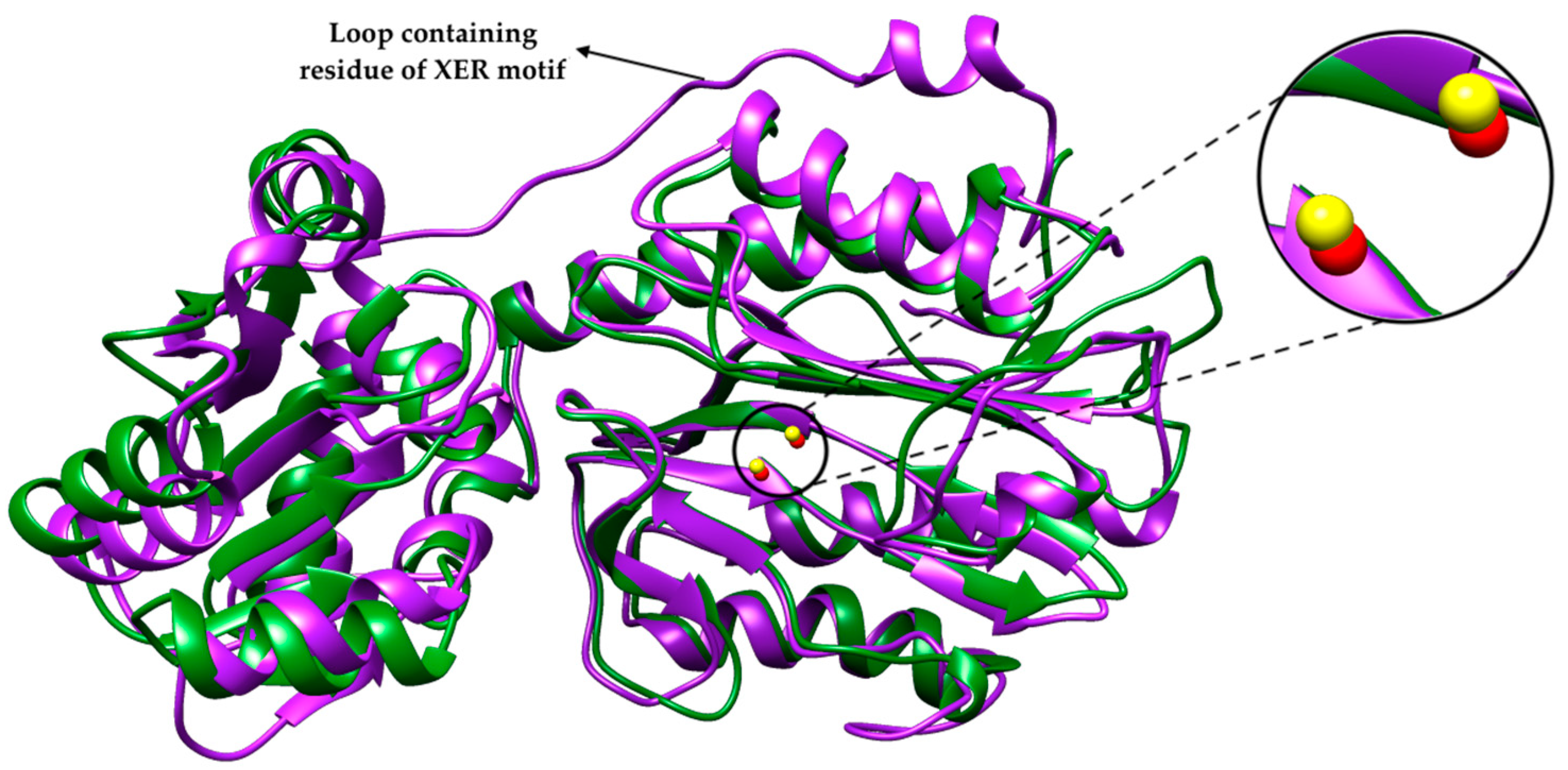
2.1. Bacterial Aminopeptidase P 3D Structure
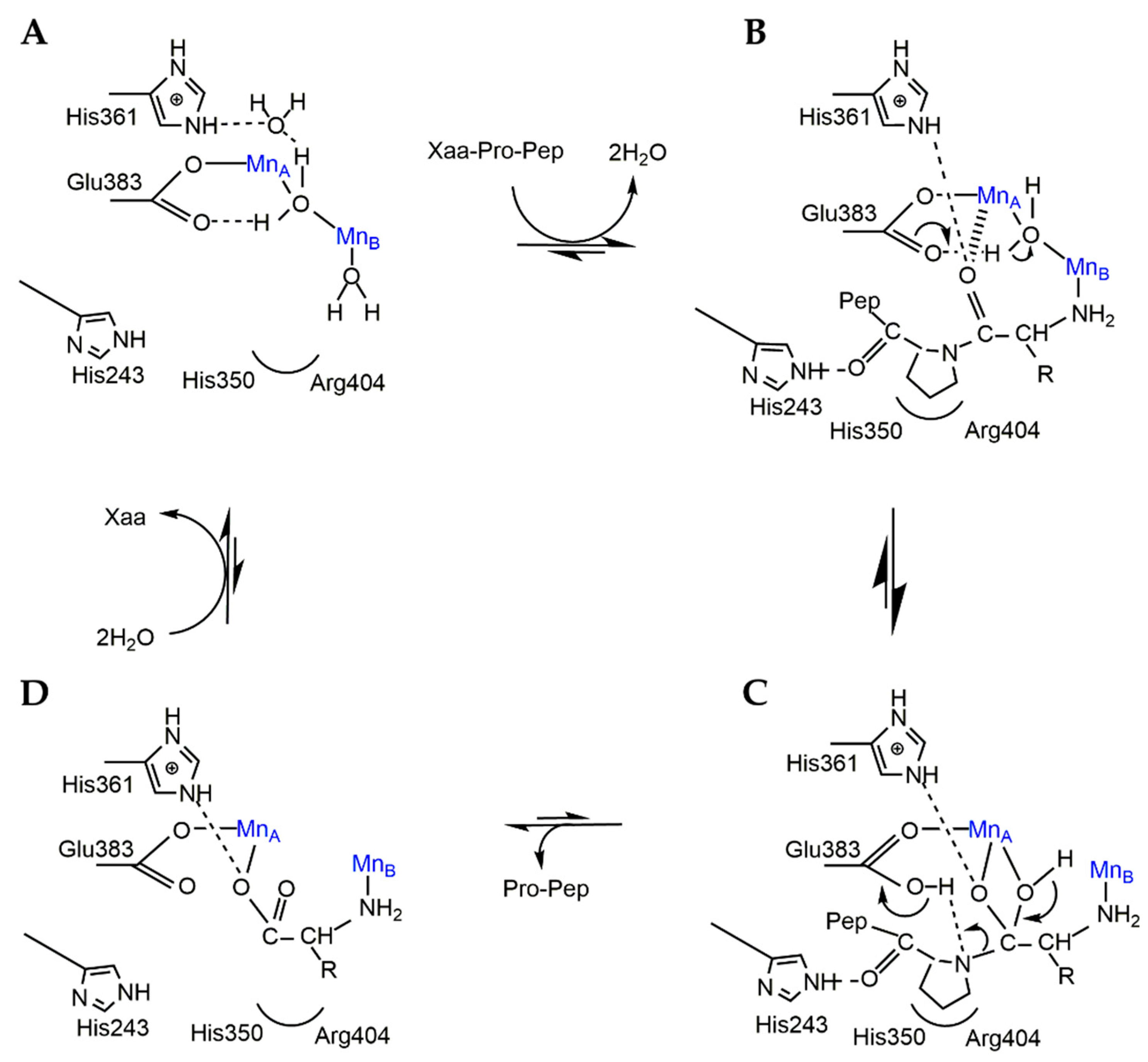
2.2. Catalytic Mechanism of APPro
3. Industrial Application of Bacterial APPro
3.1. Cheese Manufacturing Industry
3.2. Enzyme Therapy
3.3. Bioremediation and Decontamination
4. Conclusions and Future Perspectives
5. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef] [PubMed]
- Nandan, A.; Nampoothiri, K.M. Therapeutic and biotechnological applications of substrate specific microbial aminopeptidases. Appl. Microbiol. Biotechnol. 2020, 104, 5243–5257. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A. Aminopeptidase: Structure and function. FASEB J. 1993, 7, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A. Aminopeptidases: Towards a mechanism of action. Trends Biochem. Sci. 1993, 18, 167–171. [Google Scholar]
- Gonzales, T.; Robert-Baudouy, J. Bacterial aminopeptidases: Properties and functions. FEMS Microbiol. 1996, 18, 319–344. [Google Scholar] [CrossRef]
- Kunji, E.R.S.; Mierau, I.; Hagting, A.; Poolman, B.; Konings, W.N. The proteolytic systems of lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 187–221. [Google Scholar] [CrossRef]
- Christensen, J.E.; Dudley, E.G.; Pederson, J.A.; Steel, J.L. Peptidases and amino acid catabolism in lactic acid bacteria. Antonie Van Leeuwenhoek 1999, 76, 217–427. [Google Scholar] [CrossRef]
- Woessner, J.; Barret, A.; Rawlings, N. Handbook of Proteolytic Enzymes; Elsevier Academic Press: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Sanz, Y. Aminopeptidases. In Industrial Enzymes; Springer: Dordrecht, The Netherlands, 2007; pp. 243–260. [Google Scholar]
- Rahulan, R.; Manjunath, G.; Nampoothiri, K.M.; Pandey, A. Extracellular Methionine Amino peptidase ( MAP ) Production by Streptomyces gedanensis in Solid-State Fermentation. Braz. Arch. Biol. Technol. 2014, 57, 187–193. [Google Scholar] [CrossRef]
- Sanz, Y.; Toldrá, F. Purification and Characterization of an Arginine Aminopeptidase from Lactobacillus sakei. Appl. Environ. Microbiol. 2002, 68, 1980–1987. [Google Scholar] [CrossRef]
- Nandan, A.S.; Nampoothiri, K.M. Unveiling aminopeptidase P from Streptomyces lavendulae: Molecular cloning, expression and biochemical characterization. Enzyme Microb. Technol. 2014, 55, 7–13. [Google Scholar] [CrossRef]
- Niven, W.G. Purification and characterization of aminopeptidase A from Lactococcus lactis subs.p. lactis NCDO 712. J. Gen. Microbiol. 1991, 137, 1207–1212. [Google Scholar] [CrossRef]
- Nampoothiri, K.M.; Nagy, V.; Kovacs, K.; Szakacs, G.; Pandey, A. L-leucine aminopeptidase production by filamentous Aspergillus fungi. Lett. Appl. Microbiol. 2005, 41, 498–504. [Google Scholar] [CrossRef]
- Wilce, M.C.J.; Bond, C.S.; Dixon, N.E.; Freeman, H.C.; Guss, J.M.; Lilley, P.E.; Wilce, J.A. Structure and mechanism of a proline-specific aminopeptidase from Escherichia coli. Proc. Natl. Acad. Sci. USA 1998, 95, 3472–3477. [Google Scholar] [CrossRef]
