
Dietary Interventions in Cancer Treatment and Response: A Comprehensive Review

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
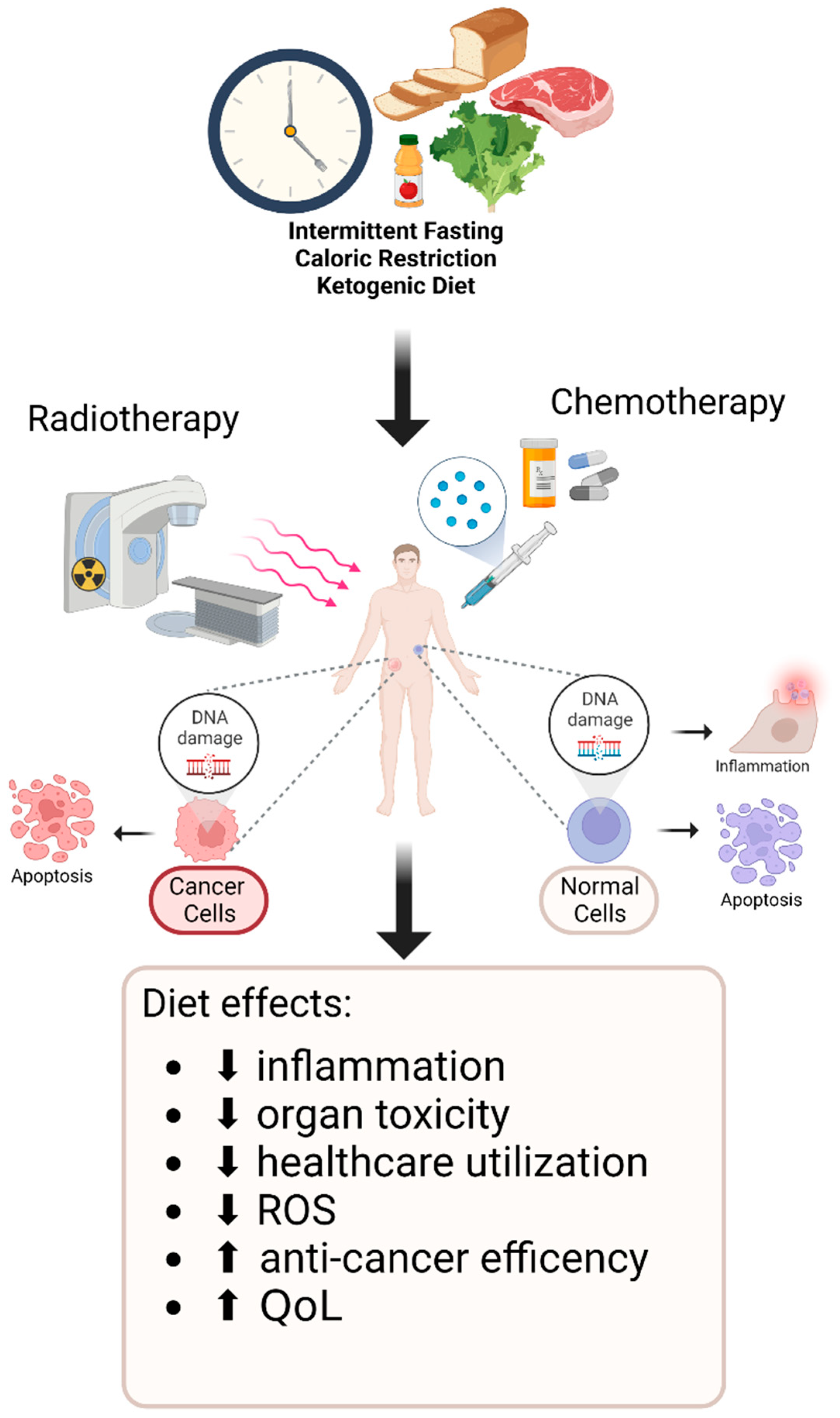
3. Dietary Interventions
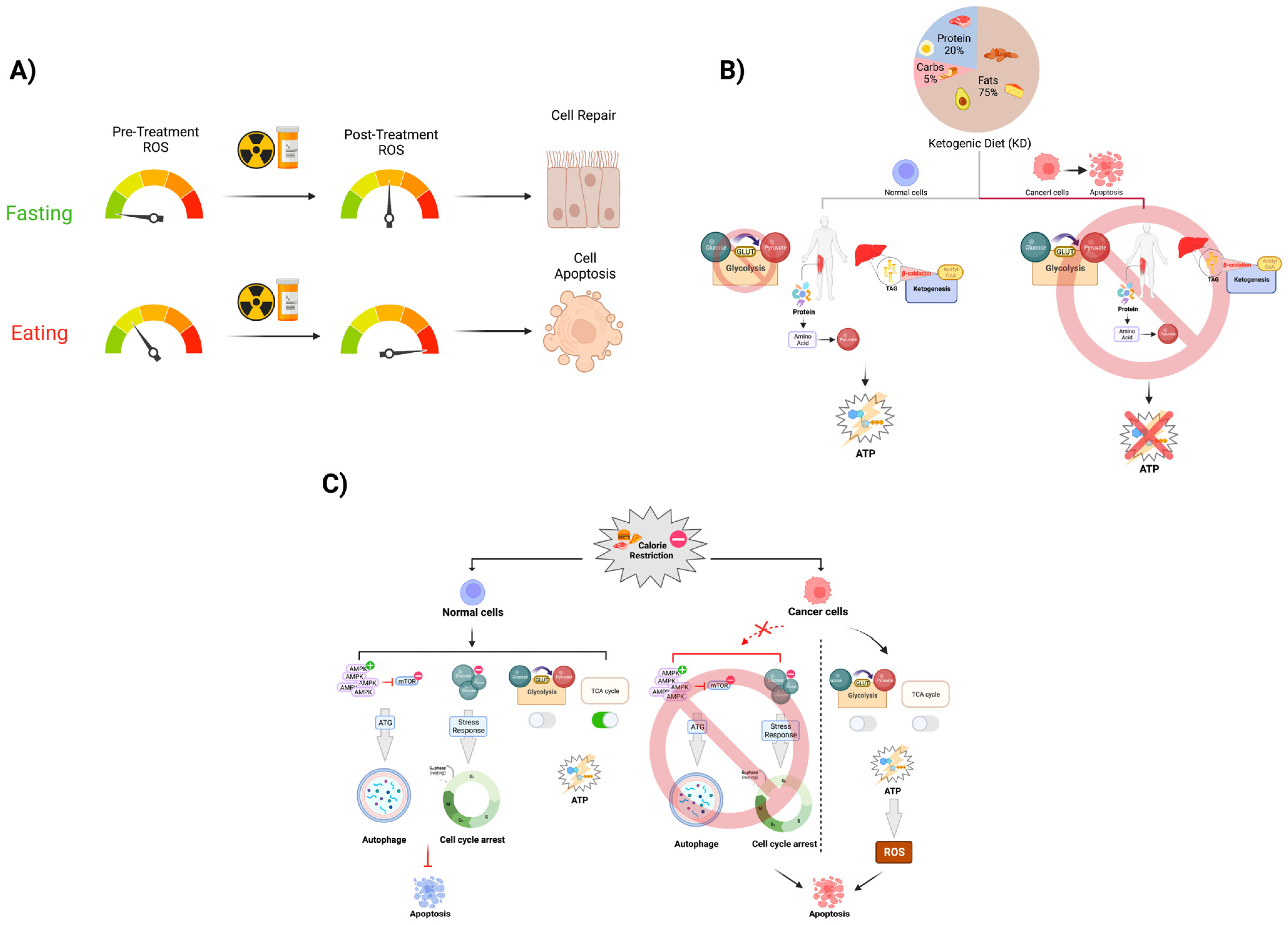
3.1. Caloric Restriction and Fasting-Mimicking Diets Assist in Further Attenuating Tumor Growth with CT/RT Treatment Regimens
3.2. Intermittent Fasting Is a Safe and Feasible Way of Increasing the Efficacy of CT/RT Treatment as Well as Decreasing Treatment Toxicity
3.3. Ketogenic Diet Has Been Shown to Maintain Non-Fat Body Mass and Increase CT/RT Treatment Efficacy in Murine Models and Human Cancer Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- DeVita, V.T., Jr.; Chu, E. A History of Cancer Chemotherapy. Cancer Res. 2008, 68, 8643–8653. [Google Scholar] [CrossRef]
- Morrison, W.b. Cancer Chemotherapy: An Annotated History. J. Vet. Intern. Med. 2010, 24, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wrzosek, A.; Mika, J.; Wordliczek, J. Mechanisms of Chemotherapy-Induced Peripheral Neuropathy. Int. J. Mol. Sci. 2019, 20, 1451. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, V.; Perry, M.C. Classical chemotherapy: Mechanisms, toxicities, and the therapeutic window. Cancer Biol. Ther. 2003, 2 (Suppl. 1), S2–S4. [Google Scholar] [CrossRef]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and Radiation Therapy: Current Advances and Future Directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Hajdu, S.I.; Vadmal, M. A note from history: Landmarks in history of cancer, Part 6. Cancer 2013, 119, 4058–4082. [Google Scholar] [CrossRef]
- Givens, D.J.; Karnell, L.H.; Gupta, A.K.; Clamon, G.H.; Pagedar, N.A.; Chang, K.E.; Van Daele, D.J.; Funk, G.F. Adverse Events Associated with Concurrent Chemoradiation Therapy in Patients with Head and Neck Cancer. Arch. Otolaryngol. Neck Surg. 2009, 135, 1209–1217. [Google Scholar] [CrossRef]
- Rocha, P.H.P.; Reali, R.M.; Decnop, M.; Souza, S.A.; Teixeira, L.A.B.; Lucas, A.; Sarpi, M.O.; Cintra, M.B.; Pinho, M.C.; Garcia, M.R.T. Adverse Radiation Therapy Effects in the Treatment of Head and Neck Tumors. RadioGraphics 2022, 42, 806–821. [Google Scholar] [CrossRef]
- Majeed, H.; Gupta, V. Adverse Effects of Radiation Therapy. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2022. Available online: http://www.ncbi.nlm.nih.gov/books/NBK563259/ (accessed on 18 July 2022).
- Klement, R.J. Fasting, Fats, and Physics: Combining Ketogenic and Radiation Therapy against Cancer. Complement. Med. Res. 2018, 25, 102–113. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef]
- Vaupel, P.; Multhoff, G. Revisiting the Warburg Effect: Historical Dogma versus Current Understanding. J. Physiol. 2021, 599, 1745–1757. [Google Scholar] [CrossRef]
- Lawenda, B.D.; Kelly, K.M.; Ladas, E.J.; Sagar, S.M.; Vickers, A.; Blumberg, J.B. Should Supplemental Antioxidant Administration Be Avoided During Chemotherapy and Radiation Therapy? JNCI J. Natl. Cancer Inst. 2008, 100, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Man, A.W.C.; Li, H.; Xia, N. Impact of Lifestyles (Diet and Exercise) on Vascular Health: Oxidative Stress and Endothelial Function. Oxid. Med. Cell Longev. 2020, 2020, 1496462. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Martella, R.; Ravera, S.; Marini, C.; Capitanio, S.; Orengo, A.; Emionite, L.; Lavarello, C.; Amaro, A.; Petretto, A.; et al. Fasting induces anti-Warburg effect that increases respiration but reduces ATP-synthesis to promote apoptosis in colon cancer models. Oncotarget 2015, 6, 11806–11819. [Google Scholar] [CrossRef] [PubMed]
- Simone, B.A.; Champ, C.E.; Rosenberg, A.L.; Berger, A.C.; Monti, D.A.; Dicker, A.P.; Simone, N.L. Selectively starving cancer cells through dietary manipulation: Methods and clinical implications. Future Oncol. 2013, 9, 959–976. [Google Scholar] [CrossRef]
- Kritchevsky, D. Caloric restriction and cancer. J. Nutr. Sci. Vitaminol. 2001, 47, 13–19. [Google Scholar] [CrossRef]
- Manukian, G.; Simone, B.; DeAngelis, T.; Kivolowitz, C.; Ko, K.; Simone, N.L. Caloric Restriction Curtails Tumor Infiltrating Regulatory T Cells and Enhances Effector T Cell Activity after Radiation Therapy. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, S164. [Google Scholar] [CrossRef]
- O’Farrell, A.M.; Abrams, T.J.; Yuen, H.A.; Ngai, T.J.; Louie, S.G.; Yee, K.W.H.; Wong, L.M.; Hong, W.; Lee, L.B.; Town, A.; et al. SU11248 is a novel FLT3 tyrosine kinase inhibitor with potent activity in vitro and in vivo. Blood 2003, 101, 3597–3605. [Google Scholar] [CrossRef]
- Hursting, S.D.; Dunlap, S.M.; Ford, N.A.; Hursting, M.J.; Lashinger, L.M. Calorie restriction and cancer prevention: A mechanistic perspective. Cancer Metab. 2013, 1, 10. [Google Scholar] [CrossRef]
- Vasim, I.; Majeed, C.N.; DeBoer, M.D. Intermittent Fasting and Metabolic Health. Nutrients 2022, 14, 631. [Google Scholar] [CrossRef]
- Lugtenberg, R.T.; de Groot, S.; Kaptein, A.A.; Fischer, M.J.; Kranenbarg, E.M.K.; Dujim-de Carpentier, M.; Cohen, D.; de Graaf, H.; Heijns, J.B.; Portielje, J.E.A.; et al. Quality of life and illness perceptions in patients with breast cancer using a fasting mimicking diet as an adjunct to neoadjuvant chemotherapy in the phase 2 DIRECT (BOOG 2013–14) trial. Breast Cancer Res. Treat. 2021, 185, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.D.; Aminzadeh-Gohari, S.; Tulipan, J.; Catalano, L.; Feichtinger, R.G.; Kofler, B. Ketogenic diet in the treatment of cancer—Where do we stand? Mol. Metab. 2020, 33, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Veggiotti, P.; Cusmai, R.; Bertoli, S.; Cardinali, S.; Dionisi-Vici, C.; Elia, M.; Lispi, M.L.; Sarnelli, C.; Tagliabue, A.; et al. The Ketogenic Diet in Children, Adolescents and Young Adults with Refractory Epilepsy: An Italian Multicentric Experience. Epilepsy Res. 2002, 48, 221–227. [Google Scholar] [CrossRef]
