
In Vitro and In Vivo Models of CLL–T Cell Interactions: Implications for Drug Testing


Abstract
Simple Summary
Abstract
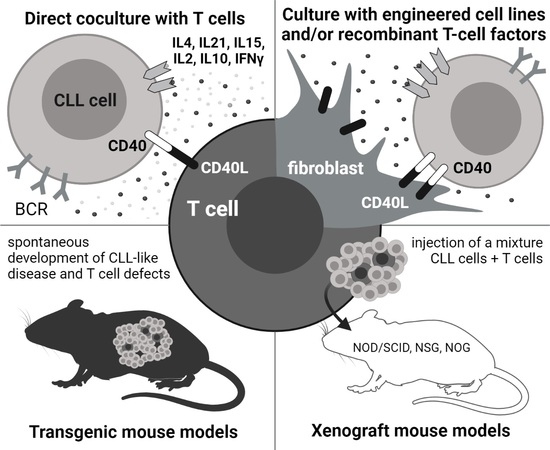
1. Introduction
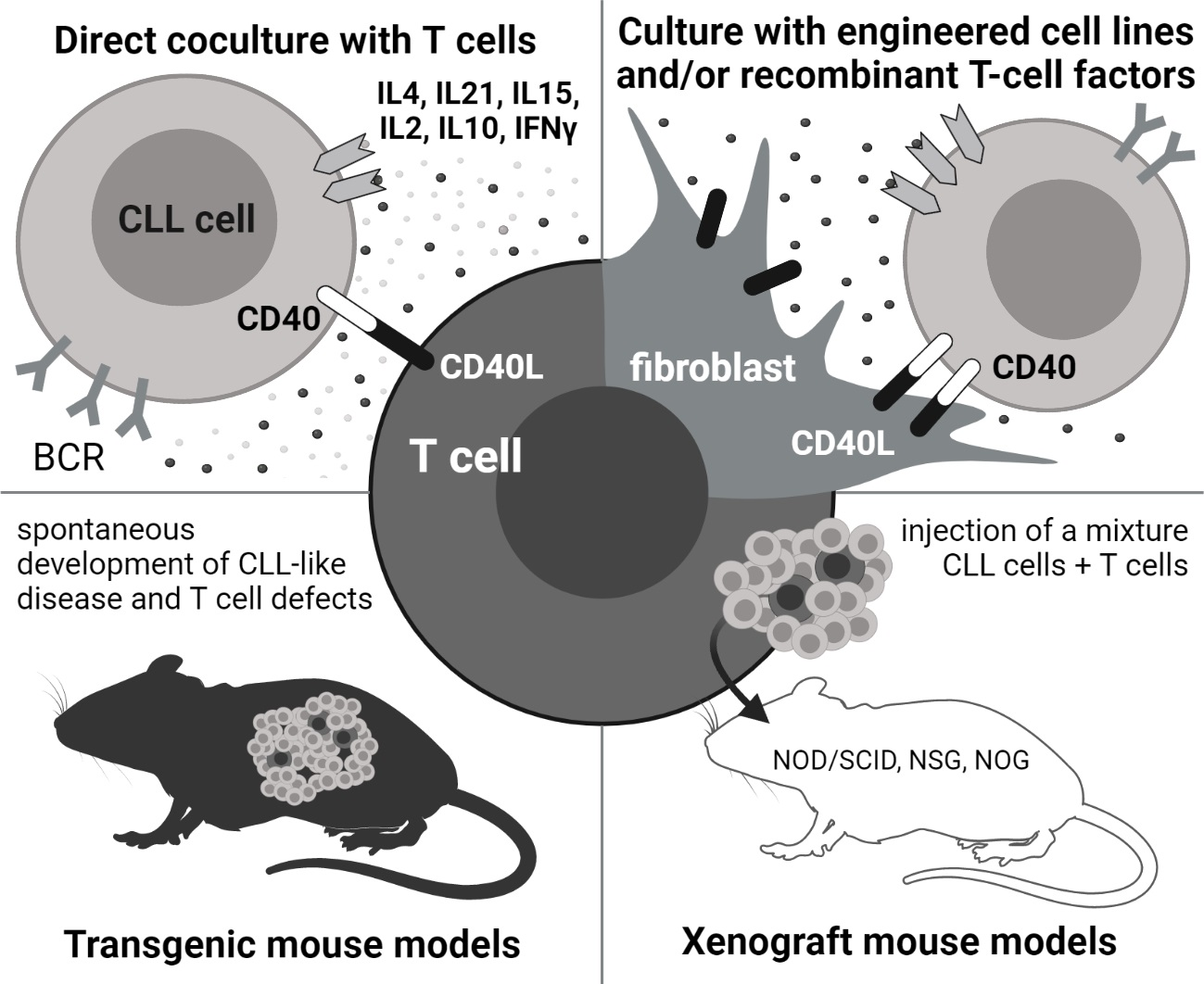
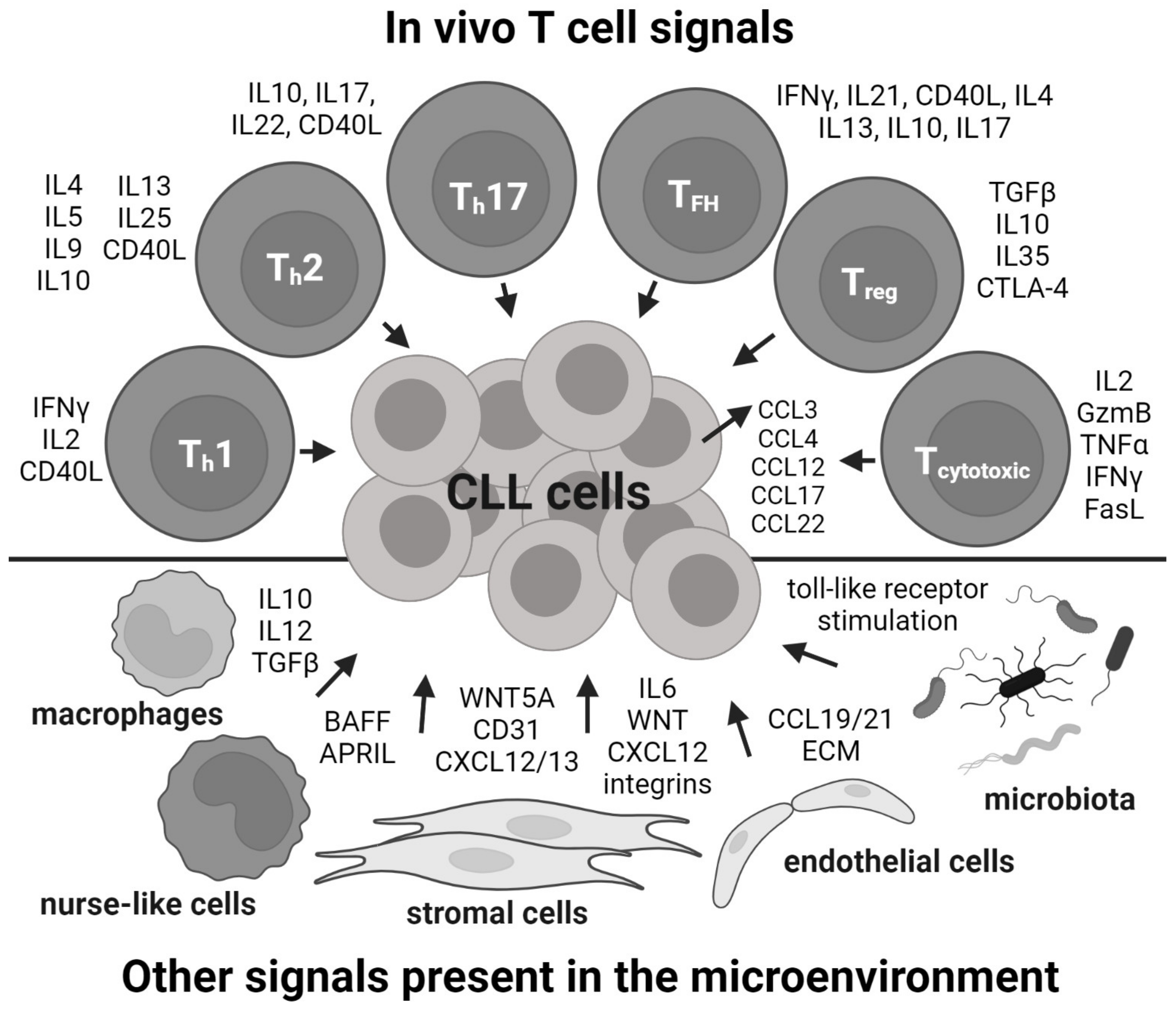
2. In Vitro: Co-Culture with T Cells

| Stimulatory T Cells | T:CLL Cells Ratio | Duration [Days] | Purification | CLL Viability | CLL Proliferation | Ref. |
|---|---|---|---|---|---|---|
| irradiated autologous/allogenic CD4+ T cells activated by anti-CD3, CLL co-stimulation by IL2 and IL10 | - | 3 (up to 9) | CLL | ~100% (~50% *) | [3H]TdR uptake, proliferation dependent on T–CLL cell contact | [35] |
| autologous activated CD3+ T cells | 1:3 | 14 | CLL | 5–20% (5–20% *) | - | [39] |
| autologous activated CD3+ T cells | 1:4 | 4–6 | PBMCs CD3-depleted | not affected | CSFE highly variable; absolute increase in number of CD19+ cells | [36] |
| autologous non-activated T cells | same as in original PBMCs | 7 | PBMCs | ~75% (~50% *) | - | [12] |
| autologous T cells and CD40L- expressing mouse fibroblasts | - | 54–154 | PBMCs | - | Ki67 (significantly increased at day 14 compared to day 0) | [40] |
| autologous activated CD4+ T cells | 1:5 | 2–6 | CLL | - | Ki67 (day 2; 2% vs. 1% in control), CSFE (day 6; 3% vs. 2% in control) | [9] |
| autologous activated T cells | 2:1 | 1 or 3 h | CLL | - | - | [33] |
3. In Vitro: Mimicking T Cells by Cell Lines and Recombinant Factors
| Stimulatory Cells | Stimulatory Soluble Factors (Concentration) | Duration in Days | Purification | CLL Viability | CLL Proliferation | Ref. |
|---|---|---|---|---|---|---|
| irradiated mouse CDw32 L cells | anti-CD40 mAb (0.5 µg/mL), IL4 (500 U/mL) | 10 | CLL | [3H]TdR uptake 10× higher (vs. control without IL4); increased cell count (until day 8) | [79] | |
| CDw32 L cells | anti-CD40 moAb (0.5 µg/mL), IL2, IL10 | 3 (up to 9) | CLL | ~80% (~50% *) at day 9 | weak [3H]TdR uptake compared to healthy B cells | [35] |
| non-irradiated 3T40L cell line | a confluent layer of 3T40L | 3 | ? | 89% (71% *) | - | [21] |
| irradiated 3T40L cell line | 1:17 (3T40L:B cells) | 3 | CLL | 77% (32% *) | - | [80] |
| 3T40L cell line | - | 1–3 | PBMCs/ CLL | upregulation of genes promoting CLL-cell survival and cell-cycle arrest | [48] | |
| irradiated 3T40L cell line | IL4 (20 ng/mL) | 6 | CLL | 35% (55% *) | cell cycle (6% cells in S-phase vs. 3% in control); [3H]TdR uptake | [42] |
| allogenic human bone marrow stromal cells | CD40L (1 μg/mL), IL2 (100 U/mL), IL10 (10 ng/mL) | 3.5 | CLL | 35–75% (10–30% *) | BrdU cell cycle (proliferation induced in 8 of 21 samples); ~4% of cells in S+G2/M phase (DAPI staining) | [61] |
| irradiated 3T40L cell line | CpG (1.5 μg/mL) | 3 | PBMCs |
75% (IGHV unmut.), 70% (IGHV mutated) | CSFE (proliferation in 80% IGHV unmutated CLL, 25% IGHV mutated CLL at day 5) | [60] |
| mouse fibroblasts expressing CD31 | IL4 (5 ng/mL) | 7 | PBMCs | 60% (30% *) | CSFE; 4–10% Ki67+ cells | [59] |
| mouse fibroblasts expressing CD40L | IL4 (5 ng/mL) | 7 | PBMCs | 50% (30% *) | CSFE; 8–14% Ki67+ cells | [59] |
| 3T40L cell line | IL21 (25 ng/mL) | 5 | CLL | ~80% (~30% *) at day 8 | CSFE (~50% divided cells, ~20% w/o IL21); Ki67 | [38] |
| non-irradiated human-bone-marrow stromal (BMSC) cell line UE6E7T-2 | CD40L (1 μg/mL), CpG (1.5 μg/mL), 1:100 (BMSC:B cells) | 3 | PBMCs | 25% (48% *); 32% (60% *) | ↑ S phase; 11% Ki67+ cells (1.18%); ↑ S phase, 7.68% Ki67+ cells (0.81%) | [34,81] |
| irradiated mouse L cells expressing CD40L | IL21 (12.5 ng/mL) | 9–10 | CLL | 94% at day 5 | CSFE (proliferation of 49% cells; 18% in control with IL4 instead of IL21) | [67] |
| non-irradiated human BMSC cell line UE6E7T-2 | CD40L (1 μg/mL), CpG (1.5 μg/mL), anti-IgM (10 μg/mL) | 2 | PMBCs | 137% (100% *) | 7% Ki67+ cells | [82] |
| 3T40L cell line | IL21 (25 ng/mL) | 5 | PBMCs | as control (~90% *) | CSFE (increased division ~10× compared to control) | [78] |
| HS5 cell line | IL2 (50 ng/mL), CpG (1 µg/mL) | 4 | PBMCs | 70% (82% *) | CSFE (proliferation of 30% cells) | [62] |
| BMF cell line # | CpG (2 µg/mL), IL15 (10 ng/mL) | 7 | CLL | - | CSFE (several generations); Ki67 | [63] |
| Recombinant Factor (Concentration) | Duration in Days | Purification | CLL Viability | CLL Proliferation | Ref. |
|---|---|---|---|---|---|
| trimeric anti-CD40L moAb, IL4 (20 ng/mL) | 4 | PBMCs | - | [3H]TdR uptake (56% of cells with a high rate of DNA synthesis) | [50] |
| trimeric anti-CD40L moAb (0.5 μg/mL) | 3 | PBMCs | 55% (22% *) | cell cycle (5.3% of proliferating cells); 31% samples non-responding | [52] |
| IL4 (2 ng/mL), CD40L plus enhancer (100 ng/mL) | 7 | CLL | ~80% (40–80% *) | - | [12] |
| CD40L or anti-CD40 moAb, IL4 (10 ng/mL), IL21 (20 ng/mL) | 1.6 | PBMCs | - | [3H]TdR uptake; BrdU cell cycle, 25% cells in S+G2/M phase (0.7–7.8% *) | [11] |
| CpG (5 μg/mL), IL21 (50 ng/mL) | 2.3 | CLL | no difference from control | [3H]TdR uptake, significantly increased proliferation with addition of IL21 | [83] |
| histidine-tagged CD40L (50 ng/mL), anti-polyhistidin mAb (5 μg/mL), CpG (10 μg/mL), IL2 (50 ng/mL), IL10 (50 ng/mL), IL15 (10 ng/mL), and IL6 (50 ng/mL), in specific time-dependent sequence | 7 | CLL | differentiation into antibody-producing cells | [84] | |
| IL15 (15 ng/mL), CpG (1.5 μg/mL) | 6 | CLL | 60–80% | CSFE (~70% divided cells) | [64] |
| IL15 (15 ng/mL), CpG (1.5 μg/mL) | 5 | PBMCs | <5% difference from control | CSFE (significantly increased cell division) | [78] |
| anti-IgM (10 µg/mL), trimeric CD40L (100 ng/mL), IL4 (10 ng/mL), and IL21 (25 ng/mL) | 6 | CLL | - | CSFE (41% of CLL samples proliferating) | [44] |
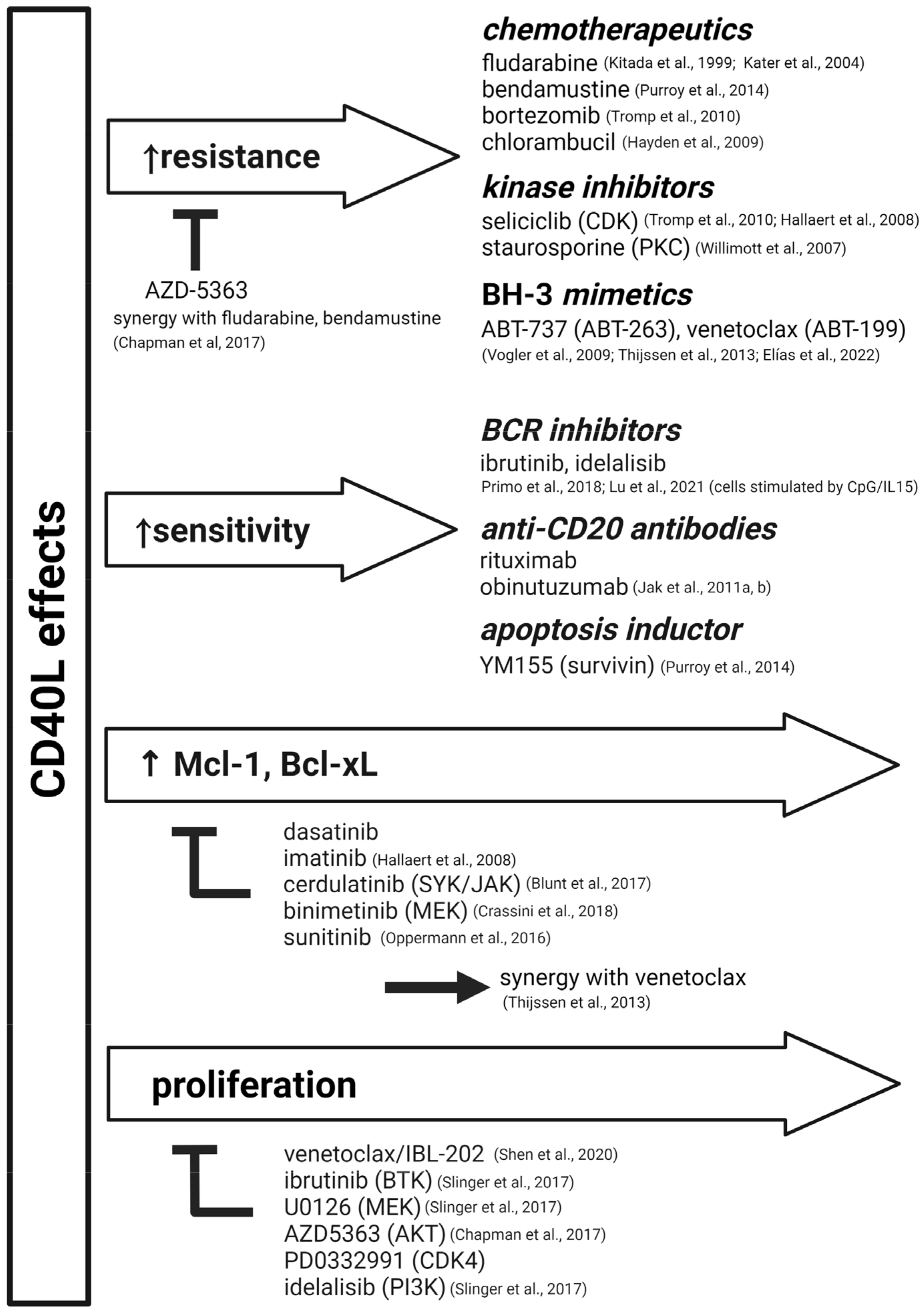
4. In Vitro: Use of CD40L-Based Systems for Drug Testing
5. In Vitro: CLL Co-Cultures in 3D
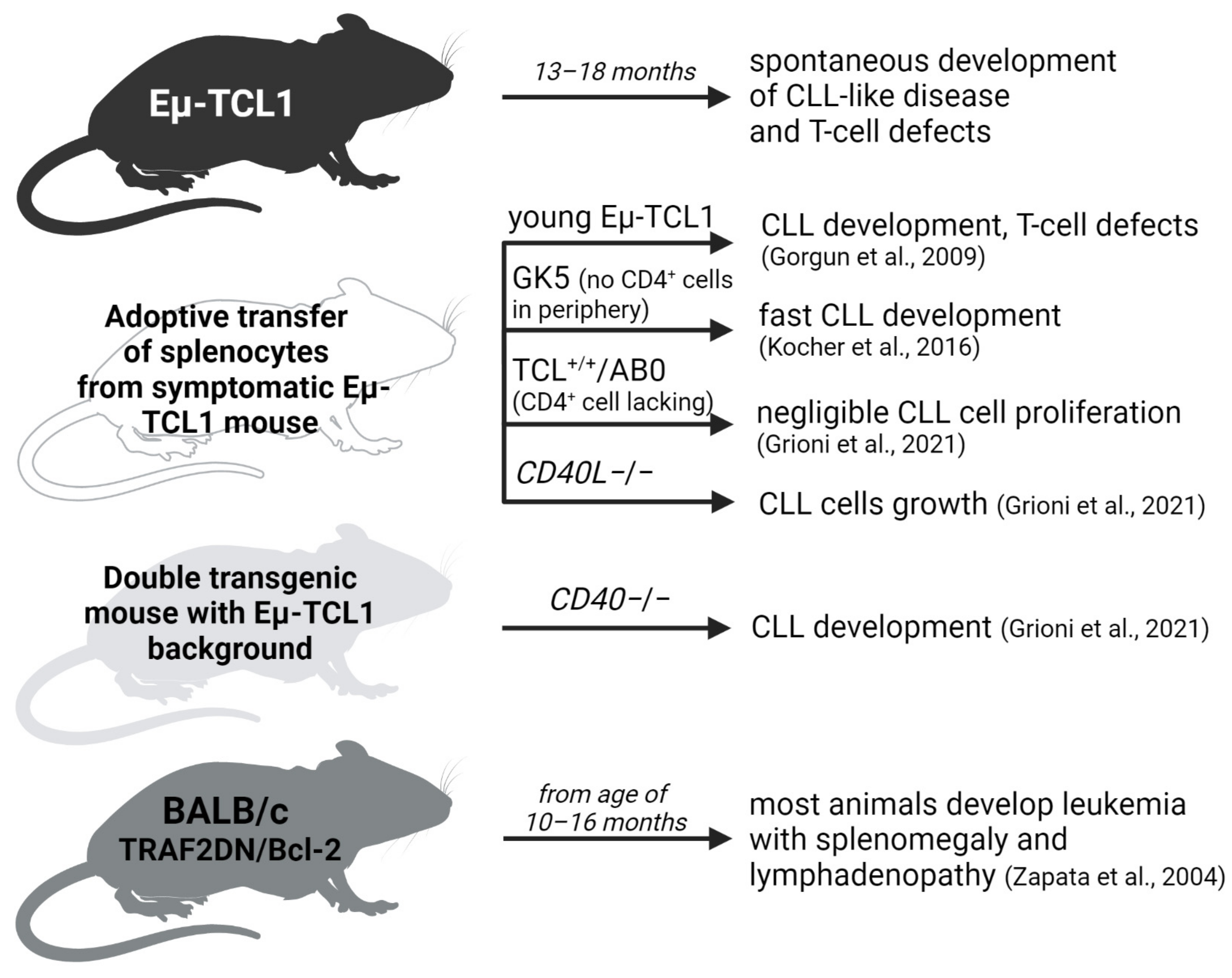
6. In Vivo: Role of T Cells in Transgenic CLL Models
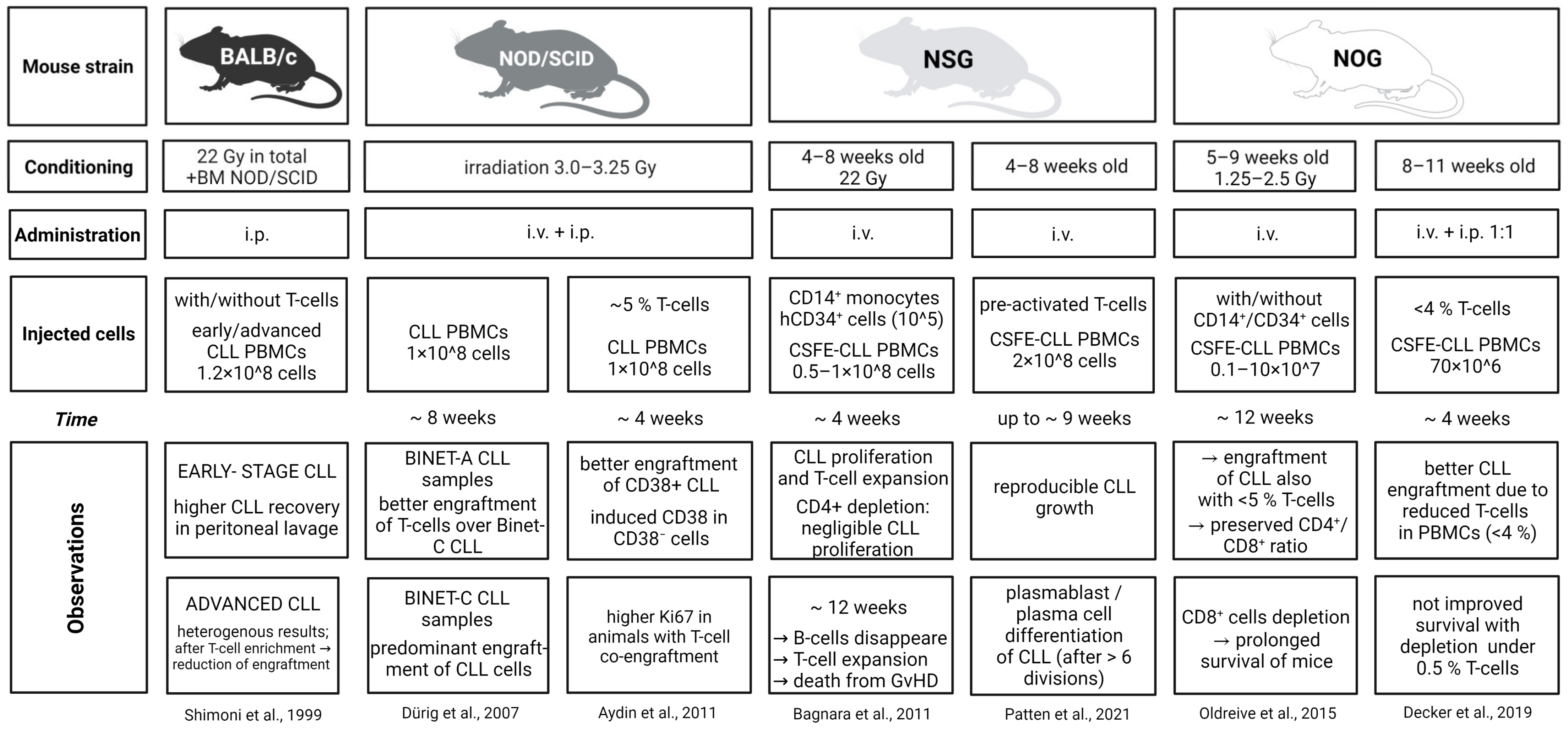
7. In Vivo: Co-Transplantation of CLL and T Cells in Xenograft Models
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smit, L.A.; Hallaert, D.Y.H.; Spijker, R.; de Goeij, B.; Jaspers, A.; Kater, A.P.; van Oers, M.H.J.; van Noesel, C.J.M.; Eldering, E. Differential Noxa/Mcl-1 Balance in Peripheral versus Lymph Node Chronic Lymphocytic Leukemia Cells Correlates with Survival Capacity. Blood 2007, 109, 1660–1668. [Google Scholar] [CrossRef] [PubMed]
