
Decoding the Complexity of Immune–Cancer Cell Interactions: Empowering the Future of Cancer Immunotherapy

Abstract
:Simple Summary
Abstract
1. Introduction
2. Cell–Cell Interactions during Tumorigenesis
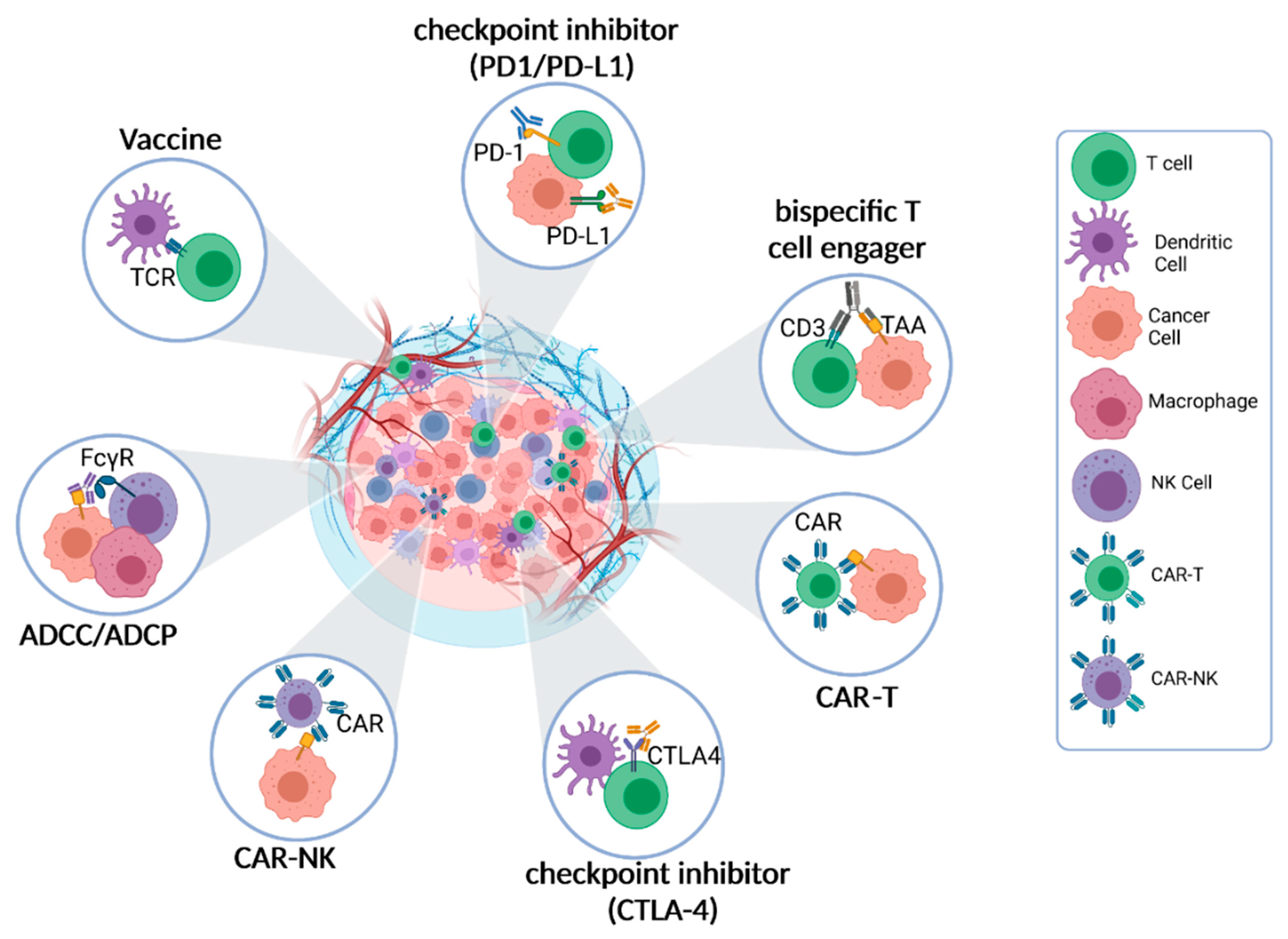
3. Cell–Cell Interactions Are the Pharmacological Basis of Immunotherapy
4. Experimental and Modeling Systems for Studying Cell–Cell Interactions
5. Proximity-Based Labeling Approaches for Studying Cell–Cell Interactions
6. Bioinformatic Techniques for Inferring Cell–Cell Interactions
7. Potential Questions to Be Addressed by These Approaches
8. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Armingol, E.; Officer, A.; Harismendy, O.; Lewis, N.E. Deciphering cell-cell interactions and communication from gene expression. Nat. Rev. Genet. 2021, 22, 71–88. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Eno, J. Immunotherapy through the years. J. Adv. Pract. Oncol. 2017, 8, 747–753. [Google Scholar]
- Ma, W.; Gilligan, B.M.; Yuan, J.; Li, T. Current status and perspectives in translational biomarker research for PD-1/PD-L1 immune checkpoint blockade therapy. J. Hematol. Oncol. 2016, 9, 47. [Google Scholar] [CrossRef]
- Lanitis, E.; Dangaj, D.; Irving, M.; Coukos, G. Mechanisms regulating T-cell infiltration and activity in solid tumors. Ann. Oncol. 2017, 28, xii18–xii32. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.; Elkord, E. FoxP3+ T regulatory cells in cancer: Prognostic biomarkers and therapeutic targets. Cancer Lett. 2020, 490, 174–185. [Google Scholar] [CrossRef]
- Blessin, N.C.; Li, W.; Mandelkow, T.; Jansen, H.L.; Yang, C.; Raedler, J.B.; Simon, R.; Büscheck, F.; Dum, D.; Luebke, A.M.; et al. Prognostic role of proliferating CD8+ cytotoxic Tcells in human cancers. Cell. Oncol. 2021, 44, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Wei, M.; Wang, S.; Dong, J.; Wei, J. Pan-Cancer Analysis of Immune Cell Infiltration Identifies a Prognostic Immune-Cell Characteristic Score (ICCS) in Lung Adenocarcinoma. Front. Immunol. 2020, 11, 1218. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Teng, M.W.L.; Smyth, M.J. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat. Rev. Clin. Oncol. 2019, 16, 151–167. [Google Scholar] [CrossRef]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef]
- Liu, T.; Han, C.; Wang, S.; Fang, P.; Ma, Z.; Xu, L.; Yin, R. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 86. [Google Scholar] [CrossRef]
- Hanley, C.J.; Thomas, G.J. Targeting cancer associated fibroblasts to enhance immunotherapy: Emerging strategies and future perspectives. Oncotarget 2021, 12, 1427–1433. [Google Scholar] [CrossRef]
- Pei, L.; Liu, Y.; Liu, L.; Gao, S.; Gao, X.; Feng, Y.; Sun, Z.; Zhang, Y.; Wang, C. Roles of cancer-associated fibroblasts (CAFs) in anti-PD-1/PD-L1 immunotherapy for solid cancers. Mol. Cancer 2023, 22, 29. [Google Scholar] [CrossRef]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z. The history and advances in cancer immunotherapy: Understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef]
- Gómez Román, V.R.; Murray, J.C.; Weiner, L.M. Antibody-Dependent Cellular Cytotoxicity (ADCC). In Antibody Fc; Academic Press: Cambridge, MA, USA, 2014; pp. 1–27. [Google Scholar]
