
LCMR1 Promotes Large-Cell Lung Cancer Proliferation and Metastasis by Downregulating HLA-Encoding Genes
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culturing
2.2. Delivery of Short Hairpin (sh) RNA Using Lentivirus
2.3. Immunofluorescence Staining
2.4. Proliferation Assay
2.5. Migration Assays
2.5.1. Wound Healing Assay
2.5.2. Transwell Migration and Invasion Assay
2.6. Cell Cycle Analysis
2.7. In Vivo Animal Assays
2.8. Protein Extraction and Immunoblotting
2.9. RNA Extraction and Quantitative PCR
2.10. Transcriptome Sequencing and Data Analysis
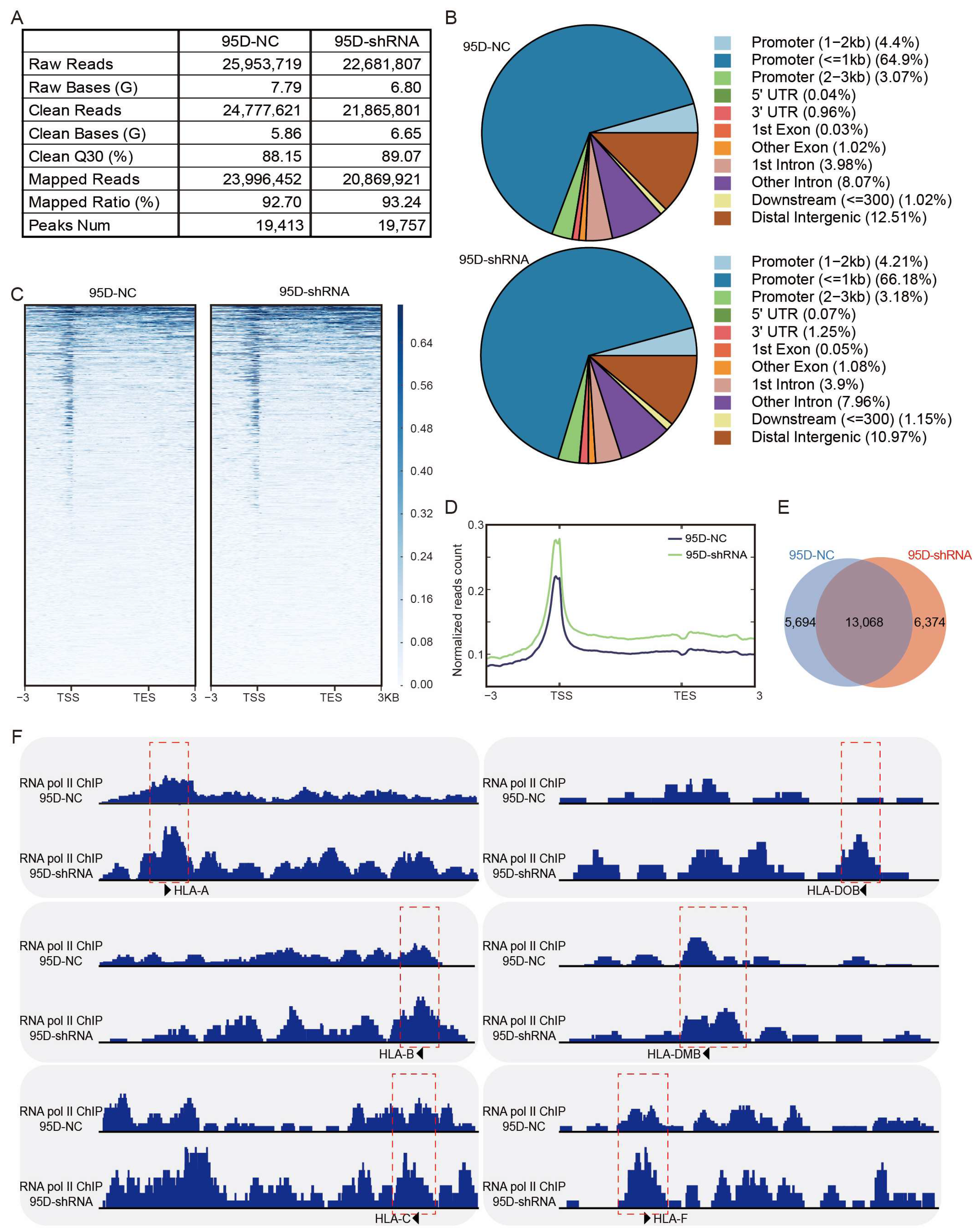
2.11. ChIP-Seq and Data Processing
2.12. Data Availability
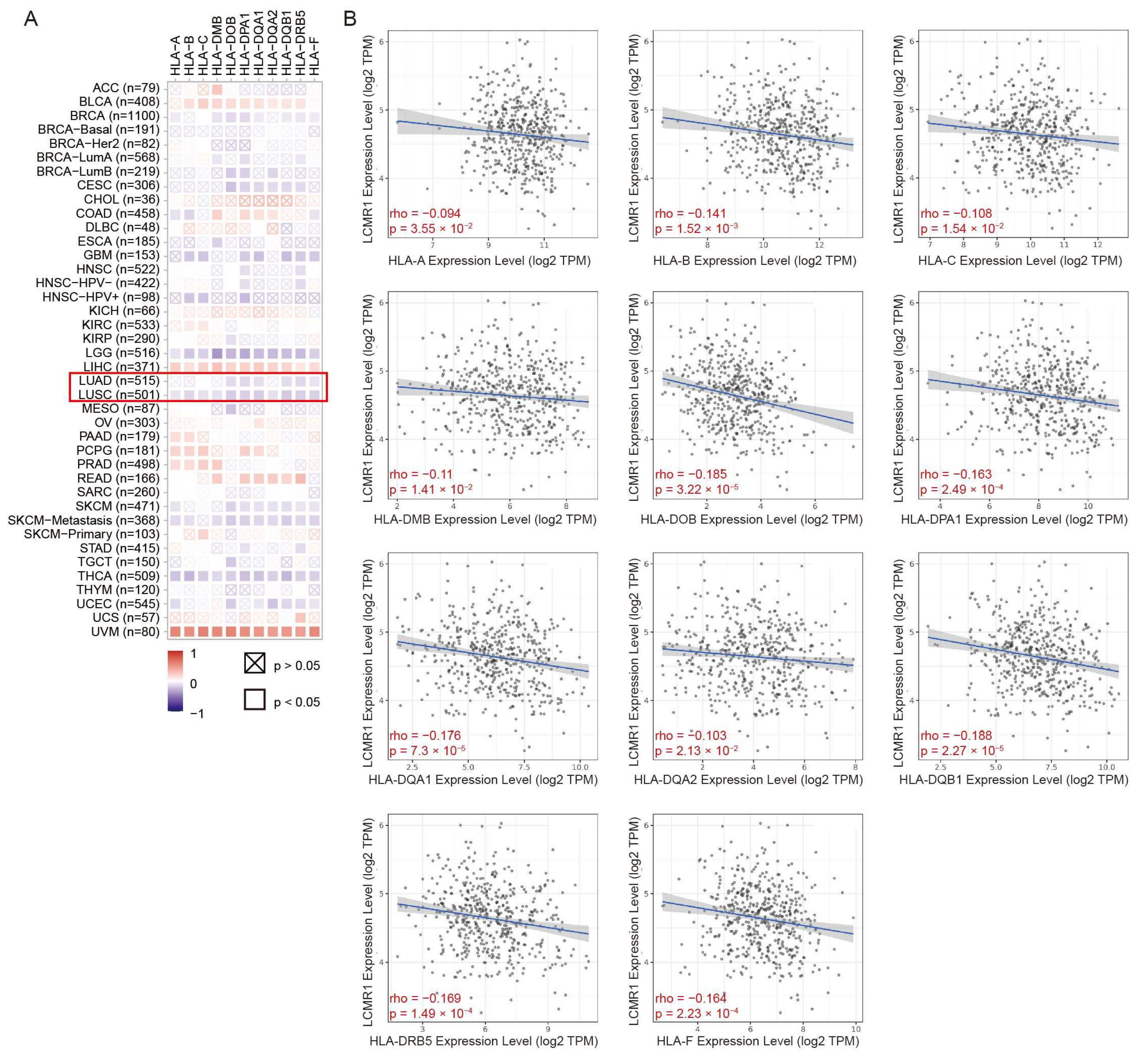
2.13. Pan-Cancer Analysis
2.14. Statistical Analysis
3. Results
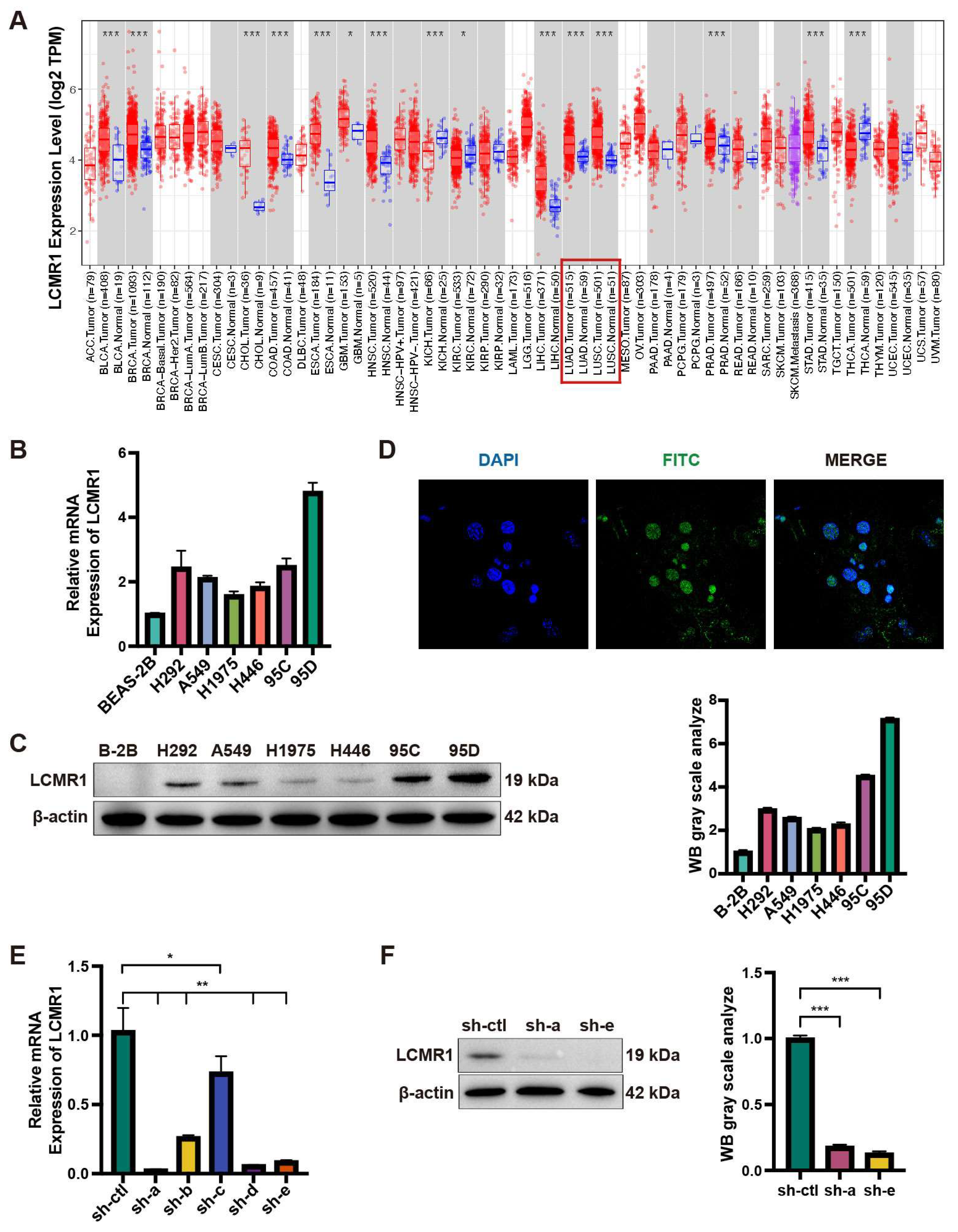
3.1. LCMR1 Is Highly Expressed in LCLC Cell Lines