- Herndon, T.M.; Chen, S.-S.; Saba, N.S.; Valdez, J.; Emson, C.; Gatmaitan, M.; Tian, X.; Hughes, T.E.; Sun, C.; Arthur, D.C.; et al. Direct In Vivo Evidence for Increased Proliferation of CLL Cells in Lymph Nodes Compared to Bone Marrow and Peripheral Blood. Leukemia 2017, 31, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Herishanu, Y.; Pérez-Galán, P.; Liu, D.; Biancotto, A.; Pittaluga, S.; Vire, B.; Gibellini, F.; Njuguna, N.; Lee, E.; Stennett, L.; et al. The Lymph Node Microenvironment Promotes B-Cell Receptor Signaling, NF-ΚB Activation, and Tumor Proliferation in Chronic Lymphocytic Leukemia. Blood 2011, 117, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Pavlasova, G.; Borsky, M.; Svobodova, V.; Oppelt, J.; Cerna, K.; Novotna, J.; Seda, V.; Fojtova, M.; Fajkus, J.; Brychtova, Y.; et al. Rituximab Primarily Targets an Intra-Clonal BCR Signaling Proficient CLL Subpopulation Characterized by High CD20 Levels. Leukemia 2018, 32, 2028–2031. [Google Scholar] [CrossRef] [PubMed]
- Seda, V.; Mraz, M. B-Cell Receptor Signalling and Its Crosstalk with Other Pathways in Normal and Malignant Cells. Eur. J. Haematol. 2015, 94, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Haselager, M.V.; Kater, A.P.; Eldering, E. Proliferative Signals in Chronic Lymphocytic Leukemia; What Are We Missing? Front. Oncol. 2020, 10, 2205. [Google Scholar] [CrossRef] [PubMed]
- Vlachonikola, E.; Stamatopoulos, K.; Chatzidimitriou, A. T Cells in Chronic Lymphocytic Leukemia: A Two-Edged Sword. Front. Immunol. 2021, 11, 612244. [Google Scholar] [CrossRef]
- Roessner, P.M.; Seiffert, M. T-Cells in Chronic Lymphocytic Leukemia: Guardians or Drivers of Disease? Leukemia 2020, 34, 2012–2024. [Google Scholar] [CrossRef]
- Wu, X.; Fajardo-Despaigne, J.E.; Zhang, C.; Neppalli, V.; Banerji, V.; Johnston, J.B.; Gibson, S.B.; Marshall, A.J. Altered T Follicular Helper Cell Subsets and Function in Chronic Lymphocytic Leukemia. Front. Oncol. 2021, 11, 674492. [Google Scholar] [CrossRef]
- Ponzoni, M.; Doglioni, C.; Caligaris-Cappio, F. Chronic Lymphocytic Leukemia: The Pathologist’s View of Lymph Node Microenvironment. Semin. Diagn. Pathol. 2011, 28, 161–166. [Google Scholar] [CrossRef]
- Ahearne, M.J.; Willimott, S.; Piñon, L.; Kennedy, D.B.; Miall, F.; Dyer, M.J.S.; Wagner, S.D. Enhancement of CD154/IL4 Proliferation by the T Follicular Helper (Tfh) Cytokine, IL21 and Increased Numbers of Circulating Cells Resembling Tfh Cells in Chronic Lymphocytic Leukaemia. Br. J. Haematol. 2013, 162, 360–370. [Google Scholar] [CrossRef]
- Coscia, M.; Pantaleoni, F.; Riganti, C.; Vitale, C.; Rigoni, M.; Peola, S.; Castella, B.; Foglietta, M.; Griggio, V.; Drandi, D.; et al. IGHV Unmutated CLL B Cells Are More Prone to Spontaneous Apoptosis and Subject to Environmental Prosurvival Signals than Mutated CLL B Cells. Leukemia 2011, 25, 828–837. [Google Scholar] [CrossRef]
- Schattner, E.J.; Mascarenhas, J.; Reyfman, I.; Koshy, M.; Woo, C.; Friedman, S.M.; Crow, M.K. Chronic Lymphocytic Leukemia B Cells Can Express CD40 Ligand and Demonstrate T-Cell Type Costimulatory Capacity. Blood 1998, 91, 2689–2697. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, A.P.; Contesti, J.; Huergo-Zapico, L.; Lopez-Soto, A.; Fernández-Guizán, A.; Acebes-Huerta, A.; Gonzalez-Huerta, A.J.; Gonzalez, E.; Fernandez-Alvarez, C.; Gonzalez, S. Prognostic Significance of CD8 and CD4 T Cells in Chronic Lymphocytic Leukemia. Leuk. Lymphoma 2010, 51, 1829–1836. [Google Scholar] [CrossRef]
- Long, M.; Beckwith, K.; Do, P.; Mundy, B.L.; Gordon, A.; Lehman, A.M.; Maddocks, K.J.; Cheney, C.; Jones, J.A.; Flynn, J.M.; et al. Ibrutinib Treatment Improves T Cell Number and Function in CLL Patients. J. Clin. Investig. 2017, 127, 3052–3064. [Google Scholar] [CrossRef]
- Davis, J.E.; Sharpe, C.; Mason, K.; Tam, C.S.; Koldej, R.M.; Ritchie, D.S. Ibrutinib Protects T Cells in Patients with CLL from Proliferation-Induced Senescence. J. Transl. Med. 2021, 19, 473. [Google Scholar] [CrossRef]
- Görgün, G.; Holderried, T.A.W.; Zahrieh, D.; Neuberg, D.; Gribben, J.G. Chronic Lymphocytic Leukemia Cells Induce Changes in Gene Expression of CD4 and CD8 T Cells. J. Clin. Investig. 2005, 115, 1797–1805. [Google Scholar] [CrossRef]
- Tötterman, T.H.; Carlsson, M.; Simonsson, B.; Bengtsson, M.; Nilsson, K. T-Cell Activation and Subset Patterns Are Altered in B-CLL and Correlate with the Stage of the Disease. Blood 1989, 74, 786–792. [Google Scholar] [CrossRef]
- Vardi, A.; Vlachonikola, E.; Karypidou, M.; Stalika, E.; Bikos, V.; Gemenetzi, K.; Maramis, C.; Siorenta, A.; Anagnostopoulos, A.; Pospisilova, S.; et al. Restrictions in the T-Cell Repertoire of Chronic Lymphocytic Leukemia: High-Throughput Immunoprofiling Supports Selection by Shared Antigenic Elements. Leukemia 2017, 31, 1555–1561. [Google Scholar] [CrossRef]
- Riches, J.C.; Davies, J.K.; McClanahan, F.; Fatah, R.; Iqbal, S.; Agrawal, S.; Ramsay, A.G.; Gribben, J.G. T Cells from CLL Patients Exhibit Features of T-Cell Exhaustion but Retain Capacity for Cytokine Production. Blood 2013, 121, 1612–1621. [Google Scholar] [CrossRef]
- Kitada, S.; Zapata, J.M.; Andreeff, M.; Reed, J.C. Bryostatin and CD40-Ligand Enhance Apoptosis Resistance and Induce Expression of Cell Survival Genes in B-Cell Chronic Lymphocytic Leukaemia. Br. J. Haematol. 1999, 106, 995–1004. [Google Scholar] [CrossRef]
- Os, A.; Bürgler, S.; Ribes, A.P.; Funderud, A.; Wang, D.; Thompson, K.M.; Tjønnfjord, G.E.; Bogen, B.; Munthe, L.A. Chronic Lymphocytic Leukemia Cells Are Activated and Proliferate in Response to Specific T Helper Cells. Cell Rep. 2013, 4, 566–577. [Google Scholar] [CrossRef]
- Kater, A.P.; Dicker, F.; Mangiola, M.; Welsh, K.; Houghten, R.; Ostresh, J.; Nefzi, A.; Reed, J.C.; Pinilla, C.; Kipps, T.J. Inhibitors of XIAP Sensitize CD40-Activated Chronic Lymphocytic Leukemia Cells to CD95-Mediated Apoptosis. Blood 2005, 106, 1742–1748. [Google Scholar] [CrossRef]
- Dicker, F.; Kater, A.P.; Prada, C.E.; Fukuda, T.; Castro, J.E.; Sun, G.; Wang, J.Y.; Kipps, T.J. CD154 Induces P73 to Overcome the Resistance to Apoptosis of Chronic Lymphocytic Leukemia Cells Lacking Functional P53. Blood 2006, 108, 3450–3457. [Google Scholar] [CrossRef][Green Version]
- Van Attekum, M.H.; Eldering, E.; Kater, A.P. Chronic Lymphocytic Leukemia Cells Are Active Participants in Microenvironmental Cross-Talk. Haematologica 2017, 102, 1469–1476. [Google Scholar] [CrossRef]
