
The Association between Serum Serine and Glycine and Related-Metabolites with Pancreatic Cancer in a Prospective Cohort Study

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Population
2.2. Case Ascertainment of Pancreatic Cancer
2.3. Nested Case-Control Study
2.4. Assessment of Serum Biomarkers
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | body mass index |
| CI | confidence interval |
| CV | coefficient of variation |
| eGFR | estimated glomerular filtration rate |
| ICD | International Classification of Diseases-Oncology |
| NF-kB | nuclear factor kappa-light-chain-enhancer of activated B cells or uclear factor kappa B |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| OR | odds ratio |
| PLP | pyridoxal 5′-phosphate |
| ROS | reactive oxygen species |
| SCH | Shanghai Cohort Study |
| SD | standard deviation |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Noone, A.; Howlader, N.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.; et al. SEER Cancer Statistics Review, 1975–2015, National Cancer Institute. Bethesda, MD. Available online: https://seer.cancer.gov/csr/1975_2015/ (accessed on 1 July 2021).
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Wu, X.; Guo, T.; Guan, N.; Liu, Y. Epidemiological Characteristics of Pancreatic Cancer in China From 1990 to 2019. Cancer Control 2021, 28, 10732748211051536. [Google Scholar] [CrossRef]
- Huang, J.; Lok, V.; Ngai, C.H.; Zhang, L.; Yuan, J.; Lao, X.Q.; Ng, K.; Chong, C.; Zheng, Z.-J.; Wong, M.C.S. Worldwide Burden of, Risk Factors for, and Trends in Pancreatic Cancer. Gastroenterology 2021, 160, 744–754. [Google Scholar] [CrossRef] [PubMed]
- World Cancer Research Fund. Diet, Nutrition, Physical Activity and Cancer: A Global Perspective; Continuous Update Project Expert Report 2018; World Cancer Research Fund: London, UK, 2018. [Google Scholar]
- Maisonneuve, P.; Lowenfels, A.B. Risk factors for pancreatic cancer: A summary review of meta-analytical studies. Int. J. Epidemiol. 2015, 44, 186–198. [Google Scholar] [CrossRef]
- Locasale, J.W. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.W.; Moskowitz, M. Arrest of cell growth in the G1 phase of the cell cycle by serine deprivation. Exp. Cell Res. 1978, 116, 127–137. [Google Scholar] [CrossRef]
- Davis, S.R.; Stacpoole, P.W.; Williamson, J.; Kick, L.S.; Quinlivan, E.P.; Coats, B.S.; Shane, B.; Bailey, L.B.; Gregory, J.F. Tracer-derived total and folate-dependent homocysteine remethylation and synthesis rates in humans indicate that serine is the main one-carbon donor. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E272–E279. [Google Scholar] [CrossRef]
