
ADAM and ADAMTS Proteins, New Players in the Regulation of Hepatocellular Carcinoma Microenvironment

Abstract
Simple Summary
Abstract
1. Adamalysins Are Multidomain Proteins with Multiple Functions



2. Adamalysins in Hepatocellular Carcinoma
2.1. Adamalysin Expression and Association with Overall Survival
2.2. Implication of Adamalysins in Inflammation and HCC Progression
2.2.1. Regulation of Pro-Tumorigenic Cytokines by Adamalysins in HCC
2.2.2. Role of Adamalysins in Tumor Escape from Immune Surveillance
2.2.3. Adamalysins Regulate Invasion in HCC
Author Contributions
Funding
Conflicts of Interest
References
- Seals, D.F.; Courtneidge, S.A. The ADAMs family of metalloproteases: Multidomain proteins with multiple functions. Genes Dev. 2003, 17, 7–30. [Google Scholar] [CrossRef]
- Apte, S.S. A Disintegrin-like and Metalloprotease (Reprolysin-Type) with Thrombospondin Type 1 Motif (ADAMTS) Superfamily: Functions and Mechanisms. J. Biol. Chem. 2009, 284, 31493–31497. [Google Scholar] [CrossRef] [PubMed]
- Kelwick, R.; Desanlis, I.; Wheeler, G.N.; Edwards, D.R. The ADAMTS (A Disintegrin and Metalloproteinase with Thrombospondin Motifs) Family. Genome Biol. 2015, 16, 113. [Google Scholar] [CrossRef]
- Huovila, A.-P.J.; Turner, A.J.; Pelto-Huikko, M.; Kärkkäinen, I.; Ortiz, R.M. Shedding Light on ADAM Metalloproteinases. Trends Biochem. Sci. 2005, 30, 413–422. [Google Scholar] [CrossRef]
- Zadka, L.; Kulus, M.J.; Piatek, K. ADAM Protein Family—Its Role in Tumorigenesis, Mechanisms of Chemoresistance and Potential as Diagnostic and Prognostic Factors. Neoplasma 2018, 65, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Blobel, C.P. ADAMs: Key Components in EGFR signalling and development. Nat. Rev. Mol. Cell Biol. 2005, 6, 32–43. [Google Scholar] [CrossRef]
- Christian, L.M. The ADAM family: Insights into notch proteolysis. Fly 2012, 6, 30–34. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zolkiewska, A. ADAM proteases: Ligand processing and modulation of the notch pathway. Cell Mol. Life Sci. 2008, 65, 2056–2068. [Google Scholar] [CrossRef]
- Atfi, A.; Dumont, E.; Colland, F.; Bonnier, D.; L’helgoualc’h, A.; Prunier, C.; Ferrand, N.; Clément, B.; Wewer, U.M.; Théret, N. The disintegrin and metalloproteinase ADAM12 contributes to TGF-Beta signaling through interaction with the Type II receptor. J. Cell Biol. 2007, 178, 201–208. [Google Scholar] [CrossRef]
- Reiss, K.; Ludwig, A.; Saftig, P. Breaking up the Tie: Disintegrin-like metalloproteinases as regulators of cell migration in inflammation and invasion. Pharmacol. Ther 2006, 111, 985–1006. [Google Scholar] [CrossRef] [PubMed]
- Dancevic, C.M.; McCulloch, D.R.; Ward, A.C. The ADAMTS hyalectanase family: Biological insights from diverse species. Biochem. J. 2016, 473, 2011–2022. [Google Scholar] [CrossRef]
- Bekhouche, M.; Leduc, C.; Dupont, L.; Janssen, L.; Delolme, F.; Vadon-Le Goff, S.; Smargiasso, N.; Baiwir, D.; Mazzucchelli, G.; Zanella-Cleon, I.; et al. Determination of the Substrate Repertoire of ADAMTS2, 3, and 14 significantly broadens their functions and identifies extracellular matrix organization and TGF-β signaling as primary targets. FASEB J. 2016, 30, 1741–1756. [Google Scholar] [CrossRef]
- Schnellmann, R.; Sack, R.; Hess, D.; Annis, D.S.; Mosher, D.F.; Apte, S.S.; Chiquet-Ehrismann, R. A selective extracellular matrix proteomics approach identifies fibronectin proteolysis by a disintegrin-like and metalloprotease domain with thrombospondin Type 1 Motifs (ADAMTS16) and its impact on spheroid morphogenesis. Mol. Cell Proteom. 2018, 17, 1410–1425. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Nandadasa, S.; Annis, D.S.; Dubail, J.; Mosher, D.F.; Willard, B.B.; Apte, S.S. A disintegrin-like and metalloproteinase domain with thrombospondin Type 1 Motif 9 (ADAMTS9) regulates fibronectin fibrillogenesis and turnover. J. Biol. Chem. 2019, 294, 9924–9936. [Google Scholar] [CrossRef]
- Colige, A.; Monseur, C.; Crawley, J.T.B.; Santamaria, S.; de Groot, R. Proteomic discovery of substrates of the cardiovascular protease ADAMTS. J. Biol. Chem. 2019, 294, 8037–8045. [Google Scholar] [CrossRef]
- Wang, L.W.; Kutz, W.E.; Mead, T.J.; Beene, L.C.; Singh, S.; Jenkins, M.W.; Reinhardt, D.P.; Apte, S.S. Adamts10 inactivation in mice leads to persistence of ocular microfibrils subsequent to reduced Fibrillin-2 cleavage. Matrix Biol. 2019, 77, 117–128. [Google Scholar] [CrossRef]
- Satz-Jacobowitz, B.; Hubmacher, D. The Quest for Substrates and Binding Partners: A Critical Barrier for Understanding the Role of ADAMTS Proteases in Musculoskeletal Development and Disease. Dev. Dyn. 2020. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S. Three-dimensional domain architecture of the ADAM family proteinases. Semin. Cell Dev. Biol. 2009, 20, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Hubmacher, D.; Apte, S.S. ADAMTS proteins as modulators of microfibril formation and function. Matrix Biol. 2015, 47, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, C.; Cormier-Daire, V. The ADAMTS(L) family and human genetic disorders. Hum. Mol. Genet. 2011, 20, R163–R167. [Google Scholar] [CrossRef] [PubMed]
- Mead, T.J.; Apte, S.S. ADAMTS proteins in human disorders. Matrix Biol. 2018, 71–72, 225–239. [Google Scholar] [CrossRef]
