
High GILT Expression Is Associated with Improved Survival in Metastatic Melanoma Patients Treated with Immune Checkpoint Inhibition

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sets
2.2. Transcriptome Data Preparation
2.3. Patients, Specimens, and Clinical Dataset
2.4. Immunohistochemistry
2.5. Survival and Statistical Analysis
3. Results
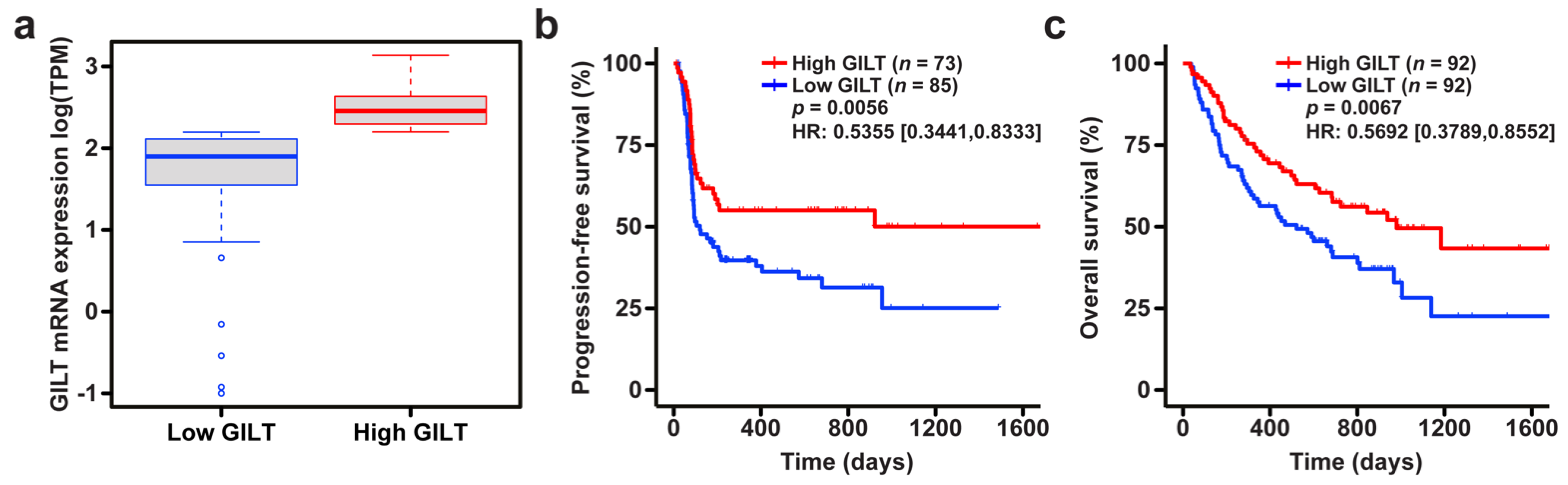
3.1. High GILT mRNA Expression Is Associated with Improved Survival in Melanoma Patients Treated with ICI
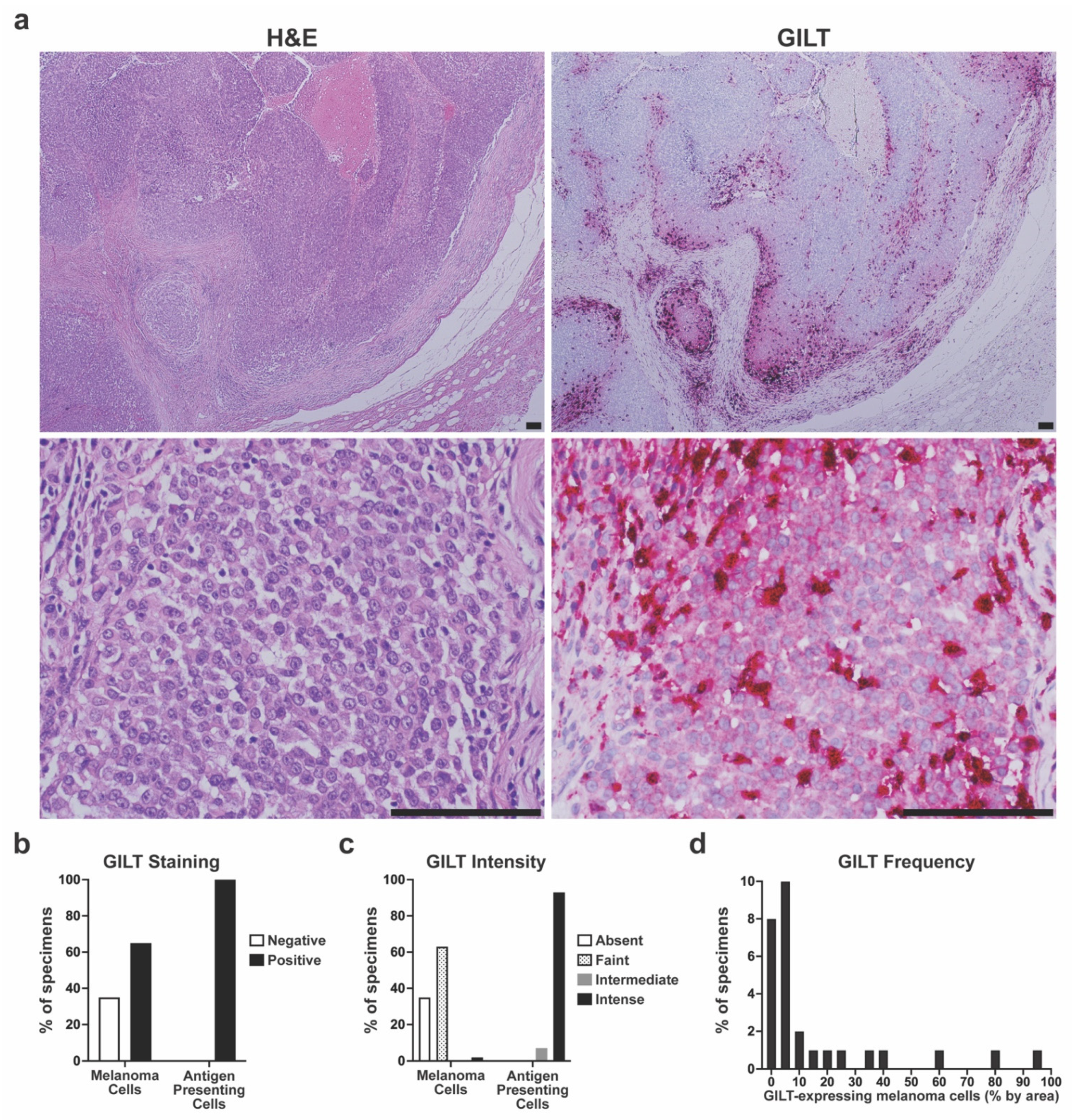
3.2. Variable GILT Protein Expression in Melanoma Cells of Metastatic Tumor Specimens
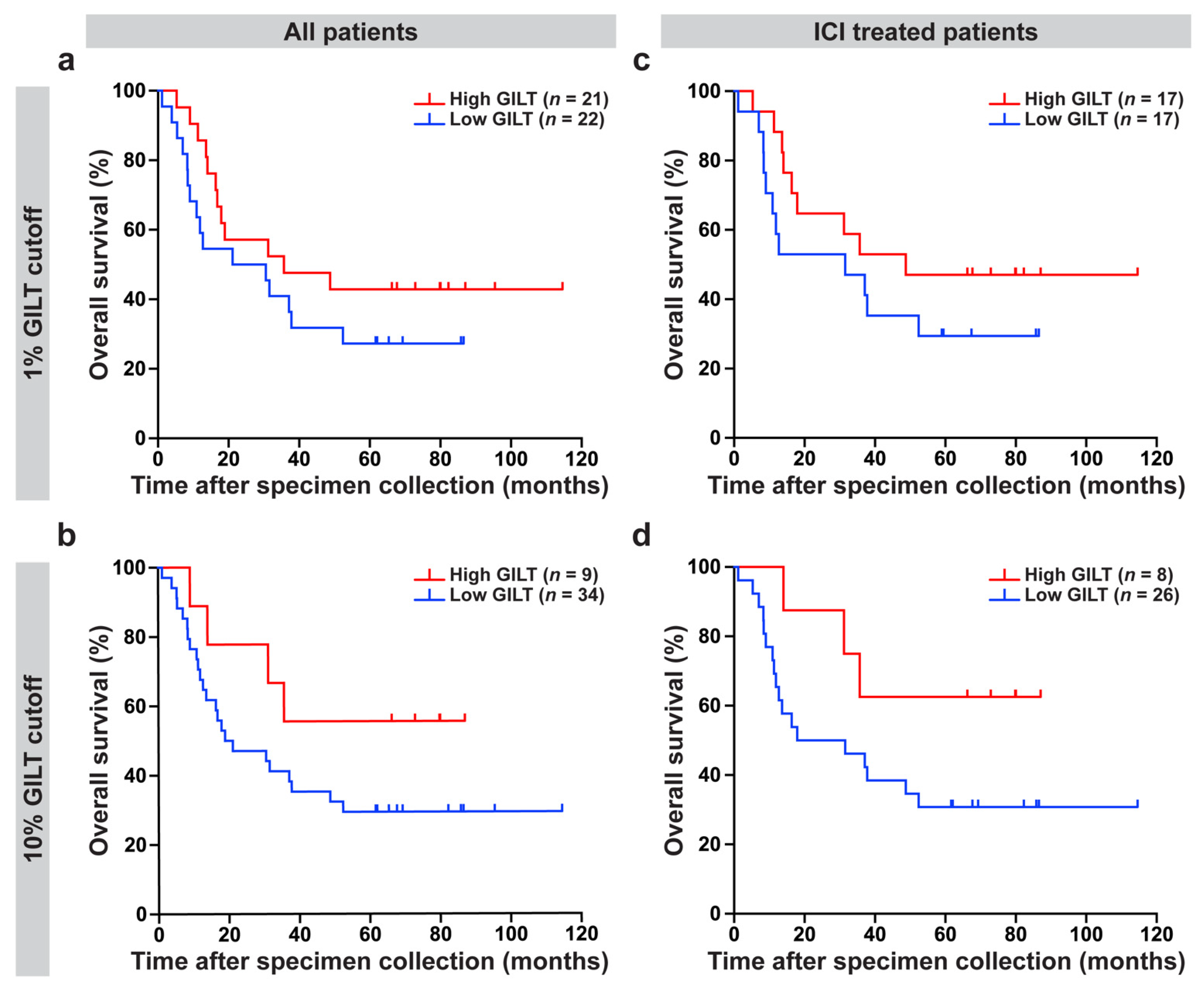
3.3. High GILT Protein Expression in Melanoma Cells Is Associated with Improved Overall Survival in Metastatic Melanoma Patients Treated with ICI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA A Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N. Engl. J. Med. 2015, 373, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, C.; Long, G.V.; Brady, B.; Dutriaux, C.; Maio, M.; Mortier, L.; Hassel, J.C.; Rutkowski, P.; McNeil, C.; Kalinka-Warzocha, E.; et al. Nivolumab in previously untreated melanoma without BRAF mutation. N. Engl. J. Med. 2015, 372, 320–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, C.; Schachter, J.; Long, G.V.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.; Lotem, M.; et al. Pembrolizumab versus Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2015, 372, 2521–2532. [Google Scholar] [CrossRef]
- Wang, Z.; Margulies, L.; Hicklin, D.J.; Ferrone, S. Molecular and functional phenotypes of melanoma cells with abnormalities in HLA class I antigen expression. Tissue Antigens 1996, 47, 382–390. [Google Scholar] [CrossRef]
- Zaretsky, J.M.; Garcia-Diaz, A.; Shin, D.S.; Escuin-Ordinas, H.; Hugo, W.; Hu-Lieskovan, S.; Torrejon, D.Y.; Abril-Rodriguez, G.; Sandoval, S.; Barthly, L.; et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N. Engl. J. Med. 2016, 375, 819–829. [Google Scholar] [CrossRef]
- Rodig, S.J.; Gusenleitner, D.; Jackson, D.G.; Gjini, E.; Giobbie-Hurder, A.; Jin, C.; Chang, H.; Lovitch, S.B.; Horak, C.; Weber, J.S.; et al. MHC proteins confer differential sensitivity to CTLA-4 and PD-1 blockade in untreated metastatic melanoma. Sci. Transl. Med. 2018, 10, eaar3342. [Google Scholar] [CrossRef] [Green Version]
- Algarra, I.; García-Lora, A.; Cabrera, T.; Ruiz-Cabello, F.; Garrido, F. The selection of tumor variants with altered expression of classical and nonclassical MHC class I molecules: Implications for tumor immune escape. Cancer Immunol. Immunother. 2004, 53, 904–910. [Google Scholar] [CrossRef]