- Fell, D.A.; Snell, K. Control analysis of mammalian serine biosynthesis. Feedback inhibition on the final step. Biochem. J. 1988, 256, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Snell, K. Enzymes of serine metabolism in normal, developing and neoplastic rat tissues. Adv. Enzym. Regul. 1984, 22, 325–400. [Google Scholar] [CrossRef]
- Snell, K.; Weber, G. Enzymic imbalance in serine metabolism in rat hepatomas. Biochem. J. 1986, 233, 617–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snell, K.; Natsumeda, Y.; Weber, G. The modulation of serine metabolism in hepatoma 3924A during different phases of cellular proliferation in culture. Biochem. J. 1987, 245, 609–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Vousden, K.H. Serine and one-carbon metabolism in cancer. Nat. Rev. Cancer 2016, 16, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Locasale, J.W.; Grassian, A.R.; Melman, T.; Lyssiotis, C.A.; Mattaini, K.R.; Bass, A.J.; Heffron, G.; Metallo, C.M.; Muranen, T.; Sharfi, H.; et al. Phosphoglycerate dehydrogenase diverts glycolytic flux and contributes to oncogenesis. Nat. Genet. 2011, 43, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Chaneton, B.; Hillmann, P.; Zheng, L.; Martin, A.C.L.; Maddocks, O.D.K.; Chokkathukalam, A.; Coyle, J.E.; Jankevics, A.; Holding, F.P.; Vousden, K.H.; et al. Serine is a natural ligand and allosteric activator of pyruvate kinase M2. Nature 2012, 491, 458–462. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite Profiling Identifies a Key Role for Glycine in Rapid Cancer Cell Proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef] [Green Version]
- Labuschagne, C.F.; Van Den Broek, N.J.F.; Mackay, G.M.; Vousden, K.H.; Maddocks, O.D.K. Serine, but Not Glycine, Supports One-Carbon Metabolism and Proliferation of Cancer Cells. Cell Rep. 2014, 7, 1248–1258. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, O.D.K.; Labuschagne, C.F.; Adams, P.D.; Vousden, K.H. Serine Metabolism Supports the Methionine Cycle and DNA/RNA Methylation through De Novo ATP Synthesis in Cancer Cells. Mol. Cell 2016, 61, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, O.D.K.; Athineos, D.; Cheung, E.C.; Lee, P.; Zhang, T.; Van Den Broek, N.J.F.; Mackay, G.M.; Labuschagne, C.F.; Gay, D.; Kruiswijk, F.; et al. Modulating the therapeutic response of tumours to dietary serine and glycine starvation. Nature 2017, 544, 372–376. [Google Scholar] [CrossRef]