- Duffy, M.J.; McKiernan, E.; O’Donovan, N.; McGowan, P.M. Role of ADAMs in cancer formation and progression. Clin. Cancer Res. 2009, 15, 1140–1144. [Google Scholar] [CrossRef]
- Binder, M.J.; McCoombe, S.; Williams, E.D.; McCulloch, D.R.; Ward, A.C. The extracellular matrix in cancer progression: Role of hyalectan proteoglycans and ADAMTS enzymes. Cancer Lett. 2017, 385, 55–64. [Google Scholar] [CrossRef]
- Cal, S.; López-Otín, C. ADAMTS proteases and cancer. Matrix Biol. 2015, 44–46, 77–85. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, J.; Yang, Z. The roles of ADAMTS in angiogenesis and cancer. Tumour Biol. 2015, 36, 4039–4051. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Ohtsuka, T.; Okada, Y.; Kanai, Y. Stromal metalloproteinases: Crucial contributors to the tumor microenvironment. Pathol. Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mullooly, M.; McGowan, P.M.; Crown, J.; Duffy, M.J. The ADAMs family of proteases as targets for the treatment of cancer. Cancer Biol. Ther. 2016, 17, 870–880. [Google Scholar] [CrossRef]
- Mazzocca, A.; Giannelli, G.; Antonaci, S. Involvement of ADAMs in tumorigenesis and progression of hepatocellular carcinoma: Is it merely fortuitous or a real pathogenic link? Biochim. Biophys. Acta 2010, 1806, 74–81. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, Y.; Yu, H.-F.; Yu, X.-T.; Zhou, S.-J.; Tan, Y.-F. Expression of ADAM8 and its clinical values in diagnosis and prognosis of hepatocellular carcinoma. Tumour Biol. 2012, 33, 2167–2172. [Google Scholar] [CrossRef]
- Zhang, Y.; Zha, T.-Z.; Hu, B.-S.; Jiang, C.; Ge, Z.-J.; Zhang, K.; Tan, Y.-F. High expression of ADAM8 correlates with poor prognosis in hepatocellular carcinoma. Surgeon 2013, 11, 67–71. [Google Scholar] [CrossRef]
- Tannapfel, A.; Anhalt, K.; Häusermann, P.; Sommerer, F.; Benicke, M.; Uhlmann, D.; Witzigmann, H.; Hauss, J.; Wittekind, C. Identification of novel proteins associated with hepatocellular carcinomas using protein microarrays. J. Pathol. 2003, 201, 238–249. [Google Scholar] [CrossRef]
- Tao, K.; Qian, N.; Tang, Y.; Ti, Z.; Song, W.; Cao, D.; Dou, K. Increased expression of a disintegrin and metalloprotease-9 in hepatocellular carcinoma: Implications for tumor progression and prognosis. Jpn. J. Clin. Oncol. 2010, 40, 645–651. [Google Scholar] [CrossRef]
- Yuan, S.; Lei, S.; Wu, S. ADAM10 is overexpressed in human hepatocellular carcinoma and contributes to the proliferation, invasion and migration of HepG2 cells. Oncol. Rep. 2013, 30, 1715–1722. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, S.; Liu, K.; Wang, Y.; Ji, B.; Zhang, X.; Liu, Y. A Disintegrin and Metalloprotease (ADAM)10 is highly expressed in Hepatocellular Carcinoma and is associated with tumour progression. J. Int. Med. Res. 2014, 42, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Le Pabic, H.; Bonnier, D.; Wewer, U.M.; Coutand, A.; Musso, O.; Baffet, G.; Clément, B.; Théret, N. ADAM12 in human liver cancers: TGF-Beta-Regulated expression in stellate cells is associated with matrix remodeling. Hepatology 2003, 37, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Yang, L.-Y.; Huang, G.-W.; Wang, W.; Lu, W.-Q. ADAM17 MRNA expression and pathological features of hepatocellular carcinoma. World J. Gastroenterol. 2004, 10, 2735–2739. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Shen, S.; Fu, S.; Preston, B.; Brandon, C.; He, S.; Shen, C.; Wu, J.; Wang, S.; Xie, W.; et al. MiR-203 Suppresses the proliferation and metastasis of Hepatocellular Carcinoma by targeting Oncogene ADAM9 and Oncogenic Long Non-Coding RNA HULC. Anticancer Agents Med. Chem. 2016, 16, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.-Y.; Ou, H.-H.; Liu, X.-C.; Chen, Z.-J.; Li, X.-H.; Huang, Y.; Yang, D.-H. Loss of Tumor Suppressor MiR-126 Contributes to the development of Hepatitis B Virus-Related Hepatocellular Carcinoma metastasis through the upregulation of ADAM. Tumour Biol. 2017, 39, 1010428317709128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, J.; Li, Y.; Liu, L.; Zhang, X.; Ma, C.; Hua, S.; Yang, M.; Yuan, Q. MicroRNA-1274a, a Modulator of Sorafenib induced a disintegrin and Metalloproteinase 9 (ADAM9) down-regulation in Hepatocellular Carcinoma. FEBS Lett. 2011, 585, 1828–1834. [Google Scholar] [CrossRef]
- Bai, S.; Nasser, M.W.; Wang, B.; Hsu, S.-H.; Datta, J.; Kutay, H.; Yadav, A.; Nuovo, G.; Kumar, P.; Ghoshal, K. MicroRNA-122 inhibits tumorigenic properties of Hepatocellular Carcinoma cells and sensitizes these cells to Sorafenib. J. Biol. Chem. 2009, 284, 32015–32027. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, K.; Zhang, W.; Wang, Y.; Jin, Z.; Jia, B.; Liu, Y. MiR-449a inhibits proliferation and invasion by regulating ADAM10 in Hepatocellular Carcinoma. Am. J. Transl. Res. 2016, 8, 2609–2619. [Google Scholar]
- Liu, Y.; Zhang, W.; Liu, S.; Liu, K.; Ji, B.; Wang, Y. MiR-365 Targets ADAM10 and suppresses the cell growth and metastasis of Hepatocellular Carcinoma. Oncol. Rep. 2017, 37, 1857–1864. [Google Scholar] [CrossRef][Green Version]
- Nakao, K.; Miyaaki, H.; Ichikawa, T. Antitumor function of MicroRNA-122 against Hepatocellular Carcinoma. J. Gastroenterol. 2014, 49, 589–593. [Google Scholar] [CrossRef]