- Huang, L.; Malu, S.; McKenzie, J.A.; Andrews, M.C.; Talukder, A.H.; Tieu, T.; Karpinets, T.; Haymaker, C.; Forget, M.A.; Williams, L.J.; et al. The RNA-binding Protein MEX3B Mediates Resistance to Cancer Immunotherapy by Downregulating HLA-A Expression. Clin. Cancer Res. 2018, 24, 3366–3376. [Google Scholar] [CrossRef] [Green Version]
- Laidlaw, B.J.; Craft, J.E.; Kaech, S.M. The multifaceted role of CD4(+) T cells in CD8(+) T cell memory. Nat. Rev. Immunol. 2016, 16, 102–111. [Google Scholar] [CrossRef]
- Janssen, E.M.; Lemmens, E.E.; Wolfe, T.; Christen, U.; von Herrath, M.G.; Schoenberger, S.P. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature 2003, 421, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.G.; Khammanivong, V.; Liu, W.J.; Leggatt, G.R.; Frazer, I.H.; Fernando, G.J. Antigen-specific CD4+ T-cell help is required to activate a memory CD8+ T cell to a fully functional tumor killer cell. Cancer Res. 2002, 62, 6438–6441. [Google Scholar] [PubMed]
- Marzo, A.L.; Kinnear, B.F.; Lake, R.A.; Frelinger, J.J.; Collins, E.J.; Robinson, B.W.; Scott, B. Tumor-specific CD4+ T cells have a major "post-licensing" role in CTL mediated anti-tumor immunity. J. Immunol. 2000, 165, 6047–6055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ossendorp, F.; Mengedé, E.; Camps, M.; Filius, R.; Melief, C.J. Specific T helper cell requirement for optimal induction of cytotoxic T lymphocytes against major histocompatibility complex class II negative tumors. J. Exp. Med. 1998, 187, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quezada, S.A.; Simpson, T.R.; Peggs, K.S.; Merghoub, T.; Vider, J.; Fan, X.; Blasberg, R.; Yagita, H.; Muranski, P.; Antony, P.A.; et al. Tumor-reactive CD4(+) T cells develop cytotoxic activity and eradicate large established melanoma after transfer into lymphopenic hosts. J. Exp. Med. 2010, 207, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Lundin, K.U.; Hofgaard, P.O.; Omholt, H.; Munthe, L.A.; Corthay, A.; Bogen, B. Therapeutic effect of idiotype-specific CD4+ T cells against B-cell lymphoma in the absence of anti-idiotypic antibodies. Blood 2003, 102, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Cachot, A.; Bilous, M.; Liu, Y.C.; Li, X.; Saillard, M.; Cenerenti, M.; Rockinger, G.A.; Wyss, T.; Guillaume, P.; Schmidt, J.; et al. Tumor-specific cytolytic CD4 T cells mediate immunity against human cancer. Sci. Adv. 2021, 7, eabe3348. [Google Scholar] [CrossRef]
- Haabeth, O.A.W.; Fauskanger, M.; Manzke, M.; Lundin, K.U.; Corthay, A.; Bogen, B.; Tveita, A.A. CD4(+) T-cell-Mediated Rejection of MHC Class II-Positive Tumor Cells Is Dependent on Antigen Secretion and Indirect Presentation on Host APCs. Cancer Res. 2018, 78, 4573–4585. [Google Scholar] [CrossRef] [Green Version]