- Mu, Y.; Zhou, Y.; Wang, Y.; Li, W.; Zhou, L.; Lu, X.; Gao, P.; Gao, M.; Zhao, Y.; Wang, Q.; et al. Serum Metabolomics Study of Nonsmoking Female Patients with Non-Small Cell Lung Cancer Using Gas Chromatography–Mass Spectrometry. J. Proteome Res. 2019, 18, 2175–2184. [Google Scholar] [CrossRef] [PubMed]
- Amara, C.S.; Ambati, C.R.; Vantaku, V.; Badrajee Piyarathna, D.W.; Donepudi, S.R.; Ravi, S.S.; Arnold, J.M.; Putluri, V.; Chatta, G.; Guru, K.A.; et al. Serum Metabolic Profiling Identified a Distinct Metabolic Signature in Bladder Cancer Smokers: A Key Metabolic Enzyme Associated with Patient Survival. Cancer Epidemiol. Prev. Biomark. 2019, 28, 770–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vogel, S.; Ulvik, A.; Meyer, K.; Ueland, P.M.; Nygård, O.; Vollset, S.E.; Tell, G.S.; Gregory, J.F.; Tretli, S.; Bjørge, T. Sarcosine and other metabolites along the choline oxidation pathway in relation to prostate cancer--a large nested case-control study within the JANUS cohort in Norway. Int. J. Cancer 2014, 134, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.; Gupta, S.; Hudes, G.R.; Egleston, B.L.; Uzzo, R.G.; Kruger, W.D. Serum Amino Acid Levels as a Biomarker for Renal Cell Carcinoma. J. Urol. 2011, 186, 1206–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Xiao, Y.; Shu, D.; Liang, X.; Hu, X.; Xie, Y.; Lin, D.; Li, H. Metabolomics Analysis in Serum from Patients with Colorectal Polyp and Colorectal Cancer by 1H-NMR Spectrometry. Dis. Markers 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, L.; Maity, S.; Coarfa, C.; Rajapakshe, K.; Chen, L.; Jin, F.; Putluri, V.; Tinker, L.F.; Mo, Q.; Chen, F.; et al. A Prospective Targeted Serum Metabolomics Study of Pancreatic Cancer in Postmenopausal Women. Cancer Prev. Res. 2019, 12, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leichtle, A.B.; Nuoffer, J.-M.; Ceglarek, U.; Kase, J.; Conrad, T.; Witzigmann, H.; Thiery, J.; Fiedler, G.M. Serum amino acid profiles and their alterations in colorectal cancer. Metabolomics 2012, 8, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.M.; Ross, R.K.; Wang, X.L.; Gao, Y.T.; Henderson, B.E.; Yu, M.C. Morbidity and mortality in relation to cigarette smoking in Shanghai, China. A prospective male cohort study. JAMA 1996, 275, 1646–1650. [Google Scholar] [CrossRef]
- Midttun, Ø.; McCann, A.; Aarseth, O.; Krokeide, M.; Kvalheim, G.; Meyer, K.; Ueland, P.M. Combined Measurement of 6 Fat-Soluble Vitamins and 26 Water-Soluble Functional Vitamin Markers and Amino Acids in 50 μL of Serum or Plasma by High-Throughput Mass Spectrometry. Anal. Chem. 2016, 88, 10427–10436. [Google Scholar] [CrossRef]