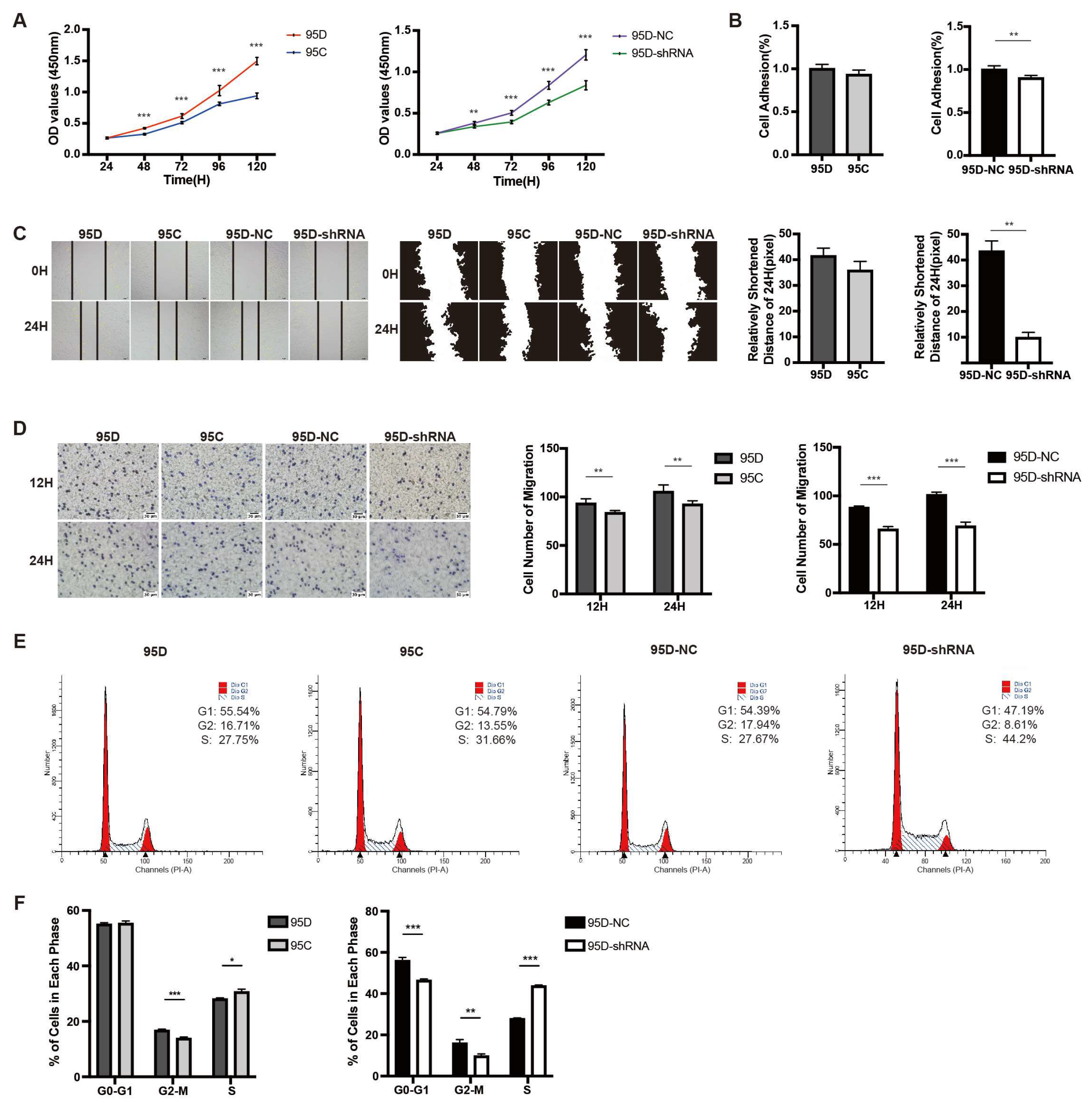
3.2. LCMR1 Contributes to LCLC Growth and Invasion In Vitro
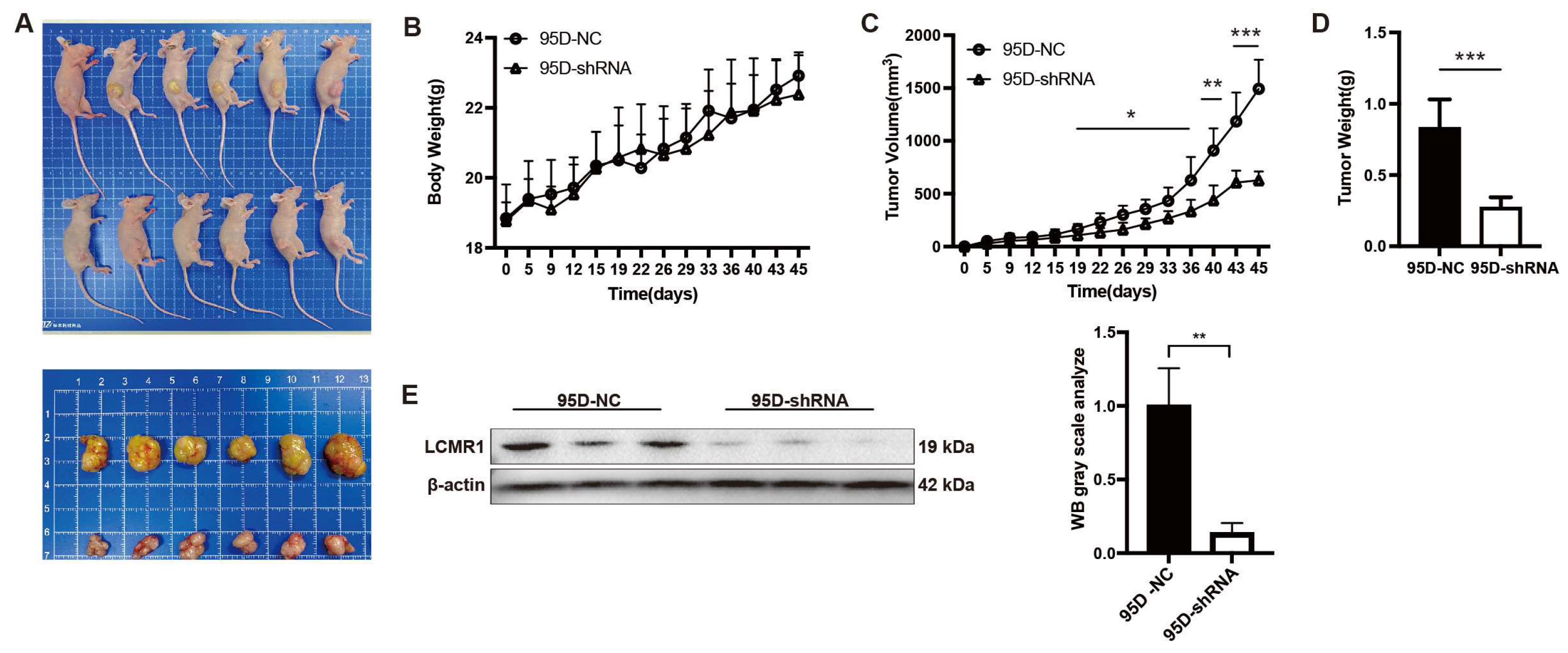
3.3. Knocking down LCMR1 Inhibits In Vivo Tumor Growth of 95D Xenografts Model
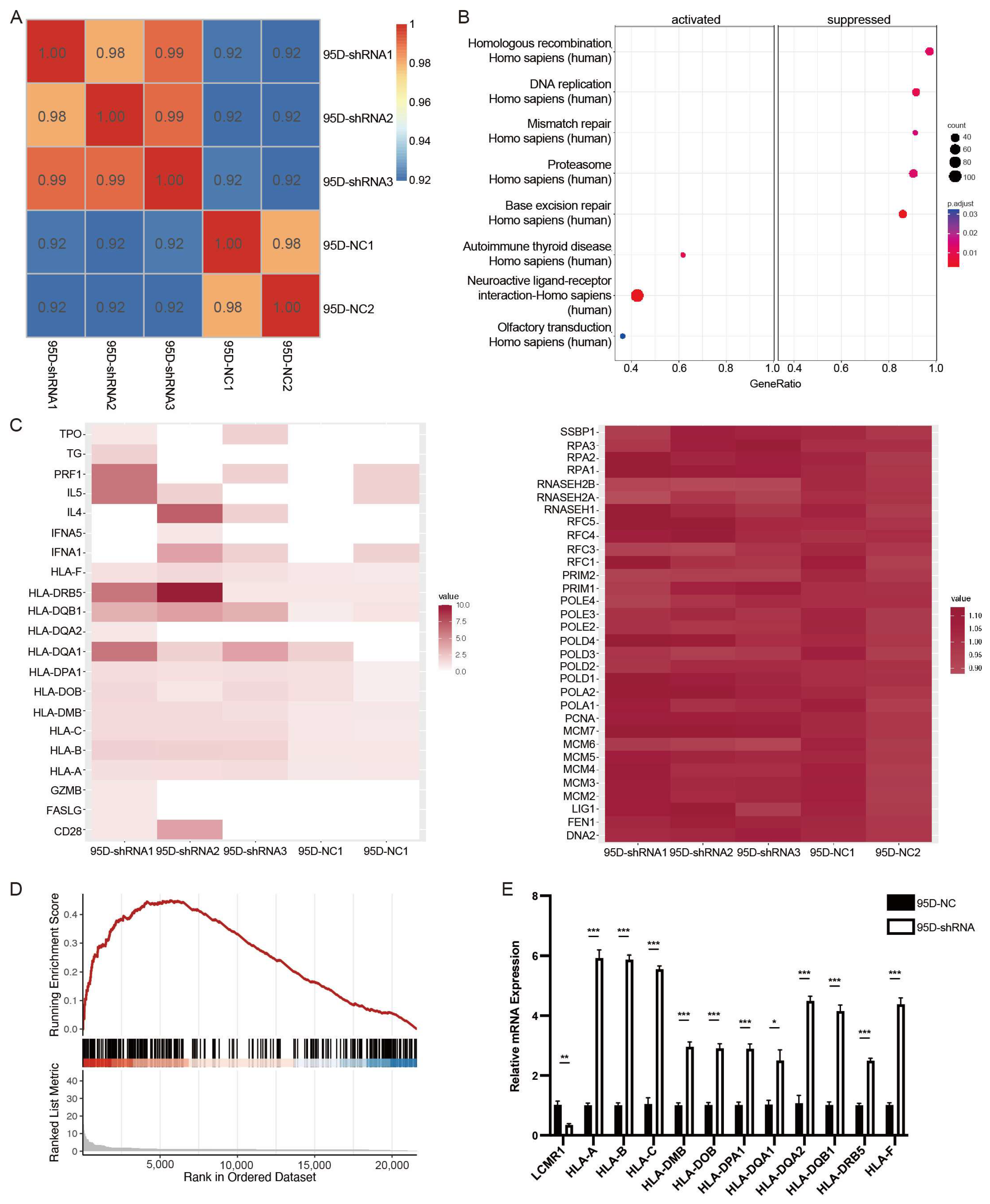
3.4. LCMR1 Negatively Regulates Human Leukocyte Antigen (HLA) Coding Genes
3.5. LCMR1 Diminishes the RNA Pol II Occupancy at the Promoter of HLA-Encoding Genes
3.6. The Expression Level of LCMR1 Negatively Correlates with HLA-Encoding Genes in NSCLC Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer treatment and survivorship statistics, 2022. CA Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Roy-Chowdhuri, S. Molecular Pathology of Lung Cancer. Surg. Pathol. Clin. 2021, 14, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liang, Z.; Tian, Q.; Li, C.; Ma, X.; Zhang, Y.; Yang, Z.; Wang, P.; Li, Y. Overexpression