- Antony, P.A.; Piccirillo, C.A.; Akpinarli, A.; Finkelstein, S.E.; Speiss, P.J.; Surman, D.R.; Palmer, D.C.; Chan, C.C.; Klebanoff, C.A.; Overwijk, W.W.; et al. CD8+ T cell immunity against a tumor/self-antigen is augmented by CD4+ T helper cells and hindered by naturally occurring T regulatory cells. J. Immunol. 2005, 174, 2591–2601. [Google Scholar] [CrossRef] [Green Version]
- Homet Moreno, B.; Zaretsky, J.M.; Garcia-Diaz, A.; Tsoi, J.; Parisi, G.; Robert, L.; Meeth, K.; Ndoye, A.; Bosenberg, M.; Weeraratna, A.T.; et al. Response to Programmed Cell Death-1 Blockade in a Murine Melanoma Syngeneic Model Requires Costimulation, CD4, and CD8 T Cells. Cancer Immunol. Res. 2016, 4, 845–857. [Google Scholar] [CrossRef] [Green Version]
- Alspach, E.; Lussier, D.M.; Miceli, A.P.; Kizhvatov, I.; DuPage, M.; Luoma, A.M.; Meng, W.; Lichti, C.F.; Esaulova, E.; Vomund, A.N.; et al. MHC-II neoantigens shape tumour immunity and response to immunotherapy. Nature 2019, 574, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Hastings, K.T.; Cresswell, P. Disulfide reduction in the endocytic pathway: Immunological functions of gamma-interferon-inducible lysosomal thiol reductase. Antioxid. Redox Signal. 2011, 15, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastings, K.T.; Lackman, R.L.; Cresswell, P. Functional requirements for the lysosomal thiol reductase GILT in MHC class II-restricted antigen processing. J. Immunol. 2006, 177, 8569–8577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, M.A.; Li, P.; Jackson, S.K.; Zarour, H.M.; Hawes, J.W.; Phan, U.T.; Maric, M.; Cresswell, P.; Blum, J.S. Absence of gamma-interferon-inducible lysosomal thiol reductase in melanomas disrupts T cell recognition of select immunodominant epitopes. J. Exp. Med. 2002, 195, 1267–1277. [Google Scholar] [CrossRef]
- Rausch, M.P.; Irvine, K.R.; Antony, P.A.; Restifo, N.P.; Cresswell, P.; Hastings, K.T. GILT accelerates autoimmunity to the melanoma antigen tyrosinase-related protein 1. J. Immunol. 2010, 185, 2828–2835. [Google Scholar] [CrossRef] [Green Version]
- Hathaway-Schrader, J.D.; Norton, D.; Hastings, K.; Doonan, B.P.; Fritz, S.T.; Bethard, J.R.; Blum, J.S.; Haque, A. GILT Expression in Human Melanoma Cells Enhances Generation of Antigenic Peptides for HLA Class II-Mediated Immune Recognition. Int. J. Mol. Sci. 2022, 23, 1066. [Google Scholar] [CrossRef]
- Buetow, K.H.; Meador, L.R.; Menon, H.; Lu, Y.K.; Brill, J.; Cui, H.; Roe, D.J.; DiCaudo, D.J.; Hastings, K.T. High GILT Expression and an Active and Intact MHC Class II Antigen Presentation Pathway Are Associated with Improved Survival in Melanoma. J. Immunol. 2019, 203, 2577–2587. [Google Scholar] [CrossRef]