- Midttun, Ø.; Kvalheim, G.; Ueland, P.M. High-throughput, low-volume, multianalyte quantification of plasma metabolites related to one-carbon metabolism using HPLC-MS/MS. Anal. Bioanal. Chem. 2013, 405, 2009–2017. [Google Scholar] [CrossRef]
- Midttun, Ø.; Hustad, S.; Ueland, P.M. Quantitative profiling of biomarkers related to B-vitamin status, tryptophan metabolism and inflammation in human plasma by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S.; Stevens, L.A.; Schmid, C.H.; Zhang, Y.L.; Castro, A.F.; Feldman, H.I.; Kusek, J.W.; Eggers, P.; Van Lente, F.; Greene, T.; et al. CKD-EPI (Chronic Kidney Disease Epidemiology Collaboration). A new equation to estimate glomerular filtration rate. Ann. Intern Med. 2009, 150, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, N.L.; Hukkanen, J.; Jacob, P. Nicotine chemistry, metabolism, kinetics and biomarkers. Handb. Exp. Pharmacol. 2009, 192, 29–60. [Google Scholar]
- Huang, J.Y.; Butler, L.M.; Midttun, Ø.; Koh, W.-P.; Ueland, P.M.; Wang, R.; Jin, A.; Gao, Y.-T.; Yuan, J.-M. Serum B6 vitamers (pyridoxal 5′-phosphate, pyridoxal, and 4-pyridoxic acid) and pancreatic cancer risk: Two nested case-control studies in Asian populations. Cancer Causes Control 2016, 27, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Luu, H.N.; Butler, L.M.; Midttun, Ø.; Ulvik, A.; Wang, R.; Jin, A.; Gao, Y.-T.; Tan, Y.; Ueland, P.M.; et al. A prospective evaluation of serum methionine-related metabolites in relation to pancreatic cancer risk in two prospective cohort studies. Int. J. Cancer 2020, 147, 1917–1927. [Google Scholar] [CrossRef]
- Obeid, R. The Metabolic Burden of Methyl Donor Deficiency with Focus on the Betaine Homocysteine Methyltransferase Pathway. Nutrients 2013, 5, 3481–3495. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.-D.; Wu, W.K.K.; Wang, H.-Y.; Li, X.-X. Serine and one-carbon metabolism, a bridge that links mTOR signaling and DNA methylation in cancer. Pharmacol. Res. 2019, 149, 104352. [Google Scholar] [CrossRef]
- Yu, L.; Teoh, S.T.; Ensink, E.; Ogrodzinski, M.P.; Yang, C.; Vazquez, A.I.; Lunt, S.Y. Cysteine catabolism and the serine biosynthesis pathway support pyruvate production during pyruvate kinase knockdown in pancreatic cancer cells. Cancer Metab. 2019, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, G.H.; Vassileva, V.; Acedo, P.; Olde Damink, S.W.M.; Malago, M.; Dhar, D.K.; Pereira, S.P. Targeting Pyruvate Kinase M2 and Lactate Dehydrogenase A Is an Effective Combination Strategy for the Treatment of Pancreatic Cancer. Cancers 2019, 11, 1372. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6770573/ (accessed on 25 November 2020). [CrossRef] [Green Version]
