
Circulating Nucleosomes and Nucleosome Modifications as Biomarkers in Cancer



Abstract
:1. Introduction
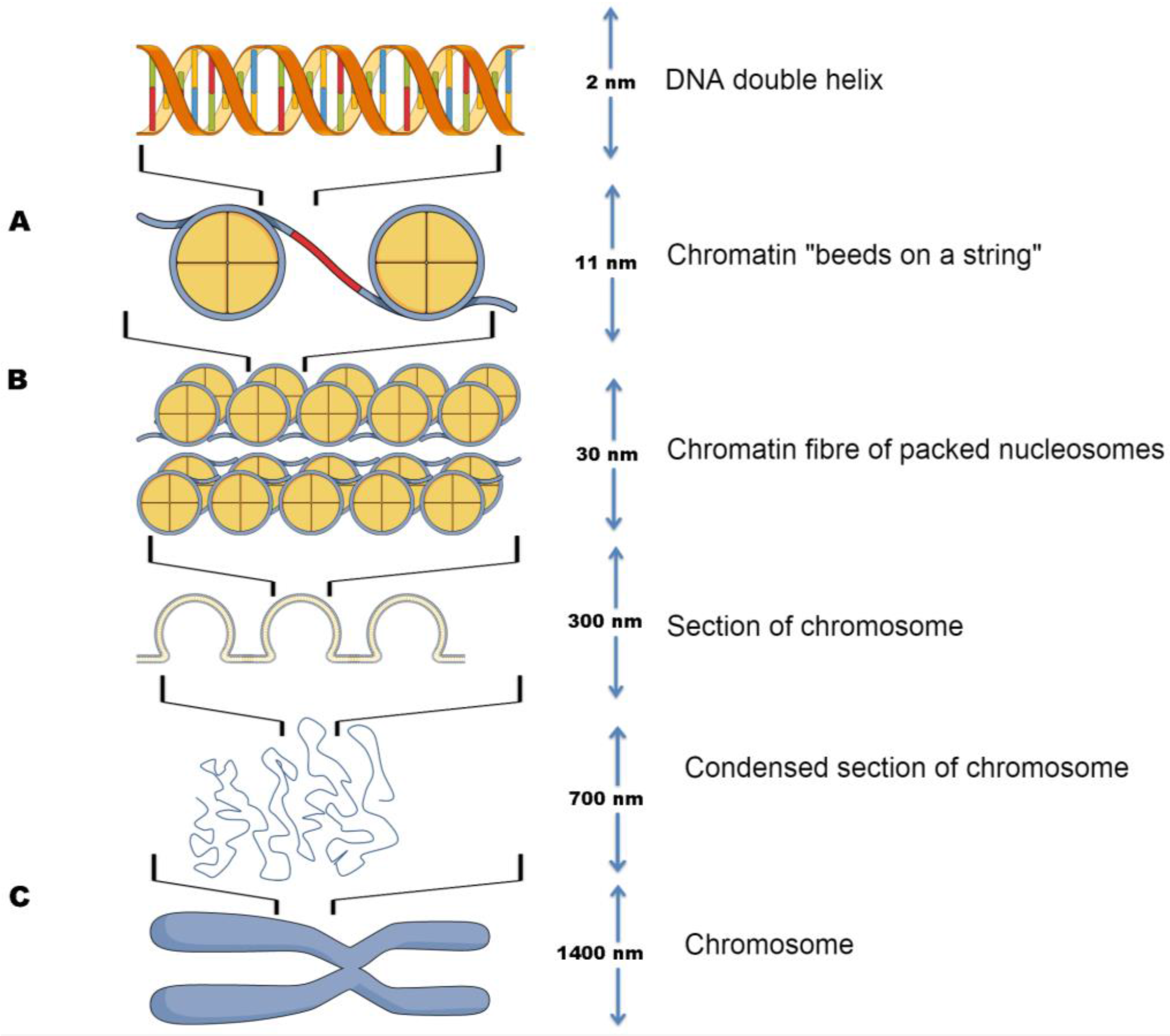
2. The Role of Nucleosomes in Packaging DNA
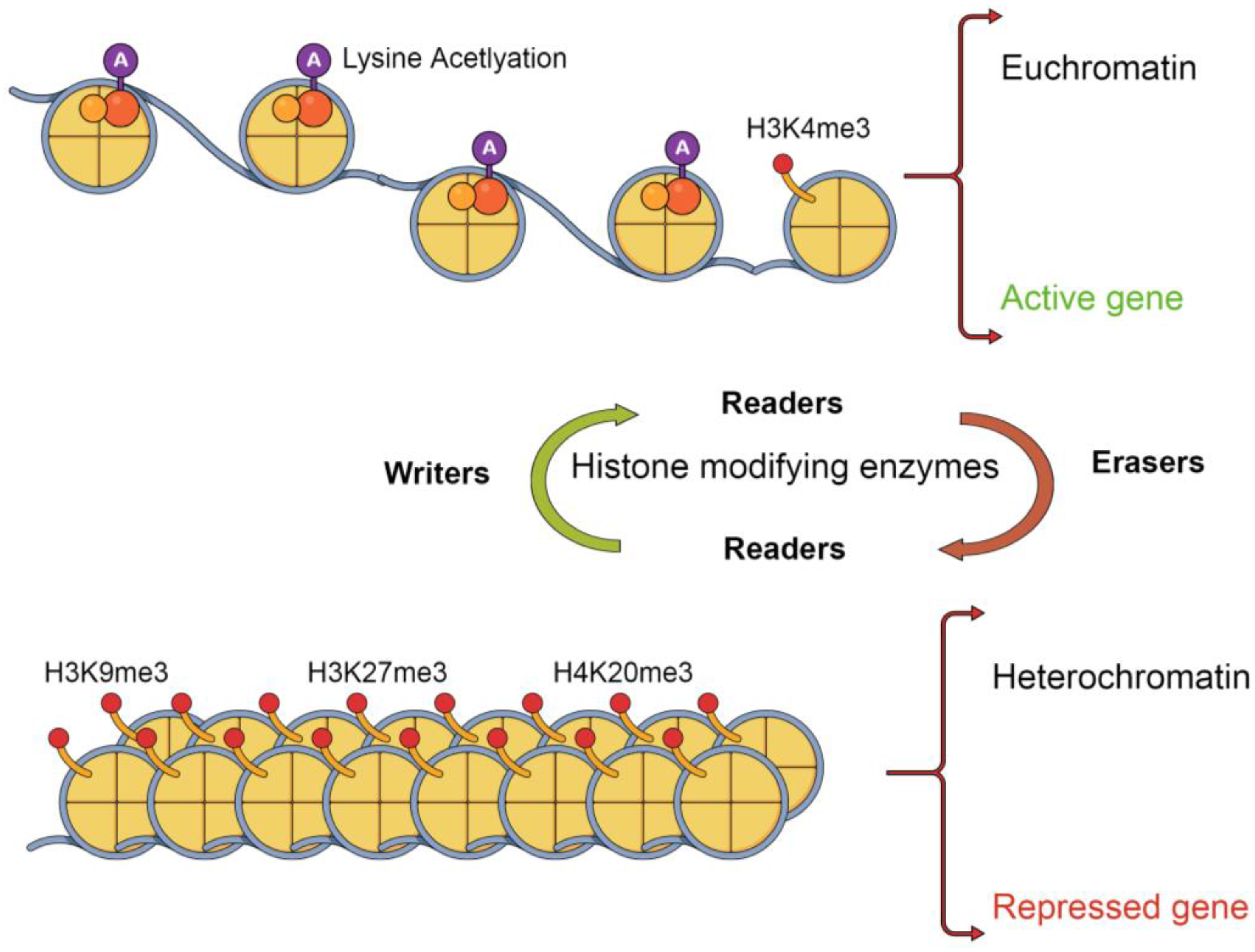
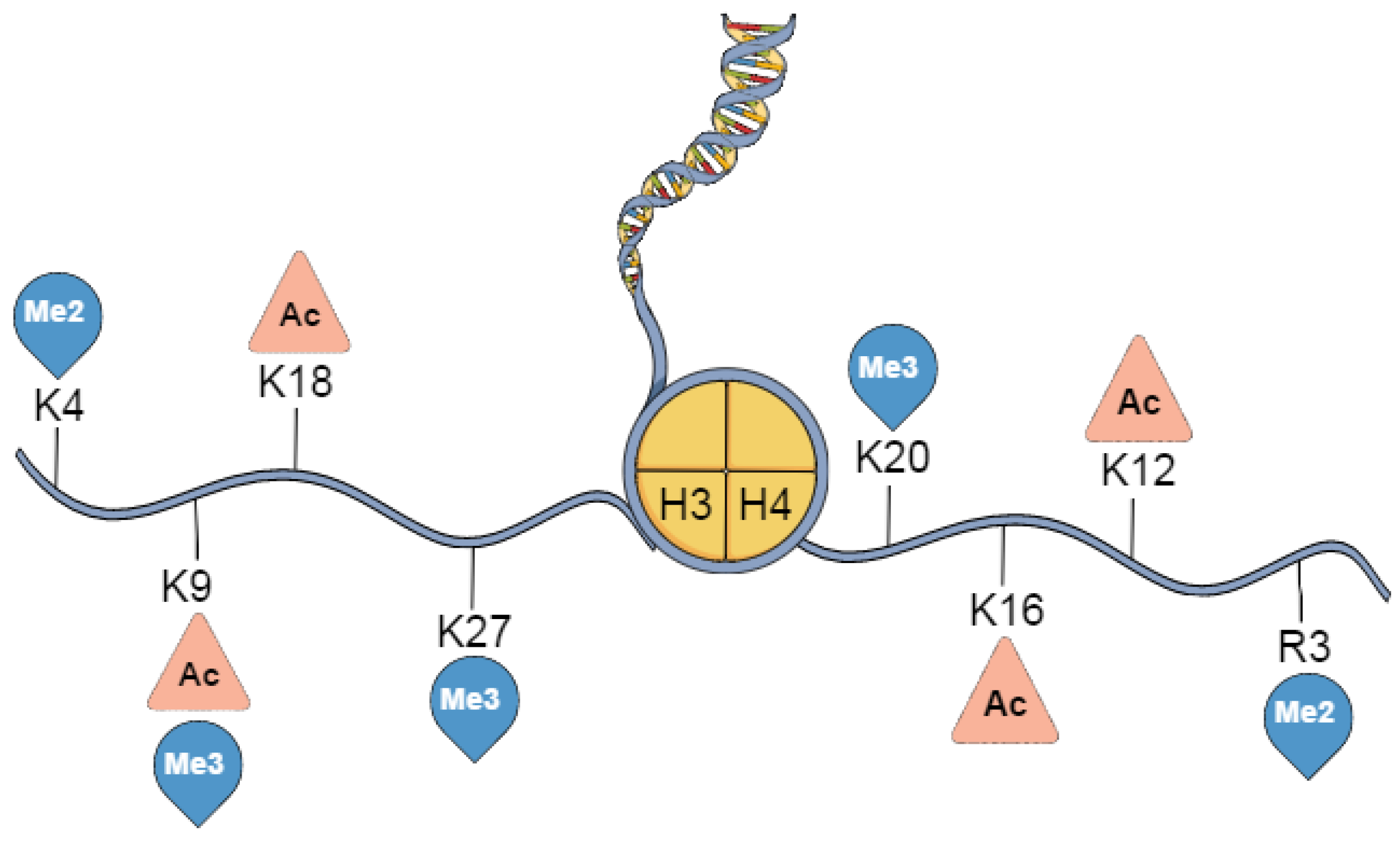
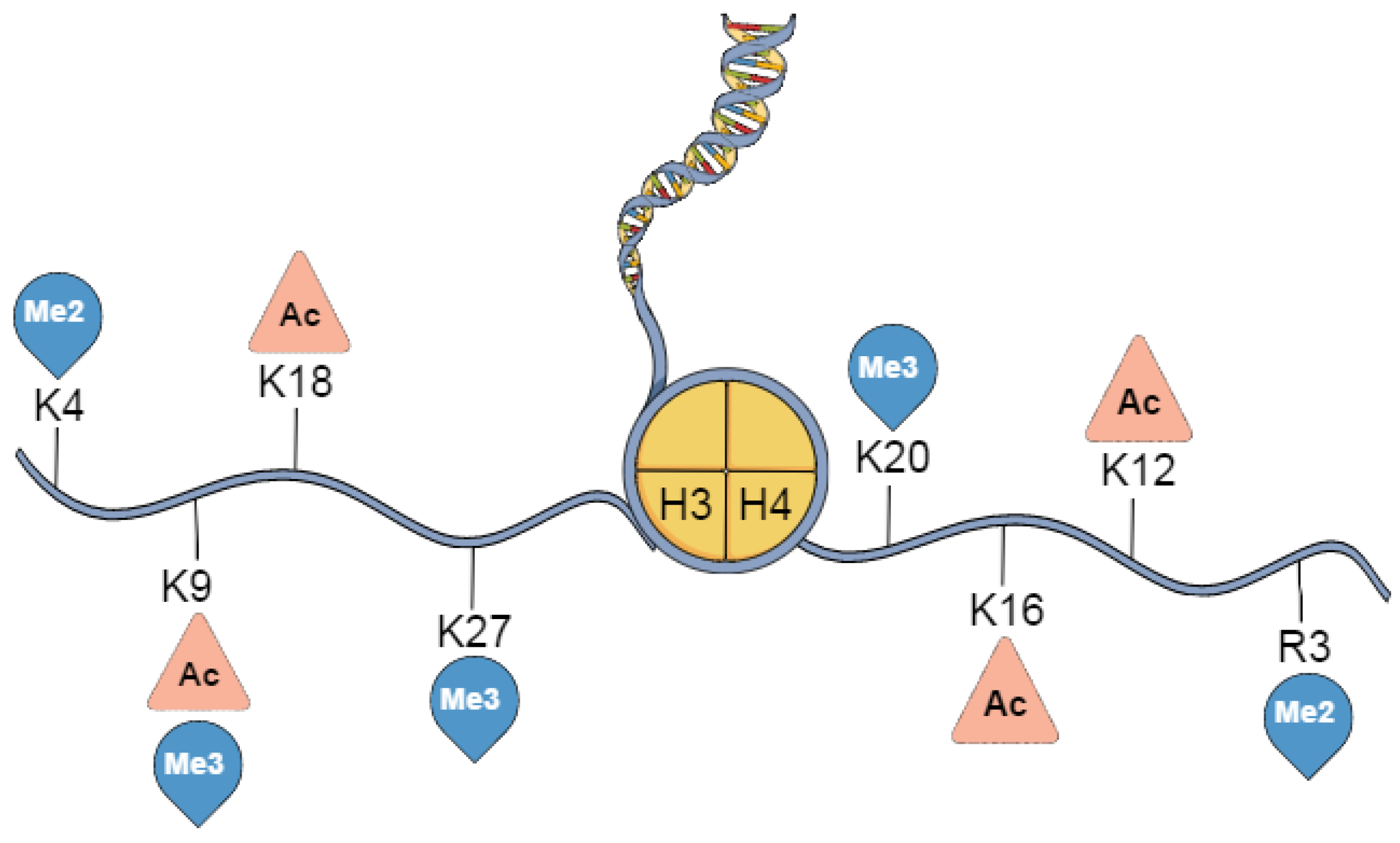
3. Histone Post-Translational Modifications
4. Quantification of Nucleosomes PTMs as Tumour Biomarkers
4.1. Breast Cancer
4.2. Esophageal Cancer
4.3. Gastric Cancer
4.4. Colorectal Cancer
5. Histone Variants
6. Circulating Nucleosomes as Cancer Biomarkers
7. Quantifying PTM of Circulating Nucleosomes as Biomarkers
8. MicroRNA Regulation of Histone PTM
9. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2012, 62, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Negrini, S.; Gorgoulis, V.G.; Halazonetis, T.D. Genomic instability—An evolving hallmark of cancer. Nat. Rev. Mol. Cell. Biol. 2010, 11, 220–228. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Dictionary—Definition of Biomarker. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms?cdrid=45618 (accessed on 24 October 2016).
- Kornberg, R.D. Chromatin structure: A repeating unit of histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.L.; Laniel, M.A. Histones and histone modifications. Curr. Biol. 2004, 14, R546–R551. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yang, F.; Shen, W.H. Genome maintenance in the context of 4D chromatin condensation. Cell. Mol. Life Sci. 2016, 73, 3137–3150. [Google Scholar] [CrossRef] [PubMed]
- Campos, E.I.; Reinberg, D. Histones: Annotating chromatin. Annu. Rev. Genet. 2009, 43, 559–599. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Grosschedl, R. Dynamics and interplay of nuclear architecture, genome organization, and gene expression. Genes Dev. 2007, 21, 3027–3043. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D.; Lorch, Y. Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell 1999, 98, 285–294. [Google Scholar] [CrossRef]
- Khorasanizadeh, S. The nucleosome: From genomic organization to genomic regulation. Cell 2004, 116, 259–272. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Taverna, S.D.; Li, H.; Ruthenburg, A.J.; Allis, C.D.; Patel, D.J. How chromatin-binding modules interpret histone modifications: Lessons from professional pocket pickers. Nat. Struct. Mol. Biol. 2007, 14, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Shogren-Knaak, M.; Ishii, H.; Sun, J.M.; Pazin, M.J.; Davie, J.R.; Peterson, C.L. Histone H4-K16 acetylation controls chromatin structure and protein interactions. Science 2006, 311, 844–847. [Google Scholar] [CrossRef] [PubMed]
- Luger, K.; Dechassa, M.L.; Tremethick, D.J. New insights into nucleosome and chromatin structure: An ordered state or a disordered affair? Nat. Rev. Mol. Cell Biol. 2012, 13, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Zlatanova, J.; Bishop, T.C.; Victor, J.-M.; Jackson, V.; van Holde, K. The nucleosome family: Dynamic and growing. Structure 2009, 17, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Andrews, A.J.; Luger, K. Nucleosome structure(s) and stability: Variations on a theme. Annu. Rev. Biophys. 