- Deffrennes, V.; Vedrenne, J.; Stolzenberg, M.C.; Piskurich, J.; Barbieri, G.; Ting, J.P.; Charron, D.; Alcaïde-Loridan, C. Constitutive expression of MHC class II genes in melanoma cell lines results from the transcription of class II transactivator abnormally initiated from its B cell-specific promoter. J. Immunol. 2001, 167, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, B.L.; Xi, H.; Tejiram, R.; Eason, D.D.; Ghosh, N.; Wright, K.L.; Nagarajan, U.; Boss, J.M.; Blanck, G. Varying functions of specific major histocompatibility class II transactivator promoter III and IV elements in melanoma cell lines. Cell Growth Differ. 2001, 12, 327–335. [Google Scholar]
- Phipps-Yonas, H.; Cui, H.; Sebastiao, N.; Brunhoeber, P.S.; Haddock, E.; Deymier, M.J.; Klapper, W.; Lybarger, L.; Roe, D.J.; Hastings, K.T. Low GILT Expression is Associated with Poor Patient Survival in Diffuse Large B-Cell Lymphoma. Front. Immunol. 2013, 4, 425. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, J.; Bernert, R.; In, K.; Kang, P.; Sebastiao, N.; Hu, C.; Hastings, K.T. Gamma-interferon-inducible lysosomal thiol reductase is upregulated in human melanoma. Melanoma Res. 2016, 26, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, S.; Spallanzani, A.; Pinzi, C.; Prignano, F.; Fabbri, P. Fibrosis in regressing melanoma versus nonfibrosis in halo nevus upon melanocyte disappearance: Could it be related to a different cytokine microenvironment? J. Cutan. Pathol. 2007, 34, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Luo, Y.; Sun, Y.; Guo, W.; Zhao, X.; Xi, Y.; Ma, Y.; Shao, M.; Tan, W.; Gao, G.; et al. Genomic and transcriptomic alterations associated with drug vulnerabilities and prognosis in adenocarcinoma at the gastroesophageal junction. Nat. Commun. 2020, 11, 6091. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Liu, J.; Wang, J.; Zhong, X.; Dong, D.; Yang, X.; Wang, Y. Development and validation of a prognostic immune-associated gene signature in clear cell renal cell carcinoma. Int. Immunopharmacol. 2020, 81, 106274. [Google Scholar] [CrossRef]
- Xiang, Y.J.; Guo, M.M.; Zhou, C.J.; Liu, L.; Han, B.; Kong, L.Y.; Gao, Z.C.; Ma, Z.B.; Wang, L.; Feng, M.; et al. Absence of gamma-interferon-inducible lysosomal thiol reductase (GILT) is associated with poor disease-free survival in breast cancer patients. PLoS ONE 2014, 9, e109449. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zhou, W.; Wang, F.; Yin, G.; Zhang, X.; Kong, L.; Gao, Z.; Feng, M.; Zhou, C.; Sun, D.; et al. Prognostic value of gamma-interferon-inducible lysosomal thiol reductase expression in female patients diagnosed with breast cancer. Int. J. Cancer 2021, 150, 705–717. [Google Scholar] [CrossRef]
- Advani, A.S.; Cooper, B.; Visconte, V.; Elson, P.; Chan, R.; Carew, J.; Wei, W.; Mukherjee, S.; Gerds, A.; Carraway, H.; et al. A Phase I/II Trial of MEC (Mitoxantrone, Etoposide, Cytarabine) in Combination with Ixazomib for Relapsed Refractory Acute Myeloid Leukemia. Clin. Cancer Res. 2019, 25, 4231–4237. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Schilling, B.; Liu, D.; Sucker, A.; Livingstone, E.; Jerby-Arnon, L.; Zimmer, L.; Gutzmer, R.; Satzger, I.; Loquai, C.; et al. Integrative molecular and clinical modeling of clinical outcomes to PD1 blockade in patients with metastatic melanoma. Nat. Med. 2019, 25, 1916–1927. [Google Scholar] [CrossRef] [Green Version]