- Marsh, E.B.; Freeman, J.M.; Kossoff, E.H.; Vining, E.P.G.; Rubenstein, J.E.; Pyzik, P.L.; Hemingway, C. The Outcome of Children with Intractable Seizures: A 3- to 6-Year Follow-up of 67 Children Who Remained on the Ketogenic Diet Less than One Year. Epilepsia 2006, 47, 425–430. [Google Scholar] [CrossRef]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bogodai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome–mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef]
- Yassin, M.A.; Ghasoub, R.S.; Aldapt, M.B.; Abdulla, M.A.; Chandra, P.; Shwaylia, H.M.; Nashwan, A.J.; Kassem, N.A.; Akiki, S.J. Effects of Intermittent Fasting on Response to Tyrosine Kinase Inhibitors (TKIs) in Patients with Chronic Myeloid Leukemia: An Outcome of European LeukemiaNet Project. Cancer Control. 2021, 28, 10732748211009256. [Google Scholar] [CrossRef]
- Schmidt, M.; Pfetzer, N.; Schwab, M.; Strauss, I.; Kämmerer, U. Effects of a ketogenic diet on the quality of life in 16 patients with advanced cancer: A pilot trial. Nutr. Metab. 2011, 8, 54. [Google Scholar] [CrossRef]
- Champ, C.E.; Palmer, J.D.; Volek, J.S.; Werner-Wasik, M.; Andrews, D.W.; Evans, J.J.; Glass, J.; Kim, L.; Shi, W. Targeting metabolism with a ketogenic diet during the treatment of glioblastoma multiforme. J. Neurooncol. 2014, 117, 125–131. [Google Scholar] [CrossRef]
- de Groot, S.; Vreeswijk, M.P.; Welters, M.J.; Gravesteijn, G.; Boei, J.J.W.A.; Jochems, A.; Hoursma, D.; Putter, H.; van der Hoeven, J.J.M.; Nortier, J.W.R.; et al. The effects of short-term fasting on tolerance to (neo) adjuvant chemotherapy in HER2-negative breast cancer patients: A randomized pilot study. BMC Cancer 2015, 15, 652. [Google Scholar] [CrossRef]
- Dorff, T.B.; Groshen, S.; Garcia, A.; Shah, M.; Tsao-Wei, D.; Pham, H.; Cheng, C.W.; Brandhorst, S.; Cohen, P.; Wei, M.; et al. Safety and Feasibility of Fasting in Combination with Platinum-Based Chemotherapy. BMC Cancer 2016, 16, 360. [Google Scholar] [CrossRef]
- Klement, R.J.; Sweeney, R.A. Impact of a ketogenic diet intervention during radiotherapy on body composition: I. Initial clinical experience with six prospectively studied patients. BMC Res. Notes 2016, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Zahra, A.; Fath, M.A.; Opat, E.; Mapuskar, K.A.; Bhatia, S.K.; Ma, D.C.; Rodman, S.N.; Snyders, T.P.; Chenard, C.A.; Eichenberger-Gilmore, J.M.; et al. Consuming a Ketogenic Diet while Receiving Radiation and Chemotherapy for Locally Advanced Lung Cancer and Pancreatic Cancer: The University of Iowa Experience of Two Phase 1 Clinical Trials. Radiat Res. 2017, 187, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Bauersfeld, S.P.; Kessler, C.S.; Wischnewsky, M.; Jaensch, A.; Steckhan, N.; Stange, R.; Kunz, B.; Bruckner, B.; Sehouli, J.; Michalsen, A. The effects of short-term fasting on quality of life and tolerance to chemotherapy in patients with breast and ovarian cancer: A randomized cross-over pilot study. BMC Cancer 2018, 18, 476. [Google Scholar] [CrossRef] [PubMed]
- Klement, R.J.; Schäfer, G.; Sweeney, R.A. A ketogenic diet exerts beneficial effects on body composition of cancer patients during radiotherapy: An interim analysis of the KETOCOMP study. J. Tradit Complement. Med. 2019, 10, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Dang, A.K.; Duong, P.T.; Phan, H.B.T.; Pham, C.T.T.; Nguyen, A.T.L.; Le, H.T. Nutrition intervention is beneficial to the quality of life of patients with gastrointestinal cancer undergoing chemotherapy in Vietnam. Cancer Med. 2021, 10, 1668–1680. [Google Scholar] [CrossRef]
- Tang, C.C.; Huang, T.C.; Tien, F.M.; Lin, J.M.; Yeh, Y.C.; Lee, C.Y. Safety, Feasibility, and Effects of Short-Term Calorie Reduction during Induction Chemotherapy in Patients with Diffuse Large B-Cell Lymphoma: A Pilot Study. Nutrients 2021, 13, 3268. [Google Scholar] [CrossRef]
- Valdemarin, F.; Caffa, I.; Persia, A.; Cremonini, A.L.; Ferrando, L.; Tagliafico, L.; Tagliafico, A.; Guijarro, A.; Carbone, F.; Ministrini, S.; et al. Safety and Feasibility of Fasting-Mimicking Diet and Effects on Nutritional Status and Circulating Metabolic and Inflammatory Factors in Cancer Patients Undergoing Active Treatment. Cancers 2021, 13, 4013. [Google Scholar] [CrossRef]
- Raffaghello, L.; Lee, C.; Safdie, F.M.; Wei, M.; Madia, F.; Bianchi, G.; Longo, V.D. Starvation-dependent differential stress resistance protects normal but not cancer cells against high-dose chemotherapy. Proc. Natl. Acad. Sci. USA 2008, 105, 8215–8220. [Google Scholar] [CrossRef]
- Otto, C.; Kaemmerer, U.; Illert, B.; Muehling, B.; Pfetzer, N.; Wittig, R.; Voelker, H.U.; Thiede, A.; Coy, J.F. Growth of human gastric cancer cells in nude mice is delayed by a ketogenic diet supplemented with omega-3 fatty acids and medium-chain triglycerides. BMC Cancer 2008, 8, 122. [Google Scholar] [CrossRef]
- Powolny, A.A.; Wang, S.; Carlton, P.S.; Hoot, D.R.; Clinton, S.K. Interrelationships between dietary restriction, the IGF-I axis, and expression of vascular endothelial growth factor by prostate adenocarcinoma in rats. Mol. Carcinog. 2008, 47, 458–465. [Google Scholar] [CrossRef]
