
Higher Integrin Alpha 3 Beta1 Expression in Papillary Thyroid Cancer Is Associated with Worst Outcome

 , , ,
, , ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Data Analysis
2.3. Cell Lines, Antibodies, Immunofluorescence and Flow Cytometric Analysis
2.4. Downregulation of α3 Expression by siRNA
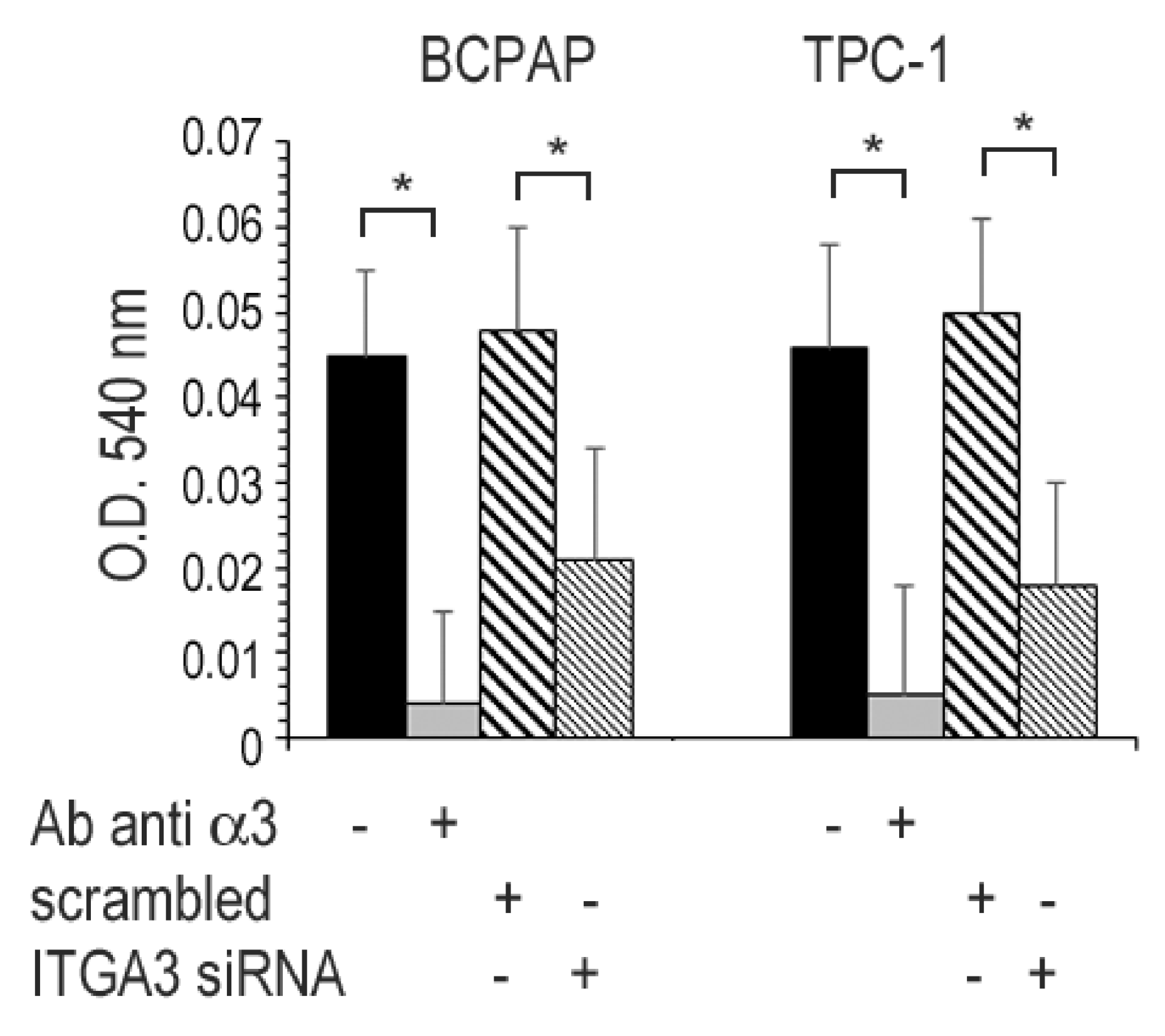
2.5. Cell Attachment Assay
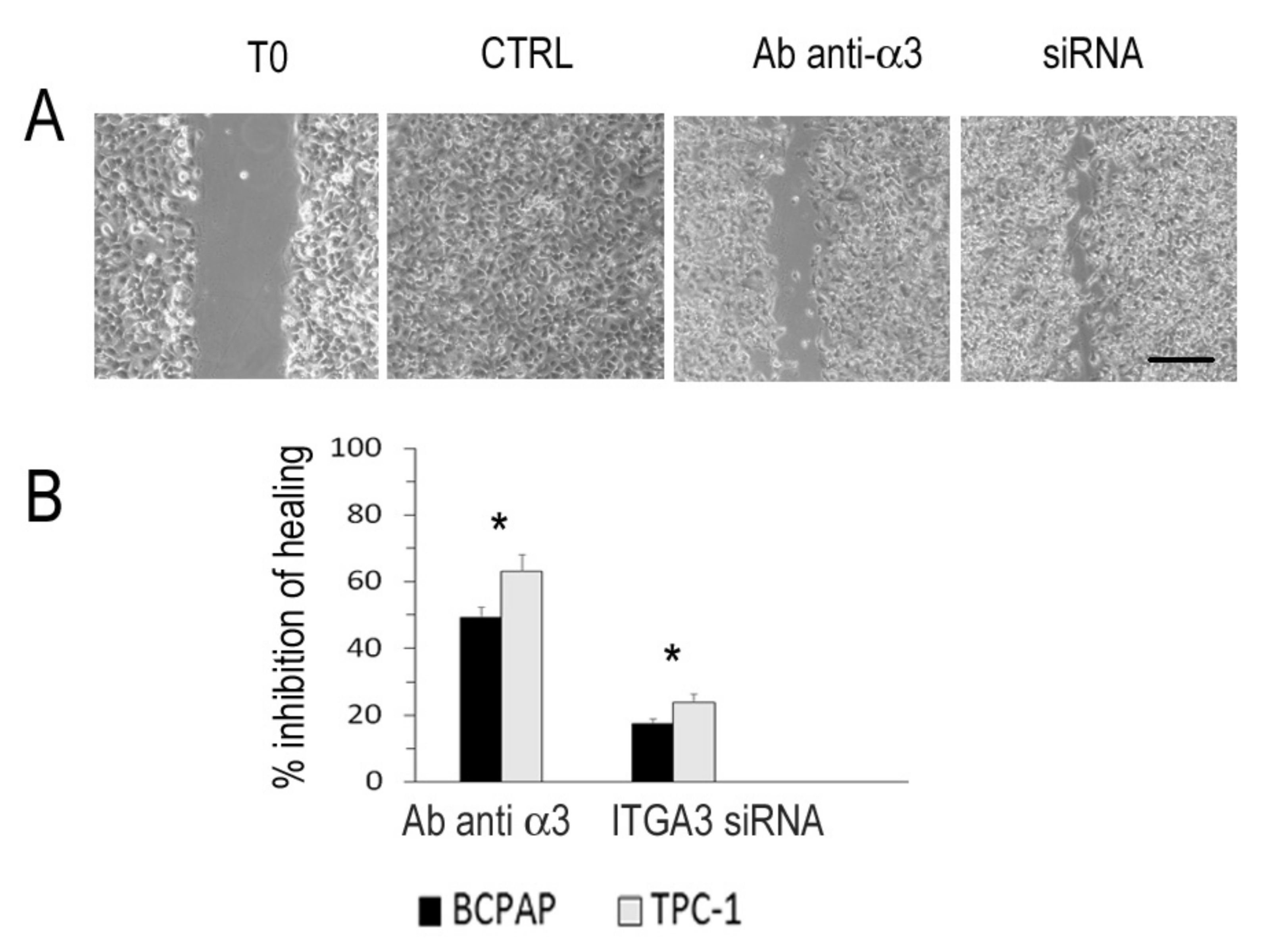
2.6. In Vitro Scratch Assay
2.7. Invasion Assay
2.8. Statistical Analysis