- Burger, J.A.; Quiroga, M.P.; Hartmann, E.; Bürkle, A.; Wierda, W.G.; Keating, M.J.; Rosenwald, A. High-Level Expression of the T-Cell Chemokines CCL3 and CCL4 by Chronic Lymphocytic Leukemia B Cells in Nurselike Cell Cocultures and after BCR Stimulation. Blood 2009, 113, 3050–3058. [Google Scholar] [CrossRef]
- Hartmann, E.M.; Rudelius, M.; Burger, J.A.; Rosenwald, A. CCL3 Chemokine Expression by Chronic Lymphocytic Leukemia Cells Orchestrates the Composition of the Microenvironment in Lymph Node Infiltrates. Leuk. Lymphoma 2016, 57, 563–571. [Google Scholar] [CrossRef]
- Man, S.; Henley, P. Chronic Lymphocytic Leukaemia: The Role of T Cells in a B Cell Disease. Br. J. Haematol. 2019, 186, 220–233. [Google Scholar] [CrossRef]
- Vaca, A.M.; Ioannou, N.; Sivina, M.; Vlachonikola, E.; Clise-Dwyer, K.; Kim, E.; Li, D.; Ma, Q.; Ferrajoli, A.; Estrov, Z.; et al. Activation and Expansion of T-Follicular Helper Cells in Chronic Lymphocytic Leukemia Nurselike Cell Co-Cultures. Leukemia 2022, 36, 1324–1335. [Google Scholar] [CrossRef]
- Burgess, M.; Cheung, C.; Chambers, L.; Ravindranath, K.; Minhas, G.; Knop, L.; Mollee, P.; McMillan, N.A.J.; Gill, D. CCL2 and CXCL2 Enhance Survival of Primary Chronic Lymphocytic Leukemia Cells in Vitro. Leuk. Lymphoma 2012, 53, 1988–1998. [Google Scholar] [CrossRef]
- Calissano, C.; Damle, R.N.; Marsilio, S.; Yan, X.-J.; Yancopoulos, S.; Hayes, G.; Emson, C.; Murphy, E.J.; Hellerstein, M.K.; Sison, C.; et al. Intraclonal Complexity in Chronic Lymphocytic Leukemia: Fractions Enriched in Recently Born/Divided and Older/Quiescent Cells. Mol. Med. 2011, 17, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Seda, V.; Vojackova, E.; Ondrisova, L.; Kostalova, L.; Sharma, S.; Loja, T.; Mladonicka Pavlasova, G.; Zicha, D.; Kudlickova Peskova, M.; Krivanek, J.; et al. FoxO1-GAB1 Axis Regulates Homing Capacity and Tonic AKT Activity in Chronic Lymphocytic Leukemia. Blood 2021, 138, 758–772. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Pavlasova, G.M.; Seda, V.; Cerna, K.A.; Vojackova, E.; Filip, D.; Ondrisova, L.; Sandova, V.; Kostalova, L.; Zeni, P.F.; et al. MiR-29 Modulates CD40 Signaling in Chronic Lymphocytic Leukemia by Targeting TRAF4: An Axis Affected by BCR Inhibitors. Blood 2021, 137, 2481–2494. [Google Scholar] [CrossRef] [PubMed]
- Purroy, N.; Abrisqueta, P.; Carabia, J.; Carpio, C.; Palacio, C.; Bosch, F.; Crespo, M. Co-Culture of Primary CLL Cells with Bone Marrow Mesenchymal Cells, CD40 Ligand and CpG ODN Promotes Proliferation of Chemoresistant CLL Cells Phenotypically Comparable to Those Proliferating In Vivo. Oncotarget 2014, 6, 7632–7643. [Google Scholar] [CrossRef]
- Tretter, T.; Schuler, M.; Schneller, F.; Brass, U.; Esswein, M.; Aman, M.J.; Huber, C.; Peschel, C. Direct Cellular Interaction with Activated CD4(+) T Cells Overcomes Hyporesponsiveness of B-Cell Chronic Lymphocytic Leukemia In Vitro. Cell Immunol. 1998, 189, 41–50. [Google Scholar] [CrossRef]
- Patten, P.E.M.; Buggins, A.G.S.; Richards, J.; Wotherspoon, A.; Salisbury, J.; Mufti, G.J.; Hamblin, T.J.; Devereux, S. CD38 Expression in Chronic Lymphocytic Leukemia Is Regulated by the Tumor Microenvironment. Blood 2008, 111, 5173–5181. [Google Scholar] [CrossRef]
- Romano, C.; De Fanis, U.; Sellitto, A.; Dalla Mora, L.; Chiurazzi, F.; Giunta, R.; Rotoli, B.; Lucivero, G. Effects of Preactivated Autologous T Lymphocytes on CD80, CD86 and CD95 Expression by Chronic Lymphocytic Leukemia B Cells. Leuk. Lymphoma 2003, 44, 1963–1971. [Google Scholar] [CrossRef]
- Pascutti, M.F.; Jak, M.; Tromp, J.M.; Derks, I.A.M.; Remmerswaal, E.B.M.; Thijssen, R.; van Attekum, M.H.A.; van Bochove, G.G.; Luijks, D.M.; Pals, S.T.; et al. IL-21 and CD40L Signals from Autologous T Cells Can Induce Antigen-Independent Proliferation of CLL Cells. Blood 2013, 122, 3010–3019. [Google Scholar] [CrossRef]
- Ghia, P.; Circosta, P.; Scielzo, C.; Vallario, A.; Camporeale, A.; Granziero, L.; Caligaris-Cappio, F. Differential Effects on CLL Cell Survival Exerted by Different Microenvironmental Elements. In Chronic Lymphocytic Leukemia; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 135–145. ISBN 978-3-540-29933-2. [Google Scholar]
- Jones, C.H.; Lin, T.T.; Walsby, E.J.; Pratt, G.E.; Fegan, C.; Baird, D.M.; Pepper, C. In Vitro Co-Culture of CLL-B Cells Reveals Long-Term Survival, Proliferation, and Maintenance of Telomere Length. Blood 2016, 128, 350. [Google Scholar] [CrossRef]
- Wiesner, M.; Zentz, C.; Mayr, C.; Wimmer, R.; Hammerschmidt, W.; Zeidler, R.; Moosmann, A. Conditional Immortalization of Human B Cells by CD40 Ligation. PLoS ONE 2008, 3, e1464. [Google Scholar] [CrossRef]
- Willimott, S.; Baou, M.; Naresh, K.; Wagner, S.D. CD154 Induces a Switch in Pro-Survival Bcl-2 Family Members in Chronic Lymphocytic Leukaemia. Br. J. Haematol. 2007, 138, 721–732. [Google Scholar] [CrossRef]
- Brian, A.A. Stimulation of B-Cell Proliferation by Membrane-Associated Molecules from Activated T Cells. Proc. Natl. Acad. Sci. USA 1988, 85, 564–568. [Google Scholar] [CrossRef]
- Schleiss, C.; Ilias, W.; Tahar, O.; Güler, Y.; Miguet, L.; Mayeur-Rousse, C.; Mauvieux, L.; Fornecker, L.-M.; Toussaint, E.; Herbrecht, R.; et al. BCR-Associated Factors Driving Chronic Lymphocytic Leukemia Cells Proliferation Ex Vivo. Sci. Rep. 2019, 9, 701. [Google Scholar] [CrossRef]
- Liebig, T.M.; Fiedler, A.; Zoghi, S.; Shimabukuro-Vornhagen, A.; von Bergwelt-Baildon, M.S. Generation of Human CD40-Activated B Cells. J. Vis. Exp. 2009, 32, 1373. [Google Scholar] [CrossRef]
- Ghamlouch, H.; Darwiche, W.; Hodroge, A.; Ouled-Haddou, H.; Dupont, S.; Singh, A.R.; Guignant, C.; Trudel, S.; Royer, B.; Gubler, B.; et al. Factors Involved in CLL Pathogenesis and Cell Survival Are Disrupted by Differentiation of CLL B-Cells into Antibody-Secreting Cells. Oncotarget 2015, 6, 18484–18503. [Google Scholar] [CrossRef]
- Grdisa, M. Influence of CD40 Ligation on Survival and Apoptosis of B-CLL Cells In Vitro. Leuk. Res. 2003, 27, 951–956. [Google Scholar] [CrossRef]
- Gricks, C.S.; Zahrieh, D.; Zauls, A.J.; Gorgun, G.; Drandi, D.; Mauerer, K.; Neuberg, D.; Gribben, J.G. Differential Regulation of Gene Expression Following CD40 Activation of Leukemic Compared to Healthy B Cells. Blood 2004, 104, 4002–4009. [Google Scholar] [CrossRef][Green Version]
