

NRP1 and GFAP Expression in the Medulloblastoma Microenvironment: Implications for Angiogenesis and Tumor Progression

 , , ,
, , ,  and
and 
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Tissue from Medulloblastoma Patients
2.2. Automated Immunohistochemistry (IHC)
2.3. Quantification of Protein Staining
2.4. Immunofluorescence Technique (IF)
2.5. RNA Extraction of MB Tissue
2.6. Reverse Transcription and Real-Time PCR
2.7. Statistical Analysis
3. Results
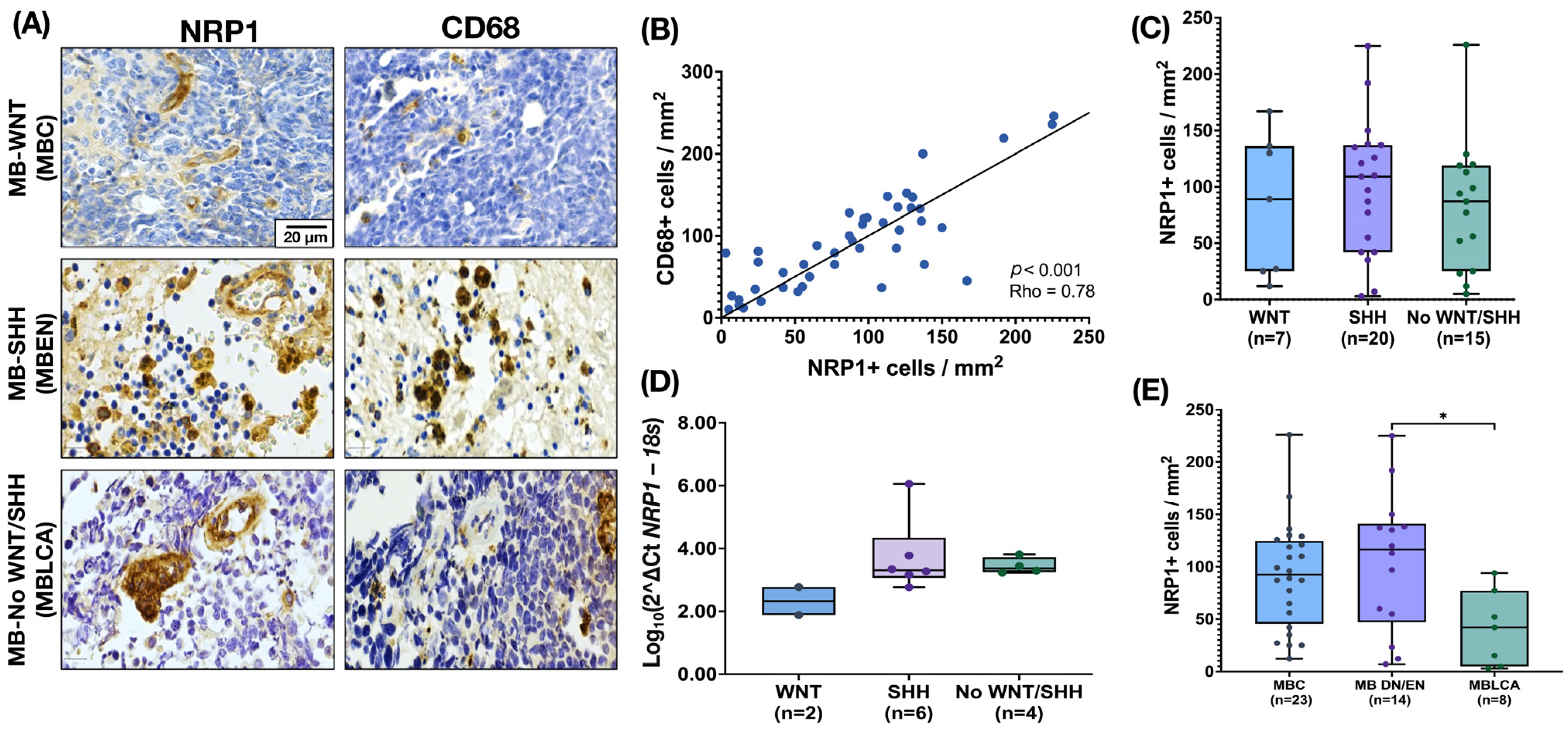
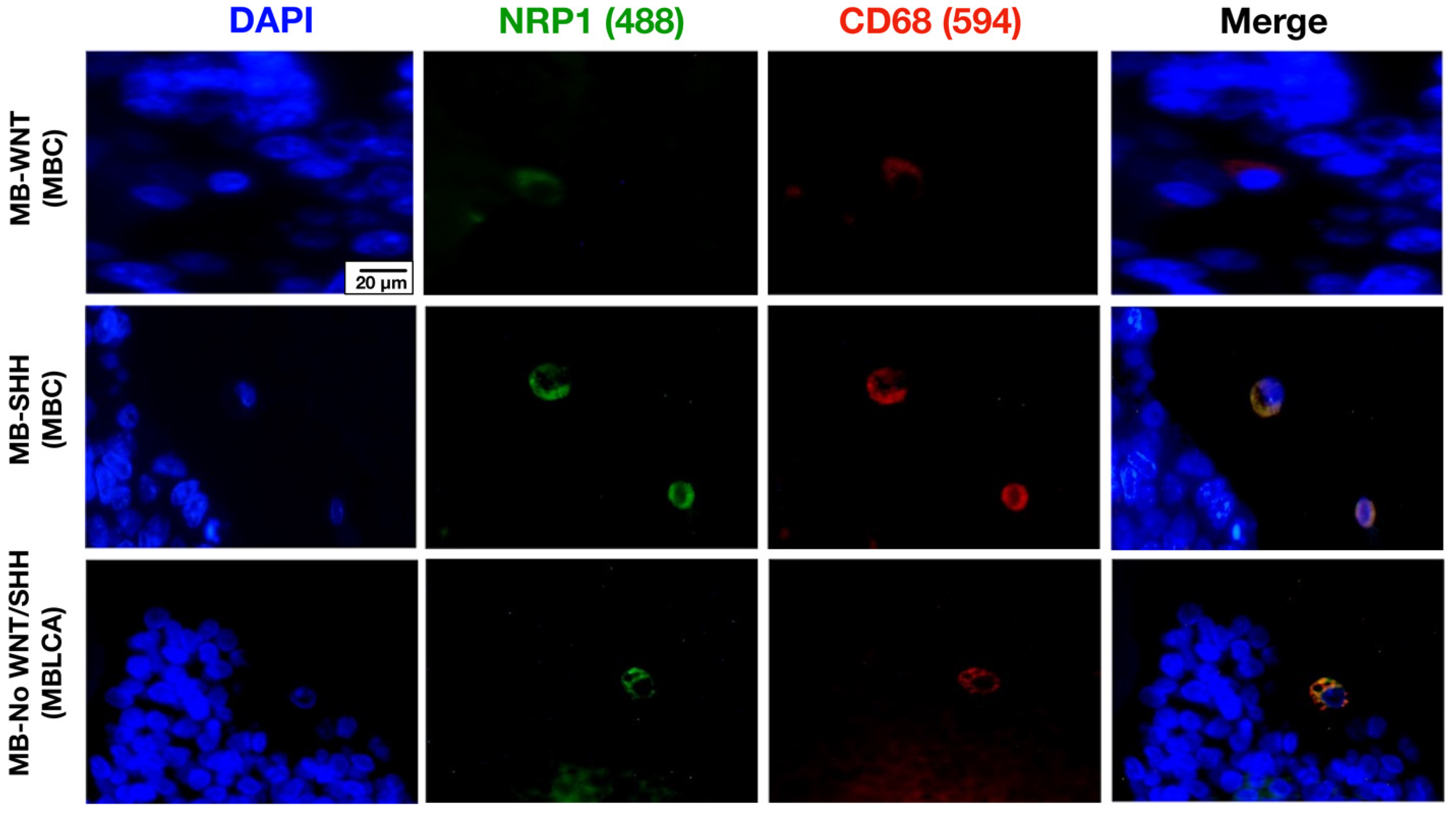
3.1. NRP1 Is Expressed by Tumor-Associated Microglia/Macrophages (TAMs) in MB Tissues Regardless of Their Molecular Subgroup
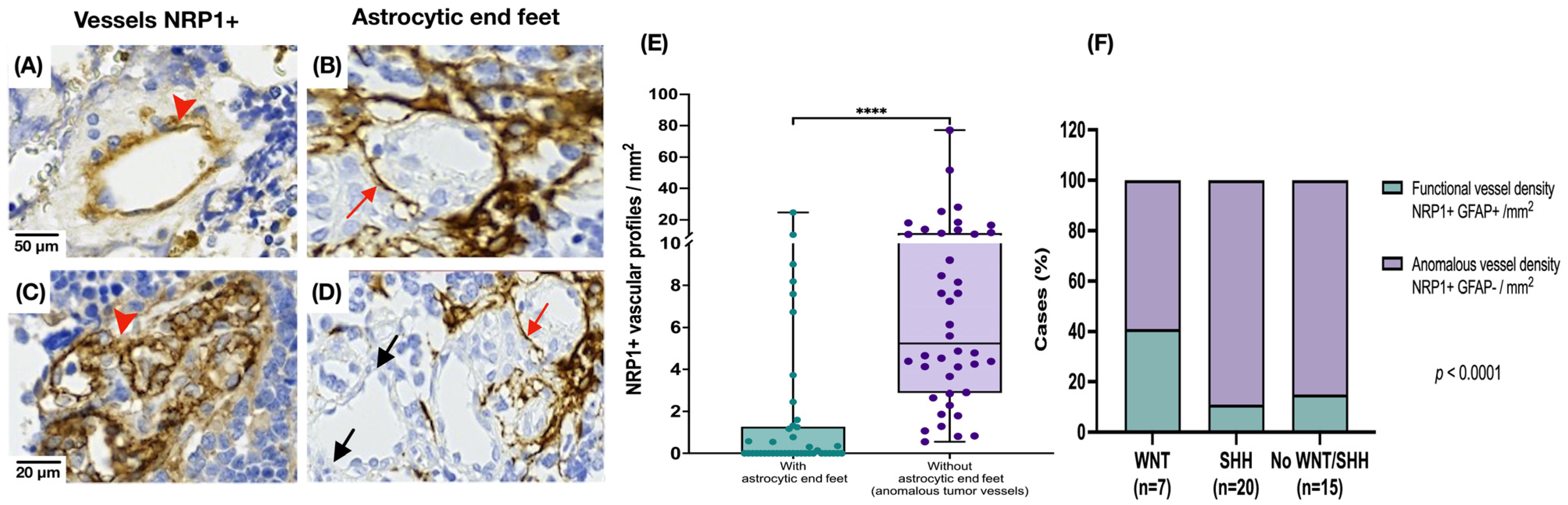
3.2. Medulloblastoma Tumor Vessels Show Strong NRP1 Expression and Limited Astrocytic Association
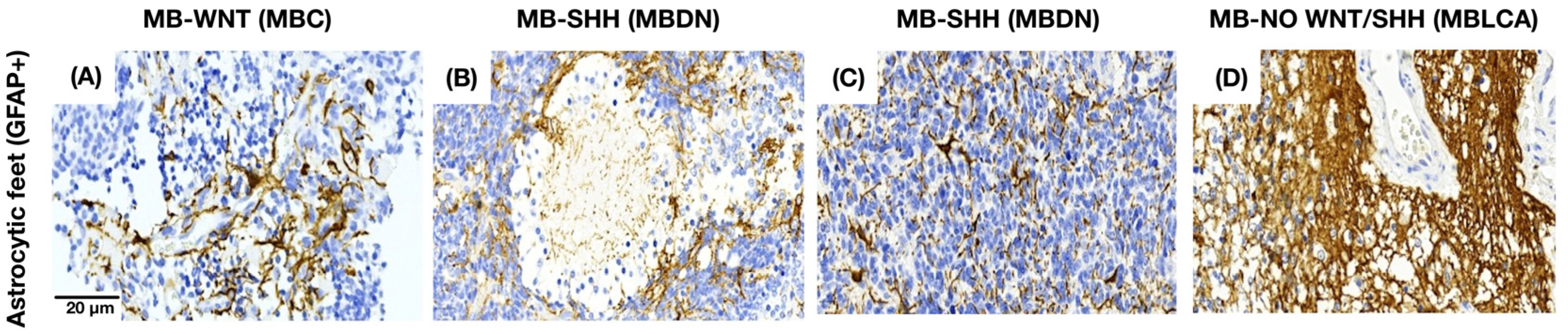
3.3. Astrocytes Show a Morphology-Dependent Distribution in the Medulloblastoma
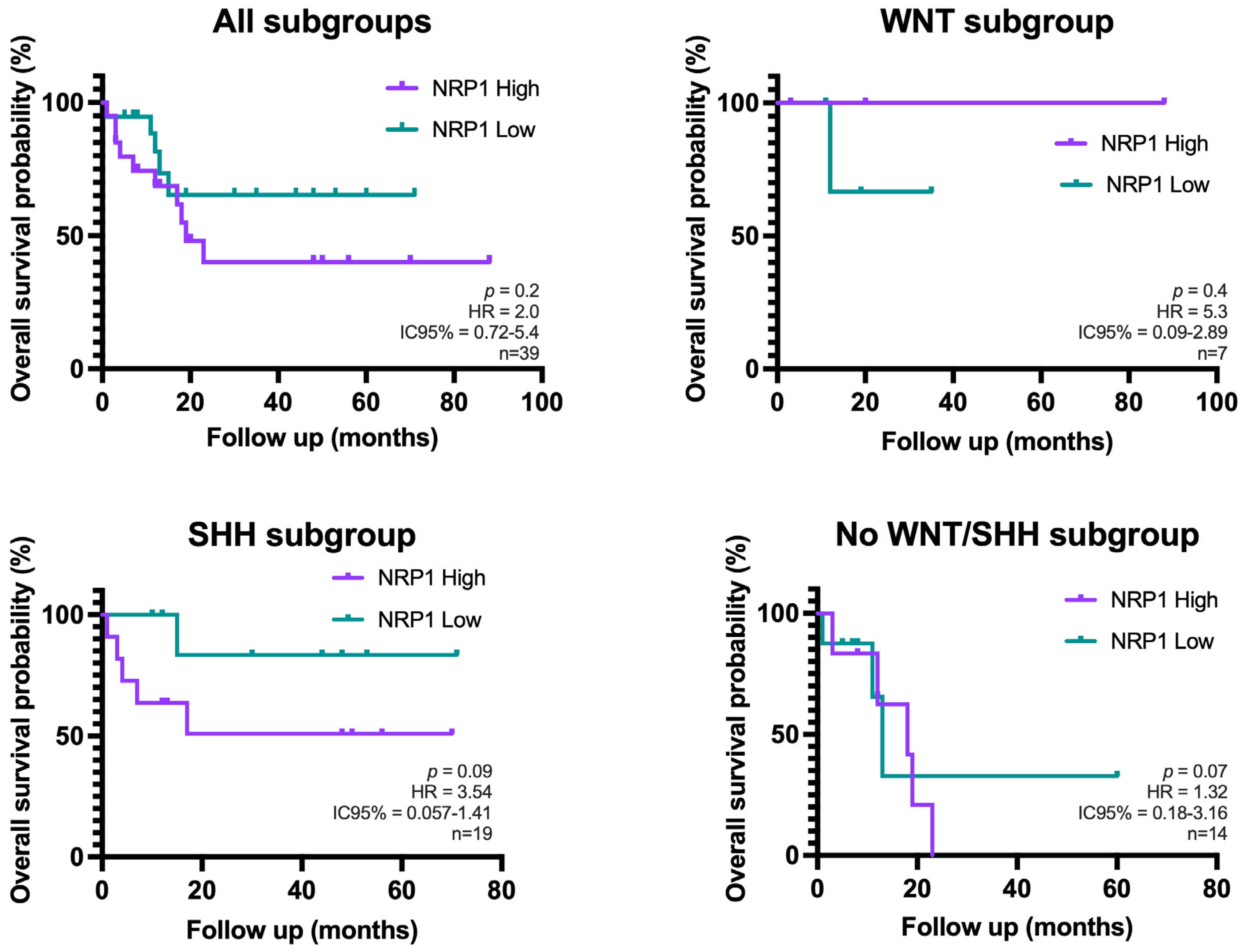
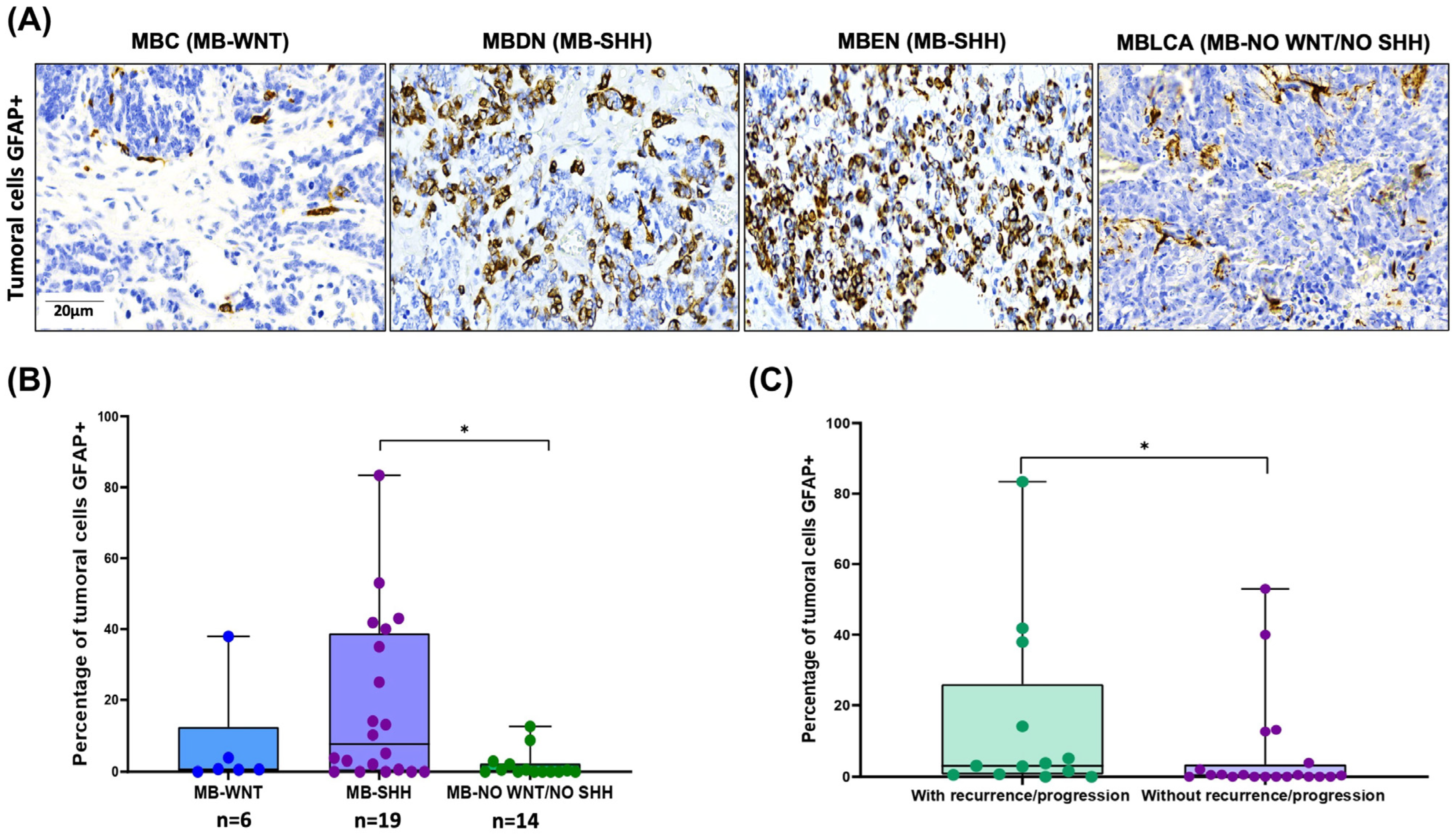
3.4. Aberrant GFAP Expression by MB Tumor Cells: Correlation with SHH Subtype and Clinical Outcome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BBB | Blood–brain barrier |
| GFAP | Glial fibrillary acidic protein |
| MB | Medulloblastoma |
| MBC | Medulloblastoma, classic |
| MBDN | Medulloblastoma, desmoplastic/nodular |
| MBEN | Medulloblastoma, extensive nodular |
| MBLCA | Medulloblastoma, large cell/anaplastic |
| NRP1 | Neuropilin-1 |
| PIGF | Placental growth factor |
| SHH | Sonic hedgehog |
| TAMs | Tumor-associated microglia/macrophages |
| TGF-β | Transforming growth factor-beta |
| TME | Tumor microenvironment |
| VEGF | Vascular endothelial growth factor |
| WNT | Wnt/Wingless |
References
- Ostrom, Q.T.; Price, M.; Neff, C.; Cioffi, G.; Waite, K.A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2015–2019. Neuro-Oncology 2022, 24, v1–v95. [Google Scholar] [CrossRef]
