
XPC Protects against Carcinogen-Induced Histologic Progression to Lung Squamous Cell Carcinoma by Reduced Basal Epithelial Cell Proliferation

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
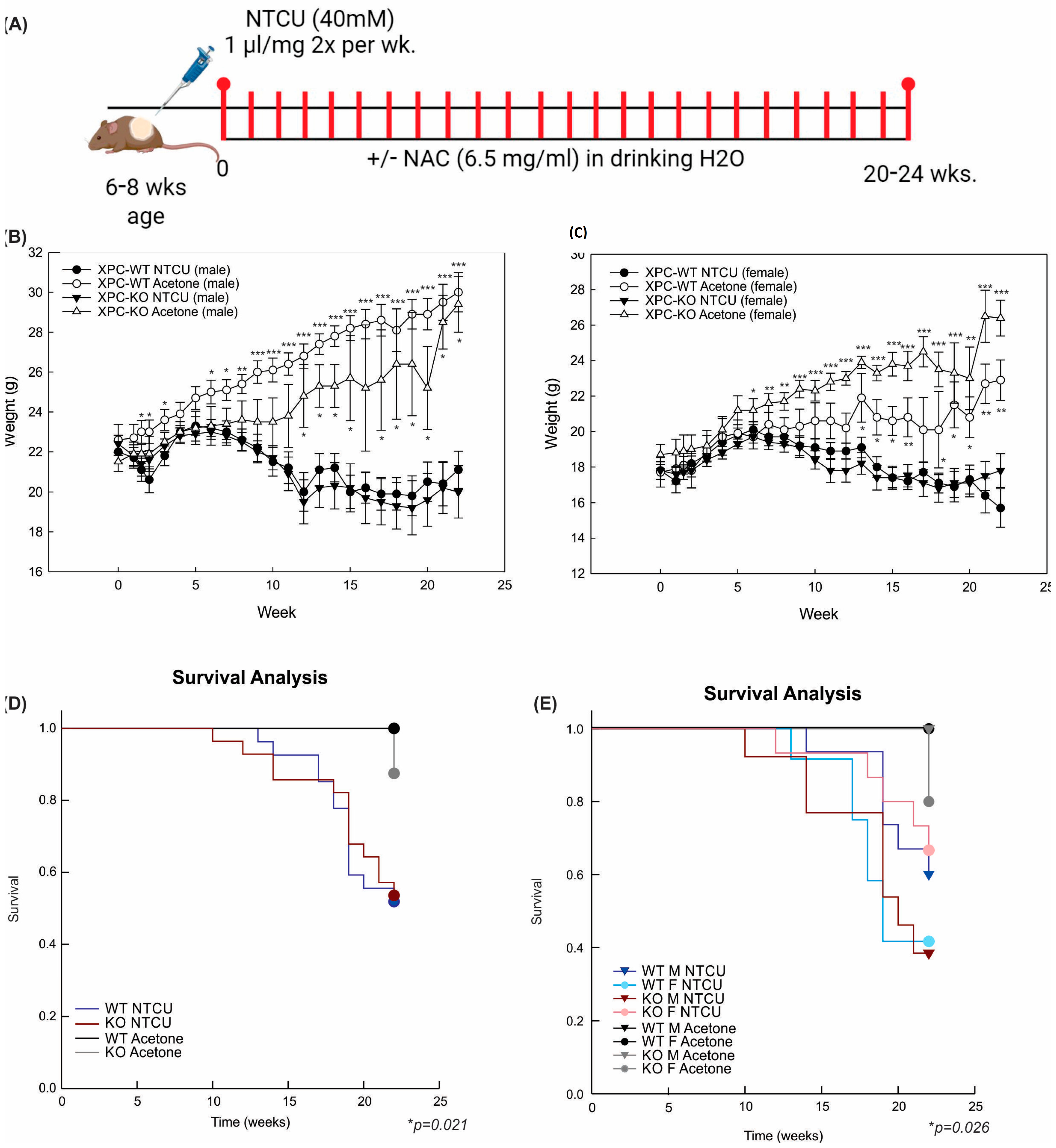