- Van Allen, E.M.; Miao, D.; Schilling, B.; Shukla, S.A.; Blank, C.; Zimmer, L.; Sucker, A.; Hillen, U.; Foppen, M.H.G.; Goldinger, S.M.; et al. Genomic correlates of response to CTLA-4 blockade in metastatic melanoma. Science 2015, 350, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Hugo, W.; Zaretsky, J.M.; Sun, L.; Song, C.; Moreno, B.H.; Hu-Lieskovan, S.; Berent-Maoz, B.; Pang, J.; Chmielowski, B.; Cherry, G.; et al. Genomic and Transcriptomic Features of Response to Anti-PD-1 Therapy in Metastatic Melanoma. Cell 2016, 165, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef] [PubMed]
- Maric, M.; Arunachalam, B.; Phan, U.T.; Dong, C.; Garrett, W.S.; Cannon, K.S.; Alfonso, C.; Karlsson, L.; Flavell, R.A.; Cresswell, P. Defective antigen processing in GILT-free mice. Science 2001, 294, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Hastings, K.T. GILT: Shaping the MHC Class II-Restricted Peptidome and CD4(+) T Cell-Mediated Immunity. Front. Immunol. 2013, 4, 429. [Google Scholar] [CrossRef] [Green Version]
- Meador, L.R.; Hastings, K.T. Gamma-Interferon-Inducible Lysosomal Thiol Reductase (GILT). In Encyclopedia of Signaling Molecules; Choi, S., Ed.; Springer International Publishing: Cham, Germany, 2018; pp. 2007–2021. [Google Scholar]
- Meyer, S.; Handke, D.; Mueller, A.; Biehl, K.; Kreuz, M.; Bukur, J.; Koehl, U.; Lazaridou, M.F.; Berneburg, M.; Steven, A.; et al. Distinct Molecular Mechanisms of Altered HLA Class II Expression in Malignant Melanoma. Cancers 2021, 13, 3907. [Google Scholar] [CrossRef]
- Harel, M.; Ortenberg, R.; Varanasi, S.K.; Mangalhara, K.C.; Mardamshina, M.; Markovits, E.; Baruch, E.N.; Tripple, V.; Arama-Chayoth, M.; Greenberg, E.; et al. Proteomics of Melanoma Response to Immunotherapy Reveals Mitochondrial Dependence. Cell 2019, 179, 236–250.e218. [Google Scholar] [CrossRef]
- Abelin, J.G.; Harjanto, D.; Malloy, M.; Suri, P.; Colson, T.; Goulding, S.P.; Creech, A.L.; Serrano, L.R.; Nasir, G.; Nasrullah, Y.; et al. Defining HLA-II Ligand Processing and Binding Rules with Mass Spectrometry Enhances Cancer Epitope Prediction. Immunity 2019, 51, 766–779.e717. [Google Scholar] [CrossRef]
- Johnson, D.B.; Estrada, M.V.; Salgado, R.; Sanchez, V.; Doxie, D.B.; Opalenik, S.R.; Vilgelm, A.E.; Feld, E.; Johnson, A.S.; Greenplate, A.R.; et al. Melanoma-specific MHC-II expression represents a tumour-autonomous phenotype and predicts response to anti-PD-1/PD-L1 therapy. Nat. Commun. 2016, 7, 10582. [Google Scholar] [CrossRef] [Green Version]