of LCMR1 is significantly associated with clinical stage in human NSCLC. J. Exp. Clin. Cancer Res. 2011, 30, 1756–9966. [Google Scholar] [CrossRef]
- Soutourina, J. Transcription regulation by the Mediator complex. Nat. Rev. Mol. Cell Biol. 2018, 19, 262–274. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, Y.; Liu, B.; Qi, X.; Guo, Z.; Li, L. Med19 promotes breast cancer cell proliferation by regulating CBFA2T3/HEB expression. Breast Cancer 2017, 24, 433–441. [Google Scholar] [CrossRef]
- Weber, H.; Ruoff, R.; Garabedian, M.J. MED19 alters AR occupancy and gene expression in prostate cancer cells, driving MAOA expression and growth under low androgen. PLoS Genet. 2021, 17, e1008540. [Google Scholar] [CrossRef]
- Ding, X.F.; Huang, G.M.; Shi, Y.; Li, J.A.; Fang, X.D. Med19 promotes gastric cancer progression and cellular growth. Gene 2012, 504, 262–267. [Google Scholar] [CrossRef]
- Zhu, L.J.; Yan, W.X.; Chen, Z.W.; Chen, Y.; Chen, D.; Zhang, T.H.; Liao, G.Q. Disruption of mediator complex subunit 19 (Med19) inhibits cell growth and migration in tongue cancer. World J. Surg. Oncol. 2013, 11, 1477–7819. [Google Scholar] [CrossRef]
- Xu, L.L.; Guo, S.L.; Ma, S.R.; Luo, Y.A. Mammalian mediator 19 mediates H1299 lung adenocarcinoma cell clone conformation, growth, and metastasis. Asian Pac. J. Cancer Prev. 2012, 13, 3695–3700. [Google Scholar] [CrossRef]
- Sun, M.; Jiang, R.; Li, J.D.; Luo, S.L.; Gao, H.W.; Jin, C.Y.; Shi, D.L.; Wang, C.G.; Wang, B.; Zhang, X.Y. MED19 promotes proliferation and tumorigenesis of lung cancer. Mol. Cell Biochem. 2011, 355, 27–33. [Google Scholar] [CrossRef]