- Amelio, I.; Cutruzzolá, F.; Antonov, A.; Agostini, M.; Melino, G. Serine and glycine metabolism in cancer. Trends Biochem. Sci. 2014, 39, 191–198. [Google Scholar] [CrossRef]
- Maddocks, O.D.K.; Berkers, C.R.; Mason, S.M.; Zheng, L.; Blyth, K.; Gottlieb, E.; Vousden, K.H. Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells. Nature 2013, 493, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.-Y.; Kwak, S.; Lee, S.; Kim, H.; Lee, S.E.; Kim, J.-H.; Kim, Y.A.; Jeon, Y.K.; Chung, D.H.; Jin, X.; et al. Psat1-Dependent Fluctuations in α-Ketoglutarate Affect the Timing of ESC Differentiation. Cell Metab. 2016, 24, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.P.; Yashinskie, J.J.; Koche, R.; Chandwani, R.; Tian, S.; Chen, C.-C.; Baslan, T.; Marinkovic, Z.S.; Sánchez-Rivera, F.J.; Leach, S.D.; et al. α-Ketoglutarate links p53 to cell fate during tumour suppression. Nature 2019, 573, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Šarenac, T.M.; Mikov, M. Bile Acid Synthesis: From Nature to the Chemical Modification and Synthesis and Their Applications as Drugs and Nutrients. Front. Pharmacol. 2018, 9, 939. Available online: https://www.frontiersin.org/articles/10.3389/fphar.2018.00939/full (accessed on 6 September 2020). [CrossRef] [PubMed]
- Kevresan, S.; Kuhajda, K.; Kandrac, J.; Fawcett, J.P.; Mikov, M. Biosynthesis of bile acids in mammalian liver. Eur. J. Drug Metab. Pharmacokinet. 2006, 31, 145–156. [Google Scholar] [CrossRef]
- Goossens, J.-F.; Bailly, C. Ursodeoxycholic acid and cancer: From chemoprevention to chemotherapy. Pharmacol. Ther. 2019, 203, 107396. [Google Scholar] [CrossRef]
- Choi, Y.H.; Im, E.O.; Suh, H.; Jin, Y.; Yoo, Y.H.; Kim, N.D. Apoptosis and modulation of cell cycle control by synthetic derivatives of ursodeoxycholic acid and chenodeoxycholic acid in human prostate cancer cells. Cancer Lett. 2003, 199, 157–167. [Google Scholar] [CrossRef]
- Khare, S.; Cerda, S.; Wali, R.K.; Von Lintig, F.C.; Tretiakova, M.; Joseph, L.; Stoiber, D.; Cohen, G.; Nimmagadda, K.; Hart, J.; et al. Ursodeoxycholic acid inhibits Ras mutations, wild-type Ras activation, and cyclooxygenase-2 expression in colon cancer. Cancer Res. 2003, 63, 3517–3523. [Google Scholar] [CrossRef]