- Meazza, R.; Comes, A.; Orengo, A.M.; Ferrini, S.; Accolla, R.S. Tumor rejection by gene transfer of the MHC class II transactivator in murine mammary adenocarcinoma cells. Eur. J. Immunol. 2003, 33, 1183–1192. [Google Scholar] [CrossRef]
- Mortara, L.; Castellani, P.; Meazza, R.; Tosi, G.; De Lerma Barbaro, A.; Procopio, F.A.; Comes, A.; Zardi, L.; Ferrini, S.; Accolla, R.S. CIITA-induced MHC class II expression in mammary adenocarcinoma leads to a Th1 polarization of the tumor microenvironment, tumor rejection, and specific antitumor memory. Clin. Cancer Res. 2006, 12, 3435–3443. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.B.; Nixon, M.J.; Wang, Y.; Wang, D.Y.; Castellanos, E.; Estrada, M.V.; Ericsson-Gonzalez, P.I.; Cote, C.H.; Salgado, R.; Sanchez, V.; et al. Tumor-specific MHC-II expression drives a unique pattern of resistance to immunotherapy via LAG-3/FCRL6 engagement. JCI Insight 2018, 3, e120360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaw, T.R.; Li, M.; Starenki, D.; Cooper, S.J.; Liu, M.; Meza-Perez, S.; Arend, R.C.; Buchsbaum, D.J.; Forero, A.; Randall, T.D. The expression of MHC class II molecules on murine breast tumors delays T-cell exhaustion, expands the T-cell repertoire, and slows tumor growth. Cancer Immunol. Immunother. 2019, 68, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Ostrand-Rosenberg, S.; Clements, V.K.; Thakur, A.; Cole, G.A. Transfection of major histocompatibility complex class I and class II genes causes tumour rejection. J. Immunogenet. 1989, 16, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Frangione, V.; Mortara, L.; Castellani, P.; De Lerma Barbaro, A.; Accolla, R.S. CIITA-driven MHC-II positive tumor cells: Preventive vaccines and superior generators of antitumor CD4+ T lymphocytes for immunotherapy. Int. J. Cancer 2010, 127, 1614–1624. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, J.; Togashi, Y.; Sugawara, T.; Itami, M.; Yamauchi, N.; Yuda, J.; Sugano, M.; Ohara, Y.; Minami, Y.; Nakamae, H.; et al. The critical role of CD4+ T cells in PD-1 blockade against MHC-II-expressing tumors such as classic Hodgkin lymphoma. Blood Adv. 2020, 4, 4069–4082. [Google Scholar] [CrossRef] [PubMed]
- Robila, V.; Ostankovitch, M.; Altrich-VanLith, M.L.; Theos, A.C.; Drover, S.; Marks, M.S.; Restifo, N.; Engelhard, V.H. MHC Class II Presentation of gp100 Epitopes in Melanoma Cells Requires the Function of Conventional Endosomes and Is Influenced by Melanosomes. J. Immunol. 2008, 181, 7843–7852. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, T.; Matsuzaki, J.; Caballero, O.L.; Jungbluth, A.A.; Ritter, G.; Odunsi, K.; Old, L.J.; Gnjatic, S. Heat shock protein 90-mediated peptide-selective presentation of cytosolic tumor antigen for direct recognition of tumors by CD4(+) T cells. J. Immunol. 2012, 188, 3851–3858. [Google Scholar] [CrossRef] [Green Version]