- Graham, S.C.; Lee, M.; Freeman, H.C.; Guss, J.M. An orthorhombic form of Escherichia coli aminopeptidase P at 2.4 Å resolution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2003, 59, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Yaron, A.; Mlynar, D. Aminopeptidase P. Biochem. Phys. Res. Commun. 1968, 32, 658–663. [Google Scholar] [CrossRef]
- Peng, C.T.; Liu, L.; Li, C.C.; He, L.H.; Li, T.; Shen, Y.L.; Gao, C.; Wang, N.Y.; Xia, Y.; Zhu, Y.B.; et al. Structure-function relationship of aminopeptidase P from Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Are, V.N.; Kumar, A.; Goyal, V.D.; Gotad, S.S.; Ghosh, B.; Gadre, R.; Jamdar, S.N.; Makde, R.D. Structures and activities of widely conserved small prokaryotic aminopeptidases-P clarify classification of M24B peptidases. Proteins Struct. Funct. Bioinforma. 2019, 87, 212–225. [Google Scholar] [CrossRef]
- Cunningham, D.F.; O’Connor, B. Proline specific peptidases. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1997, 1343, 160–186. [Google Scholar] [CrossRef]
- Phang, J.M.; Liu, W.; Zabirnyk, O. Proline metabolism and microenvironmental stress. Annu. Rev. Nutr. 2010, 30, 441–463. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Waller, M.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2014, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lowther, W.T.; Matthews, B.W. Metalloaminopeptidases: Common functional themes in disparate structural surroundings. Chem. Rev. 2002, 102, 4581–4607. [Google Scholar] [CrossRef] [PubMed]
- Feinbaum, R.L.; Urbach, J.M.; Liberati, N.T.; Djonovic, S.; Adonizio, A.; Carvunis, A.R.; Ausubel, F.M. Genome-wide identification of Pseudomonas aeruginosa virulence-related genes using a Caenorhabditis elegans infection model. PLoS Pathog. 2012, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Jao, S.C.; Huang, L.F.; Tao, Y.S.; Li, W.S. Hydrolysis of organophosphate triesters by Escherichia coli aminopeptidase P. J. Mol. Catal. B Enzym. 2004, 27, 7–12. [Google Scholar] [CrossRef]
- McDonnell, M.; Fitzgerald, R.; Ni Fhaoláin, I.; Jennings, P.V.; O’Cuinn, G. Purification and characterization of aminopeptidase P from Lactococcus lactis subsp. cremoris. J. Dairy Res. 1997, 64, 399–407. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A.J.; Johnson, A.P.; Vemulapalli, S.; Kuehn, M.J. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J. Bacteriol. 2006, 188, 5385–5392. [Google Scholar] [CrossRef]
- Vanaja, S.K.; Russo, A.J.; Behl, B.; Banerjee, I.; Yankova, M.; Deshmukh, S.D.; Rathinam, V.A.K. Bacterial Outer Membrane Vesicles Mediate Cytosolic Localization of LPS and Caspase-11 Activation. Cell 2016, 165, 1106–1119. [Google Scholar] [CrossRef]