- Zhou, S.; Liu, M.; Ren, F.; Meng, X.; Yu, J. The landscape of bispecific T cell engager in cancer treatment. Biomark. Res. 2021, 9, 38. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, J.; Kudlacek, S.; Qi, T.; Dunlap, T.; Cao, Y. Population dynamics of immunological synapse formation induced by bispecific T cell engagers predict clinical pharmacodynamics and treatment resistance. eLife 2023, 12, e83659. [Google Scholar] [CrossRef]
- Pan, K.; Farrukh, H.; Chittepu, V.C.S.R.; Xu, H.; Pan, C.-X.; Zhu, Z. CAR race to cancer immunotherapy: From CAR T, CAR NK to CAR macrophage therapy. J. Exp. Clin. Cancer Res. 2022, 41, 119. [Google Scholar] [CrossRef]
- Shalhout, S.Z.; Miller, D.M.; Emerick, K.S.; Kaufman, H.L. Therapy with oncolytic viruses: Progress and challenges. Nat. Rev. Clin. Oncol. 2023, 20, 160–177. [Google Scholar] [CrossRef]
- Cao, G.-D.; He, X.-B.; Sun, Q.; Chen, S.; Wan, K.; Xu, X.; Feng, X.; Li, P.-P.; Chen, B.; Xiong, M.-M. The oncolytic virus in cancer diagnosis and treatment. Front. Oncol. 2020, 10, 1786. [Google Scholar] [CrossRef]
- Dominiak, A.; Chełstowska, B.; Olejarz, W.; Nowicka, G. Communication in the cancer microenvironment as a target for therapeutic interventions. Cancers 2020, 12, 1232. [Google Scholar] [CrossRef] [PubMed]
- Shelton, S.E.; Nguyen, H.T.; Barbie, D.A.; Kamm, R.D. Engineering approaches for studying immune-tumor cell interactions and immunotherapy. iScience 2021, 24, 101985. [Google Scholar] [CrossRef] [PubMed]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef]
- Yuki, K.; Cheng, N.; Nakano, M.; Kuo, C.J. Organoid models of tumor immunology. Trends Immunol. 2020, 41, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Białkowska, K.; Komorowski, P.; Bryszewska, M.; Miłowska, K. Spheroids as a Type of Three-Dimensional Cell Cultures-Examples of Methods of Preparation and the Most Important Application. Int. J. Mol. Sci. 2020, 21, 6225. [Google Scholar] [CrossRef]
- Mao, Y.; Wang, X.; Huang, P.; Tian, R. Spatial proteomics for understanding the tissue microenvironment. Analyst 2021, 146, 3777–3798. [Google Scholar] [CrossRef]
- Zhong, W.; Myers, J.S.; Wang, F.; Wang, K.; Lucas, J.; Rosfjord, E.; Lucas, J.; Hooper, A.T.; Yang, S.; Lemon, L.A.; et al. Comparison of the molecular and cellular phenotypes of common mouse syngeneic models with human tumors. BMC Genom. 2020, 21, 2. [Google Scholar] [CrossRef]
- Taylor, M.A.; Hughes, A.M.; Walton, J.; Coenen-Stass, A.M.L.; Magiera, L.; Mooney, L.; Bell, S.; Staniszewska, A.D.; Sandin, L.C.; Barry, S.T.; et al. Longitudinal immune characterization of syngeneic tumor models to enable model selection for immune oncology drug discovery. J. Immunother. Cancer 2019, 7, 328. [Google Scholar] [CrossRef]
- Richmond, A.; Su, Y. Mouse xenograft models vs GEM models for human cancer therapeutics. Dis. Model. Mech. 2008, 1, 78–82. [Google Scholar] [CrossRef]
- Mestas, J.; Hughes, C.C.W. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef]
- Arnol, D.; Schapiro, D.; Bodenmiller, B.; Saez-Rodriguez, J.; Stegle, O. Modeling Cell-Cell Interactions from Spatial Molecular Data with Spatial Variance Component Analysis. Cell Rep. 2019, 29, 202–211.e6. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Chen, L.; Liu, S.; Zhao, J.; Zhang, H.; Chen, P.R. Enzyme-Mediated Intercellular Proximity Labeling for Detecting Cell-Cell Interactions. J. Am. Chem. Soc. 2019, 141, 1833–1837. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Murray, C.W.; Linde, I.L.; Kramer, N.J.; Lyu, Z.; Tsai, M.K.; Chen, L.C.; Cai, H.; Gitler, A.D.; Engleman, E.; et al. A versatile system to record cell-cell interactions. eLife 2020, 9, e61080. [Google Scholar] [CrossRef] [PubMed]