2.2. Animal Model
2.3. Mouse Harvest, Tissue Specimen Collection and Processing
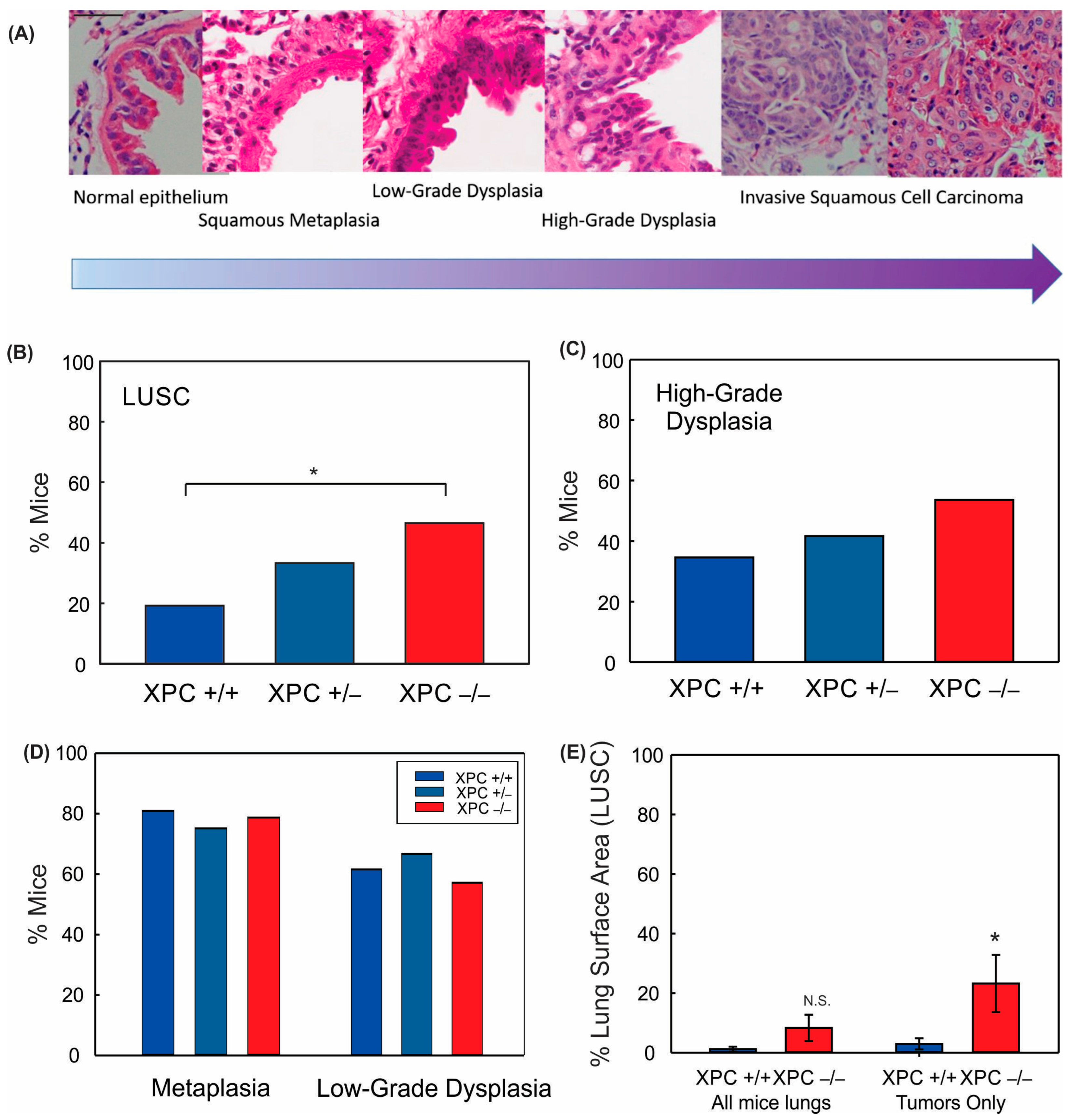
2.4. Slide Evaluation for Histology and Percent Lung Calculations
2.5. Immunofluorescence Microscopy