2011, 40, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Black, B.E.; Bassett, E.A. The histone variant CENP-A and centromere specification. Curr. Opin. Cell Biol. 2008, 20, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Nishibuchi, I.; Suzuki, H.; Kinomura, A.; Sun, J.; Liu, N.A.; Horikoshi, Y.; Shima, H.; Kusakabe, M.; Harata, M.; Fukagawa, T.; et al. Reorganization of damaged chromatin by the exchange of histone variant H2A.Z-2. Int. J. Radiat. Oncol. Biol. Phys. 2014, 89, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ayrapetov Marina, K.; Xu, C.; Gursoy-Yuzugullu, O.; Hu, Y.; Price Brendan, D. Histone H2A.Z controls a critical chromatin remodeling step required for DNA double-strand break repair. Mol. Cell 2012, 48, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Allfrey, V.G.; Faulkner, R.; Mirsky, A.E. Acetylation and methylation of histones and their possible role in the regulation of RNA synthesis. Proc. Natl. Acad. Sci. USA 1964, 51, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Luo, H.; Lee, S.; Jin, F.; Yang, J.S.; Montellier, E.; Buchou, T.; Cheng, Z.; Rousseaux, S.; Rajagopal, N.; et al. Identification of 67 histone marks and histone lysine crotonylation as a new type of histone modification. Cell 2011, 146, 1016–1028. [Google Scholar] [CrossRef] [PubMed]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.; Wu, J.; Workman, J.L.; Li, B. Readers of histone modifications. Cell Res. 2011, 21, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Fradet-Turcotte, A.; Canny, M.D.; Escribano-Diaz, C.; Orthwein, A.; Leung, C.C.; Huang, H.; Landry, M.C.; Kitevski-LeBlanc, J.; Noordermeer, S.M.; Sicheri, F.; et al. 53BP1 is a reader of the DNA-damage-induced H2A lys 15 ubiquitin mark. Nature 2013, 499, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Ikura, T.; Tashiro, S.; Kakino, A.; Shima, H.; Jacob, N.; Amunugama, R.; Yoder, K.; Izumi, S.; Kuraoka, I.; Tanaka, K.; et al. DNA damage-dependent acetylation and ubiquitination of H2AX enhances chromatin dynamics. Mol. Cell. Biol. 2007, 27, 7028–7040. [Google Scholar] [CrossRef] [PubMed]
- Ikura, M.; Furuya, K.; Matsuda, S.; Matsuda, R.; Shima, H.; Adachi, J.; Matsuda, T.; Shiraki, T.; Ikura, T. Acetylation of histone H2AX at lys 5 by the tip60 histone acetyltransferase complex is essential for the dynamic binding of NBS1 to damaged chromatin. Mol. Cell. Biol. 2015, 35, 4147–4157. [Google Scholar] [CrossRef] [PubMed]
- Cang, S.; Xu, X.; Ma, Y.; Liu, D.; Chiao, J.W. Hypoacetylation, hypomethylation, and dephosphorylation of H2B histones and excessive histone deacetylase activity in DU-145 prostate cancer cells. J. Hematol. Oncol. 2016, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Q.; Wu, M.Z.; Wu, K.J. Analysis of epigenetic regulation of hypoxia-induced epithelial-mesenchymal transition in cancer cells by quantitative chromatin immunoprecipitation of histone deacetylase 3 (HDAC3). Methods Mol. Biol. 2016, 1436, 23–29. [Google Scholar] [PubMed]
- Ellinger, J.; Kahl, P.; von der Gathen, J.; Rogenhofer, S.; Heukamp, L.C.; Gutgemann, I.; Walter, B.; Hofstadter, F.; Buttner, R.; Muller, S.C.; et al. Global levels of histone modifications predict prostate cancer recurrence. Prostate 2010, 70, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Elsheikh, S.E.; Green, A.R.; Rakha, E.A.; Powe, D.G.; Ahmed, R.A.; Collins, H.M.; Soria, D.; Garibaldi, J.M.; Paish, C.E.; Ammar, A.A.; et al. Global histone modifications in breast cancer correlate with tumor phenotypes, prognostic factors, and patient outcome. Cancer Res. 2009, 69, 3802. [Google Scholar] [CrossRef] [PubMed]