- Kim, Y.J.; Jeong, S.H.; Kim, E.-K.; Kim, E.J.; Cho, J.H. Ursodeoxycholic acid suppresses epithelial-mesenchymal transition and cancer stem cell formation by reducing the levels of peroxiredoxin II and reactive oxygen species in pancreatic cancer cells. Oncol. Rep. 2017, 38, 3632–3638. [Google Scholar] [CrossRef]
- Kalhan, S.C.; Uppal, S.O.; Moorman, J.L.; Bennett, C.; Gruca, L.L.; Parimi, P.S.; Dasarathy, S.; Serre, D.; Hanson, R.W. Metabolic and Genomic Response to Dietary Isocaloric Protein Restriction in the Rat. J. Biol. Chem. 2011, 286, 5266–5277. [Google Scholar] [CrossRef] [Green Version]
- Filho, J.C.; Hazel, S.J.; Anderstam, B.; Bergström, J.; Lewitt, M.; Hall, K. Effect of protein intake on plasma and erythrocyte free amino acids and serum IGF-I and IGFBP-1 levels in rats. Am. J. Physiol. 1999, 277, E693–E701. [Google Scholar] [CrossRef] [PubMed]
- Antflick, J.E.; Baker, G.B.; Hampson, D.R. The effects of a low protein diet on amino acids and enzymes in the serine synthesis pathway in mice. Amino Acids 2010, 39, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Nagao, K.; Bannai, M.; Seki, S.; Mori, M.; Takahashi, M. Adaptational modification of serine and threonine metabolism in the liver to essential amino acid deficiency in rats. Amino Acids 2009, 36, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, C.; Lucenteforte, E.; Silverman, D.T.; Petersen, G.; Bracci, P.M.; Ji, B.T.; Negri, E.; Li, D.; Risch, H.A.; Olson, S.; et al. Cigarette smoking and pancreatic cancer: An analysis from the International Pancreatic Cancer Case-Control Consortium (Panc4). Ann. Oncol. 2012, 23, 1880–1888. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.M.; Vrieling, A.; Lubin, J.H.; Kraft, P.; Mendelsohn, J.B.; Hartge, P.; Canzian, F.; Steplowski, E.; Arslan, A.A.; Gross, M.; et al. Cigarette smoking and pancreatic cancer: A pooled analysis from the pancreatic cancer cohort consortium. Am. J. Epidemiol. 2009, 170, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnagnarella, P.; Maisonneuve, P.; Bellomi, M.; Rampinelli, C.; Bertolotti, R.; Spaggiari, L.; Palli, D.; Veronesi, G. Red meat, Mediterranean diet and lung cancer risk among heavy smokers in the COSMOS screening study. Ann. Oncol. 2013, 24, 2606–2611. [Google Scholar] [CrossRef]
- Hawrysz, I.; Wadolowska, L.; Slowinska, M.A.; Czerwinska, A.; Golota, J.J. Adherence to Prudent and Mediterranean Dietary Patterns is Inversely Associated with Lung Cancer in Moderate but not Heavy Male Polish Smokers: A Case-Control Study. Nutrients 2020, 12, E3788. [Google Scholar] [CrossRef]