- Central Nervous System Tumours WHO Classification of Tumours. Central Nervous System Tumours. WHO Classification of Tumour; Wesseling, P., Pfister, S.M., Ellison, D.W., Eds.; World Health Organization Classification of Tumours; International Agency for Research on Cancer: Lyon, France, 2021; p. 1. ISBN 978-92-832-4508-7. [Google Scholar]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A Summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Hovestadt, V.; Smith, K.S.; Bihannic, L.; Filbin, M.G.; Shaw, M.L.; Baumgartner, A.; DeWitt, J.C.; Groves, A.; Mayr, L.; Weisman, H.R.; et al. Resolving Medulloblastoma Cellular Architecture by Single-Cell Genomics. Nature 2019, 572, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Wang, Y.; Liu, J.; Liu, Y.; Gong, X.; Sun, Y.; Sun, L.; Li, Z.; Wang, T.; et al. Tumor-Associated Macrophages Correlate with Better Outcome in SHH Medulloblastoma. Front. Oncol. 2025, 15, 1557313. [Google Scholar] [CrossRef] [PubMed]
- Du Chatinier, A.; Velilla, I.Q.; Meel, M.H.; Hoving, E.W.; Hulleman, E.; Metselaar, D.S. Microglia in Pediatric Brain Tumors: The Missing Link to Successful Immunotherapy. Cell Rep. Med. 2023, 4, 101246. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the Tumor Immune Microenvironment (TIME) for Effective Therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Margol, A.S.; Robison, N.J.; Gnanachandran, J.; Hung, L.T.; Kennedy, R.J.; Vali, M.; Dhall, G.; Finlay, J.L.; Erdreich-Epstein, A.; Krieger, M.D.; et al. Tumor-Associated Macrophages in SHH Subgroup of Medulloblastomas. Clin. Cancer Res. 2015, 21, 1457–1465. [Google Scholar] [CrossRef]
- Yao, M.; Ventura, P.B.; Jiang, Y.; Rodriguez, F.J.; Wang, L.; Perry, J.S.A.; Yang, Y.; Wahl, K.; Crittenden, R.B.; Bennett, M.L.; et al. Astrocytic Trans-Differentiation Completes a Multicellular Paracrine Feedback Loop Required for Medulloblastoma Tumor Growth. Cell 2020, 180, 502–520.e19. [Google Scholar] [CrossRef]
- Müller, S.; Kohanbash, G.; Liu, S.J.; Alvarado, B.; Carrera, D.; Bhaduri, A.; Watchmaker, P.B.; Yagnik, G.; Di Lullo, E.; Malatesta, M.; et al. Single-Cell Profiling of Human Gliomas Reveals Macrophage Ontogeny as a Basis for Regional Differences in Macrophage Activation in the Tumor Microenvironment. Genome Biol. 2017, 18, 234. [Google Scholar] [CrossRef]