- Scielzo, C.; Apollonio, B.; Scarfò, L.; Janus, A.; Muzio, M.; ten Hacken, E.; Ghia, P.; Caligaris-Cappio, F. The Functional in Vitro Response to CD40 Ligation Reflects a Different Clinical Outcome in Patients with Chronic Lymphocytic Leukemia. Leukemia 2011, 25, 1760–1767. [Google Scholar] [CrossRef]
- Jacob, A.; Pound, J.D.; Challa, A.; Gordon, J. Release of Clonal Block in B Cell Chronic Lymphocytic Leukaemia by Engagement of Co-Operative Epitopes on CD40. Leuk. Res. 1998, 22, 379–382. [Google Scholar] [CrossRef]
- Néron, S.; Nadeau, P.J.; Darveau, A.; Leblanc, J.-F. Tuning of CD40-CD154 Interactions in Human B-Lymphocyte Activation: A Broad Array of in Vitro Models for a Complex In Vivo Situation. Arch. Immunol. Ther. Exp. 2011, 59, 25–40. [Google Scholar] [CrossRef]
- Granziero, L.; Ghia, P.; Circosta, P.; Gottardi, D.; Strola, G.; Geuna, M.; Montagna, L.; Piccoli, P.; Chilosi, M.; Caligaris-Cappio, F. Survivin Is Expressed on CD40 Stimulation and Interfaces Proliferation and Apoptosis in B-Cell Chronic Lymphocytic Leukemia. Blood 2001, 97, 2777–2783. [Google Scholar] [CrossRef] [PubMed]
- Néron, S.; Racine, C.; Roy, A.; Guérin, M. Differential Responses of Human B-Lymphocyte Subpopulations to Graded Levels of CD40-CD154 Interaction. Immunology 2005, 116, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Dancescu, M.; Rubio-Trujillo, M.; Biron, G.; Bron, D.; Delespesse, G.; Sarfati, M. Interleukin 4 Protects Chronic Lymphocytic Leukemic B Cells from Death by Apoptosis and Upregulates Bcl-2 Expression. J. Exp. Med. 1992, 176, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Hernandez, M.M.; Blunt, M.D.; Dobson, R.; Yeomans, A.; Thirdborough, S.; Larrayoz, M.; Smith, L.D.; Linley, A.; Strefford, J.C.; Davies, A.; et al. IL-4 Enhances Expression and Function of Surface IgM in CLL Cells. Blood 2016, 127, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Zhang, L.; Chiorazzi, N.; Rothstein, T.L. IL-4 Rescues Surface IgM Expression in Chronic Lymphocytic Leukemia. Blood 2016, 128, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Sandova, V.; Pavlasova, G.M.; Seda, V.; Cerna, K.A.; Sharma, S.; Palusova, V.; Brychtova, Y.; Pospisilova, S.; Fernandes, S.M.; Panovska, A.; et al. IL4-STAT6 Signaling Induces CD20 in Chronic Lymphocytic Leukemia and This Axis Is Repressed by PI3Kδ Inhibitor Idelalisib. Haematologica 2021, 106, 2995–2999. [Google Scholar] [CrossRef]
- Van Kooten, C.; Rensink, I.; Aarden, L.; van Oers, R. Differentiation of Purified Malignant B Cells Induced by PMA or by Activated Normal T Cells. Leukemia 1993, 7, 1576–1584. [Google Scholar]
- Hamilton, E.; Pearce, L.; Morgan, L.; Robinson, S.; Ware, V.; Brennan, P.; Thomas, N.S.B.; Yallop, D.; Devereux, S.; Fegan, C.; et al. Mimicking the Tumour Microenvironment: Three Different Co-Culture Systems Induce a Similar Phenotype but Distinct Proliferative Signals in Primary Chronic Lymphocytic Leukaemia Cells. Br. J. Haematol. 2012, 158, 589–599. [Google Scholar] [CrossRef]
- Tromp, J.; Tonino, S.; Elias, J.; Jaspers, A.; Luijks, D.M.; Kater, A.; Lier, R.V.; Oers, M.; Eldering, E. Dichotomy in NF-ΚB Signaling and Chemoresistance in Immunoglobulin Variable Heavy-Chain-Mutated versus Unmutated CLL Cells upon CD40/TLR9 Triggering. Oncogene 2010, 29, 5071–5082. [Google Scholar] [CrossRef]
- Plander, M.; Seegers, S.; Ugocsai, P.; Diermeier-Daucher, S.; Iványi, J.; Schmitz, G.; Hofstädter, F.; Schwarz, S.; Orsó, E.; Knüchel, R.; et al. Different Proliferative and Survival Capacity of CLL-Cells in a Newly Established in Vitro Model for Pseudofollicles. Leukemia 2009, 23, 2118–2128. [Google Scholar] [CrossRef][Green Version]
- Primo, D.; Scarfò, L.; Xochelli, A.; Mattsson, M.; Ranghetti, P.; Espinosa, A.B.; Robles, A.; Gorrochategui, J.; Martínez-López, J.; de la Serna, J.; et al. A Novel Ex Vivo High-Throughput Assay Reveals Antiproliferative Effects of Idelalisib and Ibrutinib in Chronic Lymphocytic Leukemia. Oncotarget 2018, 9, 26019–26031. [Google Scholar] [CrossRef]
- Lu, P.; Wang, S.; Franzen, C.A.; Venkataraman, G.; McClure, R.; Li, L.; Wu, W.; Niu, N.; Sukhanova, M.; Pei, J.; et al. Ibrutinib and Venetoclax Target Distinct Subpopulations of CLL Cells: Implication for Residual Disease Eradication. Blood Cancer J. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Mongini, P.K.A.; Gupta, R.; Boyle, E.; Nieto, J.; Lee, H.; Stein, J.; Bandovic, J.; Stankovic, T.; Barrientos, J.; Kolitz, J.E.; et al. TLR-9 and IL-15 Synergy Promotes the In Vitro Clonal Expansion of Chronic Lymphocytic Leukemia B Cells. J. Immunol. 2015, 195, 901–923. [Google Scholar] [CrossRef]
- De Totero, D.; Capaia, M.; Fabbi, M.; Croce, M.; Meazza, R.; Cutrona, G.; Zupo, S.; Loiacono, F.; Truini, M.; Ferrarini, M.; et al. Heterogeneous Expression and Function of IL-21R and Susceptibility to IL-21−mediated Apoptosis in Follicular Lymphoma Cells. Exp. Hematol. 2010, 38, 373–383. [Google Scholar] [CrossRef]
- Cecco, L.D.; Capaia, M.; Zupo, S.; Cutrona, G.; Matis, S.; Brizzolara, A.; Orengo, A.M.; Croce, M.; Marchesi, E.; Ferrarini, M.; et al. Interleukin 21 Controls MRNA and MicroRNA Expression in CD40-Activated Chronic Lymphocytic Leukemia Cells. PLoS ONE 2015, 10, e0134706. [Google Scholar] [CrossRef]
- Chapman, E.A.; Oates, M.; Mohammad, I.S.; Davies, B.R.; Stockman, P.K.; Zhuang, J.; Pettitt, A.R. Delineating the Distinct Role of AKT in Mediating Cell Survival and Proliferation Induced by CD154 and IL-4/IL-21 in Chronic Lymphocytic Leukemia. Oncotarget 2017, 8, 102948–102964. [Google Scholar] [CrossRef][Green Version]
- De Totero, D.; Meazza, R.; Zupo, S.; Cutrona, G.; Matis, S.; Colombo, M.; Balleari, E.; Pierri, I.; Fabbi, M.; Capaia, M.; et al. Interleukin-21 Receptor (IL-21R) Is Up-Regulated by CD40 Triggering and Mediates Proapoptotic Signals in Chronic Lymphocytic Leukemia B Cells. Blood 2006, 107, 3708–3715. [Google Scholar] [CrossRef]
- Nojima, T.; Haniuda, K.; Moutai, T.; Matsudaira, M.; Mizokawa, S.; Shiratori, I.; Azuma, T.; Kitamura, D. In-Vitro Derived Germinal Centre B Cells Differentially Generate Memory B or Plasma Cells In Vivo. Nat. Commun. 2011, 2, 465. [Google Scholar] [CrossRef]
- Caeser, R.; Di Re, M.; Krupka, J.A.; Gao, J.; Lara-Chica, M.; Dias, J.M.L.; Cooke, S.L.; Fenner, R.; Usheva, Z.; Runge, H.F.P.; et al. Genetic Modification of Primary Human B Cells to Model High-Grade Lymphoma. Nat. Commun. 2019, 10, 4543. [Google Scholar] [CrossRef]