- Manuyakorn, A.; Paulus, R.; Farrell, J.; Dawson, N.A.; Tze, S.; Cheung-Lau, G.; Hines, O.J.; Reber, H.; Seligson, D.B.; Horvath, S.; et al. Cellular histone modification patterns predict prognosis and treatment response in resectable pancreatic adenocarcinoma: Results from RTOG 9704. J. Clin. Oncol. 2010, 28, 1358–1365. [Google Scholar] [CrossRef] [PubMed]
- Seligson, D.B.; Horvath, S.; McBrian, M.A.; Mah, V.; Yu, H.; Tze, S.; Wang, Q.; Chia, D.; Goodglick, L.; Kurdistani, S.K. Global levels of histone modifications predict prognosis in different cancers. Am. J. Pathol. 2009, 174, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Xu, J.; Zhang, J.; Xie, D.; Ye, H.; Xiao, Z.; Cai, M.; Xu, K.; Zeng, Y.; Li, H.; et al. High expression of trimethylated histone H3 lysine 4 is associated with poor prognosis in hepatocellular carcinoma. Hum. Pathol. 2012, 43, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Song, J.S.; Kim, Y.S.; Kim, D.K.; Park, S.I.; Jang, S.J. Global histone modification pattern associated with recurrence and disease-free survival in non-small cell lung cancer patients. Pathol. Int. 2012, 62, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Rogenhofer, S.; Kahl, P.; Holzapfel, S.; Mueller, S.C.; Ellinger, J. Decreased levels of histone H3K9ME1 indicate poor prognosis in patients with renal cell carcinoma. Anticancer Res. 2012, 32, 879–886. [Google Scholar] [PubMed]
- Ellinger, J.; Bachmann, A.; Goke, F.; Behbahani, T.E.; Baumann, C.; Heukamp, L.C.; Rogenhofer, S.; Muller, S.C. Alterations of global histone H3K9 and H3K27 methylation levels in bladder cancer. Urol. Int. 2014, 93, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Gezer, U.; Ustek, D.; Yoruker, E.E.; Cakiris, A.; Abaci, N.; Leszinski, G.; Dalay, N.; Holdenrieder, S. Characterization of H3K9ME3- and H4K20ME3-associated circulating nucleosomal DNA by high-throughput sequencing in colorectal cancer. Tumour Biol. 2013, 34, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Leszinski, G.; Gezer, U.; Siegele, B.; Stoetzer, O.; Holdenrieder, S. Relevance of histone marks H3K9ME3 and H4K20ME3 in cancer. Anticancer Res. 2012, 32, 2199–2205. [Google Scholar] [PubMed]
- Park, Y.S.; Jin, M.Y.; Kim, Y.J.; Yook, J.H.; Kim, B.S.; Jang, S.J. The global histone modification pattern correlates with cancer recurrence and overall survival in gastric adenocarcinoma. Ann. Surg. Oncol. 2008, 15, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Muller-Tidow, C.; Klein, H.U.; Hascher, A.; Isken, F.; Tickenbrock, L.; Thoennissen, N.; Agrawal-Singh, S.; Tschanter, P.; Disselhoff, C.; Wang, Y.; et al. Profiling of histone H3 lysine 9 trimethylation levels predicts transcription factor activity and survival in acute myeloid leukemia. Blood 2010, 116, 3564–3571. [Google Scholar] [CrossRef] [PubMed]
- Tzao, C.; Tung, H.J.; Jin, J.S.; Sun, G.H.; Hsu, H.S.; Chen, B.H.; Yu, C.P.; Lee, S.C. Prognostic significance of global histone modifications in resected squamous cell carcinoma of the esophagus. Mod. Pathol. 2009, 22, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Ellinger, J.; Schneider, A.C.; Bachmann, A.; Kristiansen, G.; Muller, S.C.; Rogenhofer, S. Evaluation of global histone acetylation levels in bladder cancer patients. Anticancer Res. 2016, 36, 3961–3964. [Google Scholar] [PubMed]
- Seligson, D.B.; Horvath, S.; Shi, T.; Yu, H.; Tze, S.; Grunstein, M.; Kurdistani, S.K. Global histone modification patterns predict risk of prostate cancer recurrence. Nature 2005, 435, 1262–1266. [Google Scholar] [CrossRef] [PubMed]