- Maximov, V.; Chen, Z.; Wei, Y.; Robinson, M.H.; Herting, C.J.; Shanmugam, N.S.; Rudneva, V.A.; Goldsmith, K.C.; MacDonald, T.J.; Northcott, P.A.; et al. Tumour-Associated Macrophages Exhibit Anti-Tumoural Properties in Sonic Hedgehog Medulloblastoma. Nat. Commun. 2019, 10, 2410. [Google Scholar] [CrossRef]
- Yogi, K.; Sridhar, E.; Goel, N.; Jalali, R.; Goel, A.; Moiyadi, A.; Thorat, R.; Panwalkar, P.; Khire, A.; Dasgupta, A.; et al. MiR-148a, a microRNA Upregulated in the WNT Subgroup Tumors, Inhibits Invasion and Tumorigenic Potential of Medulloblastoma Cells by Targeting Neuropilin 1. Oncoscience 2015, 2, 334–348. [Google Scholar] [CrossRef]
- Snuderl, M.; Batista, A.; Kirkpatrick, N.D.; Ruiz de Almodovar, C.; Riedemann, L.; Walsh, E.C.; Anolik, R.; Huang, Y.; Martin, J.D.; Kamoun, W.; et al. Targeting Placental Growth Factor/Neuropilin 1 Pathway Inhibits Growth and Spread of Medulloblastoma. Cell 2013, 152, 1065–1076. [Google Scholar] [CrossRef]
- Ghasemi, D.R.; Okonechnikov, K.; Rademacher, A.; Tirier, S.; Maass, K.K.; Schumacher, H.; Joshi, P.; Gold, M.P.; Sundheimer, J.; Statz, B.; et al. Compartments in Medulloblastoma with Extensive Nodularity Are Connected through Differentiation along the Granular Precursor Lineage. Nat. Commun. 2024, 15, 269. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Ransom, B.; Goldman, S.A. New Roles for Astrocytes: Redefining the Functional Architecture of the Brain. Trends Neurosci. 2003, 26, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Traiffort, E.; Charytoniuk, D.; Watroba, L.; Faure, H.; Sales, N.; Ruat, M. Discrete Localizations of Hedgehog Signalling Components in the Developing and Adult Rat Nervous System. Eur. J. Neurosci. 1999, 11, 3199–3214. [Google Scholar] [CrossRef]