- Yuan, J.-M.; Stram, D.O.; Arakawa, K.; Lee, H.-P.; Yu, M.C. Dietary cryptoxanthin and reduced risk of lung cancer: The Singapore Chinese Health Study. Cancer Epidemiol. Biomark. Prev. 2003, 12, 890–898. [Google Scholar]
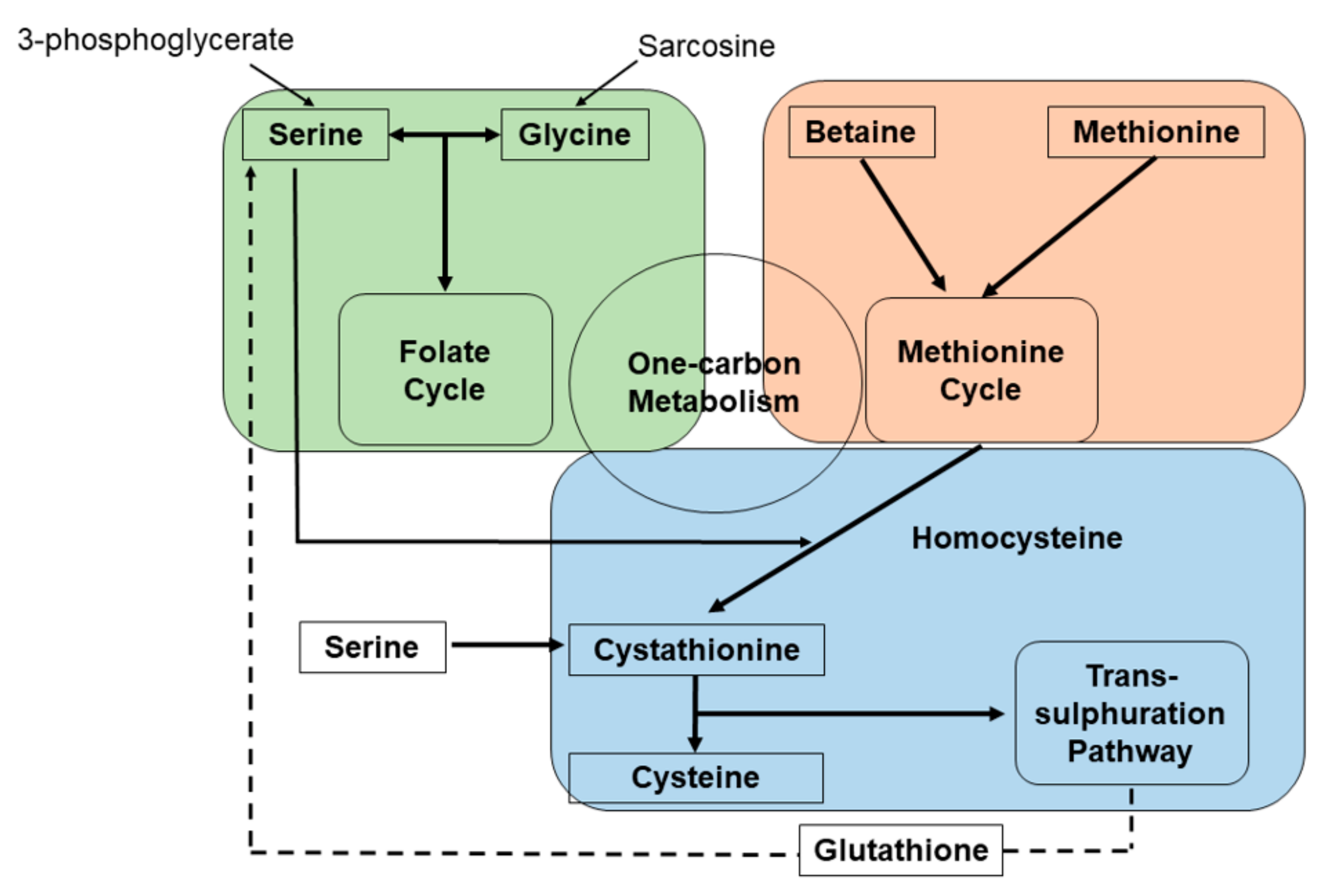

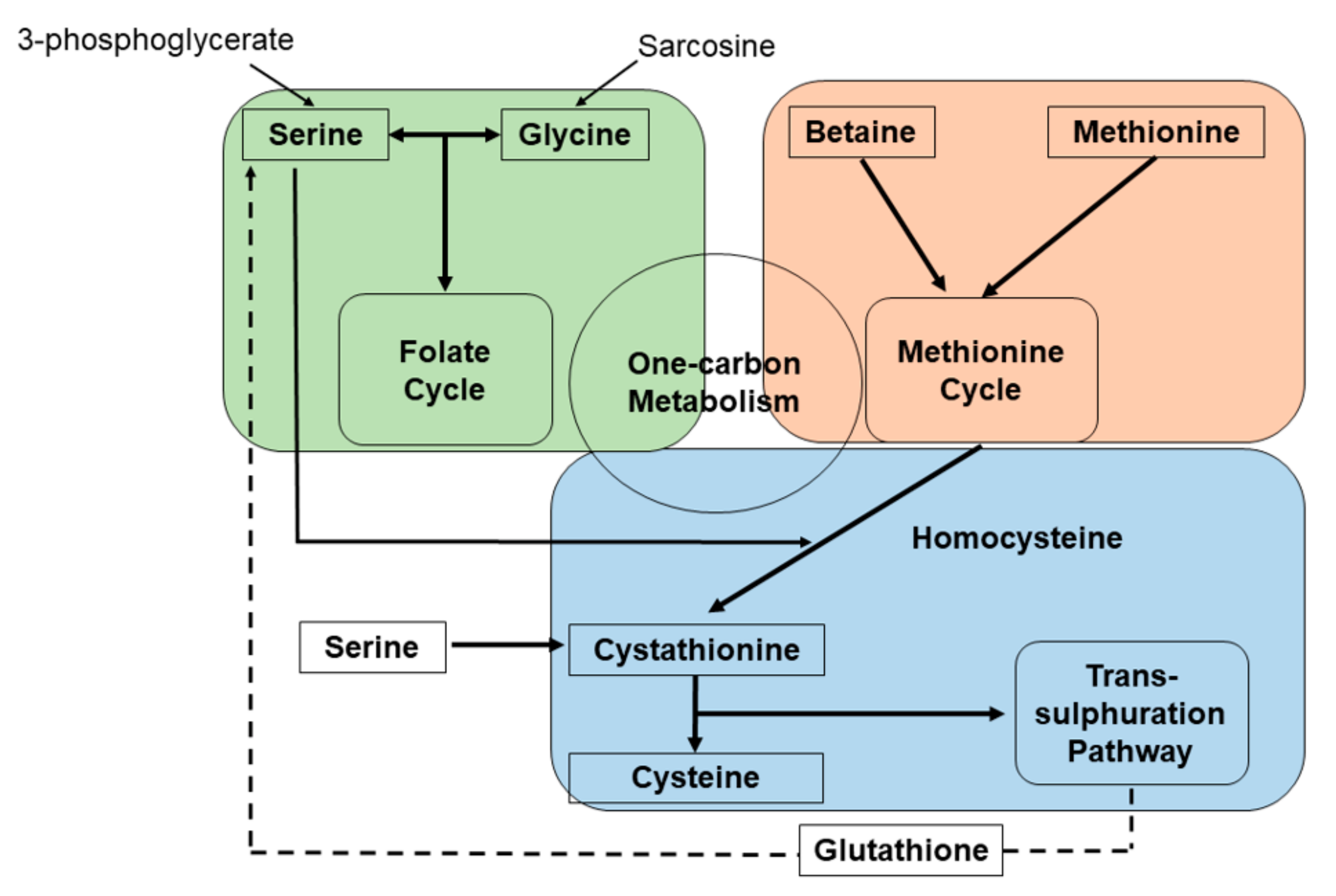{kind=link}
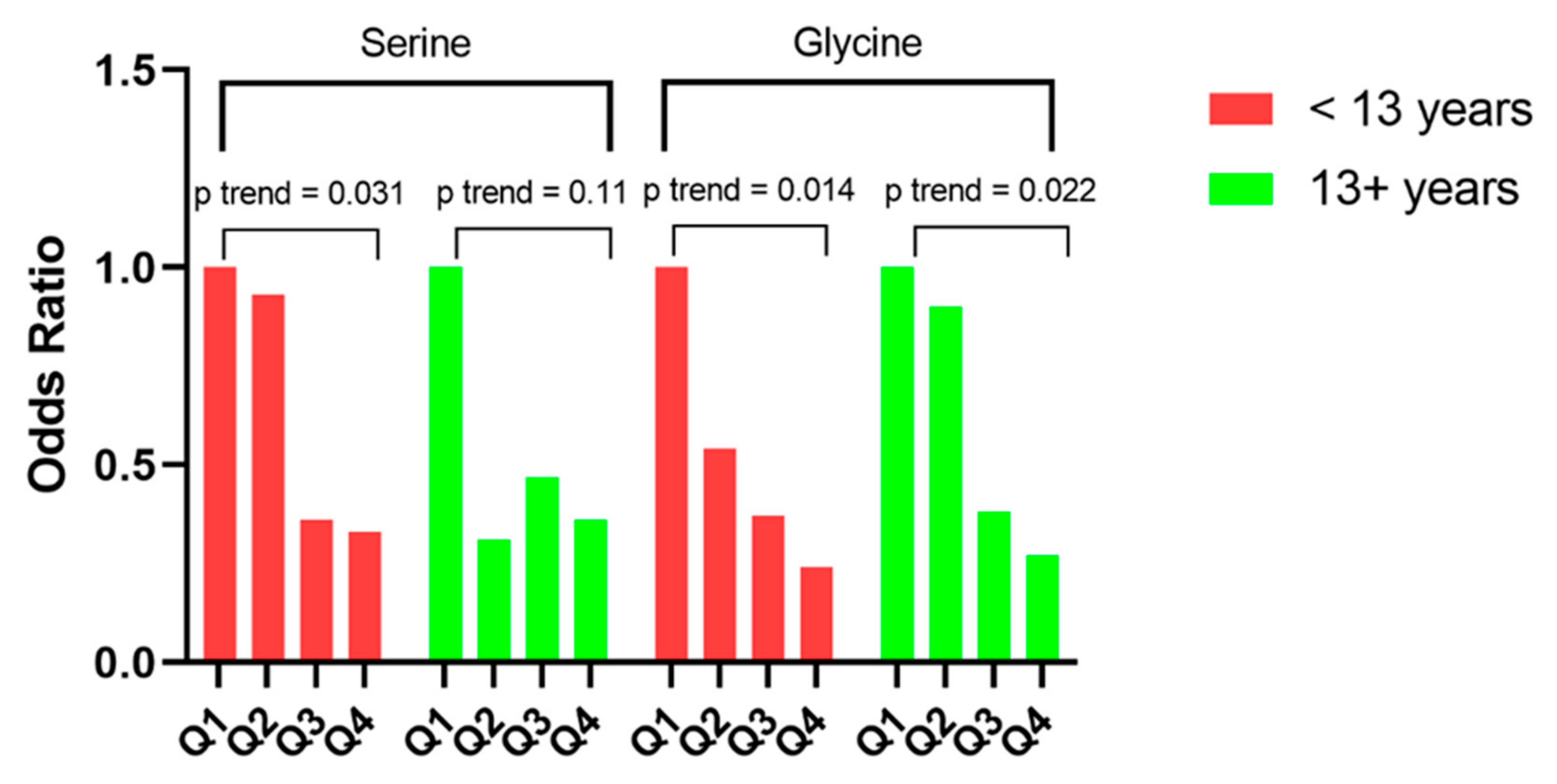
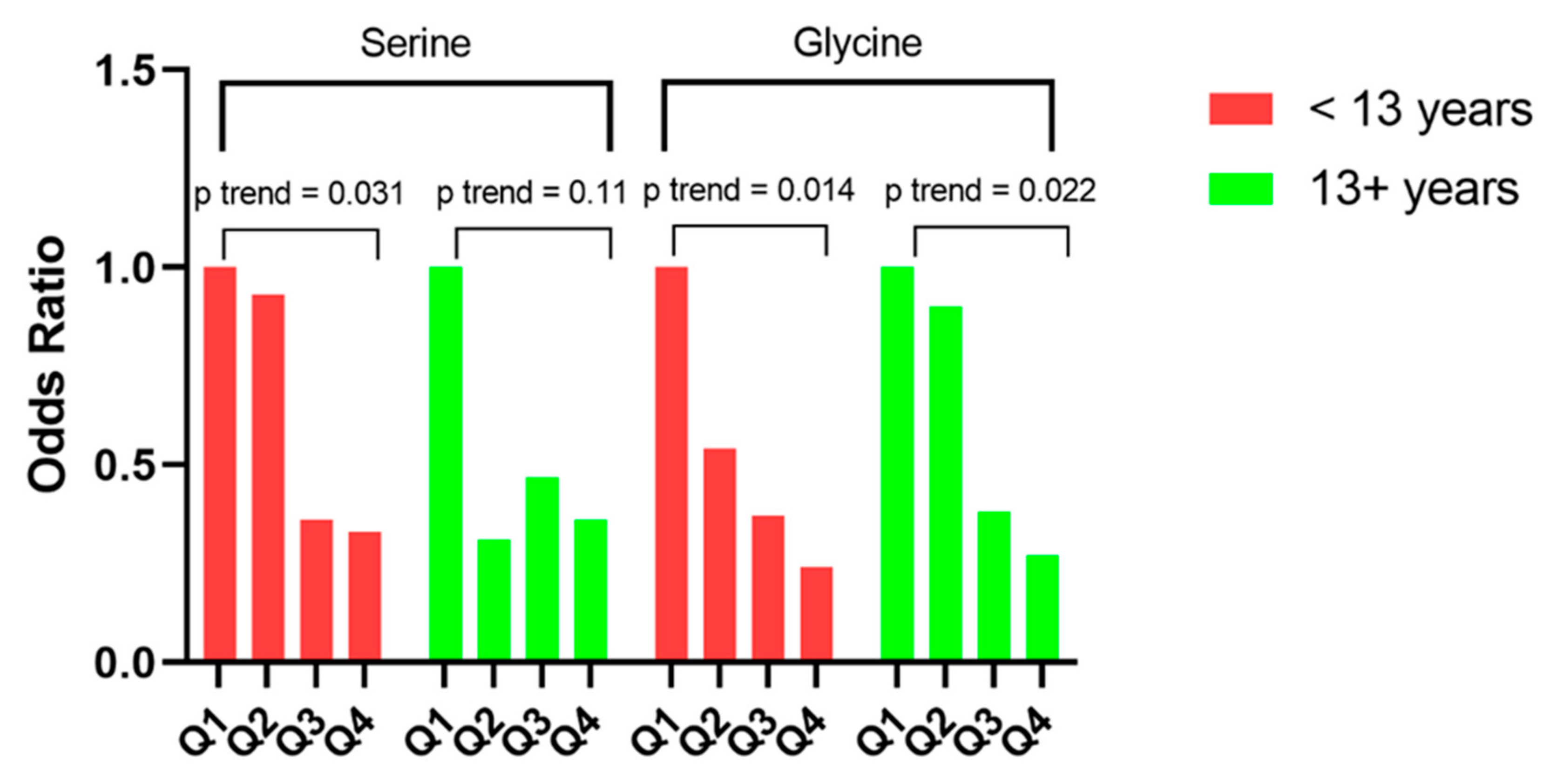{kind=link}
| Baseline Characteristics | Controls | Cases | p |
|---|---|---|---|
| Number of subjects | 258 | 129 | |
| Age (years) (Mean ± SD) | 56.4 ± 5.5 | 56.5 ± 5.5 | 0.74 |
| Body mass index (kg/m2) (Mean ± SD) | 21.9 ± 2.8 | 22.5 ± 3.0 | 0.08 |
| Level of education, n (%) | 0.36 | ||
| No formal schooling | 13 (5.0) | 3 (2.3) | |
| Primary school | 74 (28.7) | 34 (26.4) | |
| Secondary school or above | 171 (66.3) | 92 (71.3) | |
| Smoking status, n (%) | 0.003 | ||
| Never | 113 (43.8) | 35 (27.1) | |
| Former | 16 (6.2) | 6 (4.7) | |
| Current | 129 (50.0) | 88 (68.2) | |
| Cotinine (nmol/L) (Geometric mean ± SD) | 440.0 ± 573.8 | 576.4 ± 573.0 | 0.02 |
| Level of alcohol intake (drinks/week), n (%) | 0.74 | ||
| 0 | 146 (56.6) | 70 (54.3) | |
| <7 | 29 (11.2) | 18 (14.0) | |
| ≥7 | 83 (32.2) | 41 (31.8) | |
| History of diabetes, n (%) | 0.52 | ||
| No | 254 (98.5) | 128 (99.2) | |
| Yes | 4 (1.55) | 1 (0.78) | |
| eGFR (mL/min/1.73 m2) (Geometric mean ± SD) | 89.7 ± 12.4 | 91.5 ± 11.3 | 0.16 |
| PLP (nmol/L) (Geometric mean ± SD) | 35.6 ± 46.9 | 33.9 ± 60.9 | 0.76 |
| Total methyl donors µmol/L(Geometric mean ± SD) a | 114.7 ± 114.2 | 109.4 ± 112.0 | 0.02 |
| Biomarkers * (µmol/L) | Controls, n = 258 Geometric Mean (95% CI) | Cases, n = 129 Geometric Mean (95% CI) | p a |
|---|---|---|---|
| Serine | 186.37 (182.98–189.80) | 179.01 (174.33–183.80) | 0.017 |
| Glycine | 363.05 (356.96–369.24) | 345.52 (337.18–354.00) | 0.002 |
| Cystathionine | 0.28 (0.27–0.30) | 0.29 (0.27–0.31) | 0.70 |
| Cysteine | 272.00 (268.45–275.60) | 273.15 (268.01–278.30) | 0.73 |
| Sarcosine | 1.98 (1.92–2.06) | 2.03 (1.93–2.14) | 0.49 |
| Biomarkers in Quartile | Controls | Cases | OR (95% CI) a |
|---|---|---|---|
| Serine | |||
| Q1 | 65 | 49 | 1.00 |
| Q2 | 64 | 29 | 0.56 (0.30–1.10) |
| Q3 | 65 | 28 | 0.43 (0.22–0.83) |
| Q4 | 64 | 23 | 0.33 (0.14–0.75) |
| ptrend | 0.003 | ||
| Continuous (log2) | 258 | 129 | 0.28 (0.09–0.85) |
| Glycine | |||
| Q1 | 65 | 51 | 1.00 |
| Q2 | 64 | 34 | 0.68 (0.36–1.27) |
| Q3 | 65 | 23 | 0.39 (0.19–0.79) |
| Q4 | 64 | 21 | 0.25 (0.11–0.58) |
| ptrend | 0.001 | ||
| Continuous (log2) | 258 | 129 | 0.14 (0.04–0.51) |