- Wei, L.; Wang, X.W.; Sun, J.J.; Lv, L.Y.; Xie, L.; Song, X.R. Knockdown of Med19 suppresses proliferation and enhances chemo-sensitivity to cisplatin in non-small cell lung cancer cells. Asian Pac. J. Cancer Prev. 2015, 16, 875–880. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, Y.; Li, C.; Tian, Q.; Li, Y.; Yang, Z.; Liang, Z.; Chen, L. Suppression of lung cancer metastasis-related protein 1 promotes apoptosis in lung cancer cells. Int. J. Mol. Med. 2012, 30, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liang, Z.; Li, C.; Yang, Z.; Chen, L. LCMR1 interacts with DEK to suppress apoptosis in lung cancer cells. Mol. Med. Rep. 2017, 16, 4159–4164. [Google Scholar] [CrossRef] [PubMed]
- Chaulet, H.; Desgranges, C.; Renault, M.A.; Dupuch, F.; Ezan, G.; Peiretti, F.; Loirand, G.; Pacaud, P.; Gadeau, A.P. Extracellular nucleotides induce arterial smooth muscle cell migration via osteopontin. Circ. Res. 2001, 89, 772–778. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, 2013–2014. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Kaya-Okur, H.S.; Wu, S.J.; Codomo, C.A.; Pledger, E.S.; Bryson, T.D.; Henikoff, J.G.; Ahmad, K.; Henikoff, S. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 2019, 10, 1930. [Google Scholar]
- Nakato, R.; Sakata, T. Methods for ChIP-seq analysis: A practical workflow and advanced applications. Methods 2021, 187, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, 2008–2009. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, F.; Dündar, F.; Diehl, S.; Grüning, B.A.; Manke, T. deepTools: A flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 2014, 42, W187–W191. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; He, Q.Y. ChIPseeker: An R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Baidoobonso, S.M.; Guidi, B.W.; Myers, L.C. Med19(Rox3) regulates Intermodule interactions in the Saccharomyces cerevisiae mediator complex. J. Biol. Chem. 2007, 282, 5551–5559. [Google Scholar] [CrossRef]
- Koukourakis, I.M.; Giatromanolaki, A.; Mitrakas, A.; Koukourakis, M.I. Loss of HLA-class-I expression in non-small-cell lung cancer: Association with prognosis and anaerobic metabolism. Cell. Immunol. 2022, 373, 104495. [Google Scholar] [CrossRef]
- McGranahan, N.; Rosenthal, R.; Hiley, C.T.; Rowan, A.J.; Watkins, T.B.K.; Wilson, G.A.; Birkbak, N.J.; Veeriah, S.; Van Loo, P.; Herrero, J.; et al. Allele-Specific HLA Loss and Immune Escape in Lung Cancer Evolution. Cell 2017, 171, 1259–1271. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Health Organization Classification of Tumours, Volume 5: Thoracic Tumours, 5th ed.; IARC Press: Lyon, France, 2021; Available online: https://tumourclassification.iarc.who.int/ (accessed on 22 November 2022).