- Wu, G.; Zheng, K.; Xia, S.; Wang, Y.; Meng, X.; Qin, X.; Cheng, Y. MicroRNA-655-3p functions as a tumor suppressor by regulating ADAM10 and β-Catenin pathway in Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 89. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, L.; Chen, G.; Chen, L.; Li, Y.; Lai, Y.; Weng, H.; Chen, T.; Wang, L.; Ye, Y. Human umbilical cord mesenchymal stem cells-derived exosomal MicroRNA-451a represses epithelial-mesenchymal transition of hepatocellular carcinoma cells by inhibiting ADAM. RNA Biol. 2020. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, C.; Wang, Y.; Wen, S.; Wang, J.; Chen, Z.; He, Q.; Feng, D. MicroRNA-145 inhibits cell proliferation by directly targeting ADAM17 in Hepatocellular Carcinoma. Oncol. Rep. 2014, 32, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-C.; Hsu, P.W.-C.; Lai, T.-C.; Chau, G.-Y.; Lin, C.-W.; Chen, C.-M.; Lin, C.-D.; Liao, Y.-L.; Wang, J.-L.; Chau, Y.-P.; et al. MicroRNA-122, a tumor suppressor MicroRNA that regulates intrahepatic metastasis of Hepatocellular Carcinoma. Hepatology 2009, 49, 1571–1582. [Google Scholar] [CrossRef]
- Yang, B.; Wang, C.; Xie, H.; Wang, Y.; Huang, J.; Rong, Y.; Zhang, H.; Kong, H.; Yang, Y.; Lu, Y. MicroRNA-3163 Targets ADAM-17 and enhances the sensitivity of Hepatocellular Carcinoma cells to molecular targeted agents. Cell Death Dis. 2019, 10, 784. [Google Scholar] [CrossRef]
- Yang, X.-W.; Zhang, L.-J.; Huang, X.-H.; Chen, L.-Z.; Su, Q.; Zeng, W.-T.; Li, W.; Wang, Q. MiR-145 Suppresses cell invasion in Hepatocellular Carcinoma Cells: MiR-145 targets ADAM. Hepatol. Res. 2014, 44, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.-Y.; Ding, J.; Tian, W.-G.; Yu, Y.-C. MicroRNA-122 as a diagnostic biomarker for Hepatocellular Carcinoma related to Hepatitis C Virus: A meta-analysis and systematic review. J. Int. Med. Res. 2020, 48, 300060520941634. [Google Scholar] [CrossRef]
- Gao, J.; Yin, X.; Yu, X.; Dai, C.; Zhou, F. Long Noncoding LINC01551 promotes hepatocellular carcinoma cell proliferation, migration, and invasion by acting as a competing endogenous RNA of MicroRNA-122-5p to regulate ADAM10 expression. J. Cell Biochem. 2019, 120, 16393–16407. [Google Scholar] [CrossRef]
- Mazzocca, A.; Coppari, R.; De Franco, R.; Cho, J.-Y.; Libermann, T.A.; Pinzani, M.; Toker, A. A secreted form of ADAM9 promotes Carcinoma invasion through Tumor-Stromal Interactions. Cancer Res. 2005, 65, 4728–4738. [Google Scholar] [CrossRef]
- Shiu, J.-S.; Hsieh, M.-J.; Chiou, H.-L.; Wang, H.-L.; Yeh, C.-B.; Yang, S.-F.; Chou, Y.-E. Impact of ADAM10 gene polymorphisms on Hepatocellular Carcinoma development and clinical characteristics. Int. J. Med. Sci. 2018, 15, 1334–1340. [Google Scholar] [CrossRef]
- Honda, H.; Takamura, M.; Yamagiwa, S.; Genda, T.; Horigome, R.; Kimura, N.; Setsu, T.; Tominaga, K.; Kamimura, H.; Matsuda, Y.; et al. Overexpression of a disintegrin and Metalloproteinase 21 is associated with motility, metastasis, and poor prognosis in Hepatocellular Carcinoma. Sci. Rep. 2017, 7, 15485. [Google Scholar] [CrossRef] [PubMed]
- Zou, B.; Liu, X.; Gong, Y.; Cai, C.; Li, P.; Xing, S.; Pokhrel, B.; Zhang, B.; Li, J. A Novel 12-Marker panel of cancer-associated fibroblasts involved in progression of Hepatocellular Carcinoma. Cancer Manag. Res. 2018, 10, 5303–5311. [Google Scholar] [CrossRef]
- Oh, S.; Park, Y.; Lee, H.-J.; Lee, J.; Lee, S.-H.; Baek, Y.-S.; Chun, S.-K.; Lee, S.-M.; Kim, M.; Chon, Y.-E.; et al. A Disintegrin and Metalloproteinase 9 (ADAM9) in advanced Hepatocellular Carcinoma and their role as a Biomarker during Hepatocellular Carcinoma immunotherapy. Cancers 2020, 12, 745. [Google Scholar] [CrossRef] [PubMed]
- Mattern, J.; Roghi, C.S.; Hurtz, M.; Knäuper, V.; Edwards, D.R.; Poghosyan, Z. ADAM15 mediates upregulation of claudin-1 expression in breast cancer cells. Sci. Rep. 2019, 9, 12540. [Google Scholar] [CrossRef]
- Mailly, L.; Baumert, T.F. Hepatitis C Virus infection and tight junction proteins: The ties that bind. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183296. [Google Scholar] [CrossRef] [PubMed]
- Charrier-Hisamuddin, L.; Laboisse, C.L.; Merlin, D. ADAM-15: A Metalloprotease that mediates inflammation. FASEB J. 2008, 22, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, L.; Kelwick, R.; Decock, J.; Edwards, D.R. The Roles of ADAMTS Metalloproteinases in tumorigenesis and metastasis. Front. Biosci. 2011, 16, 1861–1872. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cheung, S.T.; So, S.; Fan, S.T.; Barry, C.; Higgins, J.; Lai, K.-M.; Ji, J.; Dudoit, S.; Ng, I.O.L.; et al. Gene Expression Patterns in Human Liver Cancers. Mol. Biol. Cell 2002, 13, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Masui, T.; Hosotani, R.; Tsuji, S.; Miyamoto, Y.; Yasuda, S.; Ida, J.; Nakajima, S.; Kawaguchi, M.; Kobayashi, H.; Koizumi, M.; et al. Expression of METH-1 and METH-2 in pancreatic cancer. Clin. Cancer Res. 2001, 7, 3437–3443. [Google Scholar]
- Braconi, C.; Meng, F.; Swenson, E.; Khrapenko, L.; Huang, N.; Patel, T. Candidate therapeutic agents for Hepatocellular Cancer can be identified from phenotype-associated gene expression signatures. Cancer 2009, 115, 3738–3748. [Google Scholar] [CrossRef]
- Turner, S.L.; Mangnall, D.; Bird, N.C.; Bunning, R.A.D.; Blair-Zajdel, M.E. Expression of ADAMTS-1, ADAMTS-4, ADAMTS-5 and TIMP3 by Hepatocellular Carcinoma cell lines. Int. J. Oncol. 2012, 41, 1043–1049. [Google Scholar] [CrossRef][Green Version]