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
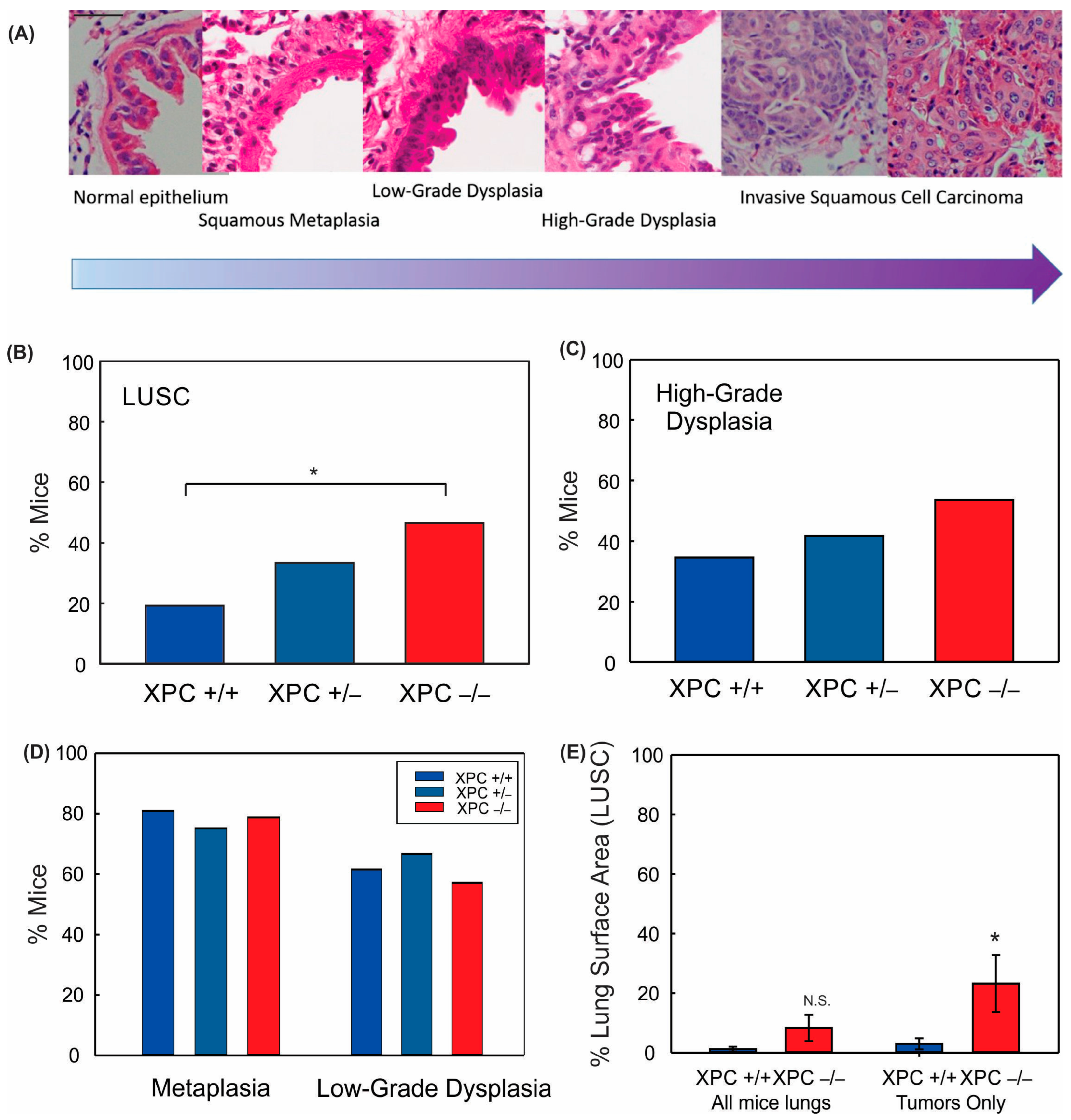
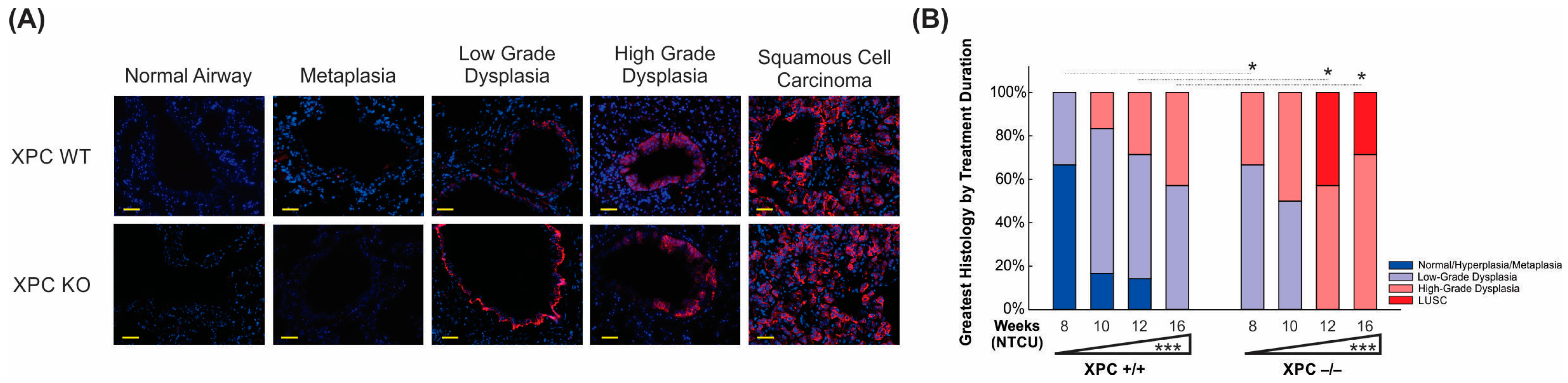
3.1. XPC Deficiency Leads to Advanced Histologic Grade of Lung Squamous Cell Carcinoma
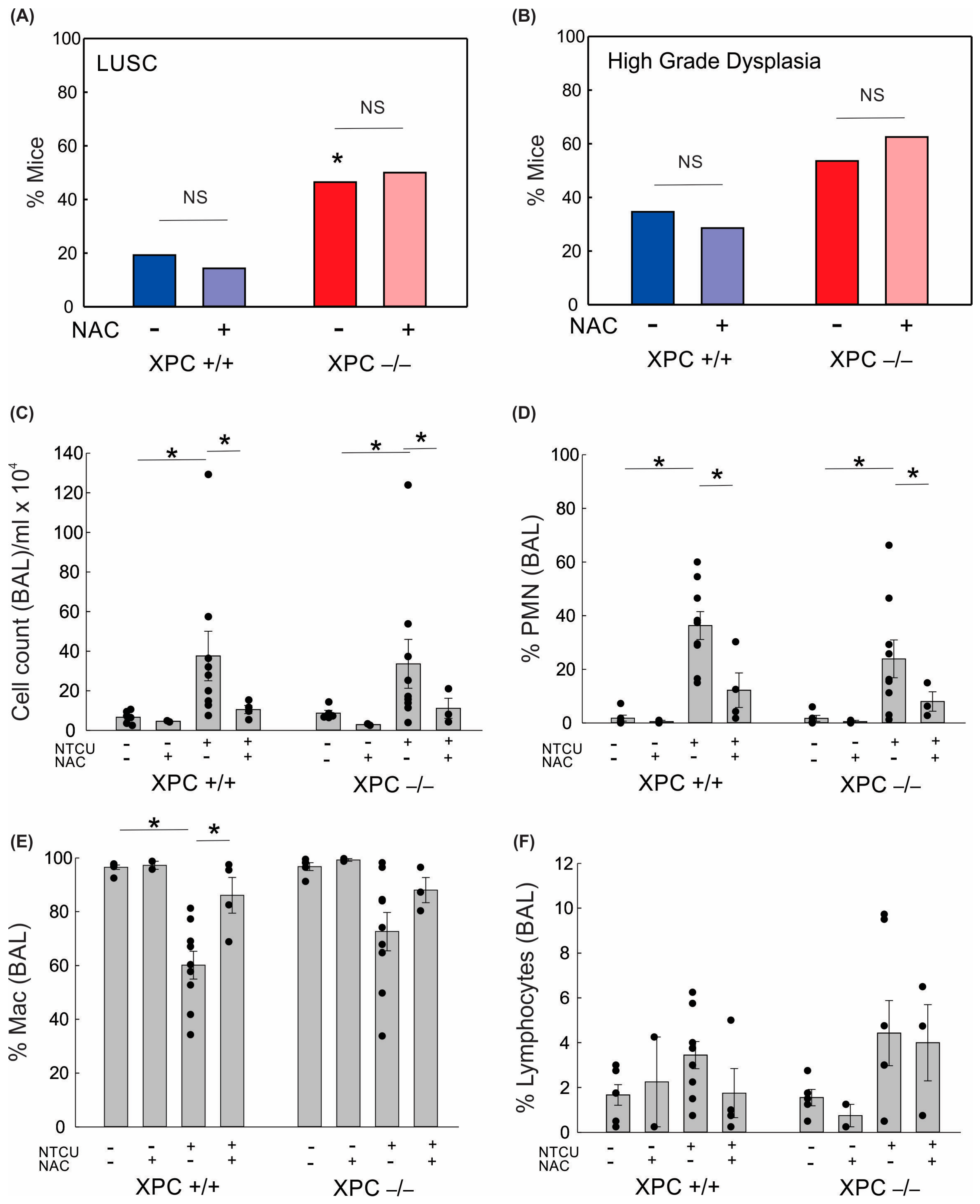