- Buschle, M.; Campana, D.; Carding, S.R.; Richard, C.; Hoffbrand, A.V.; Brenner, M.K. Interferon Gamma Inhibits Apoptotic Cell Death in B Cell Chronic Lymphocytic Leukemia. J. Exp. Med. 1993, 177, 213–218. [Google Scholar] [CrossRef]
- Xia, M.; Luo, T.Y.; Shi, Y.; Wang, G.; Tsui, H.; Harari, D.; Spaner, D.E. Effect of Ibrutinib on the IFN Response of Chronic Lymphocytic Leukemia Cells. J. Immunol. 2020, 205, 2629–2639. [Google Scholar] [CrossRef]
- Bauvois, B.; Pramil, E.; Jondreville, L.; Quiney, C.; Nguyen-Khac, F.; Susin, S.A. Activation of Interferon Signaling in Chronic Lymphocytic Leukemia Cells Contributes to Apoptosis Resistance via a JAK-Src/STAT3/Mcl-1 Signaling Pathway. Biomedicines 2021, 9, 188. [Google Scholar] [CrossRef]
- Chaouchi, N.; Wallon, C.; Goujard, C.; Tertian, G.; Rudent, A.; Caput, D.; Ferrera, P.; Minty, A.; Vazquez, A.; Delfraissy, J.F. Interleukin-13 Inhibits Interleukin-2-Induced Proliferation and Protects Chronic Lymphocytic Leukemia B Cells from In Vitro Apoptosis. Blood 1996, 87, 1022–1029. [Google Scholar] [CrossRef]
- Zhu, F.; McCaw, L.; Spaner, D.E.; Gorczynski, R.M. Targeting the IL-17/IL-6 Axis Can Alter Growth of Chronic Lymphocytic Leukemia in Vivo/in Vitro. Leuk. Res. 2018, 66, 28–38. [Google Scholar] [CrossRef]
- Ghamlouch, H.; Ouled-Haddou, H.; Damaj, G.; Royer, B.; Gubler, B.; Marolleau, J.-P. A Combination of Cytokines Rescues Highly Purified Leukemic CLL B-Cells from Spontaneous Apoptosis In Vitro. PLoS ONE 2013, 8, e60370. [Google Scholar] [CrossRef]
- Alhakeem, S.S.; McKenna, M.K.; Oben, K.Z.; Noothi, S.K.; Rivas, J.R.; Hildebrandt, G.C.; Fleischman, R.A.; Rangnekar, V.M.; Muthusamy, N.; Bondada, S. Chronic Lymphocytic Leukemia–Derived IL-10 Suppresses Antitumor Immunity. J. Immunol. 2018, 200, 4180–4189. [Google Scholar] [CrossRef]
- Slinger, E.; Thijssen, R.; Kater, A.P.; Eldering, E. Targeting Antigen-Independent Proliferation in Chronic Lymphocytic Leukemia through Differential Kinase Inhibition. Leukemia 2017, 31, 2601–2607. [Google Scholar] [CrossRef]
- Fluckiger, A.C.; Rossi, J.F.; Bussel, A.; Bryon, P.; Banchereau, J.; Defrance, T. Responsiveness of Chronic Lymphocytic Leukemia B Cells Activated via Surface Igs or CD40 to B-Cell Tropic Factors. Blood 1992, 80, 3173–3181. [Google Scholar] [CrossRef] [PubMed]
- Kater, A.P.; Evers, L.M.; Remmerswaal, E.B.M.; Jaspers, A.; Oosterwijk, M.F.; van Lier, R.A.W.; van Oers, M.H.J.; Eldering, E. CD40 Stimulation of B-Cell Chronic Lymphocytic Leukaemia Cells Enhances the Anti-Apoptotic Profile, but Also Bid Expression and Cells Remain Susceptible to Autologous Cytotoxic T-Lymphocyte Attack. Br. J. Haematol. 2004, 127, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Purroy, N.; Abrisqueta, P.; Carabia, J.; Carpio, C.; Calpe, E.; Palacio, C.; Castellví, J.; Crespo, M.; Bosch, F. Targeting the Proliferative and Chemoresistant Compartment in Chronic Lymphocytic Leukemia by Inhibiting Survivin Protein. Leukemia 2014, 28, 1993–2004. [Google Scholar] [CrossRef] [PubMed]
- Purroy, N.; Carabia, J.; Abrisqueta, P.; Egia, L.; Aguiló, M.; Carpio, C.; Palacio, C.; Crespo, M.; Bosch, F. Inhibition of BCR Signaling Using the Syk Inhibitor TAK-659 Prevents Stroma-Mediated Signaling in Chronic Lymphocytic Leukemia Cells. Oncotarget 2017, 8, 742–756. [Google Scholar] [CrossRef]
- Ghalamfarsa, G.; Jadidi-Niaragh, F.; Hojjat-Farsangi, M.; Asgarian-Omran, H.; Yousefi, M.; Tahmasebi, F.; Khoshnoodi, J.; Razavi, S.M.; Saboor-Yaraghi, A.A.; Rabbani, H.; et al. Differential Regulation of B-Cell Proliferation by IL21 in Different Subsets of Chronic Lymphocytic Leukemia. Cytokine 2013, 62, 439–445. [Google Scholar] [CrossRef]
- Ghamlouch, H.; Ouled-Haddou, H.; Guyart, A.; Regnier, A.; Trudel, S.; Claisse, J.-F.; Fuentes, V.; Royer, B.; Marolleau, J.-P.; Gubler, B. TLR9 Ligand (CpG Oligodeoxynucleotide) Induces CLL B-Cells to Differentiate into CD20+ Antibody-Secreting Cells. Front. Immunol. 2014, 5, 292. [Google Scholar] [CrossRef]
- Hallaert, D.Y.H.; Jaspers, A.; van Noesel, C.J.; van Oers, M.H.J.; Kater, A.P.; Eldering, E. C-Abl Kinase Inhibitors Overcome CD40-Mediated Drug Resistance in CLL: Implications for Therapeutic Targeting of Chemoresistant Niches. Blood 2008, 112, 5141–5149. [Google Scholar] [CrossRef]
- Oppermann, S.; Ylanko, J.; Shi, Y.; Hariharan, S.; Oakes, C.C.; Brauer, P.M.; Zúñiga-Pflücker, J.C.; Leber, B.; Spaner, D.E.; Andrews, D.W. High-Content Screening Identifies Kinase Inhibitors That Overcome Venetoclax Resistance in Activated CLL Cells. Blood 2016, 128, 934–947. [Google Scholar] [CrossRef]
- Hayden, R.E.; Pratt, G.; Davies, N.J.; Khanim, F.L.; Birtwistle, J.; Delgado, J.; Pearce, C.; Sant, T.; Drayson, M.T.; Bunce, C.M. Treatment of Primary CLL Cells with Bezafibrate and Medroxyprogesterone Acetate Induces Apoptosis and Represses the Pro-Proliferative Signal of CD40-Ligand, in Part through Increased 15dDelta12,14,PGJ2. Leukemia 2009, 23, 292–304. [Google Scholar] [CrossRef]
- Hallaert, D.Y.H.; Spijker, R.; Jak, M.; Derks, I.a.M.; Alves, N.L.; Wensveen, F.M.; de Boer, J.P.; de Jong, D.; Green, S.R.; van Oers, M.H.J.; et al. Crosstalk among Bcl-2 Family Members in B-CLL: Seliciclib Acts via the Mcl-1/Noxa Axis and Gradual Exhaustion of Bcl-2 Protection. Cell Death Differ. 2007, 14, 1958–1967. [Google Scholar] [CrossRef]
- Vogler, M.; Butterworth, M.; Majid, A.; Walewska, R.J.; Sun, X.-M.; Dyer, M.J.S.; Cohen, G.M. Concurrent Up-Regulation of BCL-XL and BCL2A1 Induces Approximately 1000-Fold Resistance to ABT-737 in Chronic Lymphocytic Leukemia. Blood 2009, 113, 4403–4413. [Google Scholar] [CrossRef]
- Thijssen, R.; Geest, C.R.; de Rooij, M.F.; Liu, N.; Florea, B.I.; Weller, K.; Overkleeft, H.S.; Van Oers, M.H.J.; Spaargaren, M.; Kater, A.P.; et al. Possible Mechanisms Of Resistance To The Novel BH3-Mimetic ABT-199 In In Vitro Lymph Node Models of CLL—The Role of Abl and Btk. Blood 2013, 122, 4188. [Google Scholar] [CrossRef]
- Elias, E.E.; Sarapura Martinez, V.J.; Amondarain, M.; Colado, A.; Cordini, G.; Bezares, R.F.; Fernandez Grecco, H.; del Rosario Custidiano, M.; Sánchez Ávalos, J.C.; Garate, G.; et al. Venetoclax-Resistant CLL Cells Show a Highly Activated and Proliferative Phenotype. Cancer Immunol. Immunother. 2022, 71, 979–987. [Google Scholar] [CrossRef]