- Gronseth, E.; Gupta, A.; Koceja, C.; Kumar, S.; Kutty, R.G.; Rarick, K.; Wang, L.; Ramchandran, R. Astrocytes Influence Medulloblastoma Phenotypes and CD133 Surface Expression. PLoS ONE 2020, 15, e0235852. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Liu, Y.; Xu, L.; Sun, Z.; Zheng, D.; Liu, X.; Song, C.; Zhang, Y.; Liang, H.; et al. Tumor-associated Astrocytes Promote Tumor Progression of Sonic Hedgehog Medulloblastoma by Secreting Lipocalin-2. Brain Pathol. 2024, 34, e13212. [Google Scholar] [CrossRef]
- Miyahara, H.; Natsumeda, M.; Kanemura, Y.; Yamasaki, K.; Riku, Y.; Akagi, A.; Oohashi, W.; Shofuda, T.; Yoshioka, E.; Sato, Y.; et al. Topoisomerase IIβ Immunoreactivity (IR) Co-Localizes with Neuronal Marker-IR but Not Glial Fibrillary Acidic Protein-IR in GLI3-Positive Medulloblastomas: An Immunohistochemical Analysis of 124 Medulloblastomas from the Japan Children’s Cancer Group. Brain Tumor Pathol. 2021, 38, 109–121. [Google Scholar] [CrossRef]
- Narayan, V.; Jaiswal, J.; Sugur, H.; Sd, S.; Rao, S.; Chatterjee, A.; Gowda, H.; Arivazhagan, A.; Somanna, S.; Santosh, V. Proteomic Profiling of Medulloblastoma Reveals Novel Proteins Differentially Expressed within Each Molecular Subgroup. Clin. Neurol. Neurosurg. 2020, 196, 106028. [Google Scholar] [CrossRef]
- Varanasi, S.M.; Gulani, Y.; Rachamala, H.K.; Mukhopadhyay, D.; Angom, R.S. Neuropilin-1: A Multifaceted Target for Cancer Therapy. Curr. Oncol. 2025, 32, 203. [Google Scholar] [CrossRef] [PubMed]
- Fearnley, G.W.; Smith, G.A.; Abdul-Zani, I.; Yuldasheva, N.; Mughal, N.A.; Homer-Vanniasinkam, S.; Kearney, M.T.; Zachary, I.C.; Tomlinson, D.C.; Harrison, M.A.; et al. VEGF-A Isoforms Program Differential VEGFR2 Signal Transduction, Trafficking and Proteolysis. Biol. Open 2016, 5, 571–583. [Google Scholar] [CrossRef] [PubMed]
- He, N.; van Iperen, L.; de Jong, D.; Szuhai, K.; Helmerhorst, F.M.; van der Westerlaken, L.A.J.; Chuva de Sousa Lopes, S.M. Human Extravillous Trophoblasts Penetrate Decidual Veins and Lymphatics before Remodeling Spiral Arteries during Early Pregnancy. PLoS ONE 2017, 12, e0169849. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Kitsukawa, T.; Takagi, S.; Fujisawa, H. Developmentally Regulated Expression of a Cell Surface Protein, Neuropilin, in the Mouse Nervous System. J. Neurobiol. 1996, 29, 1–17. [Google Scholar] [CrossRef]