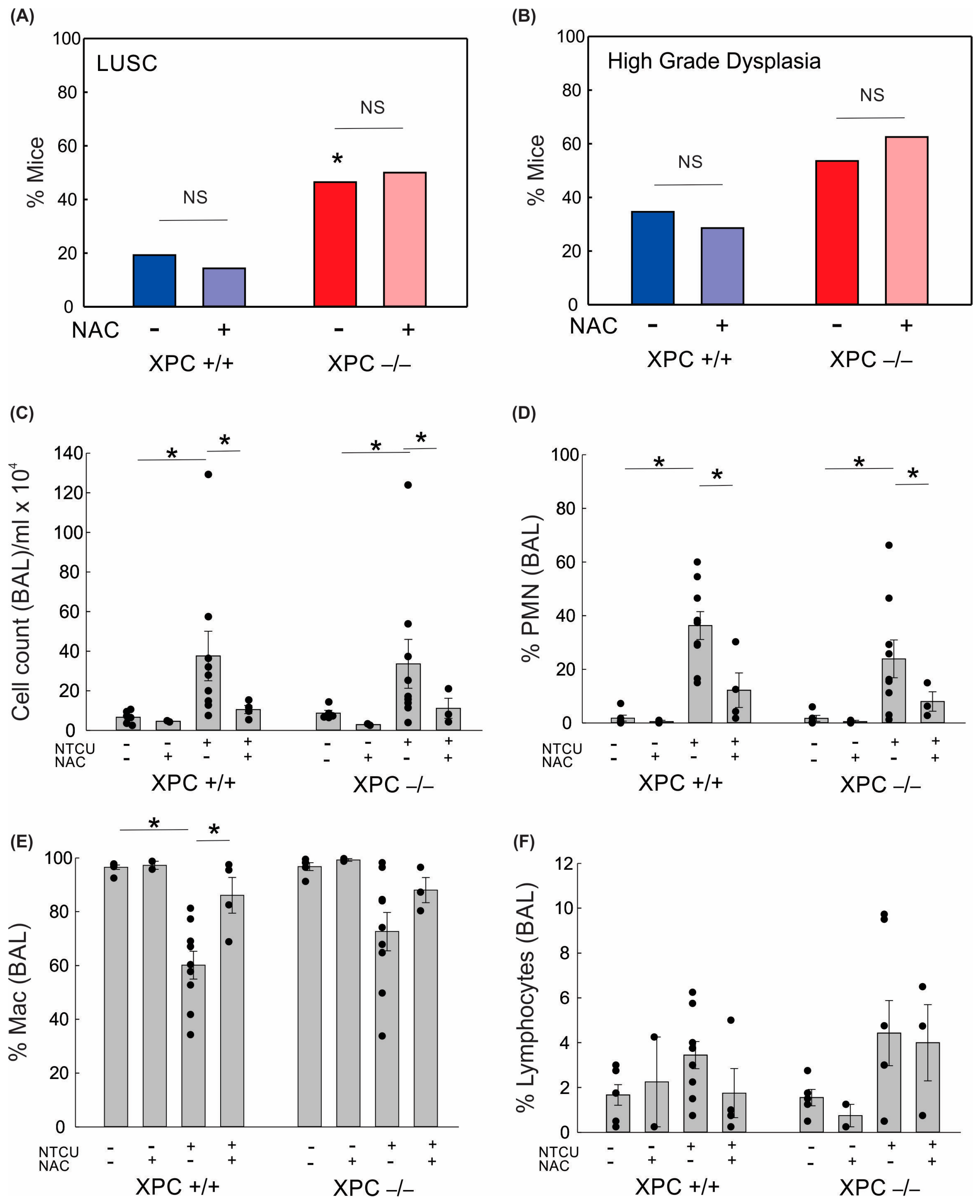
3.2. LUSC Development in XPC KO Mice Is Not Mitigated by N-Acetylcysteine or Antioxidant-Induced Changes to Alveolar Inflammation
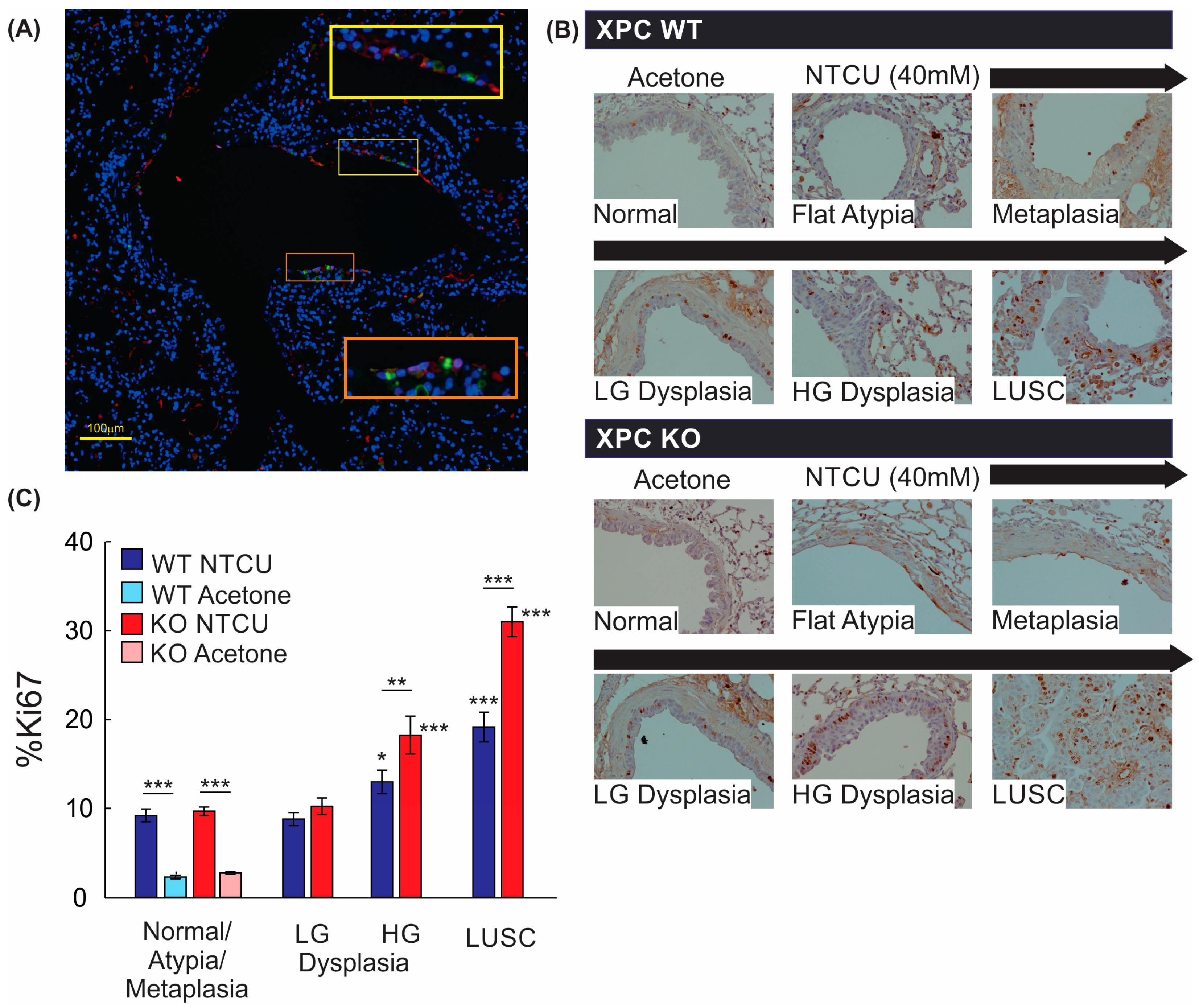