- Turkoglu, S.A.; Kockar, F. SP1 and USF differentially regulate ADAMTS1 gene expression under normoxic and hypoxic conditions in Hepatoma Cells. Gene 2016, 575, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Diamantis, I.; Lüthi, M.; Hösli, M.; Reichen, J. Cloning of the Rat ADAMTS-1 Gene and its down regulation in endothelial cells in cirrhotic rats. Liver 2000, 20, 165–172. [Google Scholar] [CrossRef]
- Bourd-Boittin, K.; Bonnier, D.; Leyme, A.; Mari, B.; Tuffery, P.; Samson, M.; Ezan, F.; Baffet, G.; Theret, N. Protease Profiling of Liver Fibrosis Reveals the ADAM Metallopeptidase with Thrombospondin Type 1 Motif, 1 as a Central Activator of Transforming Growth Factor Beta. Hepatology 2011, 54, 2173–2184. [Google Scholar] [CrossRef] [PubMed]
- Schwettmann, L.; Wehmeier, M.; Jokovic, D.; Aleksandrova, K.; Brand, K.; Manns, M.P.; Lichtinghagen, R.; Bahr, M.J. Hepatic Expression of A Disintegrin and Metalloproteinase (ADAM) and ADAMs with Thrombospondin Motives (ADAM-TS) Enzymes in patients with chronic liver diseases. J. Hepatol. 2008, 49, 243–250. [Google Scholar] [CrossRef]
- Li, C.; Xiong, Y.; Yang, X.; Wang, L.; Zhang, S.; Dai, N.; Li, M.; Ren, T.; Yang, Y.; Zhou, S.-F.; et al. Lost expression of ADAMTS5 protein associates with progression and poor prognosis of Hepatocellular Carcinoma. Drug Des. Dev. Ther. 2015, 9, 1773–1783. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, C.; Wu, J.; Nan, Y. ADAMTS8 Targets ERK to suppress cell proliferation, invasion, and metastasis of Hepatocellular Carcinoma. Onco Targets Ther. 2018, 11, 7569–7578. [Google Scholar] [CrossRef]
- Li, S.; Li, T.; Li, X.; Yao, Y.; Jiang, X.; Zhao, L.; Guo, W. MicroRNA-32 regulates development and progression of hepatocellular carcinoma by targeting ADAMTS9 and affects its prognosis. Med. Sci. Monit. Basic Res. 2018, 24, 177–187. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, H.; Hu, Y.; Wang, F. Long Non-Coding RNA ADAMTS9-AS1 exacerbates cell proliferation, migration, and invasion via triggering of the PI3K/AKT/MTOR Pathway in Hepatocellular Carcinoma Cells. Am. J. Transl. Res. 2020, 12, 5696–5707. [Google Scholar]
- Bai, K.-H.; He, S.-Y.; Shu, L.-L.; Wang, W.-D.; Lin, S.-Y.; Zhang, Q.-Y.; Li, L.; Cheng, L.; Dai, Y.-J. Identification of Cancer Stem Cell Characteristics in liver Hepatocellular Carcinoma by WGCNA analysis of transcriptome stemness index. Cancer Med. 2020, 9, 4290–4298. [Google Scholar] [CrossRef]
- Uemura, M.; Fujimura, Y.; Ko, S.; Matsumoto, M.; Nakajima, Y.; Fukui, H. Pivotal Role of ADAMTS13 Function in liver diseases. Int. J. Hematol. 2010, 91, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Uemura, M.; Fujimura, Y.; Matsumoto, M.; Ishizashi, H.; Kato, S.; Matsuyama, T.; Isonishi, A.; Ishikawa, M.; Yagita, M.; Morioka, C.; et al. Comprehensive analysis of ADAMTS13 in patients with liver cirrhosis. Thromb. Haemost. 2008, 99, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Kavakli, K.; Canciani, M.T.; Mannucci, P.M. Plasma levels of the von willebrand factor-cleaving protease in physiological and pathological conditions in children. Pediatric Hematol. Oncol. 2002, 19, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Uemura, M.; Matsuyama, T.; Ishikawa, M.; Fujimoto, M.; Kojima, H.; Sakurai, S.; Ishii, S.; Toyohara, M.; Yamazaki, M.; Yoshiji, H.; et al. Decreased Activity of Plasma ADAMTS13 May Contribute to the Development of Liver Disturbance and Multiorgan Failure in Patients with Alcoholic Hepatitis. Alcohol. Clin. Exp. Res. 2005, 29, 264S–271S. [Google Scholar] [CrossRef] [PubMed]
- Lisman, T.; Bongers, T.N.; Adelmeijer, J.; Janssen, H.L.A.; de Maat, M.P.M.; de Groot, P.G.; Leebeek, F.W.G. elevated levels of von willebrand factor in cirrhosis support platelet adhesion despite reduced functional capacity. Hepatology 2006, 44, 53–61. [Google Scholar] [CrossRef]
- Ikeda, H.; Tateishi, R.; Enooku, K.; Yoshida, H.; Nakagawa, H.; Masuzaki, R.; Kondo, Y.; Goto, T.; Shiina, S.; Kume, Y.; et al. Prediction of Hepatocellular Carcinoma development by plasma ADAMTS13 in chronic Hepatitis B and C. Cancer Epidemiol. Biomark. Prev. 2011, 20, 2204–2211. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Takaya, H.; Namisaki, T.; Shimozato, N.; Kaji, K.; Kitade, M.; Moriya, K.; Sato, S.; Kawaratani, H.; Akahane, T.; Matsumoto, M.; et al. ADAMTS13 and von willebrand factor are useful biomarkers for sorafenib treatment efficiency in patients with Hepatocellular Carcinoma. World J. Gastrointest. Oncol. 2019, 11, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Takaya, H.; Namisaki, T.; Kitade, M.; Kaji, K.; Nakanishi, K.; Tsuji, Y.; Shimozato, N.; Moriya, K.; Seki, K.; Sawada, Y.; et al. VWF/ADAMTS13 ratio as a potential biomarker for early detection of Hepatocellular Carcinoma. BMC Gastroenterol. 2019, 19, 167. [Google Scholar] [CrossRef]
- Kume, Y.; Ikeda, H.; Inoue, M.; Tejima, K.; Tomiya, T.; Nishikawa, T.; Watanabe, N.; Ichikawa, T.; Kaneko, M.; Okubo, S.; et al. Hepatic stellate cell damage may lead to decreased plasma ADAMTS13 activity in rats. FEBS Lett. 2007, 581, 1631–1634. [Google Scholar] [CrossRef][Green Version]