- Topalian, S.L.; Gonzales, M.I.; Parkhurst, M.; Li, Y.F.; Southwood, S.; Sette, A.; Rosenberg, S.A.; Robbins, P.F. Melanoma-specific CD4+ T cells recognize nonmutated HLA-DR-restricted tyrosinase epitopes. J. Exp. Med. 1996, 183, 1965–1971. [Google Scholar] [CrossRef] [Green Version]
- Manici, S.; Sturniolo, T.; Imro, M.A.; Hammer, J.; Sinigaglia, F.; Noppen, C.; Spagnoli, G.; Mazzi, B.; Bellone, M.; Dellabona, P.; et al. Melanoma cells present a MAGE-3 epitope to CD4(+) cytotoxic T cells in association with histocompatibility leukocyte antigen DR11. J. Exp. Med. 1999, 189, 871–876. [Google Scholar] [CrossRef]
- Wang, R.F.; Wang, X.; Atwood, A.C.; Topalian, S.L.; Rosenberg, S.A. Cloning genes encoding MHC class II-restricted antigens: Mutated CDC27 as a tumor antigen. Science 1999, 284, 1351–1354. [Google Scholar] [CrossRef]
- Touloukian, C.E.; Leitner, W.W.; Topalian, S.L.; Li, Y.F.; Robbins, P.F.; Rosenberg, S.A.; Restifo, N.P. Identification of a MHC class II-restricted human gp100 epitope using DR4-IE transgenic mice. J. Immunol. 2000, 164, 3535–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogunovic, B.; Srinivasan, P.; Ueda, Y.; Tomita, Y.; Maric, M. Comparative Quantitative Mass Spectrometry Analysis of MHC Class II-Associated Peptides Reveals a Role of GILT in Formation of Self-Peptide Repertoire. PLoS ONE 2010, 5, e10599. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Liu Dataset | |
|---|---|
| Number of patients | n = 122 |
| Females | n = 51 |
| Males | n = 71 |
| Length of follow up (mean ± SD) | 18.8 ± 12.8 months |
| Melanoma subtype | |
| Cutaneous | 73% (89/122) |
| Occult | 16% (19/122) |
| Acral | 7% (8/122) |
| Mucosal | 5% (6/122) |
| Stage | |
| M0 | 8% (10/122) |
| M1a | 6% (7/122) |
| M1b | 11% (14/122) |
| M1c | 76% (91/122) |
| Immune checkpoint inhibitors | |
| Anti-PD-1 therapy alone | 100% (122/122) |
| Other treatments | |
| MAPK therapy prior to anti-PD-1 therapy | 15% (18/122) |
| MAPK therapy after anti-PD-1 therapy | 9% (11/122) |
| Anti-CTLA-4 therapy prior to anti-PD-1 therapy | 39% (48/122) |
| Anti-CTLA-4 therapy after anti-PD-1 therapy | 9% (11/122) |
| Combined anti-CTLA-4/anti-PD-1 after anti-PD-1 therapy | 13% (16/122) |
| Van Allen Dataset | |
| Number of patients | n = 36 |
| Females | n = 9 |
| Males | n = 27 |
| Age of patients at diagnosis (mean ± SD) | 60.3 ± 15.5 years |
| Length of follow up (mean ± SD) | 18.5 ± 15.5 months |
| Melanoma subtype | |
| Cutaneous | 100% (36/36) |
| Stage | |
| M0 | 3% (1/36) |
| M1a | 6% (2/36) |
| M1b | 17% (6/36) |
| M1c | 75% (27/36) |
| Immune checkpoint inhibitors | |
| Anti-CTLA-4 therapy alone | 100% (36/36) |