- Graham, S.C.; Bond, C.S.; Freeman, H.C.; Guss, J.M. Structural and functional implications of metal ion selection in aminopeptidase P, a metalloprotease with a dinuclear metal center. Biochemistry 2005, 44, 13820–13836. [Google Scholar] [CrossRef]
- Graham, S.C.; Lilley, P.E.; Lee, M.; Schaeffer, P.M.; Kralicek, A.V.; Dixon, N.E.; Guss, J.M. Kinetic and crystallographic analysis of mutant Escherichia coli aminopeptidase P: Insights into substrate recognition and the mechanism of catalysis. Biochemistry 2006, 45, 964–975. [Google Scholar] [CrossRef]
- Liu, J.W.; Hadler, K.S.; Schenk, G.; Ollis, D. Using directed evolution to improve the solubility of the C-terminal domain of Escherichia coli aminopeptidase P: Implications for metal binding and protein stability. FEBS J. 2007, 274, 4742–4751. [Google Scholar] [CrossRef]
- Jeyakanthan, J.; Takada, K.; Sawano, M.; Ogasahara, K.; Mizutani, H.; Kunishima, N.; Yokoyama, S.; Yutani, K. Crystal Structural and Functional Analysis of the Putative Dipeptidase from Pyrococcus horikoshii OT3. J. Biophys. 2009, 2009, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.; Watts, T.; Li, P.; Rye, H.S. Structural basis of substrate selectivity of E. coli prolidase. PLoS ONE 2014, 9, e111531. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Bazan, J.F.; Weaver, L.H.; Roderick, S.L.; Huber, R.; Matthews, B.W. Sequence and structure comparison suggest that methionine aminopeptidase, prolidase, aminopeptidase P, and creatinase share a common fold. Proc. Natl. Acad. Sci. USA 1994, 91, 2473–2477. [Google Scholar] [CrossRef]
- Hoeffken, H.W.; Knof, S.H.; Bartlett, P.A.; Huber, R.; Moellering, H.; Schumacher, G. Crystal structure determination, refinement and molecular model of creatine amidinohydrolase from Pseudomonas putida. J. Mol. Biol. 1988, 204, 417–433. [Google Scholar] [CrossRef]
- Roderick, S.L.; Matthews, B.W. Structure of the Cobalt-Dependent Methionine Aminopeptidase from Escherichia coli: A New Type of Proteolytic Enzyme. Biochemistry 1993, 32, 3907–3912. [Google Scholar] [CrossRef]
- Coll, M.; Knof, S.H.; Ohga, Y.; Messerschmidt, A.; Huber, R.; Moellering, H.; Ru, L. Enzymatic mechanism of creatine amidinohydrolase as deduced from crystal structures. J. Mol. Biol. 1990, 214, 597–610. [Google Scholar] [CrossRef]
- Yaron, A.; Naider, F.; Scharpe, S. Proline-dependent structural and biological properties of peptides and proteins. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 31–81. [Google Scholar] [CrossRef]
- Cottrell, G.S.; Hooper, N.M.; Turner, A.J. Cloning, expression, and characterization of human cytosolic aminopeptidase P: A single manganese(II)-dependent enzyme. Biochemistry 2000, 39, 15121–15128. [Google Scholar] [CrossRef]
- Drinkwater, N.; Sivaraman, K.K.; Bamert, R.S.; Rut, W.; Mohamed, K.; Vinh, N.B.; Scammells, P.J.; Drag, M.; McGowan, S. Structure and substrate fingerprint of aminopeptidase P from Plasmodium falciparum. Biochem. J. 2016, 473, 3189–3204. [Google Scholar] [CrossRef] [PubMed]