- Uemura, M.; Tatsumi, K.; Matsumoto, M.; Fujimoto, M.; Matsuyama, T.; Ishikawa, M.; Iwamoto, T.-A.; Mori, T.; Wanaka, A.; Fukui, H.; et al. Localization of ADAMTS13 to the stellate cells of human liver. Blood 2005, 106, 922–924. [Google Scholar] [CrossRef] [PubMed]
- Niiya, M.; Uemura, M.; Zheng, X.W.; Pollak, E.S.; Dockal, M.; Scheiflinger, F.; Wells, R.G.; Zheng, X.L. Increased ADAMTS-13 proteolytic activity in rat hepatic stellate cells upon activation in vitro and in vivo. J. Thromb. Haemost. 2006, 4, 1063–1070. [Google Scholar] [CrossRef]
- Shomron, N.; Hamasaki-Katagiri, N.; Hunt, R.; Hershko, K.; Pommier, E.; Geetha, S.; Blaisdell, A.; Dobkin, A.; Marple, A.; Roma, I.; et al. A splice variant of ADAMTS13 is expressed in human hepatic stellate cells and cancerous tissues. Thromb. Haemost. 2010, 104, 531–535. [Google Scholar] [CrossRef] [PubMed]
- De Tan, I.A.; Ricciardelli, C.; Russell, D.L. The Metalloproteinase ADAMTS1: A Comprehensive review of its role in tumorigenic and Metastatic pathways. Int. J. Cancer 2013, 133, 2263–2276. [Google Scholar] [CrossRef] [PubMed]
- Sheu, M.-J.; Hsieh, M.-J.; Chou, Y.-E.; Wang, P.-H.; Yeh, C.-B.; Yang, S.-F.; Lee, H.-L.; Liu, Y.-F. Effects of ADAMTS14 genetic polymorphism and cigarette smoking on the clinicopathologic development of Hepatocellular Carcinoma. PLoS ONE 2017, 12, e0172506. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xiong, Y.; Zhong, Z.; Zhang, S.; Peng, Y.; Wang, L.; Dai, N.; Li, M.; Ren, T.; Gan, L.; et al. association between a variant in ADAMTS5 and the susceptibility to hepatocellular carcinoma in a chinese han population. Cell Biochem. Biophys. 2015, 72, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, R.; Jing, R.; Zuo, B.; Zheng, Q. Genome-Wide CRISPR Knockout Screens Identify ADAMTSL3 and PTEN Genes as suppressors of HCC proliferation and metastasis, respectively. J. Cancer Res. Clin. Oncol. 2020, 146, 1509–1521. [Google Scholar] [CrossRef]
- Arechederra, M.; Bazai, S.K.; Abdouni, A.; Sequera, C.; Mead, T.J.; Richelme, S.; Daian, F.; Audebert, S.; Dono, R.; Lozano, A.; et al. ADAMTSL5 is an epigenetically activated gene underlying tumorigenesis and drug resistance in Hepatocellular Carcinoma. J. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.-H.; Hurskainen, T.; Mielke, K.; Aung, P.P.; Casey, G.; Autio-Harmainen, H.; Apte, S.S. ADAMTSL3/Punctin-2, a Gene frequently mutated in colorectal tumors, is widely expressed in normal and malignant epithelial cells, vascular endothelial cells and other cell types, and its MRNA is reduced in colon cancer. Int. J. Cancer 2007, 121, 1710–1716. [Google Scholar] [CrossRef]
- Nonomura, Y.; Otsuka, A.; Ohtsuka, M.; Yamamoto, T.; Dummer, R.; Kabashima, K. ADAMTSL5 is upregulated in melanoma tissues in patients with idiopathic psoriasis vulgaris induced by Nivolumab. J. Eur. Acad. Dermatol. Venereol. 2017, 31, e100–e101. [Google Scholar] [CrossRef]
- Abdullah, M.; Choo, C.W.; Alias, H.; Abdul Rahman, E.J.; Mohd Ibrahim, H.; Jamal, R.; Hussin, N.H. ADAMTSL5 and CDH11: Putative epigenetic markers for therapeutic resistance in acute Lymphoblastic Leukemia. Hematology 2017, 22, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Kadalayil, L.; Khan, S.; Nevanlinna, H.; Fasching, P.A.; Couch, F.J.; Hopper, J.L.; Liu, J.; Maishman, T.; Durcan, L.; Gerty, S.; et al. Germline variation in ADAMTSL1 is associated with prognosis following breast cancer treatment in young women. Nat. Commun. 2017, 8, 1632. [Google Scholar] [CrossRef]
- Campbell, V.T.; Nadesan, P.; Ali, S.A.; Wang, C.Y.Y.; Whetstone, H.; Poon, R.; Wei, Q.; Keilty, J.; Proctor, J.; Wang, L.W.; et al. Hedgehog pathway inhibition in chondrosarcoma using the smoothened inhibitor IPI-926 directly inhibits sarcoma cell growth. Mol. Cancer Ther. 2014, 13, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.; Zhang, K.; Wang, K.; Li, G.; Huang, R.; Zhao, Z.; Liu, Y.; Chen, J. A Novel gene signature based on five glioblastoma Stem-like cell relevant genes predicts the survival of primary glioblastoma. J. Cancer Res. Clin. Oncol. 2018, 144, 439–447. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Comprehensive and integrative genomic characterization of Hepatocellular Carcinoma. Cell 2017, 169, 1327–1341. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Vanderkerken, M.; Hammad, H. The emerging role of ADAM metalloproteinases in immunity. Nat. Rev. Immunol. 2018, 18, 745–758. [Google Scholar] [CrossRef]
- Ringelhan, M.; Pfister, D.; O’Connor, T.; Pikarsky, E.; Heikenwalder, M. The Immunology of Hepatocellular Carcinoma. Nat. Immunol. 2018, 19, 222–232. [Google Scholar] [CrossRef]
- Yang, Y.M.; Kim, S.Y.; Seki, E. Inflammation and liver cancer: Molecular mechanisms and therapeutic targets. Semin. Liver Dis. 2019, 39, 26–42. [Google Scholar] [CrossRef]
- Zelová, H.; Hošek, J. TNF-α Signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- McMahan, R.S.; Riehle, K.J.; Fausto, N.; Campbell, J.S. A Disintegrin and Metalloproteinase 17 Regulates TNF and TNFR1 levels in inflammation and liver regeneration in mice. Am. J. Physiol. Gastrointest Liver Physiol. 2013, 305, G25–G34. [Google Scholar] [CrossRef] [PubMed]
- Maras, J.S.; Das, S.; Sharma, S.; Sukriti, S.; Kumar, J.; Vyas, A.K.; Kumar, D.; Bhat, A.; Yadav, G.; Choudhary, M.C.; et al. Iron-Overload triggers ADAM-17 mediated inflammation in severe alcoholic hepatitis. Sci. Rep. 2018, 8, 10264. [Google Scholar] [CrossRef]