- Colman, R.J.; Anderson, R.M.; Johnson, S.C.; Kastman, E.K.; Kosmatka, K.J.; Beasley, T.M.; Allison, D.B.; Simmons, H.A.; Kemnitz, J.W.; Wiendruch, R. Caloric Restriction Delays Disease Onset and Mortality in Rhesus Monkeys. Science 2009, 325, 201–204. [Google Scholar] [CrossRef]
- Skinner, R.; Trujillo, A.; Ma, X.; Beierle, E.A. Ketone bodies inhibit the viability of human neuroblastoma cells. J. Pediatr. Surg. 2009, 44, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Safdie, F.; Brandhorst, S.; Wei, M.; Wang, W.; Lee, C.; Hwang, S.; Conti, P.S.; Chen, T.C.; Longo, V.D. Fasting Enhances the Response of Glioma to Chemo- and Radiotherapy. PLoS ONE 2012, 7, e44603. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Simone, B.; Palazzo, J.; Savage, J.E.; Sano, Y.; Dan, T.; Jin, L.; Champ, C.; Zhao, S.; Lim, M. Caloric restriction augments radiation efficacy in breast cancer. Cell Cycle 2013, 12, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Farazi, M.; Nguyen, J.; Goldufsky, J.; Linnane, S.; Lukaesko, L.; Weinberg, A.D.; Ruby, C.E. Caloric restriction maintains OX40 agonist-mediated tumor immunity and CD4 T cell priming during aging. Cancer Immunol. Immunother. 2014, 63, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Uehara, Y.; Hosoi, Y. Activation of mTORC1 under nutrient starvation conditions increases cellular radiosensitivity in human liver cancer cell lines, HepG2 and HuH6. Biochem. Biophys. Res. Commun. 2015, 468, 684–690. [Google Scholar] [CrossRef]
- Sun, P.; Wang, H.; He, Z.; Chen, X.; Wu, Q.; Chen, W.; Sun, Z.; Weng, M.; Zhu, M.; Ma, D.; et al. Fasting inhibits colorectal cancer growth by reducing M2 polarization of tumor-associated macrophages. Oncotarget 2017, 8, 74649–74660. [Google Scholar] [CrossRef]
- Zorn, S.; Ehret, J.; Schäuble, R.; Rautenberg, B.; Ihorst, G.; Bertz, H.; Urbain, P.; Raynor, A. Impact of modified short-term fasting and its combination with a fasting supportive diet during chemotherapy on the incidence and severity of chemotherapy-induced toxicities in cancer patients—A controlled cross-over pilot study. BMC Cancer 2020, 20, 578. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, P.P.; Liu, Q.L.; Cong, M.H.; Gao, Y.; Shi, H.P.; Yu, W.N.; Miao, M.Y. Low ketolytic enzyme levels in tumors predict ketogenic diet responses in cancer cell lines in vitro and in vivo. J. Lipid Res. 2018, 59, 625–634. [Google Scholar] [CrossRef]
- Li, Y.; Dong, J.; Xiao, H.; Wang, B.; Chen, Z.; Zhang, S.; Jin, Y.; Li, Y.; Fan, S.; Cui, M. Caloric restriction alleviates radiation injuries in a sex-dependent fashion. FASEB J. 2021, 35, e21787. [Google Scholar] [CrossRef]
- Most, J.; Tosti, V.; Redman, L.M.; Fontana, L. Calorie restriction in humans: An update. Ageing Res. Rev. 2017, 39, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Norman, K.; Stobäus, N.; Zocher, D.; Bosy-Westphal, A.; Szramek, A.; Scheufele, R.; Smoliner, C.; Pirlich, M. Cutoff percentiles of bioelectrical phase angle predict functionality, quality of life, and mortality in patients with cancer. Am. J. Clin. Nutr. 2010, 92, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, M.; Roychoudhuri, R.; Restifo, N.P. Nutrient Competition: A New Axis of Tumor Immunosuppression. Cell 2015, 162, 1206–1208. [Google Scholar] [CrossRef] [PubMed]
- Klement, R.J.; Pazienza, V. Impact of Different Types of Diet on Gut Microbiota Profiles and Cancer Prevention and Treatment. Medicina 2019, 55, 84. [Google Scholar] [CrossRef] [PubMed]
- Brandhorst, S.; Longo, V.D. Fasting and Caloric Restriction in Cancer Prevention and Treatment. In Metabolism in Cancer; Recent Results in Cancer Research; Cramer, T., Schmitt, C.A., Eds.; Springer: Cham, Switzerland, 2016; Volume 207, pp. 241–266. [Google Scholar] [CrossRef]
- Nikolai, S.; Pallauf, K.; Huebbe, P.; Rimbach, G. Energy restriction and potential energy restriction mimetics. Nutr. Res. Rev. 2015, 28, 100–120. [Google Scholar] [CrossRef][Green Version]
- Vidoni, C.; Ferraresi, A.; Esposito, A.; Maheshwari, C.; Dhanasekaran, D.N.; Mollace, V.; Isidoro, C. Calorie Restriction for Cancer Prevention and Therapy: Mechanisms, Expectations, and Efficacy. J. Cancer Prev. 2021, 26, 224–236. [Google Scholar] [CrossRef]
- Caffa, I.; Spagnolo, V.; Vernieri, C.; Valdemarin, F.; Becherini, P.; Wei, M.; Brandhorst, S.; Zucal, C.; Driehuis, E.; Ferrando, L.; et al. Fasting-Mimicking Diet and Hormone Therapy Induce Breast Cancer Regression. Nature 2020, 583, 620–624. [Google Scholar] [CrossRef]
- Riedinger, C.J.; Kimball, K.J.; Kilgore, L.C.; Bell, C.; Heidel, E.; Boone, J.D. Water-only fasting and its effect on chemotherapy administration in gynecologic malignancies. Gynecol. Oncol. 2020, 159, 13. [Google Scholar] [CrossRef]
- Kozubík, A.; Pospíšil, M. Adaptation to intermittent fasting as a factor modifying the radiation resistance of mice. Experientia 1982, 38, 958–959. [Google Scholar] [CrossRef]
- Courtnay, R.; Ngo, D.C.; Malik, N.; Ververis, K.; Tortorella, S.M.; Karagiannis, T.C. Cancer metabolism and the Warburg effect: The role of HIF-1 and PI3K. Mol. Biol. Rep. 2015, 42, 841–851. [Google Scholar] [CrossRef]
- O’Flanagan, C.H.; Smith, L.A.; McDonell, S.B.; Hursting, S.D. When less may be more: Calorie restriction and response to cancer therapy. BMC Med. 2017, 15, 106. [Google Scholar] [CrossRef] [PubMed]