- Arreola, R.; Villalpando, J.L.; Puente-Rivera, J.; Morales-Montor, J.; Rudiño-Piñera, E.; Alvarez-Sánchez, M.E. Trichomonas vaginalis metalloproteinase TvMP50 is a monomeric Aminopeptidase P-like enzyme. Mol. Biotechnol. 2018, 60, 563–575. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Murayama, N.; Honda, T.; Tone, H.; Tsuru, D. Cloning and expression of aminopeptidase P gene from Escherichia coli HB101 and characterization of expressed enzyme. J. Biochem. 1988, 104, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Buchanan, T.M. Hydrolases from Neisseria gonorrhoeae. The study of gonocosin, an aminopeptidase-P, a proline iminopeptidase, and an asparaginase. J. Biol. Chem. 1980, 255, 1704–1710. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, Y.J.; Bae, S.S.; Jeon, J.H.; Lim, J.K.; Kang, S.G.; Lee, J.H. Overexpression and characterization of a carboxypeptidase from the hyperthermophilic archaeon Thermococcus sp. NA1. Biosci. Biotechnol. Biochem. 2006, 70, 1140–1147. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mars, I.; Monnet, V. An aminopeptidase P from Lactococcus lactis with original specificity. BBA Gen. Subj. 1995, 1243, 209–215. [Google Scholar] [CrossRef]
- Li, X.; Lou, Z.; Li, X.; Zhou, W.; Ma, M.; Cao, Y.; Geng, Y.; Bartlam, M.; Zhang, X.C.; Rao, Z. Structure of human cytosolic X-prolyl aminopeptidase: A double Mn(II)-dependent dimeric enzyme with a novel three-domain subunit. J. Biol. Chem. 2008, 283, 22858–22866. [Google Scholar] [CrossRef]
- Iyer, S.; La-Borde, P.J.; Payne, K.A.P.; Parsons, M.R.; Turner, A.J.; Isaac, R.E.; Acharya, K.R. Crystal structure of X-prolyl aminopeptidase from Caenorhabditis elegans: A cytosolic enzyme with a di-nuclear active site. FEBS Open Bio 2015, 5, 292–302. [Google Scholar] [CrossRef]
- Are, V.N.; Kumar, A.; Kumar, S.; Goyal, V.D.; Ghosh, B.; Bhatnagar, D.; Jamdar, S.N.; Makde, R.D. Crystal structure and biochemical investigations reveal novel mode of substrate selectivity and illuminate substrate inhibition and allostericity in a subfamily of Xaa-Pro dipeptidases. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 153–164. [Google Scholar] [CrossRef]
- Graham, S.C.; Maher, M.J.; Simmons, W.H.; Freeman, H.C.; Guss, J.M. Structure of Escherichia coli aminopeptidase P in complex with the inhibitor apstatin. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 1770–1779. [Google Scholar] [CrossRef]
- Zhang, L.; Crossley, M.J.; Dixon, N.E.; Ellis, P.J.; Fisher, M.L.; King, G.F.; Lilley, P.E.; MacLachlan, D.; Pace, R.J.; Freeman, H.C. Spectroscopic identification of a dinuclear metal centre in manganese(II)-activated aminopeptidase P from Escherichia coli: Implications for human prolidase. J. Biol. Inorg. Chem. 1998, 3, 470–483. [Google Scholar] [CrossRef]