- Karczmarski, J.; Rubel, T.; Paziewska, A.; Mikula, M.; Bujko, M.; Kober, P.; Dadlez, M.; Ostrowski, J. Histone H3 lysine 27 acetylation is altered in colon cancer. Clin. Proteom. 2014, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Rogenhofer, S.; Kahl, P.; Mertens, C.; Hauser, S.; Hartmann, W.; Buttner, R.; Muller, S.C.; von Ruecker, A.; Ellinger, J. Global histone H3 lysine 27 (H3K27) methylation levels and their prognostic relevance in renal cell carcinoma. BJU Int. 2012, 109, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Tamagawa, H.; Oshima, T.; Numata, M.; Yamamoto, N.; Shiozawa, M.; Morinaga, S.; Nakamura, Y.; Yoshihara, M.; Sakuma, Y.; Kameda, Y.; et al. Global histone modification of H3K27 correlates with the outcomes in patients with metachronous liver metastasis of colorectal cancer. Eur. J. Surg. Oncol. 2013, 39, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Xia, W.; Zhang, Z.; Liu, J.; Wang, H.; Adsay, N.V.; Albarracin, C.; Yu, D.; Abbruzzese, J.L.; Mills, G.B.; et al. Loss of trimethylation at lysine 27 of histone H3 is a predictor of poor outcome in breast, ovarian, and pancreatic cancers. Mol. Carcinog. 2008, 47, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.Y.; Hou, J.H.; Rao, H.L.; Luo, R.Z.; Li, M.; Pei, X.Q.; Lin, M.C.; Guan, X.Y.; Kung, H.F.; Zeng, Y.X.; et al. High expression of H3K27ME3 in human hepatocellular carcinomas correlates closely with vascular invasion and predicts worse prognosis in patients. Mol. Med. 2011, 17, 12–20. [Google Scholar] [PubMed]
- Benard, A.; Goossens-Beumer, I.J.; van Hoesel, A.Q.; Horati, H.; Putter, H.; Zeestraten, E.C.; van de Velde, C.J.; Kuppen, P.J. Prognostic value of polycomb proteins ezh2, bmi1 and suz12 and histone modification H3K27ME3 in colorectal cancer. PLoS ONE 2014, 9, e108265. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, N.; Matsumoto, K.; Nanashima, A.; Nagayasu, T.; Hayashi, T.; Ying, M.; Endo, D.; Wu, Z.; Koji, T. High expression of trimethylated histone H3 at lysine 27 predicts better prognosis in non-small cell lung cancer. Int. J. Oncol. 2013, 43, 1467–1480. [Google Scholar] [PubMed]
- Chen, Y.W.; Kao, S.Y.; Wang, H.J.; Yang, M.H. Histone modification patterns correlate with patient outcome in oral squamous cell carcinoma. Cancer 2013, 119, 4259–4267. [Google Scholar] [CrossRef] [PubMed]
- Benard, A.; Goossens-Beumer, I.J.; van Hoesel, A.Q.; Horati, H.; de Graaf, W.; Putter, H.; Zeestraten, E.C.; Liefers, G.J.; van de Velde, C.J.; Kuppen, P.J. Nuclear expression of histone deacetylases and their histone modifications predicts clinical outcome in colorectal cancer. Histopathology 2015, 66, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Fraga, M.F.; Ballestar, E.; Villar-Garea, A.; Boix-Chornet, M.; Espada, J.; Schotta, G.; Bonaldi, T.; Haydon, C.; Ropero, S.; Petrie, K.; et al. Loss of acetylation at lys16 and trimethylation at lys20 of histone H4 is a common hallmark of human cancer. Nat. Genet. 2005, 37, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Matsumoto, A.; Hieda, M.; Shinchi, Y.; Ogihara, E.; Hamada, M.; Nishioka, Y.; Kimura, H.; Yoshidome, K.; Tsujimoto, M.; et al. Loss of histone H4K20 trimethylation predicts poor prognosis in breast cancer and is associated with invasive activity. Breast Cancer Res. 2014, 16, R66. [Google Scholar] [CrossRef] [PubMed]