- Jak, M.; van Bochove, G.G.W.; van Lier, R.a.W.; Eldering, E.; van Oers, M.H.J. CD40 Stimulation Sensitizes CLL Cells to Rituximab-Induced Cell Death. Leukemia 2011, 25, 968–978. [Google Scholar] [CrossRef][Green Version]
- Jak, M.; van Bochove, G.G.W.; Reits, E.A.; Kallemeijn, W.W.; Tromp, J.M.; Umana, P.; Klein, C.; van Lier, R.A.W.; van Oers, M.H.J.; Eldering, E. CD40 Stimulation Sensitizes CLL Cells to Lysosomal Cell Death Induction by Type II Anti-CD20 MAb GA101. Blood 2011, 118, 5178–5188. [Google Scholar] [CrossRef][Green Version]
- Blunt, M.D.; Koehrer, S.; Dobson, R.C.; Larrayoz, M.; Wilmore, S.; Hayman, A.; Parnell, J.; Smith, L.D.; Davies, A.; Johnson, P.W.M.; et al. The Dual Syk/JAK Inhibitor Cerdulatinib Antagonizes B-Cell Receptor and Microenvironmental Signaling in Chronic Lymphocytic Leukemia. Clin. Cancer Res. 2017, 23, 2313–2324. [Google Scholar] [CrossRef]
- Crassini, K.; Shen, Y.; Stevenson, W.S.; Christopherson, R.; Ward, C.; Mulligan, S.P.; Best, O.G. MEK1/2 Inhibition by Binimetinib Is Effective as a Single Agent and Potentiates the Actions of Venetoclax and ABT-737 under Conditions That Mimic the Chronic Lymphocytic Leukaemia (CLL) Tumour Microenvironment. Br. J. Haematol. 2018, 182, 360–372. [Google Scholar] [CrossRef]
- Shen, Y.; Crassini, K.; Fatima, N.; O’Dwyer, M.; O’Neill, M.; Christopherson, R.I.; Mulligan, S.P.; Best, O.G. IBL-202 Is Synergistic with Venetoclax in CLL under in Vitro Conditions That Mimic the Tumor Microenvironment. Blood Adv. 2020, 4, 5093–5106. [Google Scholar] [CrossRef]
- Kurtova, A.V.; Balakrishnan, K.; Chen, R.; Ding, W.; Schnabl, S.; Quiroga, M.P.; Sivina, M.; Wierda, W.G.; Estrov, Z.; Keating, M.J.; et al. Diverse Marrow Stromal Cells Protect CLL Cells from Spontaneous and Drug-Induced Apoptosis: Development of a Reliable and Reproducible System to Assess Stromal Cell Adhesion-Mediated Drug Resistance. Blood 2009, 114, 4441–4450. [Google Scholar] [CrossRef]
- Trimarco, V.; Ave, E.; Facco, M.; Chiodin, G.; Frezzato, F.; Martini, V.; Gattazzo, C.; Lessi, F.; Giorgi, C.A.; Visentin, A.; et al. Cross-Talk between Chronic Lymphocytic Leukemia (CLL) Tumor B Cells and Mesenchymal Stromal Cells (MSCs): Implications for Neoplastic Cell Survival. Oncotarget 2015, 6, 42130–42149. [Google Scholar] [CrossRef]
- Haselager, M.; Thijssen, R.; West, C.; Young, L.; Van Kampen, R.; Willmore, E.; Mackay, S.; Kater, A.; Eldering, E. Regulation of Bcl-XL by Non-Canonical NF-ΚB in the Context of CD40-Induced Drug Resistance in CLL. Cell Death Differ. 2021, 28, 1658–1668. [Google Scholar] [CrossRef]
- Tromp, J.M.; Geest, C.R.; Breij, E.C.W.; Elias, J.A.; van Laar, J.; Luijks, D.M.; Kater, A.P.; Beaumont, T.; van Oers, M.H.J.; Eldering, E. Tipping the Noxa/Mcl-1 Balance Overcomes ABT-737 Resistance in Chronic Lymphocytic Leukemia. Clin. Cancer Res. 2012, 18, 487–498. [Google Scholar] [CrossRef]
- Elías, E.E.; Almejún, M.B.; Colado, A.; Cordini, G.; Vergara-Rubio, M.; Podaza, E.; Risnik, D.; Cabrejo, M.; Fernández-Grecco, H.; Bezares, R.F.; et al. Autologous T-Cell Activation Fosters ABT-199 Resistance in Chronic Lymphocytic Leukemia: Rationale for a Combined Therapy with SYK Inhibitors and Anti-CD20 Monoclonal Antibodies. Haematologica 2018, 103, e458–e461. [Google Scholar] [CrossRef]
- Romano, M.F.; Lamberti, A.; Tassone, P.; Alfinito, F.; Costantini, S.; Chiurazzi, F.; Defrance, T.; Bonelli, P.; Tuccillo, F.; Turco, M.C.; et al. Triggering of CD40 Antigen Inhibits Fludarabine-Induced Apoptosis in B Chronic Lymphocytic Leukemia Cells. Blood 1998, 92, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Kielbassa, K. Ibrutinib Treatment in CLL Interrupts CD40 Signaling Capacity and Sensitizes CLL Cells to Venetoclax. Blood 2021, 138, 1545. [Google Scholar] [CrossRef]
- Shen, Y.; Best, O.G.; Mulligan, S.P.; Christopherson, R.I. Ibrutinib and Idelalisib Block Immunophenotypic Changes Associated with the Adhesion and Activation of CLL Cells in the Tumor Microenvironment. Leuk. Lymphoma 2018, 59, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Kielbassa, K.; ter Burg, J.; Klein, C.; Trumpfheller, C.; de Heer, K.; Kater, A.P.; Eldering, E. Co-Stimulatory versus Cell Death Aspects of Agonistic CD40 Monoclonal Antibody Selicrelumab in Chronic Lymphocytic Leukemia. Cancers 2021, 13, 3084. [Google Scholar] [CrossRef]
- Mraz, M.; Zent, C.S.; Church, A.K.; Jelinek, D.F.; Wu, X.; Pospisilova, S.; Ansell, S.M.; Novak, A.J.; Kay, N.E.; Witzig, T.E.; et al. Bone Marrow Stromal Cells Protect Lymphoma B-Cells from Rituximab-Induced Apoptosis and Targeting Integrin α-4-β-1 (VLA-4) with Natalizumab Can Overcome This Resistance. Br. J. Haematol. 2011, 155, 53–64. [Google Scholar] [CrossRef]
- Buchner, M.; Baer, C.; Prinz, G.; Dierks, C.; Burger, M.; Zenz, T.; Stilgenbauer, S.; Jumaa, H.; Veelken, H.; Zirlik, K. Spleen Tyrosine Kinase Inhibition Prevents Chemokine- and Integrin-Mediated Stromal Protective Effects in Chronic Lymphocytic Leukemia. Blood 2010, 115, 4497–4506. [Google Scholar] [CrossRef]
- Steele, A.J.; Prentice, A.G.; Cwynarski, K.; Hoffbrand, A.V.; Hart, S.M.; Lowdell, M.W.; Samuel, E.R.; Wickremasinghe, R.G. The JAK3-Selective Inhibitor PF-956980 Reverses the Resistance to Cytotoxic Agents Induced by Interleukin-4 Treatment of Chronic Lymphocytic Leukemia Cells: Potential for Reversal of Cytoprotection by the Microenvironment. Blood 2010, 116, 4569–4577. [Google Scholar] [CrossRef][Green Version]
- Ondrisova, L.; Mraz, M. Genetic and Non-Genetic Mechanisms of Resistance to BCR Signaling Inhibitors in B Cell Malignancies. Front. Oncol. 2020, 10, 591577. [Google Scholar] [CrossRef]
- Gowda, A.; Roda, J.; Hussain, S.-R.A.; Ramanunni, A.; Joshi, T.; Schmidt, S.; Zhang, X.; Lehman, A.; Jarjoura, D.; Carson, W.E.; et al. IL-21 Mediates Apoptosis through up-Regulation of the BH3 Family Member BIM and Enhances Both Direct and Antibody-Dependent Cellular Cytotoxicity in Primary Chronic Lymphocytic Leukemia Cells In Vitro. Blood 2008, 111, 4723–4730. [Google Scholar] [CrossRef]
- Ramsay, A.G.; Clear, A.J.; Fatah, R.; Gribben, J.G. Multiple Inhibitory Ligands Induce Impaired T-Cell Immunologic Synapse Function in Chronic Lymphocytic Leukemia That Can Be Blocked with Lenalidomide: Establishing a Reversible Immune Evasion Mechanism in Human Cancer. Blood 2012, 120, 1412–1421. [Google Scholar] [CrossRef]