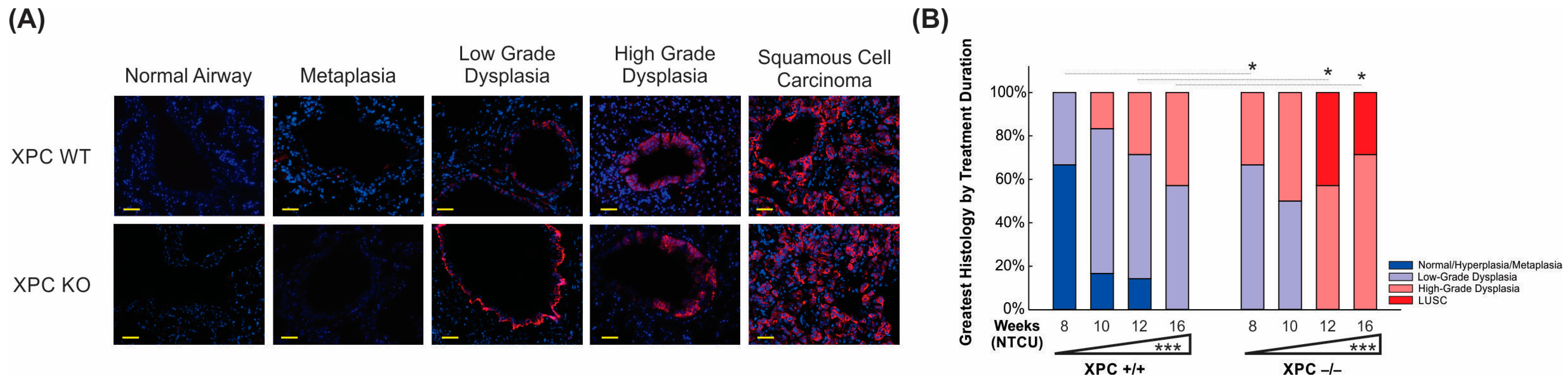
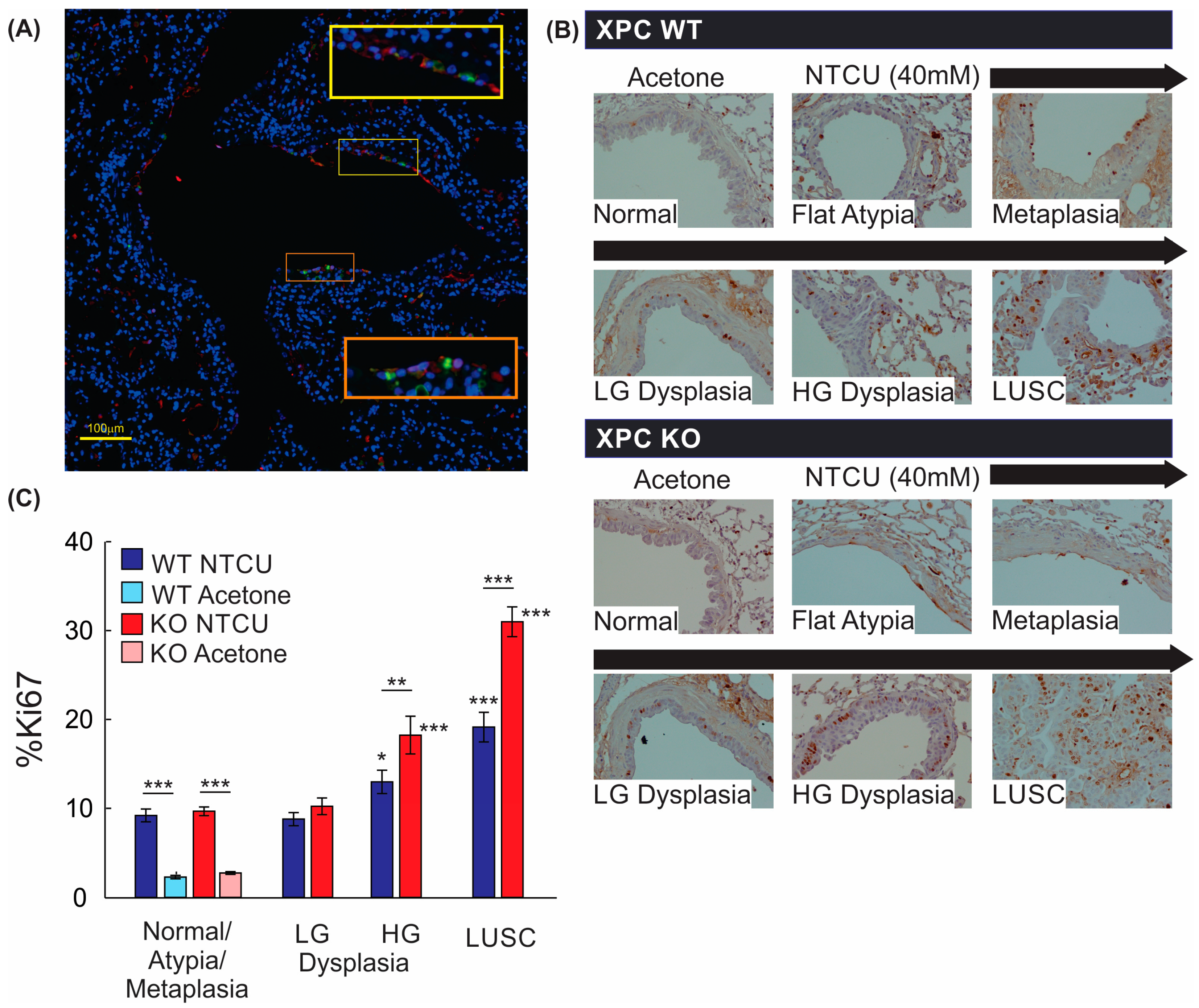
3.3. Increased Histologic Progression of LUSC Development in XPC Deficiency Is Related to Basal Cell Proliferative Activity
3.4. Airways of XPC KO Mice Show Increased Proliferation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cancer Statistics: NIH National Cancer Institute Surveillance, Epidemiology and End Results Website. Available online: https://seer.cancer.gov (accessed on 23 August 2022).