- Benard, A.; Goossens-Beumer, I.J.; van Hoesel, A.Q.; de Graaf, W.; Horati, H.; Putter, H.; Zeestraten, E.C.; van de Velde, C.J.; Kuppen, P.J. Histone trimethylation at H3K4, H3K9 and H4K20 correlates with patient survival and tumor recurrence in early-stage colon cancer. BMC Cancer 2014, 14, 531. [Google Scholar] [CrossRef] [PubMed]
- Gezer, U.; Yörüker, E.E.; Keskin, M.; Kulle, C.B.; Dharuman, Y.; Holdenrieder, S. Histone methylation marks on circulating nucleosomes as novel blood-based biomarker in colorectal cancer. Int. J. Mol. Sci. 2015, 16, 29654–29662. [Google Scholar] [CrossRef] [PubMed]
- Wysocka, J.; Swigut, T.; Xiao, H.; Milne, T.A.; Kwon, S.Y.; Landry, J.; Kauer, M.; Tackett, A.J.; Chait, B.T.; Badenhorst, P.; et al. A phd finger of nurf couples histone H3 lysine 4 trimethylation with chromatin remodelling. Nature 2006, 442, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, R. Structure of histone deacetylases: Insights into substrate recognition and catalysis. Structure 2001, 9, 1127–1133. [Google Scholar] [CrossRef]
- Zheng, Y.C.; Ma, J.; Wang, Z.; Li, J.; Jiang, B.; Zhou, W.; Shi, X.; Wang, X.; Zhao, W.; Liu, H.M. A systematic review of histone lysine-specific demethylase 1 and its inhibitors. Med. Res. Rev. 2015, 35, 1032–1071. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Smith, E.; Shilatifard, A. The language of histone crosstalk. Cell 2010, 142, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharan, M.B.; Huang, F.; Sun, Z.-W. Histone H2B ubiquitination and beyond. Epigenetics 2010, 5, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- World Cancer Research Fund International. Breast Cancer Statistics. 2012. Available online: http://www.wcrf.org/int/cancer-facts-figures/data-specific-cancers/breast-cancer-statistics (accessed on 31 December 2016).
- McCullough, A. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed]
- Polyak, K. Heterogeneity in breast cancer. J. Clin. Investig. 2011, 121, 3786–3788. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Chen, Y.Y.; Scott, G.K.; Devries, S.; Chin, K.; Benz, C.C.; Waldman, F.M.; Hwang, E.S. Protein acetylation and histone deacetylase expression associated with malignant breast cancer progression. Clin. Cancer Res. 2009, 15, 3163–3171. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hu, H.; He, L.; Yu, X.; Liu, X.; Zhong, R.; Shu, M. A novel subtype classification and risk of breast cancer by histone modification profiling. Breast Cancer Res. Treat. 2016, 157, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Pfister, S.; Rea, S.; Taipale, M.; Mendrzyk, F.; Straub, B.; Ittrich, C.; Thuerigen, O.; Sinn, H.P.; Akhtar, A.; Lichter, P. The histone acetyltransferase hmof is frequently downregulated in primary breast carcinoma and medulloblastoma and constitutes a biomarker for clinical outcome in medulloblastoma. Int J. Cancer 2008, 122, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Janzer, A.; Becker, A.; Zimmer, A.; Schule, R.; Buettner, R.; Kirfel, J. Lysine-specific demethylase 1 (LSD1) is highly expressed in er-negative breast cancers and a biomarker predicting aggressive biology. Carcinogenesis 2010, 31, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Bourke, E.; Eriksson, L.A.; Kerin, M.J. Targeting cancer using kat inhibitors to mimic lethal knockouts. Biochem. Soc. Trans. 2016, 44, 979–986. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Cancer 5 Year Survival. 2016. Available online: http://www.cancer.org/research/cancerfactsstatistics/cancerfactsfigures2016/ (accessed on 31 December 2016).