- Hu, X.V.; Chen, X.; Kee, C.H.; Mildvan, A.S.; Liu, J.O. Kinetic and mutational studies of the number of interacting divalent cations required by bacterial and human methionine aminopeptidases. Biochemistry 2007, 46, 12833–12843. [Google Scholar] [CrossRef]
- Graham, S.C.; Guss, J.M. Complexes of mutants of Escherichia coli aminopeptidase P and the tripeptide substrate ValProLeu. Arch. Biochem. Biophys. 2008, 469, 200–208. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera - A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Zheng, Y.; Roberts, R.J.; Kasif, S.; Guan, C. Characterization of two new aminopeptidases in Escherichia coli. J. Bacteriol. 2005, 187, 3671–3677. [Google Scholar] [CrossRef] [PubMed]
- Stressler, T.; Eisele, T.; Schlayer, M.; Fischer, L. Production, active staining and gas chromatography assay analysis of recombinant aminopeptidase P from Lactococcus lactis ssp. lactis DSM 20481. AMB Express 2012, 2, 1–12. [Google Scholar] [CrossRef]
- Simmons, W.H. Aminopeptidase P2. In Handbook of Proteolytic Enzyme; Elsevier: Amsterdam, The Netherlands, 2012; pp. 1528–1532. [Google Scholar]
- Hooper, N.M.; Hryszko, J.; Oppong, S.Y.; Turner, A.J. Inhibition by converting enzyme inhibitors of pig kidney aminopeptidase P. Hypertension 1992, 19, 281–285. [Google Scholar] [CrossRef][Green Version]
- Copik, A.J.; Swierczek, S.I.; Lowther, W.T.; D’Souza, V.M.; Matthews, B.W.; Holz, R.C. Kinetic and spectroscopic characterization of the H178A methionyl aminopeptidase from Escherichia coli. Biochemistry 2003, 42, 6283–6292. [Google Scholar] [CrossRef] [PubMed]
- Copik, A.J.; Nocek, B.P.; Swierczek, S.I.; Ruebush, S.; Se, B.J.; Meng, L.; D’Souza, V.M.; Peters, J.W.; Bennett, B.; Holz, R.C. EPR and X-ray crystallographic characterization of the product-bound form of the MnII-loaded methionyl aminopeptidase from Pyrococcus furiosus. Biochemistry 2005, 44, 121–129. [Google Scholar] [CrossRef]
- Li, Z.; Wan, H.; Shi, Y.; Ouyang, P. Personal experience with four kinds of chemical structure drawing software: Review on chemdraw, chemwindow, ISIS/draw, and chemsketch. J. Chem. Inf. Comput. Sci. 2004, 44, 1886–1890. [Google Scholar] [CrossRef] [PubMed]
- Jao, S.C.; Huang, L.F.; Hwang, S.M.; Li, W.S. Tyrosine 387 and arginine 404 are critical in the hydrolytic mechanism of Escherichia coli aminopeptidase P. Biochemistry 2006, 45, 1547–1553. [Google Scholar] [CrossRef]
- Nandan, A.; Nampoothiri, K.M. Biochemical and structural analysis of a site directed mutant of manganese dependent aminopeptidase P from Streptomyces lavendulae. J. Biosci. Biotechnol. 2015, 4, 109–116. [Google Scholar]
- Doi, E.; Shibata, D.; Matoba, T. Modified colorimetric ninhydrin methods for peptidase assay. Anal. Biochem. 1981, 118, 173–184. [Google Scholar] [CrossRef]