- Icard, P.; Ollivier, L.; Forgez, P.; Otz, J.; Alifano, M.; Fournel, L.; Loi, M.; Thariat, J. Perspective: Do Fasting, Caloric Restriction, and Diets Increase Sensitivity to Radiotherapy? A Literature Review. Adv. Nutr. 2020, 11, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Cho, G.S.; Park, Y.H.; Kim, S.K. Comparison of Quality of Life and Nutritional Status in Gastric Cancer Patients Undergoing Gastrectomies. Clin. Nutr Res. 2015, 4, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Safdie, F.M.; Dorff, T.; Quinn, D.; Fontana, L.; Wei, M.; Lee, C.; Cohen, P.; Longo, V.D. Fasting and cancer treatment in humans: A case series report. Aging 2009, 1, 988–1007. [Google Scholar] [CrossRef]
- Lo Re, O.; Panebianco, C.; Porto, S.; Cervi, C.; Rappa, F.; Di Biase, S.; Caraglia, M.; Pazienza, V.; Vinciguerra, M. Fasting inhibits hepatic stellate cells activation and potentiates anti-cancer activity of Sorafenib in hepatocellular cancer cells. J. Cell Physiol. 2018, 233, 1202–1212. [Google Scholar] [CrossRef]
- Paoli, A.; Rubini, A.; Volek, J.S.; Grimaldi, K.A. Beyond weight loss: A review of the therapeutic uses of very-low-carbohydrate (ketogenic) diets. Eur. J. Clin. Nutr. 2013, 67, 789–796. [Google Scholar] [CrossRef]
- Plotti, F.; Terranova, C.; Luvero, D.; Bartolone, M.; Messina, G.; Feole, L.; Cianci, S.; Scaletta, G.; Marchetti, C.; Di Donato, V.; et al. Diet and Chemotherapy: The Effects of Fasting and Ketogenic Diet on Cancer Treatment. Chemotherapy 2020, 65, 77–84. [Google Scholar] [CrossRef]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; Le Moan, N.; Gruetter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 2013, 339, 211–214. [Google Scholar] [CrossRef]
- Vergara, N.; Montoya, J.E.; Luna, H.G.; Amparo, J.R.; Cristal-Luna, G. Quality of Life and Nutritional Status Among Cancer Patients on Chemotherapy. Oman Med. J. 2013, 28, 270–274. [Google Scholar] [CrossRef]
- Berrill, J.W.; Turner, J.K.; Hurley, J.J.; Swift, G.; Dolwani, S.; Green, J.T. Upper gastrointestinal cancer in its early stages is predominantly asymptomatic. Frontline Gastroenterol. 2012, 3, 47–51. [Google Scholar] [CrossRef]
- Muscaritoli, M.; Arends, J.; Bachmann, P.; Baracos, V.; Barthelemy, N.; Bertz, H.; Bozzetti, F.; Hütterer, E.; Isenring, E.; Kaasa, S.; et al. ESPEN Practical Guideline: Clinical Nutrition in Cancer. Clin. Nutr. 2021, 40, 2898–2913. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.A.; Ingram, D.K.; Roth, G.S. 2-Deoxy-D-Glucose Feeding in Rats Mimics Physiologic Effects of Calorie Restriction. J. Anti-Aging Med. 1998, 1, 327–337. [Google Scholar] [CrossRef]
- Minor, R.K.; Smith, D.L.; Sossong, A.M.; Kaushik, S.; Poosala, S.; Spangler, E.L.; Roth, G.S.; Lane, M.; Allison, D.B.; de Cabo, R.; et al. Chronic ingestion of 2-deoxy-D-glucose induces cardiac vacuolization and increases mortality in rats. Toxicol. Appl. Pharmacol. 2010, 243, 332–339. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Author | Year Published | Years Conducted | Patient Population | Type(s) of Cancer | Treatment Regimen | Diet Regimen | Outcome: QoL | Outcome: Therapy Toxicity/Efficacy |
|---|---|---|---|---|---|---|---|---|
| Safdie et al. [27] | 2009 | April 2008–August 2009 | n = 10 | Breast, esophageal, prostate, lung, ovary, uterus | CT | IF (fasting 48–140 h before) | Six fasting patients reported reduction in fatigue, weakness, and gastrointestinal side effects. | NA |
| Schmidt et al. [28] | 2011 | Could not determine | n = 16 | Breast, esophageal, prostate, lung, ovary, granulosa cell tumor, parotid carcinoma, osteosarcoma (jaw) thyroid carcinoma, colon carcinoma, endometrial | CRT | KD | Five out of sixteen patients completed the study and one dropped out due to diet intolerance. No severe adverse effects were observed, and insomnia improved. | Blood cholesterol was reduced significantly. Blood leukocytes increased significantly. |
| Champ et al. [29] | 2014 | August 2010–April 2013 | n = 53 (6 underwent dieting) | Glioma | CRT | KD | NA | Serum glucose levels were reduced, and diet was safe/feasible. |
| de Groot et al. [30] | 2015 | May 2011–December 2012 | n = 13 | Breast | CT | IF (48 h short-term fast) | NA | Mean erythrocyte and thrombocyte counts were higher in the IF group. Non-hematological toxicity did not significantly differ, but γ-H2AX levels were significantly higher in CD45 + CD3- cells in the non-IF group. |
| Dorff et al. [31] | 2016 | October 2009–November 2012 | n = 20 | Urothelial, NSCLC, ovarian, uterine, breast | CT | IF (24 h, 48 h, and 72 h STS) | Tolerance to fasting was assessed and limited to ≤grade 2 dizziness, fatigue, and headache. Symptoms tended to increase as fasting period increased. | DNA damage in leukocytes for patients who fasted 48 h or more was reduced. IGF-1 levels decreased by 30%, 33%, and 8% in the 24 h, 48 h, and 72 h cohorts, respectively. |
| Klement et al. [32] | 2016 | 2014 | n = 6 | Breast, prostate, rectum, primary lung | CRT | KD | QoL was stable and good through CRT, but eventually decreased, possibly from prolonged radiation exposure. Some patients experienced significant weight loss with KD at a caloric deficit. | Tumor regression occurred as expected in 5 of 6 patients, with 1 seeing slight progression of small cell lung cancer. Once KD ended, their disease progressed rapidly. |
| Zahra et al. [33] | 2017 | July 2011–June 2014 | n = 9 | Lung, pancreatic | CRT | KD | 2 of 7 lung cancer patients and 1 of 2 pancreatic cancer patients maintained KD. All who dropped out noted compliance difficulties. | Compliance was 33%, too difficult to assess outcome. |