- He, L.R.; Liu, M.Z.; Li, B.K.; Rao, H.L.; Liao, Y.J.; Guan, X.Y.; Zeng, Y.X.; Xie, D. Prognostic impact of H3K27ME3 expression on locoregional progression after chemoradiotherapy in esophageal squamous cell carcinoma. BMC Cancer 2009, 9, 461. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Shimada, H.; Noie, T.; Ohashi, M.; Oba, K.; Takahashi, Y. Clinical significance of serum tumor markers for gastric cancer: A systematic review of literature by the task force of the japanese gastric cancer association. Gastric Cancer 2014, 17, 26–33. [Google Scholar] [CrossRef] [PubMed]
- He, L.J.; Cai, M.Y.; Xu, G.L.; Li, J.J.; Weng, Z.J.; Xu, D.Z.; Luo, G.Y.; Zhu, S.L.; Xie, D. Prognostic significance of overexpression of EZH2 and H3K27ME3 proteins in gastric cancer. Asian Pac. J. Cancer Prev. 2012, 13, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Lech, G.; Słotwiński, R.; Słodkowski, M.; Krasnodębski, I.W. Colorectal cancer tumour markers and biomarkers: Recent therapeutic advances. World J. Gastroenterol. 2016, 22, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Phipps, A.I.; Buchanan, D.D.; Makar, K.W.; Win, A.K.; Baron, J.A.; Lindor, N.M.; Potter, J.D.; Newcomb, P.A. Kras-mutation status in relation to colorectal cancer survival: The joint impact of correlated tumour markers. Br. J. Cancer 2013, 108, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Barras, D. Braf mutation in colorectal cancer: An update. Biomark. Cancer 2015, 7, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, J.; Sidransky, D. DNA methylation markers in colorectal cancer. Cancer Metast. Rev. 2010, 29, 181–206. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Hieda, M.; Nishioka, Y.; Matsumoto, A.; Higashi, S.; Kimura, H.; Yamamoto, H.; Mori, M.; Matsuura, S.; Matsuura, N. Cancer-associated upregulation of histone H3 lysine 9 trimethylation promotes cell motility in vitro and drives tumor formation in vivo. Cancer Sci. 2013, 104, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Kondo, T.; Ma, D.; Niu, D.; Mochizuki, K.; Kawasaki, T.; Yamane, T.; Iino, H.; Fujii, H.; Katoh, R. Global histone modification of histone H3 in colorectal cancer and its precursor lesions. Hum. Pathol. 2012, 43, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Goossens-Beumer, I.J.; Benard, A.; van Hoesel, A.Q.; Zeestraten, E.C.M.; Putter, H.; Böhringer, S.; Liefers, G.-J.; Morreau, H.; van de Velde, C.J.H.; Kuppen, P.J.K. Age-dependent clinical prognostic value of histone modifications in colorectal cancer. Transl. Res. 2015, 165, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Yuen, B.T.; Knoepfler, P.S. Histone H3.3 mutations: A variant path to cancer. Cancer Cell 2013, 24, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Goldberg, M.S.; Cumberland, L.K.; Ratnakumar, K.; Segura, M.F.; Emanuel, P.O.; Menendez, S.; Vardabasso, C.; Leroy, G.; Vidal, C.I.; et al. The histone variant macroH2A suppresses melanoma progression through regulation of CDK8. Nature 2010, 468, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.; Kallen, C.B.; Dhar, R.; Baquero, M.T.; Mason, C.E.; Russell, B.A.; Shah, P.K.; Liu, J.; Khramtsov, A.; Tretiakova, M.S.; et al. Genomic analysis of estrogen cascade reveals histone variant H2A.Z associated with breast cancer progression. Mol. Syst. Biol. 2008, 4, 188. [Google Scholar] [CrossRef] [PubMed]
- Vardabasso, C.; Hasson, D.; Ratnakumar, K.; Chung, C.-Y.; Duarte, L.F.; Bernstein, E. Histone variants: Emerging players in cancer biology. Cell. Mol. Life Sci. 2014, 71, 379–404. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-H.; Miyai, K.; Sporn, J.C.; Luo, L.; Wang, J.Y.J.; Cosman, B.; Ramamoorthy, S. Loss of histone variant macroh2a2 expression associates with progression of anal neoplasm. J. Clin. Pathol. 2016, 69, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerke, A.; van Kuijk, S.J.; Sweep, F.C.; Nagtegaal, I.D.; Hoogerbrugge, N.; Martens, J.W.; Timmermans, M.A.; van Laarhoven, H.W.; Bussink, J.; Span, P.N. Constitutive expression of gamma-H2AX has prognostic relevance in triple negative breast cancer. Radiother. Oncol. 2011, 101, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Yin, T.C.; Chen, Y.T.; Chai, C.Y.; Wang, J.Y.; Liu, M.C.; Lin, Y.C.; Kan, J.Y. High expression of phospho-H2AX predicts a poor prognosis in colorectal cancer. Anticancer Res. 2015, 35, 2447–2453. [Google Scholar] [PubMed]
- Lichtenstein, A.V.; Melkonyan, H.S.; Tomei, L.D.; Umansky, S.R. Circulating nucleic acids and apoptosis. Ann. N. Y. Acad. Sci. 2001, 945, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Jahr, S.; Hentze, H.; Englisch, S.; Hardt, D.; Fackelmayer, F.O.; Hesch, R.D.; Knippers, R. DNA fragments in the blood plasma of cancer patients: Quantitations and evidence for their origin from apoptotic and necrotic cells. Cancer Res. 2001, 61, 1659–1665. [Google Scholar] [PubMed]
- Chan, K.C.A.; Zhang, J.; Chan, A.T.C.; Lei, K.I.K.; Leung, S.-F.; Chan, L.Y.S.; Chow, K.C.K.; Lo, Y.M.D. Molecular characterization of circulating EBV DNA in the plasma of nasopharyngeal carcinoma and lymphoma patients. Cancer Res. 2003, 63, 2028. [Google Scholar] [PubMed]
- Diaz, L.A.; Bardelli, A. Liquid biopsies: Genotyping circulating tumor DNA. J. Clin. Oncol. 2014, 32, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Kinzler, K.W. Digital PCR. Proc. Natl. Acad. Sci. USA 1999, 96, 9236–9241. [Google Scholar] [CrossRef] [PubMed]
- Dressman, D.; Yan, H.; Traverso, G.; Kinzler, K.W.; Vogelstein, B. Transforming single DNA molecules into fluorescent magnetic particles for detection and enumeration of genetic variations. Proc. Natl. Acad. Sci. USA 2003, 100, 8817–8822. [Google Scholar] [CrossRef] [PubMed]
- Forshew, T.; Murtaza, M.; Parkinson, C.; Gale, D.; Tsui, D.W.; Kaper, F.; Dawson, S.J.; Piskorz, A.M.; Jimenez-Linan, M.; Bentley, D.; et al. Noninvasive identification and monitoring of cancer mutations by targeted deep sequencing of plasma DNA. Sci. Transl. Med. 2012, 4, 136ra168. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Pantel, K. Circulating DNA as biomarker in breast cancer. Breast Cancer Res. 2015, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Karachaliou, N.; Mayo-de-las-Casas, C.; Molina-Vila, M.A.; Rosell, R. Real-time liquid biopsies become a reality in cancer treatment. Ann. Transl. Med. 2015, 3, 36. [Google Scholar] [PubMed]