- Lasch, J.; Moschner, S.; Sann, H.; Zellmer, S.; Koelsch, R. Aminopeptidase P--a cell-surface antigen of endothelial and lymphoid cells: Catalytic and immuno-histotopical. Biol. Chem. 1998, 379, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.T.; Su, C.Y.; Du, H.C.; Jao, S.C.; Li, W.S. Evaluation of organophosphorus chemicals-degrading enzymes: A comparison of Escherichia coli and human cytosolic aminopeptidase P. Chem. Biodivers. 2008, 5, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Fleminger, G.; Carmel, A.; Goldenberg, D.; Yaron, A. Fluorogenic Substrates for Bacterial Aminopeptidase P and Its Analogs Detected in Human Serum and Calf Lung. Eur. J. Biochem. 1982, 125, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Holtzman, E.J.; Pillay, G.; Rosenthal, T.; Yaron, A. Aminopeptidase P activity in rat organs and human serum. Anal. Biochem. 1987, 162, 476–484. [Google Scholar] [CrossRef]
- Hawthorne, S.J.; Harriott, P.; Lim, J.; Turner, A.J.; Walker, B.; Williams, C.H. Evaluation of some fluorogenic substrates for continuous assay of aminopeptidase P. Anal. Biochem. 1997, 253, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Stöckel-Maschek, A.; Stiebitz, B.; Koelsch, R.; Neubert, K. A continuous fluorimetric assay for aminopeptidase P detailed analysis of product inhibition. Anal. Biochem. 2003, 322, 60–67. [Google Scholar] [CrossRef]
- Visser, S. Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview. J. Dairy Sci. 1993, 76, 329–350. [Google Scholar] [CrossRef]
- Gatti, M.; Fornasari, M.E.; Mucchetti, G.; Addeo, F.; Neviani, E. Presence of peptidase activities in different varieties of cheese. Lett. Appl. Microbiol. 1999, 28, 368–372. [Google Scholar] [CrossRef]
- Martins, J.M.; Galinari, É.; Pimentel-Filho, N.J.; Ribeiro, J.I.; Furtado, M.M.; Ferreira, C.L.L.F. Determining the minimum ripening time of artisanal Minas cheese, a traditional Brazilian cheese. Braz. J. Microbiol. 2015, 46, 219–230. [Google Scholar] [CrossRef]
- Turner, A.J.; Hyde, R.J.; Lim, J.; Hooper, N.M. Structural Studies of Aminopeptidase P. In Cellular Peptidases in Immune Functions and Diseases; Springer: Berlin/Heidelberg, Germany, 1997; pp. 7–16. [Google Scholar]
- Meller, S.T.; Gebhart, G.F. A critical review of the afferent pathways and the potential chemical mediators involved in cardiac pain. Neuroscience 1992, 48, 501–524. [Google Scholar] [CrossRef]
- Kidd, B.L.; Urban, L.A. Mechanism of inflammatory. Br. J. Anaesth. 2001, 87, 3–11. [Google Scholar] [CrossRef]
- Lind, L.; Granstam, S.O.; Millgård, J. Endothelium-dependent vasodilation in hypertension: A review. Blood Press. 2000, 9, 4–15. [Google Scholar] [CrossRef]
- Gavras, I. Bradykinin-mediated effects of ACE inhibition. Kidney Int. 1992, 42, 1020–1029. [Google Scholar] [CrossRef]