- Argast, G.M.; Campbell, J.S.; Brooling, J.T.; Fausto, N. Epidermal growth factor receptor transactivation mediates tumor necrosis factor-induced hepatocyte replication. J. Biol. Chem. 2004, 279, 34530–34536. [Google Scholar] [CrossRef]
- Berasain, C.; Nicou, A.; Garcia-Irigoyen, O.; Latasa, M.U.; Urtasun, R.; Elizalde, M.; Salis, F.; Perugorría, M.J.; Prieto, J.; Recio, J.A.; et al. Epidermal growth factor receptor signaling in Hepatocellular Carcinoma: Inflammatory activation and a new intracellular regulatory mechanism. Dig. Dis. 2012, 30, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Lokau, J.; Schoeder, V.; Haybaeck, J.; Garbers, C. Jak-Stat signaling induced by Interleukin-6 family cytokines in Hepatocellular Carcinoma. Cancers 2019, 11, 1704. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Arras, D.; Rose-John, S. Regulation of fibrotic processes in the liver by ADAM proteases. Cells 2019, 8, 1226. [Google Scholar] [CrossRef]
- Giannitrapani, L.; Cervello, M.; Soresi, M.; Notarbartolo, M.; La Rosa, M.; Virruso, L.; D’Alessandro, N.; Montalto, G. Circulating IL-6 and SIL-6R in patients with Hepatocellular Carcinoma. Ann. N. Y. Acad. Sci. 2002, 963, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Soresi, M.; Giannitrapani, L.; D’Antona, F.; Florena, A.-M.; La Spada, E.; Terranova, A.; Cervello, M.; D’Alessandro, N.; Montalto, G. Interleukin-6 and its soluble receptor in patients with liver cirrhosis and Hepatocellular Carcinoma. World J. Gastroenterol. 2006, 12, 2563–2568. [Google Scholar] [CrossRef]
- Bergmann, J.; Müller, M.; Baumann, N.; Reichert, M.; Heneweer, C.; Bolik, J.; Lücke, K.; Gruber, S.; Carambia, A.; Boretius, S.; et al. IL-6 Trans-signaling is essential for the development of hepatocellular carcinoma in mice. Hepatology 2017, 65, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Dyczynska, E.; Sun, D.; Yi, H.; Sehara-Fujisawa, A.; Blobel, C.P.; Zolkiewska, A. Proteolytic processing of Delta-like 1 by ADAM proteases. J. Biol. Chem. 2007, 282, 436–444. [Google Scholar] [CrossRef]
- Huang, Q.; Li, J.; Zheng, J.; Wei, A. The carcinogenic role of the notch signaling pathway in the development of Hepatocellular Carcinoma. J. Cancer 2019, 10, 1570–1579. [Google Scholar] [CrossRef]
- Lu, J.; Xia, Y.; Chen, K.; Zheng, Y.; Wang, J.; Lu, W.; Yin, Q.; Wang, F.; Zhou, Y.; Guo, C. Oncogenic role of the notch pathway in primary liver cancer. Oncol. Lett. 2016, 12, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, S.; Erhardt, A.; Claass, B.; Huber, S.; Adler, G.; Jacobs, T.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S.; Karimi, K.; et al. Hepatocytes contribute to immune regulation in the liver by activation of the notch signaling pathway in T cells. J. Immunol. 2013, 191, 5574–5582. [Google Scholar] [CrossRef]
- Wang, R.; Li, Y.; Tsung, A.; Huang, H.; Du, Q.; Yang, M.; Deng, M.; Xiong, S.; Wang, X.; Zhang, L.; et al. INOS promotes CD24+CD133+ liver cancer stem cell phenotype through a TACE/ADAM17-Dependent notch signaling pathway. Proc. Natl. Acad. Sci. USA 2018, 115, E10127–E10136. [Google Scholar] [CrossRef]
- Chalupský, K.; Kanchev, I.; Žbodáková, O.; Buryová, H.; Jiroušková, M.; Kořínek, V.; Gregor, M.; Sedláček, R. ADAM10/17-dependent release of soluble c-Met correlates with hepatocellular damage. Folia Biol. 2013, 59, 76–86. [Google Scholar]
- Barisione, G.; Fabbi, M.; Gino, A.; Queirolo, P.; Orgiano, L.; Spano, L.; Picasso, V.; Pfeffer, U.; Mosci, C.; Jager, M.J.; et al. Potential Role of Soluble C-Met as a new candidate biomarker of metastatic uveal melanoma. JAMA Ophthalmol. 2015, 133, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-F.; Li, A.-N.; Yang, J.-J.; Chen, Z.-H.; Xie, Z.; Zhang, X.-C.; Su, J.; Lou, N.-N.; Yan, H.-H.; Han, J.-F.; et al. Soluble C-Met levels correlated with tissue c-Met protein expression in patients with advanced non-small-cell lung cancer. Clin. Lung Cancer 2017, 18, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J.; Yang, J.H.; Kim, J.; Ma, S.H.; Cho, L.Y.; Ko, K.-P.; Shin, A.; Choi, B.Y.; Kim, H.J.; Han, D.S.; et al. Soluble C-Met protein as a susceptible biomarker for gastric cancer risk: A nested case-control study within the korean multicenter cancer cohort. Int. J. Cancer 2013, 132, 2148–2156. [Google Scholar] [CrossRef]
- Moosavi, F.; Giovannetti, E.; Saso, L.; Firuzi, O. HGF/MET pathway aberrations as diagnostic, prognostic, and predictive biomarkers in human cancers. Crit. Rev. Clin. Lab. Sci. 2019, 56, 533–566. [Google Scholar] [CrossRef]
- Xing, S.; Ferrari de Andrade, L. NKG2D and MICA/B shedding: A “tag Game” between NK cells and Malignant cells. Clin. Transl. Immunol. 2020, 9, e1230. [Google Scholar] [CrossRef]
- Arai, J.; Goto, K.; Tanoue, Y.; Ito, S.; Muroyama, R.; Matsubara, Y.; Nakagawa, R.; Kaise, Y.; Lim, L.A.; Yoshida, H.; et al. Enzymatic inhibition of MICA sheddase ADAM17 by lomofungin in Hepatocellular Carcinoma Cells. Int. J. Cancer 2018, 143, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Kohga, K.; Takehara, T.; Tatsumi, T.; Miyagi, T.; Ishida, H.; Ohkawa, K.; Kanto, T.; Hiramatsu, N.; Hayashi, N. Anticancer chemotherapy inhibits MHC Class I-Related chain a ectodomain shedding by downregulating ADAM10 expression in Hepatocellular Carcinoma. Cancer Res. 2009, 69, 8050–8057. [Google Scholar] [CrossRef]