- Imamura, T.; Komatsu, S.; Ichikawa, D.; Kawaguchi, T.; Miyamae, M.; Okajima, W.; Ohashi, T.; Arita, T.; Konishi, H.; Shiozaki, A.; et al. Liquid biopsy in patients with pancreatic cancer: Circulating tumor cells and cell-free nucleic acids. World J. Gastroenterol. 2016, 22, 5627–5641. [Google Scholar] [CrossRef] [PubMed]
- Hocking, J.; Mithraprabhu, S.; Kalff, A.; Spencer, A. Liquid biopsies for liquid tumors: Emerging potential of circulating free nucleic acid evaluation for the management of hematologic malignancies. Cancer Biol. Med. 2016, 13, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ramírez, C.; Cañadas-Garre, M.; Robles, A.I.; Molina, M.Á.; Faus-Dáder, M.J.; Calleja-Hernández, M.Á. Liquid biopsy in early stage lung cancer. Transl. Lung Cancer Res. 2016, 5, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Riethdorf, S.; Pantel, K. Liquid biopsies for surveillance and monitoring treatment response of bladder cancer. Ann. Transl. Med. 2016, 4, 379. [Google Scholar] [CrossRef] [PubMed]
- Holdenrieder, S.; von Pawel, J.; Nagel, D.; Stieber, P. Long-term stability of circulating nucleosomes in serum. Anticancer Res. 2010, 30, 1613–1615. [Google Scholar] [PubMed]
- Snyder, M.W.; Kircher, M.; Hill, A.J.; Daza, R.M.; Shendure, J. Cell-free DNA comprises an in vivo nucleosome footprint that informs its tissues-of-origin. Cell 2016, 164, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Hoon, D.S.B.; Pantel, K. Cell-free nucleic acids as biomarkers in cancer patients. Nat. Rev. Cancer 2011, 11, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Holdenrieder, S.; Nagel, D.; Schalhorn, A.; Heinemann, V.; Wilkowski, R.; von Pawel, J.; Raith, H.; Feldmann, K.; Kremer, A.E.; Müller, S.; et al. Clinical relevance of circulating nucleosomes in cancer. Ann. N. Y. Acad. Sci. 2008, 1137, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Holdenrieder, S.; Stieber, P.; Bodenmller, H.; Busch, M.; Fertig, G.; Forst, H.; Schalhorn, A.; Schmeller, N.; Untch, M.; Seidel, D. Nucleosomes in serum of patients with benign and malignant diseases. Int. J. Cancer 2001, 95, 114–120. [Google Scholar] [CrossRef]
- Kuroi, K.; Tanaka, C.; Toi, M. Plasma Nucleosome Levels in Node-Negative Breast Cancer Patients. Breast Cancer 1999, 6, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Stoetzer, O.J.; Fersching, D.M.I.; Salat, C.; Steinkohl, O.; Gabka, C.J.; Hamann, U.; Braun, M.; Feller, A.-M.; Heinemann, V.; Siegele, B.; et al. Prediction of response to neoadjuvant chemotherapy in breast cancer patients by circulating apoptotic biomarkers nucleosomes, dnase, cytokeratin-18 fragments and survivin. Cancer Lett. 2013, 336, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Holdenrieder, S.; Stieber, P.; von Pawel, J.; Raith, H.; Nagel, D.; Feldmann, K.; Seidel, D. Circulating nucleosomes predict the response to chemotherapy in patients with advanced non-small cell lung cancer. Clin. Cancer Res. 2004, 10, 5981–5987. [Google Scholar] [CrossRef] [PubMed]
- Fahmueller, Y.N.; Nagel, D.; Hoffmann, R.-T.; Tatsch, K.; Jakobs, T.; Stieber, P.; Holdenrieder, S. Predictive and prognostic value of circulating nucleosomes and serum biomarkers in patients with metastasized colorectal cancer undergoing selective internal radiation therapy. BMC Cancer 2012, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Deligezer, U.; Akisik, E.Z.; Akisik, E.E.; Kovancilar, M.; Bugra, D.; Erten, N.; Holdenrieder, S.; Dalay, N. H3K9me3/H4K20me3 ratio in circulating nucleosomes as potential biomarker for colorectal cancer. In Proceedings of the 6th International Conference on Circulating Nucleic Acids in Plasma and Serum, Hong Kong, China, 9–11 November 2009; Circulating Nucleic Acids in Plasma and Serum. Gahan, B.P., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 97–103. [Google Scholar]
- Poruk, K.E.; Gay, D.Z.; Brown, K.; Mulvihill, J.D.; Boucher, K.M.; Scaife, C.L.; Firpo, M.A.; Mulvihill, S.J. The clinical utility of CA 19-9 in pancreatic adenocarcinoma: Diagnostic and prognostic updates. Curr. Mol. Med. 2013, 13, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Bauden, M.; Pamart, D.; Ansari, D.; Herzog, M.; Eccleston, M.; Micallef, J.; Andersson, B.; Andersson, R. Circulating nucleosomes as epigenetic biomarkers in pancreatic cancer. Clin. Epigenet. 2015, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.; Herzog, M.; Romer, E.; Micallef, J.; Bulut, O.; Wilhelmsen, M.; Christensen, I.J.; Nielsen, H.J. Pre-analytical variables of circulating cell-free nucleosomes containing 5-methylcytosine DNA or histone modification H3K9ME3. Scand. J. Clin Lab. Investig. 2016, 76, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Casey, M.C.; Sweeney, K.J.; Brown, J.A.; Kerin, M.J. Exploring circulating micro-RNA in the neoadjuvant treatment of breast cancer. Int. J. Cancer 2016, 139, 12–22. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.; Brown, J.A.L.; Kerin, M.J. Metastatic breast cancer: The potential of miRNA for diagnosis and treatment monitoring. Cancer Metast. Rev. 2015, 34, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, H.M.; Miller, N.; Kelly, R.; Newell, J.; Kerin, M.J. Systemic miRNA-195 differentiates breast cancer from other malignancies and is a potential biomarker for detecting noninvasive and early stage disease. Oncologist 2010, 15, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M.; Calin, G.A. miRNAs, cancer, and stem cell division. Cell 2005, 122, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Liu, Z.; Zhao, Y.; Ding, Y.; Liu, H.; Xi, Y.; Xiong, W.; Li, G.; Lu, J.; Fodstad, O.; et al. microRNA-125b confers the resistance of breast cancer cells to paclitaxel through suppression of pro-apoptotic bcl-2 antagonist killer 1 (Bak1) expression. J. Biol. Chem. 2010, 285, 21496–21507. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Soberanis, F.; Andonegui, M.A.; Herrera, L.A. mir-125b-1 is repressed by histone modifications in breast cancer cell lines. Springerplus 2016, 5, 959. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Modification | Writer | Eraser | Function | Cancer role | Reference |
|---|---|---|---|---|---|
| Histone 2B | |||||
| Global hypoacetylation & hypomethylation | P300, ATF2 | Transcriptional activation | Low level-Prostate | [28] | |