- Santana-Bejarano, M.B.; Grosso-Martínez, P.R.; Puebla-Mora, A.G.; Martínez-Silva, M.G.; Nava-Villalba, M.; Márquez-Aguirre, A.L.; Ortuño-Sahagún, D.; Godínez-Rubí, M. Pleiotrophin and the Expression of Its Receptors during Development of the Human Cerebellar Cortex. Cells 2023, 12, 1733. [Google Scholar] [CrossRef]
- Gong, C.; Valduga, J.; Chateau, A.; Richard, M.; Pellegrini-Moïse, N.; Barberi-Heyob, M.; Chastagner, P.; Boura, C. Stimulation of Medulloblastoma Stem Cells Differentiation by a Peptidomimetic Targeting Neuropilin-1. Oncotarget 2018, 9, 15312–15325. [Google Scholar] [CrossRef]
- Rodrigues, E.M.; Giovanini, A.F.; Ribas, C.A.P.M.; Malafaia, O.; Roesler, R.; Isolan, G.R. The Nervous System Development Regulator Neuropilin-1 as a Potential Prognostic Marker and Therapeutic Target in Brain Cancer. Cancers 2023, 15, 4922. [Google Scholar] [CrossRef]
- Chen, X.; Wu, S.; Yan, R.; Fan, L.; Yu, L.; Zhang, Y.; Wei, W.; Zhou, C.; Wu, X.; Zhong, M.; et al. The Role of the hypoxia-Nrp-1 Axis in the Activation of M2-like Tumor-associated Macrophages in the Tumor Microenvironment of Cervical Cancer. Mol. Carcinog. 2019, 58, 388–397. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Suzuki, E.; Nishie, M.; Kii, I.; Kataoka, T.R.; Hirata, M.; Inoue, M.; Pu, F.; Iwaisako, K.; Tsuda, M.; et al. Downregulation of Neuropilin-1 on Macrophages Modulates Antibody-Mediated Tumoricidal Activity. Cancer Immunol. Immunother. 2017, 66, 1131–1142. [Google Scholar] [CrossRef]
- Jiang, X.; Andjelkovic, A.V.; Zhu, L.; Yang, T.; Bennett, M.V.L.; Chen, J.; Keep, R.F.; Shi, Y. Blood-Brain Barrier Dysfunction and Recovery after Ischemic Stroke. Progress. Neurobiol. 2018, 163–164, 144–171. [Google Scholar] [CrossRef]
- Xu, X.; Chen, G.; Zhou, H.; Liu, Y.; Ding, H.; Wang, Z.; Shen, H.; Li, X.; Li, H. Downregulation of Nrp1 Transcription Promotes Blood–Brain Barrier Disruption Following Experimental Cerebral Ischemia–Reperfusion. Neurosci. Lett. 2024, 818, 137553. [Google Scholar] [CrossRef] [PubMed]