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- SEER*Explorer: An Interactive Website for SEER Cancer Statistics. Available online: https://seer.cancer.gov/explorer/ (accessed on 23 January 2024).
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Aberle, D.R.; Adams, A.M.; Berg, C.D.; Black, W.C.; Clapp, J.D.; Fagerstrom, R.M.; Gareen, I.F.; Gatsonis, C.; Marcus, P.M.; Sicks, J.D. The National Lung Screening Trial Research Team. Reduced Lung-Cancer Mortality with Low-Dose Computed Tomographic Screening. N. Engl. J. Med. 2011, 265, 395–409. [Google Scholar]
- de Koning, H.J.; van der Aalst, C.M.; de Jong, P.A.; Scholten, E.T.; Nackaerts, K.; Heuvelmans, M.A.; Lammers, J.J.; Weenink, C.; Yousaf-Khan, U.; Horeweg, N.; et al. Reduced Lung-Cancer Mortality with Volume CT Screening in a Randomized Trial. N. Engl. J. Med. 2020, 382, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, U.; Silva, M.; Sestini, S.; Sabia, F.; Boeri, M.; Cantarutti, A.; Sverzellati, N.; Sozzi, G.; Corrao, G.; Marchiano, A. Prolonged lung cancer screening reduced 10-year mortality in the MILD trial: New confirmation of lung cancer screening efficacy. Ann. Oncol. 2019, 30, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, P.F.; Church, T.R.; Izmirlian, G.; Kramer, B.S. The National Lung Screening Trial: Results stratified by demographics, smoking history, and lung cancer histology. Cancer 2013, 119, 3976–3983. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Borresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Ju, Y.S.; Haase, K.; Van Loo, P.; Martincorena, I.; Nik-Zainal, S.; Totoki, Y.; Fujimoto, A.; Nakagawa, H.; Shibata, T.; et al. Mutational signatures associated with tobacco smoking in human cancer. Science 2016, 354, 618–622. [Google Scholar] [CrossRef]
- Zhou, H.; Saliba, J.; Sandusky, G.E.; Sears, C.R. XPC protects against smoking and carcinogen-induced lung adenocarcinoma. Carcinogenesis 2019, 40, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Melis, J.P.; Luijten, M.; Mullenders, L.H.; van Steeg, H. The role of XPC: Implications in cancer and oxidative DNA damage. Mutat. Res. 2011, 728, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, N.A.; Wiese, B.M.; Sears, C.R. Xeroderma Pigmentosum Complementation Group C (XPC): Emerging Roles in Non-Dermatologic Malignancies. Front. Oncol. 2022, 12, 846965. [Google Scholar] [CrossRef] [PubMed]
- Hollander, M.C.; Philburn, R.T.; Patterson, A.D.; Velasco-Miguel, S.; Friedberg, E.C.; Linnoila, R.I.; Fornace, A.J., Jr. Deletion of XPC leads to lung tumors in mice and is associated with early events in human lung carcinogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 13200–13205. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.R.; Zhou, H.; Justice, M.J.; Fisher, A.J.; Saliba, J.; Lamb, I.; Wicker, J.; Schweitzer, K.S.; Petrache, I. Xeroderma Pigmentosum Group C Deficiency Alters Cigarette Smoke DNA Damage Cell Fate and Accelerates Emphysema Development. Am. J. Respir. Cell Mol. Biol. 2018, 58, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Hudish, T.M.; Opincariu, L.I.; Mozer, A.B.; Johnson, M.S.; Cleaver, T.G.; Malkoski, S.P.; Merrick, D.T.; Keith, R.L. N-nitroso-tris-chloroethylurea induces premalignant squamous dysplasia in mice. Cancer Prev. Res. 2012, 5, 283–289. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Yan, Y.; Lemon, W.J.; LaRegina, M.; Morrison, C.; Lubet, R.; You, M. A Chemically Induced Model for Squamous Cell Carcinoma of the Lung in Mice: Histophatology and Strain Susceptibility. Cancer Res. 2004, 64, 1647–1654. [Google Scholar] [CrossRef]
- Ghosh, M.; Dwyer-Nield, L.D.; Kwon, J.B.; Barthel, L.; Janssen, W.J.; Merrick, D.T.; Keith, R.L. Tracheal dysplasia precedes bronchial dysplasia in mouse model of N-nitroso trischloroethylurea induced squamous cell lung cancer. PLoS ONE 2015, 10, e0122823. [Google Scholar] [CrossRef]
- Melis, J.P.; Kuiper, R.V.; Zwart, E.; Robinson, J.; Pennings, J.L.; van Oostrom, C.T.; Luijten, M.; van Steeg, H. Slow accumulation of mutations in Xpc mice upon induction of oxidative stress. DNA Repair. 2013, 12, 1081–1086. [Google Scholar] [CrossRef]
- Wang, M.; Herbst, R.S.; Boshoff, C. Toward personalized treatment approaches for non-small-cell lung cancer. Nat. Med. 2021, 27, 1345–1356. [Google Scholar] [CrossRef]