| Bauersfeld et al. [34] | 2018 | November 2013–August 2015 | n = 34 | Breast, ovarian | CT | IF (started 36 h before and ended 24 h after) | FACT-G and FACIT-F scores increased in the IF group compared to the non-IF group. Fasting QoL scores increased while non-fasting QoL scores decreased. QoL scores in cross-over fasting groups remained similar. | NA |
| Klement et al. [35] | 2019 | Could not determine | n = 81 | Head and neck, rectum, breast | RT | KD (also supplemented with 10 g of essential amino acids on days with RT) | No significant adverse effects reported. | In rectal and breast cancer patients, KD significantly associated with fat mass loss. No significant changes in nonfat and skeletal muscle mass occurred. HNC patients who were under CRT had KD-associated gain in fat-free and skeletal muscle mass |
| Nguyen et al. [36] | 2021 | 2016–2019 | n = 120 | Gastrointestinal | CT | Nutritional Intervention | Maintenance of adequate nutrition in GI cancer patients saw significantly higher QoL assessment scores compared to control groups. | Nutritional intervention was associated with significantly better treatment prognoses. |
| Tang et al. [37] | 2021 | October 2020–July 2021 | n = 12 | Diffuse Large B-cell lymphoma | CT | CR (short term) | Diet was tolerated and compliance was high. Mean phase angle increased compared to the baseline. | CR intervention was associated with higher erythrocyte and lymphocyte counts. |
| Valdemarin et al. [38] | 2021 | November 2017–August 2020 | n = 90 | 18 different types of cancer, plurality was breast | CRT | CR/IF (fasting-mimicking) | Diet was found safe/feasible. Increase in phase angle and fat-free mass was noted while fat mass decreased. | Serum c peptide, IGF-1, IGFBP3 and leptin levels decreased while IGFBP1 increased. |
| Author | Year Published | Animal/Cellular Model | Type(s) of Cancer | Sample Size | Condition Tested | Outcome |
|---|---|---|---|---|---|---|
| Raffaghello et al. [39] | 2008 | S. cerevisiae with RAS2 and/or SCH9 deletion. Rat cell lines: primary rat mixed glial cells (astrocytes + 5–10% microglia), C6, RG2, A10-85, and 9 L Human cell lines: LN229, SH-SY5Y | Glioma, neuroblastoma | ~260 million S. cerevisiae cells | 48-h STS in S. cerevisiae; noncontrol cells were treated with either hydrogen peroxide or menadione. MMS and CP were used to mimic chemotherapy treatment. | STS cells with RAS2/SCH9 deletions were up to 1000-fold more resistant to oxidizing treatments. Expression of the oncogene-like RAS2^(Val19) negated this protection. |
| Rat cell lines: primary rat mixed glial cells (astrocytes + 5–10% microglia), C6, RG2, A10-85, and 9 L Human cell lines: LN229, SH-SY5Y | Glioma, neuroblastoma | 5 different in vitro cell lines | Concentration of glucose in experimental group was reduced to mimic STS, STS induction of DSR in mammalian cell lines, cells were treated with H2O2, menadione, and CP | 80% STS survival rate vs. <10% for control glial cells. No difference between cell survival in H2O2, but STS induced DSR in normal cells treated with methadione, causing cancer cell sensitization/lower survival rates. No difference was observed in CP treated STS cells. | ||
| A/J mice | NA | n = 30, 17 under STS, 23 AL; n = 10 for 60 h STS | Mouse models were injected with EtO to generate cytotoxic conditions, 80 mg/kg dose of EtO was administered | 43% of control mice died by day 10; only 1 of 17 STS-treated mice died after EtO treatment. STS mice showed no visible signs of stress or pain after EtO treatment; control mice showed reduced mobility and increased stress. No STS mice died in 60 h STS trials. | ||
| A/J mice | Neuroblastoma | n = 46 | Mouse models were inoculated with NXS2 cells. Survival rates of 48 h STS, non-STS, and STS/EtO administered mice were compared. | NXS2/STS/EtO had a significantly larger survival rate than NXS2/STS or NSX2 mice | ||
| Otto et al. [40] | 2008 | NMRI strain mice | Gastric adenocarcinoma | n = 24 (total), n = 12 (KD group), n = 12 (standard diet) | Mice were injected subcutaneously with cell line 23132/87 and randomly split into KD or standard diet groups. | KD group tumors had significantly delayed growth. After day 20, KD tumor growth increased, yet remained significantly lower. KD group tumors had significantly larger necrotic areas. |
| Powolny et al. [41] | 2008 | Copenhagen rats | Prostate adenocarcinoma | n = 10 (total), n = 5 (AL), n = 5 (40% CR) | Rats were fed either AL or 40% CR. After 8 weeks, rats were implanted with AT6.3 cells. 2 weeks after, serum concentrations and protein expression of IDF-I, IGFBP2, and VEGF were measured. | CR reduced IGF-1 (35%) and increased IGBP3 by 7-fold. mRNA expression of IGF-1 and its receptor, as well as VEGF mRNA/protein secretion, were significantly lowered. CR reduced endocrine/autocrine IGF-1 expression, contributing to reduced VEGF/angiogenesis. |
| Colman et al. [42] | 2009 | Rhesus macaques | NA | n = 76 (total), n = 30 (1st cohort), n = 46 (2nd cohort) | Rhesus macaques were studied longitudinally over the course of 20 years, with some undergoing moderate CR and compared to a control-fed group. | 50% (control) vs. 80% (CR) overall survival |