- Browning, R.L.; Byrd, W.H.; Gupta, N.; Jones, J.; Mo, X.; Hertlein, E.; Yu, L.; Muthusamy, N.; Byrd, J.C. Lenalidomide Induces Interleukin-21 Production by T Cells and Enhances IL21-Mediated Cytotoxicity in Chronic Lymphocytic Leukemia B Cells. Cancer Immunol. Res. 2016, 4, 698–707. [Google Scholar] [CrossRef]
- Fanale, M.; Assouline, S.; Kuruvilla, J.; Solal-Céligny, P.; Heo, D.S.; Verhoef, G.; Corradini, P.; Abramson, J.S.; Offner, F.; Engert, A.; et al. Phase IA/II, Multicentre, Open-Label Study of the CD40 Antagonistic Monoclonal Antibody Lucatumumab in Adult Patients with Advanced Non-Hodgkin or Hodgkin Lymphoma. Br. J. Haematol. 2014, 164, 258–265. [Google Scholar] [CrossRef]
- Scielzo, C.; Ghia, P. Modeling the Leukemia Microenviroment In Vitro. Front. Oncol. 2020, 10, 607608. [Google Scholar] [CrossRef]
- Abarrategi, A.; Mian, S.A.; Passaro, D.; Rouault-Pierre, K.; Grey, W.; Bonnet, D. Modeling the Human Bone Marrow Niche in Mice: From Host Bone Marrow Engraftment to Bioengineering Approaches. J. Exp. Med. 2018, 215, 729–743. [Google Scholar] [CrossRef]
- Antonelli, A.; Noort, W.A.; Jaques, J.; de Boer, B.; de Jong-Korlaar, R.; Brouwers-Vos, A.Z.; Lubbers-Aalders, L.; van Velzen, J.F.; Bloem, A.C.; Yuan, H.; et al. Establishing Human Leukemia Xenograft Mouse Models by Implanting Human Bone Marrow–like Scaffold-Based Niches. Blood 2016, 128, 2949–2959. [Google Scholar] [CrossRef]
- Shah, S.B.; Singh, A. Creating Artificial Lymphoid Tissues to Study Immunity and Hematological Malignancies. Curr. Opin. Hematol 2017, 24, 377–383. [Google Scholar] [CrossRef]
- Tavakol, D.N.; Tratwal, J.; Bonini, F.; Genta, M.; Campos, V.; Burch, P.; Hoehnel, S.; Béduer, A.; Alessandrini, M.; Naveiras, O.; et al. Injectable, Scalable 3D Tissue-Engineered Model of Marrow Hematopoiesis. Biomaterials 2020, 232, 119665. [Google Scholar] [CrossRef]
- Bello, A.B.; Park, H.; Lee, S.-H. Current Approaches in Biomaterial-Based Hematopoietic Stem Cell Niches. Acta Biomater. 2018, 72, 1–15. [Google Scholar] [CrossRef]
- Sbrana, F.V.; Pinos, R.; Barbaglio, F.; Ribezzi, D.; Scagnoli, F.; Scarfò, L.; Redwan, I.N.; Martinez, H.; Farè, S.; Ghia, P.; et al. 3D Bioprinting Allows the Establishment of Long-Term 3D Culture Model for Chronic Lymphocytic Leukemia Cells. Front. Immunol. 2021, 12, 639572. [Google Scholar] [CrossRef]
- Svozilová, H.; Plichta, Z.; Proks, V.; Studená, R.; Baloun, J.; Doubek, M.; Pospíšilová, Š.; Horák, D. RGDS-Modified Superporous Poly(2-Hydroxyethyl Methacrylate)-Based Scaffolds as 3D In Vitro Leukemia Model. Int. J. Mol. Sci. 2021, 22, 2376. [Google Scholar] [CrossRef]
- Roh, K.-H.; Song, H.W.; Pradhan, P.; Bai, K.; Bohannon, C.D.; Dale, G.; Leleux, J.; Jacob, J.; Roy, K. A Synthetic Stroma-Free Germinal Center Niche for Efficient Generation of Humoral Immunity Ex Vivo. Biomaterials 2018, 164, 106–120. [Google Scholar] [CrossRef]
- Purwada, A.; Singh, A. Immuno-Engineered Organoids for Regulating the Kinetics of B-Cell Development and Antibody Production. Nat. Protoc. 2017, 12, 168–182. [Google Scholar] [CrossRef]
- Walsby, E.; Buggins, A.; Devereux, S.; Jones, C.; Pratt, G.; Brennan, P.; Fegan, C.; Pepper, C. Development and Characterization of a Physiologically Relevant Model of Lymphocyte Migration in Chronic Lymphocytic Leukemia. Blood 2014, 123, 3607–3617. [Google Scholar] [CrossRef]
- Pasikowska, M.; Walsby, E.; Apollonio, B.; Cuthill, K.; Phillips, E.; Coulter, E.; Longhi, M.S.; Ma, Y.; Yallop, D.; Barber, L.D.; et al. Phenotype and Immune Function of Lymph Node and Peripheral Blood CLL Cells Are Linked to Transendothelial Migration. Blood 2016, 128, 563–573. [Google Scholar] [CrossRef]
- Barbaglio, F.; Belloni, D.; Scarfò, L.; Sbrana, F.V.; Ponzoni, M.; Bongiovanni, L.; Pavesi, L.; Zambroni, D.; Stamatopoulos, K.; Caiolfa, V.R.; et al. Three-Dimensional Co-Culture Model of Chronic Lymphocytic Leukemia Bone Marrow Microenvironment Predicts Patient-Specific Response to Mobilizing Agents. Haematologica 2021, 106, 2334–2344. [Google Scholar] [CrossRef]
- Lamaison, C.; Latour, S.; Hélaine, N.; Le Morvan, V.; Saint-Vanne, J.; Mahouche, I.; Monvoisin, C.; Dussert, C.; Andrique, L.; Deleurme, L.; et al. A Novel 3D Culture Model Recapitulates Primary FL B-Cell Features and Promotes Their Survival. Blood Adv. 2021, 5, 5372–5386. [Google Scholar] [CrossRef]
- Simonetti, G.; Bertilaccio, M.T.S.; Ghia, P.; Klein, U. Mouse Models in the Study of Chronic Lymphocytic Leukemia Pathogenesis and Therapy. Blood 2014, 124, 1010–1019. [Google Scholar] [CrossRef]
- Bresin, A.; D’Abundo, L.; Narducci, M.G.; Fiorenza, M.T.; Croce, C.M.; Negrini, M.; Russo, G. TCL1 Transgenic Mouse Model as a Tool for the Study of Therapeutic Targets and Microenvironment in Human B-Cell Chronic Lymphocytic Leukemia. Cell Death Dis. 2016, 7, e2071. [Google Scholar] [CrossRef]
- Bichi, R.; Shinton, S.A.; Martin, E.S.; Koval, A.; Calin, G.A.; Cesari, R.; Russo, G.; Hardy, R.R.; Croce, C.M. Human Chronic Lymphocytic Leukemia Modeled in Mouse by Targeted TCL1 Expression. Proc. Natl. Acad. Sci. USA 2002, 99, 6955–6960. [Google Scholar] [CrossRef]
- Gorgun, G.; Ramsay, A.G.; Holderried, T.A.W.; Zahrieh, D.; Le Dieu, R.; Liu, F.; Quackenbush, J.; Croce, C.M.; Gribben, J.G. Eµ-TCL1 Mice Represent a Model for Immunotherapeutic Reversal of Chronic Lymphocytic Leukemia-Induced T-Cell Dysfunction. Proc. Natl. Acad. Sci. USA 2009, 106, 6250–6255. [Google Scholar] [CrossRef]
- Hofbauer, J.P.; Heyder, C.; Denk, U.; Kocher, T.; Holler, C.; Trapin, D.; Asslaber, D.; Tinhofer, I.; Greil, R.; Egle, A. Development of CLL in the TCL1 Transgenic Mouse Model Is Associated with Severe Skewing of the T-Cell Compartment Homologous to Human CLL. Leukemia 2011, 25, 1452–1458. [Google Scholar] [CrossRef] [PubMed]
- Kocher, T.; Asslaber, D.; Zaborsky, N.; Flenady, S.; Denk, U.; Reinthaler, P.; Ablinger, M.; Geisberger, R.; Bauer, J.W.; Seiffert, M.; et al. CD4+ T Cells, but Not Non-Classical Monocytes, Are Dispensable for the Development of Chronic Lymphocytic Leukemia in the TCL1-Tg Murine Model. Leukemia 2016, 30, 1409–1413. [Google Scholar] [CrossRef] [PubMed]
- Grioni, M.; Brevi, A.; Cattaneo, E.; Rovida, A.; Bordini, J.; Bertilaccio, M.T.S.; Ponzoni, M.; Casorati, G.; Dellabona, P.; Ghia, P.; et al. CD4+ T Cells Sustain Aggressive Chronic Lymphocytic Leukemia in Eμ-TCL1 Mice through a CD40L-Independent Mechanism. Blood Adv. 2021, 5, 2817–2828. [Google Scholar] [CrossRef] [PubMed]