- Jamal-Hanjani, M.; Wilson, G.A.; McGranahan, N.; Birkbak, N.J.; Watkins, T.B.K.; Veeriah, S.; Shafi, S.; Johnson, D.H.; Mitter, R.; Rosenthal, R.; et al. Tracking the Evolution of Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 376, 2109–2121. [Google Scholar] [CrossRef] [PubMed]
- Melis, J.P.; Wijnhoven, S.W.; Beems, R.B.; Roodbergen, M.; van den Berg, J.; Moon, H.; Friedberg, E.; van der Horst, G.T.; Hoeijmakers, J.H.; Vijg, J.; et al. Mouse models for xeroderma pigmentosum group A and group C show divergent cancer phenotypes. Cancer Res. 2008, 68, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Melis, J.P.; Speksnijder, E.N.; Kuiper, R.V.; Salvatori, D.C.; Schaap, M.M.; Maas, S.; Robinson, J.; Verhoef, A.; van Benthem, J.; Luijten, M.; et al. Detection of genotoxic and non-genotoxic carcinogens in Xpc(−/−)p53(+/−) mice. Toxicol. Appl. Pharmacol. 2013, 266, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, H.R.; Kim, A.L.; Rossignol, R.; Ali, N.; Daly, M.; Mahfouf, W.; Bellance, N.; Taieb, A.; de Verneuil, H.; Mazurier, F.; et al. XPC silencing in normal human keratinocytes triggers metabolic alterations that drive the formation of squamous cell carcinomas. J. Clin. Investig. 2011, 121, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, M.A.; Rajab, N.F.; Chua, E.W.; Selvarajah, G.T.; Masre, S.F. NTCU induced pre-malignant and malignant stages of lung squamous cell carcinoma in mice model. Sci. Rep. 2021, 11, 22500. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.A. Formation, repair, and genotoxic properties of bulky DNA adducts formed from tobacco-specific nitrosamines. J. Nucleic Acids 2010, 2010, 284935. [Google Scholar] [CrossRef] [PubMed]
- Haines, D.C.; Chattopadhyay, S.; Ward, J.M. Pathology of Aging B6;129 Mice. Toxicol. Pathol. 2001, 29, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Ide, F.; Iida, N.; Nakatsuru, Y.; Oda, H.; Tanaka, K.; Ishikawa, T. Mice deficient in the nucleotide excision repair gene XPA have elevated sensitivity to benzo[a]pyrene induction of lung tumors. Carcinogenesis 2000, 21, 1263–1265. [Google Scholar]
- Friedberg, E.C.; Bond, J.P.; Burns, D.K.; Cheo, D.L.; Greenblatt, M.S.; Meira, L.B.; Nahari, D.; Reis, A.M.C. Defective nucleotide excision repair in XPC mutant mice and its association with cancer predisposition. Mutat. Res. 2000, 459, 99–108. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Deaths/Sac Prior to 19.5 Weeks (Total Mice) | Mice with LUSC/Total * | Mice with LUSC or High-Grade Dysplasia/Total * | Other Findings |
|---|---|---|---|---|---|
| XPC +/+ | Acetone (control) | 0 (13) | 0/13 | 0/13 | |
| XPC +/+ | NTCU | 11 (37) Death: 3 Sac: 8 Δ | 5/26 M: 3/17 F: 2/9 | 8/26 M: 5/17 F: 4/9 | Granuloma (1), early lymphoma (2) |
| XPC +/− | Acetone (control) | 0 (8) | 0/8 | 0/8 | |
| XPC +/− | NTCU | 5 (17) Death: 5 | 4/12 M: 2/4 F: 2/8 | 5/12 M: 2/4 F: 3/8 | Early lymphoma (2) |
| XPC −/− | Acetone (control) | 0 (11) | 0/11 | 0/11 | |
| XPC −/− | NTCU | 12 (40) Death: 5 Sac: 7 Δ | 13/28 M: 6/12 F: 7/16 | 16/28 M: 7/12 F: 9/16 | Early lymphoma (1), Granulomas (3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sears, C.R.; Zhou, H.; Hulsey, E.; Aidoo, B.A.; Sandusky, G.E.; Al Nasrallah, N. XPC Protects against Carcinogen-Induced Histologic Progression to Lung Squamous Cell Carcinoma by Reduced Basal Epithelial Cell Proliferation. Cancers 2024, 16, 1495. https://doi.org/10.3390/cancers16081495
Sears CR, Zhou H, Hulsey E, Aidoo BA, Sandusky GE, Al Nasrallah N. XPC Protects against Carcinogen-Induced Histologic Progression to Lung Squamous Cell Carcinoma by Reduced Basal Epithelial Cell Proliferation. Cancers. 2024; 16(8):1495. https://doi.org/10.3390/cancers16081495
Chicago/Turabian StyleSears, Catherine R., Huaxin Zhou, Emily Hulsey, Bea A. Aidoo, George E. Sandusky, and Nawar Al Nasrallah. 2024. "XPC Protects against Carcinogen-Induced Histologic Progression to Lung Squamous Cell Carcinoma by Reduced Basal Epithelial Cell Proliferation" Cancers 16, no. 8: 1495. https://doi.org/10.3390/cancers16081495
APA StyleSears, C. R., Zhou, H., Hulsey, E., Aidoo, B. A., Sandusky, G. E., & Al Nasrallah, N. (2024). XPC Protects against Carcinogen-Induced Histologic Progression to Lung Squamous Cell Carcinoma by Reduced Basal Epithelial Cell Proliferation. Cancers, 16(8), 1495. https://doi.org/10.3390/cancers16081495