- Pasqual, G.; Chudnovskiy, A.; Tas, J.M.J.; Agudelo, M.; Schweitzer, L.D.; Cui, A.; Hacohen, N.; Victora, G.D. Monitoring T cell-dendritic cell interactions in vivo by intercellular enzymatic labelling. Nature 2018, 553, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, J.P.; Chen, M.; Wu, M.; Shi, Y.; Li, W.; Teijaro, J.R.; Wu, P. Detecting Tumor Antigen-Specific T Cells via Interaction-Dependent Fucosyl-Biotinylation. Cell 2020, 183, 1117–1133.e19. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Zhao, Z.; Wu, M.; Xue, Q.; Yang, Y.; Ouyang, S.; Li, W.; Zhong, L.; Wang, W.; Yang, R.; et al. Use of intercellular proximity labeling to quantify and decipher cell-cell interactions directed by diversified molecular pairs. Cancer 2022, 8, eadd2337. [Google Scholar] [CrossRef]
- Liu, Q.; Zheng, J.; Sun, W.; Huo, Y.; Zhang, L.; Hao, P.; Wang, H.; Zhuang, M. A proximity-tagging system to identify membrane protein-protein interactions. Nat. Methods 2018, 15, 715–722. [Google Scholar] [CrossRef]
- Patel, S.J.; Sanjana, N.E.; Kishton, R.J.; Eidizadeh, A.; Vodnala, S.K.; Cam, M.; Gartner, J.J.; Jia, L.; Steinberg, S.M.; Yamamoto, T.N.; et al. Identification of essential genes for cancer immunotherapy. Nature 2017, 548, 537–542. [Google Scholar] [CrossRef]
- Cho, K.F.; Gillespie, S.M.; Kalogriopoulos, N.A.; Quezada, M.A.; Jacko, M.; Monje, M.; Ting, A.Y. A light-gated transcriptional recorder for detecting cell-cell contacts. eLife 2022, 11, e70881. [Google Scholar] [CrossRef]
- Ombrato, L.; Nolan, E.; Kurelac, I.; Mavousian, A.; Bridgeman, V.L.; Heinze, I.; Chakravarty, P.; Horswell, S.; Gonzalez-Gualda, E.; Matacchione, G.; et al. Metastatic-niche labelling reveals parenchymal cells with stem features. Nature 2019, 572, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, W.B.; Jones, K.A.; McCutcheon, D.C.; Prescher, J.A. A “Caged” Luciferin for Imaging Cell-Cell Contacts. J. Am. Chem. Soc. 2015, 137, 8656–8659. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, Y.; Nystrom, N.N.; Liu, S.; Fu, Y.; Martinez, F.M.; Scholl, T.J.; Ronald, J.A. Visualizing cell-cell communication using synthetic notch activated MRI. Proc. Natl. Acad. Sci. USA 2023, 120, e2216901120. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Q.; Liu, Z.; Liu, K.; He, L.; Lui, K.O.; Wang, L.; Zhou, B. Genetic dissection of intercellular interactions in vivo by membrane-permeable protein. Proc. Natl. Acad. Sci. USA 2023, 120, e2120582120. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cohen, J.; Song, X.; Zhao, A.; Ye, Z.; Feulner, C.J.; Doonan, P.; Somers, W.; Lin, L.; Chen, P.R. Improved variants of SrtA for site-specific conjugation on antibodies and proteins with high efficiency. Sci. Rep. 2016, 6, 31899. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Liao, J.; Li, C.; Lu, X.; Cheng, J.; Fan, X. CellTalkDB: A manually curated database of ligand-receptor interactions in humans and mice. Brief. Bioinform. 2021, 22, bbaa269. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Zhang, S.; Song, S.; Jiang, C.; Han, G.; Wang, M.; Ajani, J.; Futreal, A.; Wang, L. iTALK: An R Package to Characterize and Illustrate Intercellular Communication. BioRxiv 2019, 507871. [Google Scholar] [CrossRef]