| Cystathionine | |||
| Q1 | 65 | 34 | 1.00 |
| Q2 | 64 | 33 | 0.94 (0.48–1.83) |
| Q3 | 65 | 30 | 0.91 (0.46–1.83) |
| Q4 | 64 | 32 | 1.46 (0.72–2.93) |
| ptrend | 0.77 | ||
| Continuous (log2) | 258 | 129 | 1.09 (0.75–1.59) |
| Cysteine | |||
| Q1 | 65 | 32 | 1.00 |
| Q2 | 64 | 28 | 1.05 (0.54–2.06) |
| Q3 | 65 | 38 | 1.52 (0.78–2.99) |
| Q4 | 64 | 31 | 1.41 (0.69–2.88) |
| ptrend | 0.26 | ||
| Continuous (log2) | 258 | 129 | 1.37 (0.31–6.01) |
| Sarcosine | |||
| Q1 | 65 | 34 | 1.00 |
| Q2 | 64 | 29 | 0.91 (0.47–1.78) |
| Q3 | 65 | 34 | 1.18 (0.62–2.24) |
| Q4 | 64 | 32 | 1.27 (0.65–2.47) |
| ptrend | 0.39 | ||
| Continuous (log2) | 258 | 129 | 1.26 (0.72–2.19) |
| Glycine | Serine | |||||
|---|---|---|---|---|---|---|
| Low (<184.2) | High (≥184.2) | Total | ||||
| Cases/Control | OR (95% CI) | Cases/Control | OR (95% CI) | Cases/Control | OR (95% CI) | |
| Low (<353.3) | 61/90 | 1.00 | 24/39 | 0.77 (0.41–1.46) | 85/129 | 1.00 |
| High (≥353.3) | 17/39 | 0.56 (0.26–1.22) | 27/90 | 0.24 (0.11–0.63) | 44/129 | 0.38 (0.21–0.70) |
| Total | 78/129 | 1.00 | 51/129 | 0.52 (0.31–0.87) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luu, H.N.; Paragomi, P.; Wang, R.; Huang, J.Y.; Adams-Haduch, J.; Midttun, Ø.; Ulvik, A.; Nguyen, T.C.; Brand, R.E.; Gao, Y.; et al. The Association between Serum Serine and Glycine and Related-Metabolites with Pancreatic Cancer in a Prospective Cohort Study. Cancers 2022, 14, 2199. https://doi.org/10.3390/cancers14092199
Luu HN, Paragomi P, Wang R, Huang JY, Adams-Haduch J, Midttun Ø, Ulvik A, Nguyen TC, Brand RE, Gao Y, et al. The Association between Serum Serine and Glycine and Related-Metabolites with Pancreatic Cancer in a Prospective Cohort Study. Cancers. 2022; 14(9):2199. https://doi.org/10.3390/cancers14092199
Chicago/Turabian StyleLuu, Hung N., Pedram Paragomi, Renwei Wang, Joyce Y. Huang, Jennifer Adams-Haduch, Øivind Midttun, Arve Ulvik, Tin C. Nguyen, Randall E. Brand, Yutang Gao, and et al. 2022. "The Association between Serum Serine and Glycine and Related-Metabolites with Pancreatic Cancer in a Prospective Cohort Study" Cancers 14, no. 9: 2199. https://doi.org/10.3390/cancers14092199
APA StyleLuu, H. N., Paragomi, P., Wang, R., Huang, J. Y., Adams-Haduch, J., Midttun, Ø., Ulvik, A., Nguyen, T. C., Brand, R. E., Gao, Y., Ueland, P. M., & Yuan, J.-M. (2022). The Association between Serum Serine and Glycine and Related-Metabolites with Pancreatic Cancer in a Prospective Cohort Study. Cancers, 14(9), 2199. https://doi.org/10.3390/cancers14092199