- Liang, Z.; Wang, W.; Hu, Q.; Zhou, P.; Zhang, Y.; Tang, Y.; Wu, Q.; Fu, Y.; Li, X.; Shao, Y.; et al. Pulmonary large cell carcinoma with neuroendocrine morphology shows genetic similarity to large cell neuroendocrine carcinoma. Diagn. Pathol. 2022, 17, 26. [Google Scholar] [CrossRef] [PubMed]
- Rekhtman, N.; Tafe, L.J.; Chaft, J.E.; Wang, L.; Arcila, M.E.; Colanta, A.; Moreira, A.L.; Zakowski, M.F.; Travis, W.D.; Sima, C.S.; et al. Distinct profile of driver mutations and clinical features in immunomarker-defined subsets of pulmonary large-cell carcinoma. Mod. Pathol. 2013, 26, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Rekhtman, N.; Travis, W.D. Large No More: The Journey of Pulmonary Large Cell Carcinoma from Common to Rare Entity. J. Thorac. Oncol. 2019, 14, 1125–1127. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Y.; Wang, L.; Liu, H. Role of RACK1 on cell proliferation, adhesion, and bortezomib-induced apoptosis in multiple myeloma. Int. J. Biol. Macromol. 2019, 121, 1077–1085. [Google Scholar] [CrossRef]
- Hsiao, Y.T.; Fan, M.J.; Huang, A.C.; Lien, J.C.; Lin, J.J.; Chen, J.C.; Hsia, T.C.; Wu, R.S.; Chung, J.G. Deguelin Impairs Cell Adhesion, Migration and Invasion of Human Lung Cancer Cells through the NF-κB Signaling Pathways. Am. J. Chin. Med. 2018, 46, 209–229. [Google Scholar] [CrossRef]
- Jhunjhunwala, S.; Hammer, C.; Delamarre, L. Antigen presentation in cancer: Insights into tumour immunogenicity and immune evasion. Nat. Rev. Cancer 2021, 21, 298–312. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, J.; Hu, C.; Zheng, X.; Guo, Z.; Li, L. CXCL11 negatively regulated by MED19 favours antitumour immune infiltration in breast cancer. Cytokine 2023, 162, 156106. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Li, C.; Wu, Z.; Li, Y.; Yu, H.; Li, T.; Wang, Y.; Zhao, W.; Chen, L. LCMR1 Promotes Large-Cell Lung Cancer Proliferation and Metastasis by Downregulating HLA-Encoding Genes. Cancers 2023, 15, 5445. https://doi.org/10.3390/cancers15225445
Liu L, Li C, Wu Z, Li Y, Yu H, Li T, Wang Y, Zhao W, Chen L. LCMR1 Promotes Large-Cell Lung Cancer Proliferation and Metastasis by Downregulating HLA-Encoding Genes. Cancers. 2023; 15(22):5445. https://doi.org/10.3390/cancers15225445
Chicago/Turabian StyleLiu, Lu, Chunsun Li, Zhen Wu, Yanqin Li, Hang Yu, Tao Li, Yueming Wang, Wei Zhao, and Liangan Chen. 2023. "LCMR1 Promotes Large-Cell Lung Cancer Proliferation and Metastasis by Downregulating HLA-Encoding Genes" Cancers 15, no. 22: 5445. https://doi.org/10.3390/cancers15225445
APA StyleLiu, L., Li, C., Wu, Z., Li, Y., Yu, H., Li, T., Wang, Y., Zhao, W., & Chen, L. (2023). LCMR1 Promotes Large-Cell Lung Cancer Proliferation and Metastasis by Downregulating HLA-Encoding Genes. Cancers, 15(22), 5445. https://doi.org/10.3390/cancers15225445