- Tyler, S.R.; Rotti, P.G.; Sun, X.; Yi, Y.; Xie, W.; Winter, M.C.; Flamme-Wiese, M.J.; Tucker, B.A.; Mullins, R.F.; Norris, A.W.; et al. PyMINEr Finds Gene and Autocrine-Paracrine Networks from Human Islet scRNA-Seq. Cell Rep. 2019, 26, 1951–1964.e8. [Google Scholar] [CrossRef]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.-H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef]
- Efremova, M.; Vento-Tormo, M.; Teichmann, S.A.; Vento-Tormo, R. CellPhoneDB: Inferring cell-cell communication from combined expression of multi-subunit ligand-receptor complexes. Nat. Protoc. 2020, 15, 1484–1506. [Google Scholar] [CrossRef]
- Dries, R.; Zhu, Q.; Dong, R.; Eng, C.-H.L.; Li, H.; Liu, K.; Fu, Y.; Zhao, T.; Sarkar, A.; Bao, F.; et al. Giotto: A toolbox for integrative analysis and visualization of spatial expression data. Genome Biol. 2021, 22, 78. [Google Scholar] [CrossRef] [PubMed]
- Noël, F.; Massenet-Regad, L.; Carmi-Levy, I.; Cappuccio, A.; Grandclaudon, M.; Trichot, C.; Kieffer, Y.; Mechta-Grigoriou, F.; Soumelis, V. ICELLNET: A transcriptome-based framework to dissect intercellular communication. BioRxiv 2020. [Google Scholar] [CrossRef]
- Cabello-Aguilar, S.; Alame, M.; Kon-Sun-Tack, F.; Fau, C.; Lacroix, M.; Colinge, J. SingleCellSignalR: Inference of intercellular networks from single-cell transcriptomics. Nucleic Acids Res. 2020, 48, e55. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Sheng, J.; Gao, D.; Li, F.; Durrans, A.; Ryu, S.; Lee, S.B.; Narula, N.; Rafii, S.; Elemento, O.; et al. Transcriptome analysis of individual stromal cell populations identifies stroma-tumor crosstalk in mouse lung cancer model. Cell Rep. 2015, 10, 1187–1201. [Google Scholar] [CrossRef] [PubMed]
- Browaeys, R.; Saelens, W.; Saeys, Y. NicheNet: Modeling intercellular communication by linking ligands to target genes. Nat. Methods 2020, 17, 159–162. [Google Scholar] [CrossRef]
- Wang, S.; Karikomi, M.; MacLean, A.L.; Nie, Q. Cell lineage and communication network inference via optimization for single-cell transcriptomics. Nucleic Acids Res. 2019, 47, e66. [Google Scholar] [CrossRef]
- Cang, Z.; Nie, Q. Inferring spatial and signaling relationships between cells from single cell transcriptomic data. Nat. Commun. 2020, 11, 2084. [Google Scholar] [CrossRef] [PubMed]
- Tsuyuzaki, K.; Ishii, M.; Nikaido, I. Uncovering hypergraphs of cell-cell interaction from single cell RNA-sequencing data. BioRxiv 2019, 566182. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Model System | Sample | Method Description | Reference |
|---|---|---|---|
| 2D cell culture | Cells | Cells grow in a monolayer if adherent or suspended in a culture flask. These cultures are a straightforward, cost-effective, and low-maintenance approach. Within the controlled environment, it is possible to investigate the interactions between different cell lines and observe their behavior and responses to treatments. | [24] |
| 3D cell culture | Cells | Cell growth and interactions occur in 3D space, where cells interact with their surrounding environment and neighboring cells. Two approaches: scaffold-based methods using hydrogels or structural scaffolds and scaffold-free techniques (spheroids). | [24] |