- Zapata, J.M.; Krajewska, M.; Morse, H.C.; Choi, Y.; Reed, J.C. TNF Receptor-Associated Factor (TRAF) Domain and Bcl-2 Cooperate to Induce Small B Cell Lymphoma/Chronic Lymphocytic Leukemia in Transgenic Mice. Proc. Natl. Acad. Sci. USA 2004, 101, 16600–16605. [Google Scholar] [CrossRef]
- Munzert, G.; Kirchner, D.; Stobbe, H.; Bergmann, L.; Schmid, R.M.; Döhner, H.; Heimpel, H. Tumor Necrosis Factor Receptor-Associated Factor 1 Gene Overexpression in B-Cell Chronic Lymphocytic Leukemia: Analysis of NF-ΚB/Rel–Regulated Inhibitors of Apoptosis. Blood 2002, 100, 3749–3756. [Google Scholar] [CrossRef]
- Elgueta, R.; Benson, M.J.; de Vries, V.C.; Wasiuk, A.; Guo, Y.; Noelle, R.J. Molecular Mechanism and Function of CD40/CD40L Engagement in the Immune System. Immunol. Rev. 2009, 229, 152–172. [Google Scholar] [CrossRef]
- Lee, S.Y.; Reichlin, A.; Santana, A.; Sokol, K.A.; Nussenzweig, M.C.; Choi, Y. TRAF2 Is Essential for JNK but Not NF-KappaB Activation and Regulates Lymphocyte Proliferation and Survival. Immunity 1997, 7, 703–713. [Google Scholar] [CrossRef]
- Shimoni, A.; Marcus, H.; Canaan, A.; Ergas, D.; David, M.; Berrebi, A.; Reisner, Y. A Model for Human B-Chronic Lymphocytic Leukemia in Human/Mouse Radiation Chimera: Evidence for Tumor-Mediated Suppression of Antibody Production in Low-Stage Disease. Blood 1997, 89, 2210–2218. [Google Scholar] [CrossRef]
- Shimoni, A.; Marcus, H.; Dekel, B.; Shkarchi, R.; Arditti, F.; Shvidel, L.; Shtalrid, M.; Bucher, W.; Canaan, A.; Ergas, D.; et al. Autologous T Cells Control B-Chronic Lymphocytic Leukemia Tumor Progression in Human→Mouse Radiation Chimera. Cancer Res. 1999, 59, 5968–5974. [Google Scholar]
- Dürig, J.; Ebeling, P.; Grabellus, F.; Sorg, U.R.; Möllmann, M.; Schütt, P.; Göthert, J.; Sellmann, L.; Seeber, S.; Flasshove, M.; et al. A Novel Nonobese Diabetic/Severe Combined Immunodeficient Xenograft Model for Chronic Lymphocytic Leukemia Reflects Important Clinical Characteristics of the Disease. Cancer Res. 2007, 67, 8653–8661. [Google Scholar] [CrossRef]
- Aydin, S.; Grabellus, F.; Eisele, L.; Möllmann, M.; Hanoun, M.; Ebeling, P.; Moritz, T.; Carpinteiro, A.; Nückel, H.; Sak, A.; et al. Investigating the Role of CD38 and Functionally Related Molecular Risk Factors in the CLL NOD/SCID Xenograft Model. Eur. J. Haematol. 2011, 87, 10–19. [Google Scholar] [CrossRef]
- Bagnara, D.; Kaufman, M.S.; Calissano, C.; Marsilio, S.; Patten, P.E.M.; Simone, R.; Chum, P.; Yan, X.-J.; Allen, S.L.; Kolitz, J.E.; et al. A Novel Adoptive Transfer Model of Chronic Lymphocytic Leukemia Suggests a Key Role for T Lymphocytes in the Disease. Blood 2011, 117, 5463–5472. [Google Scholar] [CrossRef]
- Patten, P.E.M.; Ferrer, G.; Chen, S.-S.; Kolitz, J.E.; Rai, K.R.; Allen, S.L.; Barrientos, J.C.; Ioannou, N.; Ramsay, A.G.; Chiorazzi, N. A Detailed Analysis of Parameters Supporting the Engraftment and Growth of Chronic Lymphocytic Leukemia Cells in Immune-Deficient Mice. Front. Immunol. 2021, 12, 627020. [Google Scholar] [CrossRef]
- Oldreive, C.E.; Skowronska, A.; Davies, N.J.; Parry, H.; Agathanggelou, A.; Krysov, S.; Packham, G.; Rudzki, Z.; Cronin, L.; Vrzalikova, K.; et al. T-Cell Number and Subtype Influence the Disease Course of Primary Chronic Lymphocytic Leukaemia Xenografts in Alymphoid Mice. Dis. Models Mech. 2015, 8, 1401–1412. [Google Scholar] [CrossRef]
- Decker, S.; Zwick, A.; Khaja Saleem, S.; Kissel, S.; Rettig, A.; Aumann, K.; Dierks, C. Optimized Xenograft Protocol for Chronic Lymphocytic Leukemia Results in High Engraftment Efficiency for All CLL Subgroups. Int. J. Mol. Sci. 2019, 20, 6277. [Google Scholar] [CrossRef]
- Davies, N.J.; Kwok, M.; Gould, C.; Oldreive, C.E.; Mao, J.; Parry, H.; Smith, E.; Agathanggelou, A.; Pratt, G.; Taylor, A.M.R.; et al. Dynamic Changes in Clonal Cytogenetic Architecture during Progression of Chronic Lymphocytic Leukemia in Patients and Patient-Derived Murine Xenografts. Oncotarget 2017, 8, 44749–44760. [Google Scholar] [CrossRef]
- Herman, S.E.M.; Sun, X.; McAuley, E.M.; Hsieh, M.M.; Pittaluga, S.; Raffeld, M.; Liu, D.; Keyvanfar, K.; Chapman, C.M.; Chen, J.; et al. Modeling Tumor-Host Interactions of Chronic Lymphocytic Leukemia in Xenografted Mice to Study Tumor Biology and Evaluate Targeted Therapy. Leukemia 2013, 27, 2311–2321. [Google Scholar] [CrossRef]
- Patten, P.E.M.; Ferrer, G.; Chen, S.-S.; Simone, R.; Marsilio, S.; Yan, X.-J.; Gitto, Z.; Yuan, C.; Kolitz, J.E.; Barrientos, J.; et al. Chronic Lymphocytic Leukemia Cells Diversify and Differentiate in Vivo via a Nonclassical Th1-Dependent, Bcl-6-Deficient Process. JCI Insight 2016, 1, e86288. [Google Scholar] [CrossRef]
- Nagatani, M.; Kodera, T.; Suzuki, D.; Igura, S.; Fukunaga, Y.; Kanemitsu, H.; Nakamura, D.; Mochizuki, M.; Kemi, M.; Tamura, K.; et al. Comparison of Biological Features between Severely Immuno-Deficient NOD/Shi-Scid Il2rgnull and NOD/LtSz-Scid Il2rgnull Mice. Exp. Anim. 2019, 68, 471–482. [Google Scholar] [CrossRef]
- Kikushige, Y.; Ishikawa, F.; Miyamoto, T.; Shima, T.; Urata, S.; Yoshimoto, G.; Mori, Y.; Iino, T.; Yamauchi, T.; Eto, T.; et al. Self-Renewing Hematopoietic Stem Cell Is the Primary Target in Pathogenesis of Human Chronic Lymphocytic Leukemia. Cancer Cell 2011, 20, 246–259. [Google Scholar] [CrossRef]
- Burack, W.R.; Spence, J.M.; Spence, J.P.; Spence, S.A.; Rock, P.J.; Shenoy, G.N.; Shultz, L.D.; Bankert, R.B.; Bernstein, S.H. Patient-Derived Xenografts of Low-Grade B-Cell Lymphomas Demonstrate Roles of the Tumor Microenvironment. Blood Adv. 2017, 1, 1263–1273. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoferkova, E.; Kadakova, S.; Mraz, M. In Vitro and In Vivo Models of CLL–T Cell Interactions: Implications for Drug Testing. Cancers 2022, 14, 3087. https://doi.org/10.3390/cancers14133087
Hoferkova E, Kadakova S, Mraz M. In Vitro and In Vivo Models of CLL–T Cell Interactions: Implications for Drug Testing. Cancers. 2022; 14(13):3087. https://doi.org/10.3390/cancers14133087
Chicago/Turabian StyleHoferkova, Eva, Sona Kadakova, and Marek Mraz. 2022. "In Vitro and In Vivo Models of CLL–T Cell Interactions: Implications for Drug Testing" Cancers 14, no. 13: 3087. https://doi.org/10.3390/cancers14133087
APA StyleHoferkova, E., Kadakova, S., & Mraz, M. (2022). In Vitro and In Vivo Models of CLL–T Cell Interactions: Implications for Drug Testing. Cancers, 14(13), 3087. https://doi.org/10.3390/cancers14133087