- Ceconi, C.; Francolini, G.; Bastianon, D.; Gitti, G.L.; Comini, L.; Ferrari, R. Differences in the effect of angiotensin-converting enzyme inhibitors on the rate of endothelial cell apoptosis: In vitro and in vivo studies. Cardiovasc. Drugs Ther. 2007, 21, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Van Vark, L.C.; Bertrand, M.; Akkerhuis, K.M.; Brugts, J.J.; Fox, K.; Mourad, J.J.; Boersma, E. Angiotensin-converting enzyme inhibitors reduce mortality in hypertension: A meta-analysis of randomized clinical trials of renin-angiotensin-aldosterone system inhibitors involving 158 998 patients. Eur. Heart J. 2012, 33, 2088–2097. [Google Scholar] [CrossRef]
- Karlberg, B.E. Cough and inhibition of the renin-angiotensin system. J. Hypertens. Suppl. 1993, 11, 3–11. [Google Scholar]
- Fox, A.J.; Lalloo, U.G.; Belvisi, M.G.; Bernareggi, M.; Chung, K.F.; Barnes, P.J. Bradykinin-evoked sensitization of airway sensory nerves: A mechanism for ACE-inhibitor cough. Nat. Med. 1996, 2, 814–817. [Google Scholar] [CrossRef]
- Bas, M.; Adams, V.; Suvorava, T.; Niehues, T.; Hoffmann, T.K.; Kojda, G. Nonallergic angioedema: Role of bradykinin. Allergy Eur. J. Allergy Clin. Immunol. 2007, 62, 842–856. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.A.; Perego, F.; Zanichelli, A.; Cicardi, M. Angioedema Phenotypes: Disease Expression and Classification. Clin. Rev. Allergy Immunol. 2016, 51, 162–169. [Google Scholar] [CrossRef]
- Habif, T.P. Clinical Dermatolgy E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Bernstein, J.A.; Cremonesi, P.; Hoffmann, T.K.; Hollingsworth, J. Angioedema in the emergency department: A practical guide to differential diagnosis and management. Int. J. Emerg. Med. 2017, 10, 15. [Google Scholar] [CrossRef]
- Kuoppala, A.; Lindstedt, K.A.; Saarinen, J.; Kovanen, P.T.; Kokkonen, J.O. Inactivation of bradykinin by angiotensin-converting enzyme and by carboxypeptidase N in human plasma. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Dendorfer, A.; Wolfrum, S.; Wagemann, M.; Qadri, F.; Dominiak, P. Pathways of bradykinin degradation in blood and plasma of normotensive and hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, 2182–2188. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.C.; Defrank, J.J. Hydrolysis of Organophosphorus Compounds by Bacterial Prolidases; Springer: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Singh, B.K.; Walker, A. Microbial degradation of organophosphorus compounds. FEMS Microbiol. Rev. 2006, 30, 428–471. [Google Scholar] [CrossRef]
- Singh, B.K. Organophosphorus-degrading bacteria: Ecology and industrial applications. Nat. Rev. Microbiol. 2009, 7, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.B.; Sutherland, T.D.; Gresham, C.; Oakeshott, J.; Scott, C.; Eddleston, M. OpdA, a bacterial organophosphorus hydrolase, prevents lethality in rats after poisoning with highly toxic organophosphorus pesticides. Toxicology 2008, 247, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Jeyaratnam, J. Acute pesticide poisoning: A major global health problem. World Health Stat Q. 1990, 43, 139–144. [Google Scholar]