| Other treatments | |
| BRAF inhibitor prior to anti-CTLA-4 therapy | 11% (4/36) |
| BRAF inhibitor after anti-CTLA-4 therapy | 33% (6/36) |
| Hugo Dataset | |
| Number of patients | n = 26 |
| Females | n = 8 |
| Males | n = 18 |
| Age of patients at diagnosis (mean ± SD) | 59.3 ± 15.1 years |
| Length of follow up (mean ± SD) | 17.3 ± 10.4 months |
| Melanoma subtype | Not reported |
| Stage | |
| M0 | 4% (1/27) |
| M1a | 7% (2/27) |
| M1b | 11% (3/27) |
| M1c | 78% (21/27) |
| Other treatments | |
| MAPK therapy prior to anti-PD-1 therapy | 15% (12/27) |
| Metastatic Melanoma Specimen Dataset | |
|---|---|
| Number of patients | n = 43 |
| Females | n = 11 |
| Males | n = 32 |
| Average age of all patients at time of collection (mean ± SD) | 64.1 ± 14.3 years |
| Average length of follow up (mean ± SD) | 39.6 ± 31.9 months |
| Stage at the time of specimen collection | |
| II | 2% (1/43) |
| III | 28% (12/43) |
| IV | 70% (30/43) |
| BRAF V600E status | |
| Positive | 35% (15/43) |
| Negative | 56% (24/43) |
| Not tested | 9% (4/43) |
| Immune checkpoint inhibitors | |
| Anti-CTLA-4 therapy alone | 35% (15/43) |
| Anti-PD-1 therapy alone | 30% (13/43) |
| Anti-CTLA-4 and anti-PD-1 therapy | 14% (6/43) |
| Other treatments | |
| BRAF inhibitor therapy | 21% (9/43) |
| Chemotherapy | 33% (14/43) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adams, A.C.; Borden, E.S.; Macy, A.M.; Thomson, N.; Cui, H.; Gimbel, M.I.; Wilson, M.A.; Buetow, K.H.; Roe, D.J.; DiCaudo, D.J.; et al. High GILT Expression Is Associated with Improved Survival in Metastatic Melanoma Patients Treated with Immune Checkpoint Inhibition. Cancers 2022, 14, 2200. https://doi.org/10.3390/cancers14092200
Adams AC, Borden ES, Macy AM, Thomson N, Cui H, Gimbel MI, Wilson MA, Buetow KH, Roe DJ, DiCaudo DJ, et al. High GILT Expression Is Associated with Improved Survival in Metastatic Melanoma Patients Treated with Immune Checkpoint Inhibition. Cancers. 2022; 14(9):2200. https://doi.org/10.3390/cancers14092200
Chicago/Turabian StyleAdams, Anngela C., Elizabeth S. Borden, Anne M. Macy, Nick Thomson, Haiyan Cui, Mark I. Gimbel, Melissa A. Wilson, Kenneth H. Buetow, Denise J. Roe, David J. DiCaudo, and et al. 2022. "High GILT Expression Is Associated with Improved Survival in Metastatic Melanoma Patients Treated with Immune Checkpoint Inhibition" Cancers 14, no. 9: 2200. https://doi.org/10.3390/cancers14092200
APA StyleAdams, A. C., Borden, E. S., Macy, A. M., Thomson, N., Cui, H., Gimbel, M. I., Wilson, M. A., Buetow, K. H., Roe, D. J., DiCaudo, D. J., Homsi, J., & Hastings, K. T. (2022). High GILT Expression Is Associated with Improved Survival in Metastatic Melanoma Patients Treated with Immune Checkpoint Inhibition. Cancers, 14(9), 2200. https://doi.org/10.3390/cancers14092200