- Phoenix, T.N.; Patmore, D.M.; Boop, S.; Boulos, N.; Jacus, M.O.; Patel, Y.T.; Roussel, M.F.; Finkelstein, D.; Goumnerova, L.; Perreault, S.; et al. Medulloblastoma Genotype Dictates Blood Brain Barrier Phenotype. Cancer Cell 2016, 29, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, A.; Okonechnikov, K.; Stichel, D.; Ryzhova, M.; Schrimpf, D.; Sahm, F.; Sievers, P.; Absalyamova, O.; Zheludkova, O.; Golanov, A.; et al. Integrated Molecular Analysis of Adult Sonic Hedgehog (SHH)-Activated Medulloblastomas Reveals Two Clinically Relevant Tumor Subsets with VEGFA as Potent Prognostic Indicator. Neuro-Oncology 2021, 23, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.M.; Keir, S.T.; Venkatraman, T.; Lascola, C.; Yeom, K.W.; Nixon, A.B.; Liu, Y.; Picard, D.; Remke, M.; Bigner, D.D.; et al. The Role of Angiogenesis in Group 3 Medulloblastoma Pathogenesis and Survival. Neuro-Oncology 2017, 19, 1217–1227. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef]
- Hu, C.; Jiang, X. Role of NRP-1 in VEGF-VEGFR2-Independent Tumorigenesis. Targ. Oncol. 2016, 11, 501–505. [Google Scholar] [CrossRef]
- He, Z.; Tessier-Lavigne, M. Neuropilin Is a Receptor for the Axonal Chemorepellent Semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef]
- Miao, H.; Lee, P.; Lin, H.; Soker, S.; Klagsbrun, M. Neuropilin-1 Expression by Tumor Cells Promotes Tumor Angiogenesis and Progression. FASEB J. 2000, 14, 2532–2539. [Google Scholar] [CrossRef]
- Caponegro, M.D.; Moffitt, R.A.; Tsirka, S.E. Expression of Neuropilin-1 Is Linked to Glioma Associated Microglia and Macrophages and Correlates with Unfavorable Prognosis in High Grade Gliomas. Oncotarget 2018, 9, 35655–35665. [Google Scholar] [CrossRef]
- Miyauchi, J.T.; Chen, D.; Choi, M.; Nissen, J.C.; Shroyer, K.R.; Djordevic, S.; Zachary, I.C.; Selwood, D.; Tsirka, S.E. Ablation of Neuropilin 1 from Glioma-Associated Microglia and Macrophages Slows Tumor Progression. Oncotarget 2016, 7, 9801–9814. [Google Scholar] [CrossRef]