- Kohga, K.; Takehara, T.; Tatsumi, T.; Ishida, H.; Miyagi, T.; Hosui, A.; Hayashi, N. Sorafenib Inhibits the Shedding of Major Histocompatibility Complex Class I-Related chain a on hepatocellular carcinoma cells by down-regulating a disintegrin and metalloproteinase. Hepatology 2010, 51, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Arai, J.; Goto, K.; Stephanou, A.; Tanoue, Y.; Ito, S.; Muroyama, R.; Matsubara, Y.; Nakagawa, R.; Morimoto, S.; Kaise, Y.; et al. Predominance of regorafenib over sorafenib: Restoration of membrane-bound MICA in hepatocellular carcinoma cells. J. Gastroenterol. Hepatol. 2018, 33, 1075–1081. [Google Scholar] [CrossRef]
- Gérard, C.; Hubeau, C.; Carnet, O.; Bellefroid, M.; Sounni, N.E.; Blacher, S.; Bendavid, G.; Moser, M.; Fässler, R.; Noel, A.; et al. Microenvironment-Derived ADAM28 prevents cancer dissemination. Oncotarget 2018, 9, 37185–37199. [Google Scholar] [CrossRef]
- Zha, Z.; Jia, F.; Hu, P.; Mai, E.; Lei, T. MicroRNA-574-3p inhibits the malignant behavior of liver cancer cells by targeting ADAM. Oncol. Lett. 2020, 20, 3015–3023. [Google Scholar] [CrossRef] [PubMed]
- Hubeau, C.; Rocks, N.; Cataldo, D. ADAM28: Another ambivalent protease in cancer. Cancer Lett. 2020, 494, 18–26. [Google Scholar] [CrossRef]
- Higashiyama, S.; Nanba, D. ADAM-Mediated Ectodomain shedding of HB-EGF in receptor Cross-Talk. Biochim. Biophys. Acta 2005, 1751, 110–117. [Google Scholar] [CrossRef]
- Sanderson, M.P.; Dempsey, P.J.; Dunbar, A.J. Control of ErbB signaling through metalloprotease mediated ectodomain shedding of EGF-like factors. Growth Factors 2006, 24, 121–136. [Google Scholar] [CrossRef]
- Itabashi, H.; Maesawa, C.; Oikawa, H.; Kotani, K.; Sakurai, E.; Kato, K.; Komatsu, H.; Nitta, H.; Kawamura, H.; Wakabayashi, G.; et al. Angiotensin II and epidermal growth factor receptor cross-talk mediated by a disintegrin and metalloprotease accelerates tumor cell proliferation of hepatocellular carcinoma cell lines. Hepatol. Res. 2008, 38, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-J.; Feng, C.-W.; Li, M. ADAM17 mediates hypoxia-induced drug resistance in hepatocellular carcinoma cells through activation of EGFR/PI3K/Akt pathway. Mol. Cell Biochem. 2013, 380, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, G.; Koudelkova, P.; Dituri, F.; Mikulits, W. Role of epithelial to mesenchymal transition in hepatocellular carcinoma. J. Hepatol. 2016, 65, 798–808. [Google Scholar] [CrossRef]
- Pinzani, M. Epithelial-Mesenchymal transition in chronic liver disease: Fibrogenesis or escape from death? J. Hepatol. 2011, 55, 459–465. [Google Scholar] [CrossRef]
- Dong, Y.; Wu, Z.; He, M.; Chen, Y.; Chen, Y.; Shen, X.; Zhao, X.; Zhang, L.; Yuan, B.; Zeng, Z. ADAM9 mediates the Interleukin-6-Induced epithelial-mesenchymal transition and metastasis through ROS production in hepatoma cells. Cancer Lett. 2018, 421, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-Y.; Chu, H.-X.; Tan, Y.-X.; Qin, X.-C.; Liu, M.-Y.; Li, J.-D.; Ren, T.-S.; Zhang, Y.-S.; Zhao, Q.-C. Novel ADAM-17 inhibitor ZLDI-8 inhibits the metastasis of hepatocellular carcinoma by reversing epithelial-mesenchymal transition in vitro and in vivo. Life Sci. 2020, 244, 117343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, D.; Jiang, Q.; Cao, S.; Sun, H.; Chai, Y.; Li, X.; Ren, T.; Yang, R.; Feng, F.; et al. Novel ADAM-17 inhibitor ZLDI-8 enhances the in vitro and in vivo chemotherapeutic effects of sorafenib on hepatocellular carcinoma cells. Cell Death Dis. 2018, 9, 743. [Google Scholar] [CrossRef]
- Hong, S.W.; Hur, W.; Choi, J.E.; Kim, J.-H.; Hwang, D.; Yoon, S.K. Role of ADAM17 in invasion and migration of CD133-expressing liver cancer stem cells after irradiation. Oncotarget 2016, 7, 23482–23497. [Google Scholar] [CrossRef]
- Li, Y.; Ren, Z.; Wang, Y.; Dang, Y.-Z.; Meng, B.-X.; Wang, G.-D.; Zhang, J.; Wu, J.; Wen, N. ADAM17 promotes cell migration and invasion through the integrin β1 pathway in hepatocellular carcinoma. Exp. Cell Res. 2018, 370, 373–382. [Google Scholar] [CrossRef]
- Saha, S.K.; Choi, H.Y.; Yang, G.-M.; Biswas, P.K.; Kim, K.; Kang, G.-H.; Gil, M.; Cho, S.-G. GPR50 promotes hepatocellular carcinoma progression via the notch signaling pathway through direct interaction with ADAM. Mol. Ther. Oncolytics 2020, 17, 332–349. [Google Scholar] [CrossRef]
- Wu, B.; Cui, J.; Yang, X.-M.; Liu, Z.-Y.; Song, F.; Li, L.; Jiang, J.-L.; Chen, Z.-N. Cytoplasmic fragment of CD147 generated by regulated intramembrane proteolysis contributes to HCC by promoting autophagy. Cell Death Dis. 2017, 8, e2925. [Google Scholar] [CrossRef]
- Fabregat, I.; Moreno-Càceres, J.; Sánchez, A.; Dooley, S.; Dewidar, B.; Giannelli, G.; Ten Dijke, P. IT-LIVER Consortium TGF-β signalling and liver disease. FEBS J. 2016, 283, 2219–2232. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Murata, M.; Yamaguchi, T.; Matsuzaki, K. TGF-β/Smad signaling during hepatic fibro-carcinogenesis (Review). Int. J. Oncol. 2014, 45, 1363–1371. [Google Scholar] [CrossRef]
- Le Pabic, H.; L’Helgoualc’h, A.; Coutant, A.; Wewer, U.M.; Baffet, G.; Clément, B.; Théret, N. Involvement of the serine/threonine P70S6 kinase in TGF-Beta1-induced ADAM12 expression in cultured human hepatic stellate cells. J. Hepatol. 2005, 43, 1038–1044. [Google Scholar] [CrossRef]