| Skinner et al. [43] | 2009 | Human foreskin fibroblasts (control) and human neuroblastoma cells (SK-N-AS) | Neuroblastoma | 1 control and 1 neuroblastoma cell line | 2 different cell lines were grown in standard media with or without glucose (glc +/glc-), glc- with acetoacetate, or glc- with BHB. This was to determine cell viability in ketone-rich environments. | Neuroblastoma viability decreased significantly in BHB and acetoacetate media. Inability for neuroblastoma cells to utilize ketone bodies was likely due to decreased expression of the SCOT protein. |
| Safdie et al. [44] | 2012 | Primary mouse glia, murine GL26, rat C6, and human U251, LN229, and A172 glioma cells | Neuroblastoma | 6 different cell lines: 2 mouse, 1 rat, and 3 human | TMZ was tested in both AL and STS-mimicking conditions | STS-mimicking sensitized glioma cell lines, not glial cells, to TMZ. |
| C57BL/6N murine models injected with GL26 | Neuroblastoma | n = 30 (subcutaneous), n = 33 (intracranial) | TMZ administered in both STS and AL conditions on in vivo murine models with GL26 injected both subcutaneously and intracranially | 48 h STS caused a significant decrease in blood glucose and IGF-1 levels and significant sensitization to CRT treatment. | ||
| Saleh et al. [45] | 2013 | Balb/c murine models | Triple negative breast (TNBC) | n = 80 | 67NR/4TI cell lines were injected into mice part of either a standard diet group, or an ADF group. Desired CR was not achieved, so a second cohort of mice were injected with 4TI TNBC cells. All groups were subject to RT. | ADF caused tumor growth delays in 67NR and 4TI tumors. CR + RT saw the greatest growth delay. No difference between the first and second ADF cohorts was specified. |
| Farazi et al. [46] | 2014 | Balb/c and C57BL/6 murine models | Mouse fibrosarcoma | NA | Aged mice were placed on CR or dietary supplementation with RES, a CR mimetic. Tumor immune responses were induced and observed starting at 4 months until the mice reached ~12 months in age. | CR resulted in fully sustained OX40-mediated anti-tumor immunity and antigen-specific CD4 T cell priming in aged hosts. RES supplementation was unable to mimic such effects. |
| Murata et al. [47] | 2015 | HepG2 and HuH6 cell lines; SV-0-transformed human fibroblast line (LM217) | Hepatoma | 3 cell lines | Nutrient starvation was induced in all 3 cell lines to observe effects on mTORC1 activity and other cellular signaling pathways. | mTORC1 activity was suppressed in LM217 cells under nutrient starvation. Hep2 and HuH6 cells increased in mTORC1 activity and radiosensitivity. mTOR inhibition by siRNA/rapamycin suppressed this in HepG2. |
| Bianchi et al. [15] | 2015 | CT26 colorectal tumor cells, Balb/c murine models | Colorectal | 1 cell line, n = 28 | Cells were subject to 48 h STS or given standard nutrients. In either group, half were given OXP. The same analysis was performed on mouse models, with CT26 injections in their lower back. | In both in vivo and in vitro models, STS was shown to exhibit an anti-Warburg effect, causing tumor cells to move into an uncoupled oxidative phosphorylation model of metabolism. |
| Sun et al. [48] | 2017 | CT26 colorectal tumor cells, Balb/c murine models | Colorectal | n = 12, n = 6(ADF), n = 6 (control) | M2 TAMs promote cancer cell proliferation. Experimental in vitro and in vivo models were both placed under fasting conditions and compared to control cell lines/mouse models. | Fasting was shown to inhibit tumor growth via the reduction in M2 polarization of TAMs in mouse models. In vitro evidence reinforced this. Fasting was shown to induce cancer cell autophagy. |
| Zahra et al. [33] | 2017 | MIA PaCa-2 injected athymic nu/nu murine models | Pancreatic | n = 6 (total), n = 3 (standard chow), n = 3 (4:1 KetoCal) | Mice injected with MIA PaCa-2 cells were fed 2 different diets: a standard isocaloric diet and a specific KetoCal diet (KD analogue). Mice were then exposed to 6 fractions of 2 Gy for 25 days. Tumor progression was measured. | KD enhanced radiation response in mouse xenograft models without increasing radiation toxicity. |
| Lo Re et al. [49] | 2017 | LX-2 human hepatic stellate cell (HSC) line | NA | 1 cell line | LX-2 cells were exposed to either NM or FM, and some were then subsequently treated with LPS. | LX-2 cells exposed to FM had inhibited DNA synthesis/proliferation rates. LPS managed to increase LX-2 cell proliferation by, regardless of media. |
| C57BL/6J murine models | NA | n = 24 (total), n = 12 (72 h fast), n = 12 (control) | Mice were administered streptozotocin to simulate human diabetes and non-alcoholic steatohepatitis (NASH) in vivo. One group of mice fasted for 72 h, which was followed by 10 days of high fat diet refeeding. | Cyclic fasting did not have beneficial effect in reducing fibrotic burden of NASH in the model. | ||
| HepG2 and Huh-7 cell lines | Hepatoma | 2 cell lines | Cytostatic ability of sorafenib was assessed for HepG2 and Huh-7 cells in either starvation-mimicking (72 h fast) or non-fasting conditions. | Fasting synergized with and increased efficacy of the anti-proliferative effects of Sorafenib. | ||
| Zhang et al. [50] | 2018 | Various human cancer cell lines | Glioma, leukemia, pancreatic, thyroid, cervical, liver, renal, lung, gastric, colorectal, prostate, breast, and ovarian | 33 cell lines | Expression levels of 4 enzymes were examined: BDH 1/2 and OXCT 1/2. | No correlation between glycolysis reliance and ketolytic enzyme quantity. BHB-supplemented HeLa cells saw improved growth rates; PANC-1 saw no change. BHB no longer benefitted HeLa growth with BDH1/OXCT1 downregulation via siRNA. |