- Dang, M.T.; Gonzalez, M.V.; Gaonkar, K.S.; Rathi, K.S.; Young, P.; Arif, S.; Zhai, L.; Alam, Z.; Devalaraja, S.; Jerrick To, T.K.; et al. Macrophages in SHH Subgroup Medulloblastoma Display Dynamic Heterogeneity That Varies with Treatment Modality. Cell Rep. 2023, 42, 112600. [Google Scholar] [CrossRef]
- Ellison, D.W.; Dalton, J.; Kocak, M.; Nicholson, S.L.; Fraga, C.; Neale, G.; Kenney, A.M.; Brat, D.J.; Perry, A.; Yong, W.H.; et al. Medulloblastoma: Clinicopathological Correlates of SHH, WNT, and Non-SHH/WNT Molecular Subgroups. Acta Neuropathol. 2011, 121, 381–396. [Google Scholar] [CrossRef]
- De Araújo, M.A.; Malafaia, O.; Ribas Filho, J.M.; Fratini, L.; Roesler, R.; Isolan, G.R. Low Expression of the NRP1 Gene Is Associated with Shorter Overall Survival in Patients with Sonic Hedgehog and Group 3 Medulloblastoma. Int. J. Mol. Sci. 2023, 24, 11601. [Google Scholar] [CrossRef]
- Northcott, P.A.; Robinson, G.W.; Kratz, C.P.; Mabbott, D.J.; Pomeroy, S.L.; Clifford, S.C.; Rutkowski, S.; Ellison, D.W.; Malkin, D.; Taylor, M.D.; et al. Medulloblastoma. Nat. Rev. Dis. Prim. 2019, 5, 11. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, Y.; Mangalam, A.K.; Guo, Y.; LaFrance-Corey, R.G.; Gamez, J.D.; Atanga, P.A.; Clarkson, B.D.; Zhang, Y.; Wang, E.; et al. Neuropilin-1 Modulates Interferon-γ-Stimulated Signaling in Brain Microvascular Endothelial Cells. J. Cell Sci. 2016, 129, 3911–3921. [Google Scholar] [CrossRef]
- Nduom, E.K.; Yang, C.; Merrill, M.J.; Zhuang, Z.; Lonser, R.R. Characterization of the Blood-Brain Barrier of Metastatic and Primary Malignant Neoplasms: Laboratory Investigation. J. Neurosurg. 2013, 119, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Watkins, S.; Robel, S.; Kimbrough, I.F.; Robert, S.M.; Ellis-Davies, G.; Sontheimer, H. Disruption of Astrocyte–Vascular Coupling and the Blood–Brain Barrier by Invading Glioma Cells. Nat. Commun. 2014, 5, 4196. [Google Scholar] [CrossRef] [PubMed]