- Gruel, J.; Leborgne, M.; LeMeur, N.; Théret, N. In silico investigation of ADAM12 effect on TGF-Beta Receptors trafficking. BMC Res. Notes 2009, 2, 193. [Google Scholar] [CrossRef]
- Ruff, M.; Leyme, A.; Le Cann, F.; Bonnier, D.; Le Seyec, J.; Chesnel, F.; Fattet, L.; Rimokh, R.; Baffet, G.; Théret, N. The disintegrin and metalloprotease ADAM12 is associated with TGF-β-Induced epithelial to mesenchymal transition. PLoS ONE 2015, 10, e0139179. [Google Scholar] [CrossRef]
- Dekky, B.; Ruff, M.; Bonnier, D.; Legagneux, V.; Théret, N. Proteomic screening identifies the zonula occludens protein ZO-1 as a new partner for ADAM12 in invadopodia-like structures. Oncotarget 2018, 9, 21366–21382. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Eckert, M.A.; Santiago-Medina, M.; Lwin, T.M.; Kim, J.; Courtneidge, S.A.; Yang, J. ADAM12 Induction by Twist1 promotes tumor invasion and metastasis via regulation of invadopodia and focal adhesions. J. Cell Sci. 2017, 130, 2036–2048. [Google Scholar] [CrossRef] [PubMed]
- Laurent, M.-A.; Bonnier, D.; Théret, N.; Tufféry, P.; Moroy, G. In Silico Characterization of the interaction between LSKL peptide, a LAP-TGF-Beta derived peptide, and ADAMTS. Comput. Biol. Chem. 2016, 61, 155–161. [Google Scholar] [CrossRef]
- Amann, T.; Bataille, F.; Spruss, T.; Mühlbauer, M.; Gäbele, E.; Schölmerich, J.; Kiefer, P.; Bosserhoff, A.-K.; Hellerbrand, C. Activated hepatic stellate cells promote tumorigenicity of hepatocellular carcinoma. Cancer Sci. 2009, 100, 646–653. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
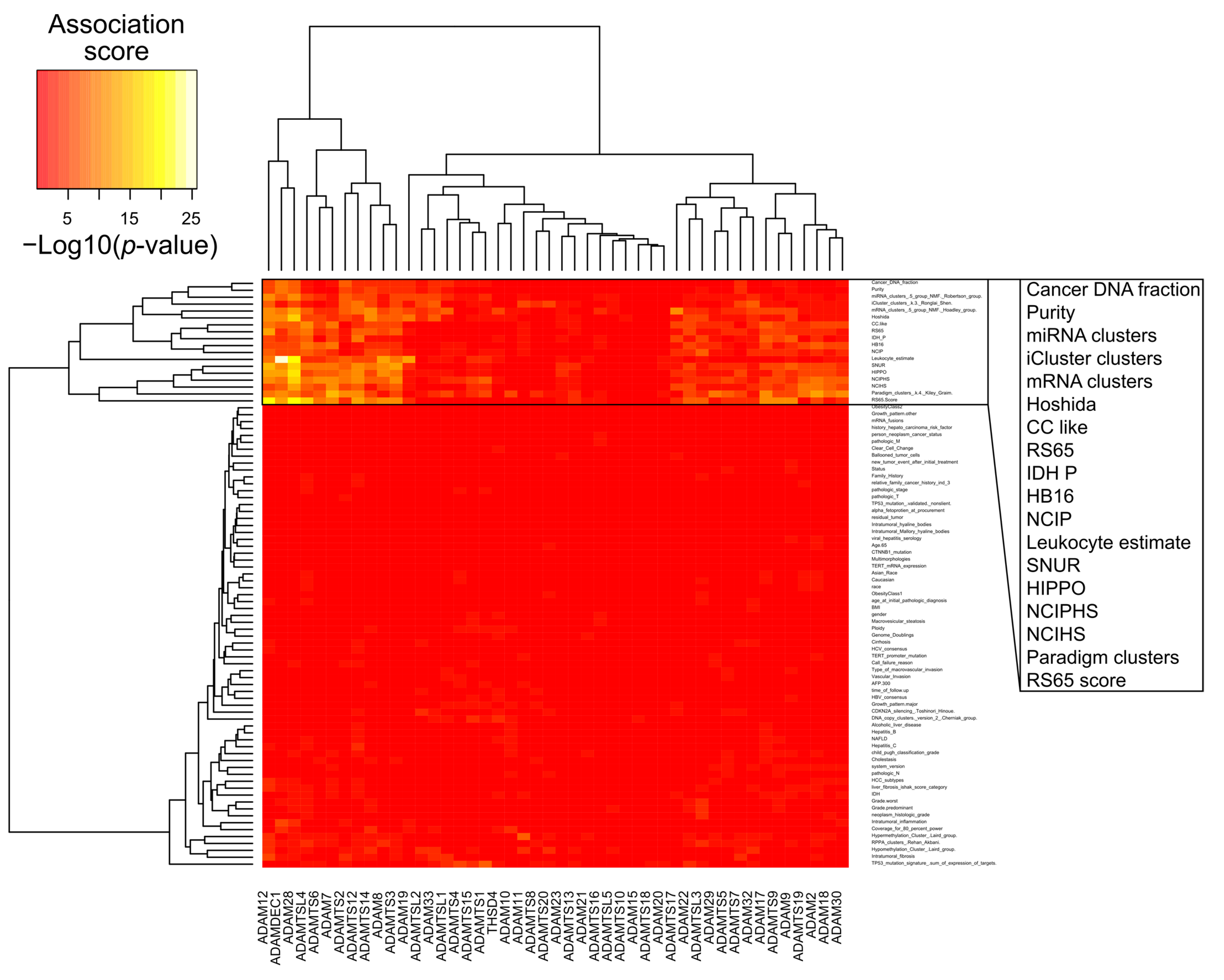{kind=link}
| T versus Adjacent NT Tissue (n = 48) | Kaplan–Meier Survival Analysis (n = 364) | |||
|---|---|---|---|---|
| Name | p-Value | Worse Prognosis (Expression Low–High) | p-Value | |
| ADAM2 | nd | - | Low | 2.00 × 10−9 |
| ADAM7 | nd | - | Low | 1.40 × 10−9 |
| ADAM8 | ns | 7.10 × 10−1 | Low | 1.20 × 10−2 |
| ADAM9 | increase | 5.60 × 10−13 | High | 3.50 × 10−4 |
| ADAM10 | increase | 7.20 × 10−6 | ns | 6.30 × 10−2 |
| ADAM11 | increase | 1.50 × 10−5 | Low | 5.60 × 10−3 |
| ADAM12 | increase | 2.20 × 10−5 | High | 4.00 × 10−5 |
| ADAM15 | increase | 8.20 × 10−19 | High | 7.40 × 10−3 |
| ADAM17 | increase | 2.10 × 10−6 | High | 3.70 × 10−2 |
| ADAM18 | nd | - | Low | 8.30 × 10−9 |
| ADAM19 | ns | 1.80 × 10−1 | High | 2.20 × 10−2 |
| ADAM20 | ns | 2.00 × 10−1 | Low | 1.90 × 10−3 |
| ADAM21 | increase | 1.50 × 10−9 | ns | 1.80 × 10−1 |
| ADAM22 | increase | 4.80 × 10−4 | ns | 6.90 × 10−2 |
| ADAM23 | increase | 1.30 × 10−6 | ns | 9.30 × 10−2 |
| ADAM28 | ns | 8.90 × 10−1 | ns | 1.70 × 10−1 |
| ADAM29 | nd | - | Low | 2.30 × 10−9 |
| ADAM30 | nd | - | Low | 4.00 × 10−8 |
| ADAM32 | nd | - | Low | 1.30 × 10−2 |
| ADAM33 | ns | 5.90 × 10−1 | Low | 1.00 × 10−3 |
| ADAMDEC1 | increase | 3.30 × 10−3 | ns | 2.70 × 10−1 |
| ADAMTS1 | decrease | 5.90 × 10−5 | ns | 4.10 × 10−2 |
| ADAMTS2 | decrease | 1.00 × 10−4 | ns | 4.80 × 10−1 |
| ADAMTS3 | ns | 8.00 × 10−1 | high | 1.10 × 10−2 |
| ADAMTS4 | ns | 7.80 × 10−2 | ns | 4.80 × 10−2 |
| ADAMTS5 | increase | 2.40 × 10−4 | High | 1.00 × 10−5 |
| ADAMTS6 | increase | 5.00 × 10−03 | ns | 1.70 × 10−1 |
| ADAMTS7 | increase | 1.30 × 10−15 | High | 3.30 × 10−2 |
| ADAMTS8 | ns | 2.30 × 10−1 | Low | 7.90 × 10−4 |
| ADAMTS9 | increase | 2.60 × 10−10 | High | 3.20 × 10−2 |
| ADAMST10 | increase | 6.10 × 10−7 | Low | 1.90 × 10−3 |
| ADAMTS12 | ns | 2.60 × 10−2 | ns | 2.60 × 10−1 |
| ADAMTS13 | decrease | 3.80 × 10−21 | ns | 6.70 × 10−2 |
| ADAMTS14 | increase | 1.70 × 10−6 | High | 1.40 × 10−2 |
| ADAMTS15 | ns | 7.10 × 10−1 | ns | 6.50 × 10−2 |
| ADAMTS16 | increase | 8.40 × 10−3 | ns | 7.10 × 10−2 |
| ADAMTS17 | ns | 2.20 × 10−1 | Low | 3.90 × 10−2 |
| ADAMTS18 | increase | 9.30 × 10−6 | Low | 2.40 × 10−2 |
| ADAMTS19 | ns | - | Low | 6.50 × 10−9 |
| ADAMTS20 | ns | - | Low | 5.30 × 10−6 |
| ADAMTSL1 | ns | 1.20 × 10−1 | Low | 1.90 × 10−3 |
| ADAMTSL2 | decrease | 9.70 × 10−6 | Low | 3.30 × 10−3 |
| ADAMTSL3 | decrease | 1.90 × 10−2 | Low | 1.10 × 10−2 |
| ADAMTSL4 | ns | 1.20 × 10−1 | Low | 2.30 × 10−2 |
| ADAMTSL5 | increase | 3.10 × 10−5 | ns | 6.80 × 10−2 |
| ADAMTSL6 (THSD4) | ns | 3.10 × 10−1 | Low | 1.40 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Théret, N.; Bouezzeddine, F.; Azar, F.; Diab-Assaf, M.; Legagneux, V. ADAM and ADAMTS Proteins, New Players in the Regulation of Hepatocellular Carcinoma Microenvironment. Cancers 2021, 13, 1563. https://doi.org/10.3390/cancers13071563
Théret N, Bouezzeddine F, Azar F, Diab-Assaf M, Legagneux V. ADAM and ADAMTS Proteins, New Players in the Regulation of Hepatocellular Carcinoma Microenvironment. Cancers. 2021; 13(7):1563. https://doi.org/10.3390/cancers13071563
Chicago/Turabian StyleThéret, Nathalie, Fidaa Bouezzeddine, Fida Azar, Mona Diab-Assaf, and Vincent Legagneux. 2021. "ADAM and ADAMTS Proteins, New Players in the Regulation of Hepatocellular Carcinoma Microenvironment" Cancers 13, no. 7: 1563. https://doi.org/10.3390/cancers13071563
APA StyleThéret, N., Bouezzeddine, F., Azar, F., Diab-Assaf, M., & Legagneux, V. (2021). ADAM and ADAMTS Proteins, New Players in the Regulation of Hepatocellular Carcinoma Microenvironment. Cancers, 13(7), 1563. https://doi.org/10.3390/cancers13071563