| Spheroids | Cells | Organoids, also known as multicellular spheroids, are self-assembled structures that mimic the physiological environment and interactions found in vivo. They provide a more physiologically relevant context, allowing the investigation of intercellular interactions and responses within a 3D microenvironment resembling in vivo conditions. | [25] |
| Organoids | Patient-derived cells and tissues | Primary patient-derived microtissues grown in a 3D extracellular matrix that represents in vivo physiology and genetic diversity, allowing the investigation of intercellular interactions and responses in a patient-specific manner. | [26] |
| Tissue Slices | Tumor Tissue | Tumor biopsy taken from patients or xenograft models, stained to assess tumor morphology and spatial location of cells. | [27] |
| Animal models | Tumor Tissue | Compatible with intravital and intercellular imaging/labeling techniques, as well as other genetic systems designed to detect cell–cell interactions upon contact or external stimulation, including UV or fluorescent light. | [28] |
| System | Scale | Application | Method | Reference |
|---|---|---|---|---|
| EXCELL | In vitro | Labeling Imaging | EXCELL (enzyme-mediated intercellular proximity labeling) is a method that utilizes a variant of SrtA, mgSrtA, to enable the non-specific labeling of cell surface proteins containing a monoglycine residue at the N-terminus. Unlike other methods, EXCELL does not require pre-engineering of acceptor cells and was applied in in vitro studies. | [33] |
| G-BaToN | In vitro In vivo Ex vivo | Labeling Imaging | G-BaToN is a versatile system for physical contact labeling between cells. Sender cells express surface-bound GFP, while receiver cells carry a synthetic element that selectively binds to GFP. Upon cell contact, GFP is transferred from sender to receiver cells, leading to fluorescence labeling of the receiver cells. This method requires pre-engineering of both sender and receiver cells and can be used for in vitro and ex vivo studies. | [34] |
| LIPSTIC | In vitro | Labeling | LIPSTIC (Labelling Immune Partnerships by SorTagging Intercellular Contacts) is a proximity-dependent labeling method that employs bacterial sortase (SrtA) to detect receptor–ligand interactions between cells. It involves the attachment of biotin to cell surface proteins, which can be detected using flow cytometry. LIPSTIC can be used in both in vitro and in vivo settings by pre-engineering the cells on both sides of the interaction. | [35] |
| FucoID | In vitro Ex vivo | Labeling | FucoID is a method for identifying antigen-specific T cells using interaction-dependent fucosyl biotinylation. This technique enables the isolation of endogenous tumor antigen T cells from tumor digests without prior knowledge of the tumor-specific antigens and has been used for ex vivo studies. | [36,37] |
| PUP-IT | In vitro | Labeling | PUP-IT (pupylation-based interaction tagging) is a method used to identify membrane protein interactions. In this approach, a small protein tag, Pup, is applied to proteins that interact with a PafA-fused bait, enabling transient and weak interactions to be enriched and detected by mass spectrometry. PUP-IT enables the identification and analysis of protein–protein interactions occurring at the membrane level. | [38] |