- Okano-Uchida, T.; Himi, T.; Komiya, Y.; Ishizaki, Y. Cerebellar Granule Cell Precursors Can Differentiate into Astroglial Cells. Proc. Natl. Acad. Sci. USA 2004, 101, 1211–1216. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | n/Median | Percentage/Range |
|---|---|---|
| Sex (Male/Female) | 33/12 | 73.33/26.66 |
| Age (median, range) Follow-up time (months) | 7 12 | 1–33 1–88 |
| Risk stratification | ||
| Standard | 20 | 44.44 |
| High | 17 | 37.77 |
| Not available | 8 | 17.77 |
| Recurrence/Progression | 15 | 33.33 |
| Death | 15 | 33.33 |
| Histological subtypes | ||
| MBC | 23 | 51.11 |
| MBDN | 11 | 24.44 |
| MBEN | 3 | 6.66 |
| MBLCA | 8 | 17.77 |
| Molecular subtypes | ||
| MB-WNT | 7 | 15.55 |
| MB-SHH | 20 | 44.44 |
| MB-No WNT/No SHH | 15 | 33.33 |
| MB-NOS | 3 | 6.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana-Bejarano, M.B.; Reyes-Mata, M.P.; Guerrero-García, J.d.J.; Ortuño-Sahagún, D.; Godínez-Rubí, M. NRP1 and GFAP Expression in the Medulloblastoma Microenvironment: Implications for Angiogenesis and Tumor Progression. Cancers 2025, 17, 2417. https://doi.org/10.3390/cancers17152417
Santana-Bejarano MB, Reyes-Mata MP, Guerrero-García JdJ, Ortuño-Sahagún D, Godínez-Rubí M. NRP1 and GFAP Expression in the Medulloblastoma Microenvironment: Implications for Angiogenesis and Tumor Progression. Cancers. 2025; 17(15):2417. https://doi.org/10.3390/cancers17152417
Chicago/Turabian StyleSantana-Bejarano, Margarita Belem, María Paulina Reyes-Mata, José de Jesús Guerrero-García, Daniel Ortuño-Sahagún, and Marisol Godínez-Rubí. 2025. "NRP1 and GFAP Expression in the Medulloblastoma Microenvironment: Implications for Angiogenesis and Tumor Progression" Cancers 17, no. 15: 2417. https://doi.org/10.3390/cancers17152417
APA StyleSantana-Bejarano, M. B., Reyes-Mata, M. P., Guerrero-García, J. d. J., Ortuño-Sahagún, D., & Godínez-Rubí, M. (2025). NRP1 and GFAP Expression in the Medulloblastoma Microenvironment: Implications for Angiogenesis and Tumor Progression. Cancers, 17(15), 2417. https://doi.org/10.3390/cancers17152417