- LeJeune, K.E.; Wild, J.R.; Russel, A.J. Nerve agents degraded by enzymatic foams. Nature 1998, 395, 27–28. [Google Scholar] [CrossRef]
- Chen, W.; Richins, R.D.; Mulchandani, P.; Kaneva, I.; Mulchandani, A. Biodgradation of organophosphorus nerve agents by surface expressed organophosphorus hydrolase. In Enzymes in Action; Springer: Dordrecht, The Netherlands, 2000; pp. 211–221. [Google Scholar]
- DeFrank, J.J.; White, W.E. Phosphofluoridates: Biological Activity and Biodegradation. Organofluorines 2006, 3, 295–343. [Google Scholar] [CrossRef]
- Theriot, C.M.; Grunden, A.M. Hydrolysis of organophosphorus compounds by microbial enzymes. Appl. Microbiol. Biotechnol. 2011, 89, 35–43. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sources | PDB ID | Resolution (Å) | Metal Ions (MAMB) | Inhibitor | Substrate | Crystal Assembly | References |
|---|---|---|---|---|---|---|---|
| Escherichia coli | 1AZ9 | 2.0 | MnMn | - | - | Tetramer | [15] |
| 1A16 | 2.3 | MnMn | ProLeu | - | |||
| IJAW (low pH) | 2.7 | MnMn | - | - | |||
| 1N51 | 2.3 | MnMn | Apstatin | - | Tetramer | [51] | |
| 1WL9 | 1.9 | MnMn | - | - | Tetramer | [30] | |
| 1WLR | 2.1 | - | - | - | |||
| 1WL6 | 2.0 | MgMg | - | - | |||
| 1W2M | 2.4 | Ca- | - | - | |||
| 2BHC | 2.4 | Na- | - | - | |||
| 2BHA | 2.4 | - | - | ValProLeu | |||
| 2BHD | 2.5 | MgMg | - | ProLeu 1 | |||
| 2BHB | 2.4 | ZnZn | - | - | |||
| No crystal structure | 2.8 | ZnZn, Zn 2 | - | ProLeu 1 | |||
| 2BH3 | 2.4 | ZnZn, Zn 2 | ProLeu | - | |||
| 2BN7 | 2.4 | MnMn, Zn 2 | ProLeu | - | |||
| 1WBQ | 2.3 | ZnMg | - | - | |||
| 1W7V | 2.0 | ZnMg | - | ProLeu 1 | |||
| 5CNX | 2.6 | ZnZn | - | - | Dimer | [19] | |
| Deinococcus radiodurans | 5G1Q | 1.8 | - | - | |||
| Pseudomonas aeruginosa | 5WZE | 2.0 | MnMn | - | - | Tetramer | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omar, M.N.; Raja Abd Rahman, R.N.Z.; Muhd Noor, N.D.; Latip, W.; Knight, V.F.; Ali, M.S.M. Structure-Function and Industrial Relevance of Bacterial Aminopeptidase P. Catalysts 2021, 11, 1157. https://doi.org/10.3390/catal11101157
Omar MN, Raja Abd Rahman RNZ, Muhd Noor ND, Latip W, Knight VF, Ali MSM. Structure-Function and Industrial Relevance of Bacterial Aminopeptidase P. Catalysts. 2021; 11(10):1157. https://doi.org/10.3390/catal11101157
Chicago/Turabian StyleOmar, Muhamad Nadzmi, Raja Noor Zaliha Raja Abd Rahman, Noor Dina Muhd Noor, Wahhida Latip, Victor Feizal Knight, and Mohd Shukuri Mohamad Ali. 2021. "Structure-Function and Industrial Relevance of Bacterial Aminopeptidase P" Catalysts 11, no. 10: 1157. https://doi.org/10.3390/catal11101157
APA StyleOmar, M. N., Raja Abd Rahman, R. N. Z., Muhd Noor, N. D., Latip, W., Knight, V. F., & Ali, M. S. M. (2021). Structure-Function and Industrial Relevance of Bacterial Aminopeptidase P. Catalysts, 11(10), 1157. https://doi.org/10.3390/catal11101157