| 2CT-CRISPR | In vitro Ex vivo | Genetic influence | Two-cell type CRISPR assay. This assay can genetically manipulate T cells to interact with cancer cells ex vivo to determine the genes that influence T cell effector function on cancer cells. | [39] |
| TRACC | In vitro | Labeling Imaging | TRACC (Transcriptional Readout Activated by Cell–Cell Contacts) is a system that utilizes light gating to detect cell–cell contacts based on transcriptional activity (TF). Cells are engineered to express a light-responsive TF that regulates the expression of a reporter gene. When two cells come into contact, a light signal is applied to activate the TF, resulting in the activation of the reporter gene and subsequent detection of the cell–cell contact, monitoring cell–cell interactions in a controlled and dynamic manner. | [40] |
| Cherry-niche | In vivo | Labeling Imaging | Cherry-niche is an innovative method that allows cells expressing a fluorescent protein to selectively label their surrounding cells in the tumor niche. This technique involves generating cancer cells capable of transferring a liposoluble fluorescent protein to their neighboring cells within the tumor microenvironment. | [41] |
| Caged luciferins | In vitro In vivo | Imaging | Caged luciferins are utilized for bioluminescent activity-based sensing. Activator cells expressing β-galactosidase catalyze the cleavage of caged luciferin, known as Lugal, resulting in the release of D-luciferin. The liberated D-luciferin can then enter nearby reporter cells, where it serves as a substrate for the luciferase enzyme, leading to the production of light and allowing for the identification and visualization of cells that are in close proximity to the sender cells. | [42] |
| SynNotch-activated MRI | In vivo | Imaging | The SynNotch system is utilized to induce the expression of an MRI contrast agent in recipient cells when they interact with sender cells expressing the corresponding synthetic notch receptor, enabling the detection and visualization of cell–cell communication events in real time. | [43] |
| CLIP | In vivo | Labeling Imaging | CLIP (cre-induced intercellular labeling protein) secretes a membrane-permeable fluorescent protein (mCherry) from a donor cell that can mark neighboring receptor cells. This method can label both direct cell contact receptor cells and receptor cells at a close-range distance. | [44] |
| Platform | Data Source | Method | Reference |
|---|---|---|---|
| CellTalkDB | scRNA-seq | Manually curated database of ligand–receptor pairs from both human and mouse samples. | [46] |
| iTalk | scRNA-seq | Identifying and illustrating alterations in intercellular signaling network. R package made to analyze and visualize ligand–receptor pair. | [47] |
| PyMINeR | scRNA-seq | Python maximal information network exploration resource. Fully automates cell type-specific identification, and pathways as well as in silico detection of autocrine and paracrine signaling networks | [48] |
| CellChat | scRNA-seq | Open source R package that is able to visualize, analyze, and deduce intercellular communications from a data input. Uses mass action models and differential expression analysis to deduce cell state-specific signaling communications. Also provides visualization outputs to compare intercellular communication methods. | [49] |