| nu/nu murine models | Pancreatic | NA | BDH1/2 and OXCT1/2 levels were investigated in PANC-1 and HeLa xenograft tumors. | In vivo KD results were consistent with in vitro data. PANC-1 xenograft tumor growth rate was heavily attenuated compared to HeLa. Mean survival of KD HeLa mice was significantly lower than that of standard diet HeLa mice. | ||
| Manukian et al. [18] | 2019 | Balb/c murine models | TNBC | NA | Mouse models were injected with 4TI cells and randomized into AL or CR. CR mice were treated with either only CR or CR + RT (12 Gy in 3 fractions). | Increased expression activity related to integrin signaling and inflammation pathways was observed. CD4 + CD25 + Foxp3+ Tregs were significantly decreased with CR. Effector T cells from CR mice produced significantly more interferon-y than AL controls. |
| Li et al. [51] | 2021 | C57BL/6J murine models | NA | NA | Mouse models were fed ad libitum or subjected to 30% CR before TBI or TAI treatment. | CR improved intestinal architecture and restored hematogenic organs in all mice. Male mice noted improved gut barrier function; female mice had mitigated systemic and enteric inflammation. There were notable differences in gut microbiome bacterial species enrichment in either sex. |
| ClinicalTrials.gov Identifier | Responsible Party | Status of Trial | Trial Start Date | Trial Completion Date | Patient Eligibility Criteria | Type(s) of Cancer | Treatment Regimen | Diet | Outcome Measure(s) |
|---|---|---|---|---|---|---|---|---|---|
| NCT05359848 | Roberto Pili, State University of New York at Buffalo | Recruiting | 15 April 2022 | 1 November 2023 | Adult cancer patients eligible for chemotherapy with adequate hematologic, renal, and liver function with >6 months life expectancy | any | CT | CR/IF | Feasibility and compliance of IF/CR plant-based diet in CT patients |
| NCT03535701 | Jeff Volek, Ohio State University Comprehensive Cancer Center | Completed | 20 October 2017 | 1 May 2020 | Adult; BMI ≥ 22; FDG-18 PET avid tumor; life expectancy >6 m; ECOG performance status 0–1 | metastatic or stage IV breast cancer | CT | KD | Feasibility and compliance to KD (tumor progression and biologic and behavioral health markers) |
| NCT03591861 | Washington University School of Medicine | Recruiting | 1 May 2019 | 31 May 2034 | <21 y/o; life expectancy >12 w; Karnofsky/Lansky performance score ≥60 | recurrent primary brain tumor with no curative therapy | CT | KD | Feasibility of combining KD with CT in children with relapsed brain tumors |
| NCT05234502 | Mehmet Artac, Necmettin Erbakan University | Not yet recruiting | 30 January 2022 | 30 January 2025 | 19–64 y/o; female; Dx with breast cancer 1st time; plan to receive neoadjuvant treatment; BMI 25–29.9 or ≥30; Karnofsky Performance Score ≥70 | breast cancer | CT | KD | Interventional; Effects of KD on metabolism and polyneuropathy on patients receiving neoadjuvant CT (polyneuropathy, polyneuropathy severity of symptoms, tumor size, QoL, 5-year survival rate) |
| NCT04631445 | Translational Drug Development | Recruiting | 2 December 2020 | 1 June 2023 | adult; Karnofsky Performance Status of ≥70; Life expectancy ≥12 w | metastatic pancreatic ductal adenocarcinoma | CT | KD | Evaluate the progression free survival, changes in serum metabolites and quality of life (KD + triplet therapy) |
| NCT01535911 | Kenneth Schwartz, MD, Michigan State University | Active, not recruiting | 1 April 2012 | 1 June 2024 | 18–90 y/o; ECOG performance status ≤2; Life expectancy >3 m | GBM | CRT | Energy restricted KD | Effect of KD with RT/CT on tumor size |
| NCT04730869 | Waikato Hospital | Recruiting | 26 May 2021 | 1 November 2022 | ≥18 y/o; ECOG Performance Status 0–2 | GBM | CRT | KD/IF | Feasibility, safety, and efficacy of a Metabolic Therapy Program in Conjunction with Standard Treatment for Glioblastoma Multiforme |
| NCT05428852 | Jeff Volek, Ohio State University Comprehensive Cancer Center | Recruiting | 1 July 2022 | 1 July 2024 | 18–75 y/o; Graded Prognostic Assessment >1.5; BMI ≥ 18; ECOG performance status 0–1 | brain metastasis | RT | KD | Feasibility, metabolic outcome, effect on cognitive function, QOL of KD versus AICR diet on brain metastasis patient selected for radiosurgery or undergoing RT |
| NCT03451799 | Jethro Hu, Cedars-Sinai Medical Center | Active, not recruiting | 13 April 2018 | 1 October 2023 | ≥18 y/o; Karnofsky Performance Status >70; BMI >22 | GBM | CRT | KD | Safety and feasibility of KD in combination with standard-of-care radiation and temozolomide for patients with glioblastoma |
| NCT02302235 | Mid-Atlantic Epilepsy and Sleep Center, LLC | Completed | 1 February 2014 | 1 May 2022 | 18–65 y/o; Documented surgical resection/debulking; Karnovsky Performance Score ≥70 | GBM | CRT | KD | Efficacy, safety, tolerability of KD as adjunctive treatment of RT in GBM |
| NCT05327608 | Thomas Jefferson University | Recruiting | 28 July 2022 | 1 May 2024 | ≥18 y/o; BMI ≥ 25; HER2-negative; ER/PR <10%; clinical stage I-III; size ≥1.5 cm; | primary invasive breast carcinoma | CT | IF | Feasibility of IF for patients with HER2- negative and ER/PR <10% breast cancer and body mass index ≥ 25 receiving neoadjuvant CT |
| NCT03162289 | Andreas Michalsen, Charite University, Berlin, Germany | Recruiting | 10 May 2017 | 10 June 2022 | 18–75 y/o; conventional adjuvant or neo-adjuvant protocol with at least 4 CT cycles | non-metastatic ovarian or breast cancer | CT | IF | Tolerance and effectiveness of CT through accompanying intense nutritional therapy interventions |