| Modification | Writer | Eraser | Function | Cancer Role | Reference |
|---|---|---|---|---|---|
| Histone 3 | |||||
| H3K4Ac | HDAC 3 | Transcriptional activation | Low level—poorer prognosis in oral squamous cell carcinoma | [29] | |
| H3K4Me | SETD 7 | KDM 1A | Transcriptional activation | High levels in locally confined prostate cancer | [30] |
| H3K4Me2 | NSD 3 | KDM 1A | Transcriptional activation | Low levels in breast, pancreatic, renal and lung carcinomas of worse prognosis Low levels—higher rate of prostate cancer recurrence | [31,32,33] |
| KDM 5A | |||||
| KDM 5D | |||||
| H3K4Me3 | KDM 2B | Transcriptional activation Transcriptional elongation | High expression associated with poorer prognosis in hepatocellular carcinoma | [34] | |
| MLL | KDM | ||||
| MLL 3 | KDM 5A | ||||
| MLL 4 | KDM 5B | ||||
| PRDM 9 | KDM 5C | ||||
| SETD 1A | KDM 5D | ||||
| SETD 1B | PHF 8 | ||||
| NO 66 | |||||
| H3K9Ac | Gcn 5 | SIRT 1 | Transcriptional activation Telomere metabolism and function | Low level—breast cancer of poorer prognostic subtype Low levels—poorer prognosis in non-small cell lung cancer | [31,35] |
| SIRT 6 | |||||
| H3K9Me | G9a | KDM 1A | Transcriptional repression | Decreased levels indicates poor prognosis in renal cell cancer Decreased levels in bladder cancer | [36,37] |
| KDM 3 A & B | |||||
| H3K9Me2 | PHF 8 | Transcriptional repression and initiation | Low levels in prostate & pancreatic cancer Low levels—poorer outcome in prostate and renal cancer High levels in bladder cancer, correlates with pT stage and grade | [30,32,33,37] | |
| KMD 1A | |||||
| Glp 1 | KDM 1B | ||||
| G9a | KDM 3A | ||||
| PRDM 2 | KDM 3B | ||||
| KDM 4C | |||||
| KDM 4D | |||||
| KDM 7 | |||||
| H3K9Me3 | SETDB 1 | KDM 4A | Transcriptional repression and initiation Indexing of pericentromeric chromatin | Low levels in circulating nucleosomes in colorectal cancer, high in breast cancer High levels correlates with poorer survival and increased recurrence in gastric cancer Predicts survival in acute myeloid leukemia High levels in bladder cancer, correlates with pT stage and grade | [37,38,39,40,41] |
| SETDB 2 | KDM 4B | ||||
| SUV39H 1 & 2 | KDM 4C | ||||
| KDM 4D | |||||
| H3K18Ac | P300 | Transcriptional activation | Low levels—breast cancer of poorer prognostic subtype Low levels—better prognosis in esophageal squamous cell carcinoma, especially early Lower levels in muscle-invasive bladder cancer compared to non-muscle invasive and normal tissue Correlates to higher tumor grade in prostate cancer Low levels—poorer survival in pancreatic cancer | [31,32,42,43,44] | |
| CBP | |||||
| Elp3 | |||||
| H3K27Ac | P300 | Transcriptional activation | Up-regulated in colorectal cancer | [45] | |
| CBP | |||||
| H3K27Me | EZH 1 | Transcriptional activation | Low levels correlate to decreased survival in renal cell carcinoma (in addition to H3K27me2 &H3K27me3) | [46] | |
| EZH 2 | |||||
| Glp 1 | |||||
| G9a | |||||
| H3K27Me2 | EZH 1 | KDM 6B | Transcriptional repression | Lower levels correlate with poorer survival in colorectal cancer with liver metastases | [47] |
| EZH 2 | KDM 7 | ||||
| NSD 3 | PHF 8 | ||||
| H3K27Me3 | EZH 2 | KDM 6A | Transcriptional repression | Low levels—poorer prognosis in breast, ovarian and pancreatic cancer High expression correlates with vascular invasion and poorer prognosis in hepatocellular cancer High expression—increased survival in colorectal cancer High expression—better prognosis in non-small cell lung cancer High expression—poorer prognosis in oral squamous cell carcinoma | [48,49,50,51,52] |
| NSD 3 | KDM 6B | ||||
| H3K36Me2 | SETMAR | KDM 2A | Double strand repair & Non-homologous end joining | High level—correlates to histological subtype in primary colorectal cancer | [47] |
| NSD 1 | KDM 2B | ||||
| SMYD 2 | KDM 8 | ||||
| ASH 1L | |||||
| H3K36Me3 | SET D2 | KDM 4A | Antagonises PRC2-mediated H3K27 methylation | High level—correlates to lymph node spread in primary colorectal cancer | [47] |
| NSD 2 | NO66 | ||||
| H3K56Ac | HDAC 1 | DNA double stranded break repair | High expression—poorer prognosis in colorectal cancer | [53] | |
| CBP | SIRT 1 | ||||
| P300 | SIRT 3 | ||||
| SIRT 6 | |||||
| Modification | Writer | Eraser | Function | Cancer Role | Reference |
|---|---|---|---|---|---|
| Histone 4 | |||||
| H4Rme2 | PRMT 1 | JMJD 6 | Facilitates acetylation of H3 and H4 resulting in gene activation | Low levels in breast cancer of poorer prognosis | [31] |
| PRMT 5 | |||||
| PRMT 6 | |||||
| H4K12Ac | Gcn 5 | Histone deposition, telomere silencing Transcriptional activation, DNA repair Transcriptional activation | Low levels in breast cancer of poorer prognosis | [31] | |
| KAT1 | |||||
| Tip60 | |||||
| P300 | |||||
| CBP | |||||
| MYST2 | |||||
| H4K16Ac | Gcn 5 | DNA damage response & double-strand repair | Loss—Hallmark of cancer Low/absent in majority of breast cancers | [31,54] | |
| P300 | SIRT 1 | ||||
| CBP | SIRT 2 | ||||
| hMOF | |||||
| H4K20Me3 | SUV420H1 | Gene silencing Indexing of pericentromeric chromatin | Loss—Hallmark of cancer Reduced levels – independently associated with lower disease-free survival in breast cancer High expression (in combination with high H3K9Me3 & low H3K4Me3) correlates to better prognosis in colorectal cancer Significantly lower levels in circulating nucleosomes of colorectal cancer patients | [54,55,56,57] | |
| SUV420H2 | |||||
| NSD2 | |||||
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McAnena, P.; Brown, J.A.L.; Kerin, M.J. Circulating Nucleosomes and Nucleosome Modifications as Biomarkers in Cancer. Cancers 2017, 9, 5. https://doi.org/10.3390/cancers9010005
McAnena P, Brown JAL, Kerin MJ. Circulating Nucleosomes and Nucleosome Modifications as Biomarkers in Cancer. Cancers. 2017; 9(1):5. https://doi.org/10.3390/cancers9010005
Chicago/Turabian StyleMcAnena, Peter, James A. L. Brown, and Michael J. Kerin. 2017. "Circulating Nucleosomes and Nucleosome Modifications as Biomarkers in Cancer" Cancers 9, no. 1: 5. https://doi.org/10.3390/cancers9010005
APA StyleMcAnena, P., Brown, J. A. L., & Kerin, M. J. (2017). Circulating Nucleosomes and Nucleosome Modifications as Biomarkers in Cancer. Cancers, 9(1), 5. https://doi.org/10.3390/cancers9010005