| CellPhoneDB | scRNA-seq | Identifies biologically relevant interacting ligand–receptor pairs. Cells with the same cluster are pooled together as one cell state. Ligand–receptor interactions are derived based on the expression of a receptor of one state and a ligand of the other state. | [50] |
| Giotto | scRNA-seq | Open source spatial analysis platform that contains two modules, Giotto analyzer and Giotto viewer, which are both independent and fully integrated. Analyzer provides instructions about steps in analyzing single-cell expression data, and the viewer provides an interactive view of the data. | [51] |
| ICellNET | RNA-seq, scRNA-seq, and microarray | Transcriptomic-based framework that integrates a database of ligand–receptor interactions, communication scores, and connections of cell populations of interest with 31 human reference cell types and three visualization methods. | [52] |
| SingleCellSignalR | scRNA-seq | Open source R platform. Relies on a database of known ligand–receptor interactions called LRdb. | [53] |
| CCC Explorer | Transcriptome profiles | Java-based software. Uses a computational model to look at cell–cell communications ranging from ligand–receptor interactions to transcription factors and target genes. | [54] |
| NicheNet | Gene expression data | Open source R platform. Uses a database of ligand–receptor interactions to identify ligand–receptor interactions that could drive gene expression changes | [55] |
| SoptSC | RNA-seq | Similarity matrix-based optimization for single-cell data analysis. Uses a cell-to-cell similarity matrix via gene marker identification, lineage reference, clustering, and pseudo-temporal ordering. From this information, it predicts cell–cell communication networks. | [56] |
| SpaoTSC | scRNA-seq | Spatially optimal transporting of the single cells. The method has two major components: (1) constructing spatial metric for cells from scRNA-seq data and (2) reconstructing the cell–cell communication networks from the data and identifying relationships between genes from intercellular relationships. Uses python. | [57] |
| scTensor | scRNA-seq | Open source R package. Instead of looking at one-to-one cell–cell interactions, this software focuses on many-to-many cell–cell interactions. scTensor looks at a three-way relationship (hypergraph) between ligand expression, receptor expression, and ligand–receptor pairs. | [58] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maffuid, K.; Cao, Y. Decoding the Complexity of Immune–Cancer Cell Interactions: Empowering the Future of Cancer Immunotherapy. Cancers 2023, 15, 4188. https://doi.org/10.3390/cancers15164188
Maffuid K, Cao Y. Decoding the Complexity of Immune–Cancer Cell Interactions: Empowering the Future of Cancer Immunotherapy. Cancers. 2023; 15(16):4188. https://doi.org/10.3390/cancers15164188
Chicago/Turabian StyleMaffuid, Kaitlyn, and Yanguang Cao. 2023. "Decoding the Complexity of Immune–Cancer Cell Interactions: Empowering the Future of Cancer Immunotherapy" Cancers 15, no. 16: 4188. https://doi.org/10.3390/cancers15164188
APA StyleMaffuid, K., & Cao, Y. (2023). Decoding the Complexity of Immune–Cancer Cell Interactions: Empowering the Future of Cancer Immunotherapy. Cancers, 15(16), 4188. https://doi.org/10.3390/cancers15164188