| NCT05259410 | Kelsey Nicole Dipman Gabel, University of Illinois at Chicago | Not yet recruiting | 1 July 2022 | 1 March 2026 | 18–99 y/o; ECOG 0 or 1; BMI 25–40 | Breast cancer (histologically confirmed Stage I-III) | CT | IF | Feasibility, safety, and efficacy of time-restricted eating combined with CT during breast cancer treatment |
| NCT02710721 | Andreas Michalsen, Charite University, Berlin, Germany | Active, not recruiting | 1 April 2016 | 1 December 2021 | 25–89 y/o male; BMI ≥ 20 | CRPC or hormone-sensitive prostate cancer with high metastatic load | CT | IF | Efficacy of fasting and nutritional therapy as a complementary treatment of advanced metastatic prostate cancer undergoing CT |
| NCT05503108 | J.R. Kroep, Leiden University Medical Center | Not yet recruiting | 15 October 2022 | 15 April 2032 | ≥18 y/o; WHO performance status 0–2; Clinical stage II-III (cT1cN+ or ≥T2 any cN, cM0); BMI ≥ 18.5 | HR+, HER2- Breast Cancer | CT | IF | Response rate, effect, side effects and QoL of IF during neoadjuvant CT |
| NCT04626843 | Eleah Stringer, British Columbia Cancer Agency | Active, not recruiting | 3 February 2021 | 31 December 2022 | 19–85 y/o; BMI ≥ 20; ECOG Performance Status ≤2; Lymphocytes ≥40 and <150; Hb ≥ 100 g/L | CLL and SLL | CT | IF | Feasibility and efficacy of IF in CLL/SLL Patients |
| NCT05023967 | National Cancer Institute (NCI) | Not yet recruiting | 30 January 2023 | 30 November 2024 | ≥18 y/o; ECOG performance status ≤ 1 (Karnofsky ≥ 70%); BMI >18.5 | Invasive Breast Cancer or DCIS | CT | IF | Combined effect of prolonged nightly fasting and metformin in decreasing breast tumor cell proliferation and other biomarkers of breast cancer |
| NCT01819233 | Thomas Jefferson University (Sidney Kimmel Cancer Center at Thomas Jefferson University) | Completed | 8 March 2013 | 1 September 2019 | ≥18 y/o; female; BMI ≥ 21; Weight ≥ 100 lbs | DCIS or invasive breast cancer | RT | CR | Feasibility of RT with CR for the treatment of breast cancer; measurable changes of patient characteristics and tissue and serum from CR conditions to determine a metric for evaluating this treatment in future studies |
| NCT04959474 | Thomas Jefferson University | Recruiting | 23 August 2021 | 1 January 2026 | ≥40 y/o; BMI ≥ 21; KPS score 70–100; tumor size ≤ 3.0 cm; unifocal | DCIS or invasive breast cancer | RT | CR | Compare percent reduction in cellularity of breast tumor between pre-operative SABR with and without CR |
| NCT01802346 | University of Southern California | Recruiting | 29 January 2013 | 29 January 2024 | ≥19 y/o; BMI ≥ 18.5; ECOG performance status 0–1 | breast cancer or metastatic prostate adenocarcinoma | CT | CR | Impact on toxicity and efficacy of CR prior to CT in treatment of breast and prostate cancer |
| NCT02827370 | Thomas Jefferson University (Sidney Kimmel Cancer Center at Thomas Jefferson University) | Active, not recruiting | 16 June 2016 | 1 April 2022 | ≥18 y/o; female; BMI ≥ 21; Weight ≥ 120 lbs; Karnofsky Performance Status of 80–100% | invasive breast cancer (Non-metastatic and non-inflammatory) | CT | CR | CR impact on pathologic complete response rate, incidence of adverse events and efficacy of CT for breast cancer |
| NCT03340935 | Filippo de Braud, Fondazione IRCCS Istituto Nazionale dei Tumori, Milano | Completed | 1 February 2017 | 31 July 2020 | ≥18 y/o; BMI ≥ 20 | any, except small cell neuroendocrine tumors | standard adjuvant treatments or therapies for advanced disease | CR | Safety, feasibility and metabolic effects of the fasting-mimicking diet (FMD) in cancer patients |
| NCT05082519 | Etan Orgel, Therapeutic Advances in Childhood Leukemia Consortium | Recruiting | 12 March 2022 | 15 October 2031 | 10–25 y/o; Karnofsky > 50% for patients > 16 y/o and Lansky > 50% for patients ≤16 y/o; BMI% ≥ 5th percentile for age for patients aged 10–19 years, BMI ≥ 18.5 in patients 20–29 years | B-ALL | CT | CR | Efficacy of the IDEAL2 (Improving Diet and Exercise in ALL) CR and activity intervention integrated into HR B-ALL induction to reduce incidence of end of induction MRD ≥ 0.01%; efficacy of the IDEAL2 intervention to reduce gain in fat mass during induction |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercier, B.D.; Tizpa, E.; Philip, E.J.; Feng, Q.; Huang, Z.; Thomas, R.M.; Pal, S.K.; Dorff, T.B.; Li, Y.R. Dietary Interventions in Cancer Treatment and Response: A Comprehensive Review. Cancers 2022, 14, 5149. https://doi.org/10.3390/cancers14205149
Mercier BD, Tizpa E, Philip EJ, Feng Q, Huang Z, Thomas RM, Pal SK, Dorff TB, Li YR. Dietary Interventions in Cancer Treatment and Response: A Comprehensive Review. Cancers. 2022; 14(20):5149. https://doi.org/10.3390/cancers14205149
Chicago/Turabian StyleMercier, Benjamin D., Eemon Tizpa, Errol J. Philip, Qianhua Feng, Ziyi Huang, Reeny M. Thomas, Sumanta K. Pal, Tanya B. Dorff, and Yun R. Li. 2022. "Dietary Interventions in Cancer Treatment and Response: A Comprehensive Review" Cancers 14, no. 20: 5149. https://doi.org/10.3390/cancers14205149
APA StyleMercier, B. D., Tizpa, E., Philip, E. J., Feng, Q., Huang, Z., Thomas, R. M., Pal, S. K., Dorff, T. B., & Li, Y. R. (2022). Dietary Interventions in Cancer Treatment and Response: A Comprehensive Review. Cancers, 14(20), 5149. https://doi.org/10.3390/cancers14205149