3. Results
3.1. Effect of Non-Tumoral Cells on Integrin Expression Assessment
3.2. Expression of Integrin Subunits mRNA in Normal Thyroid Tissue
3.3. Comparative Analysis of Integrin Subunits in PTC Variants
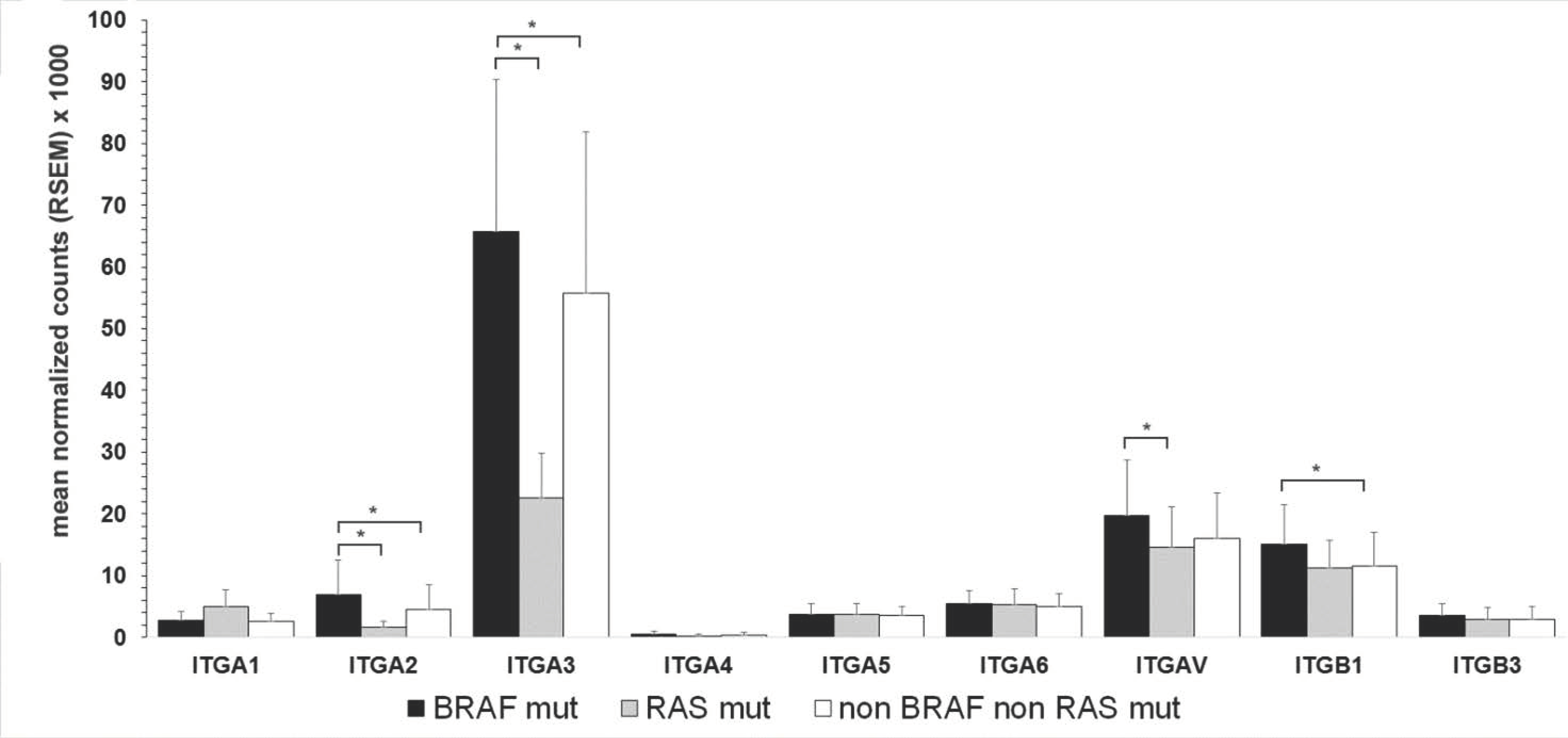
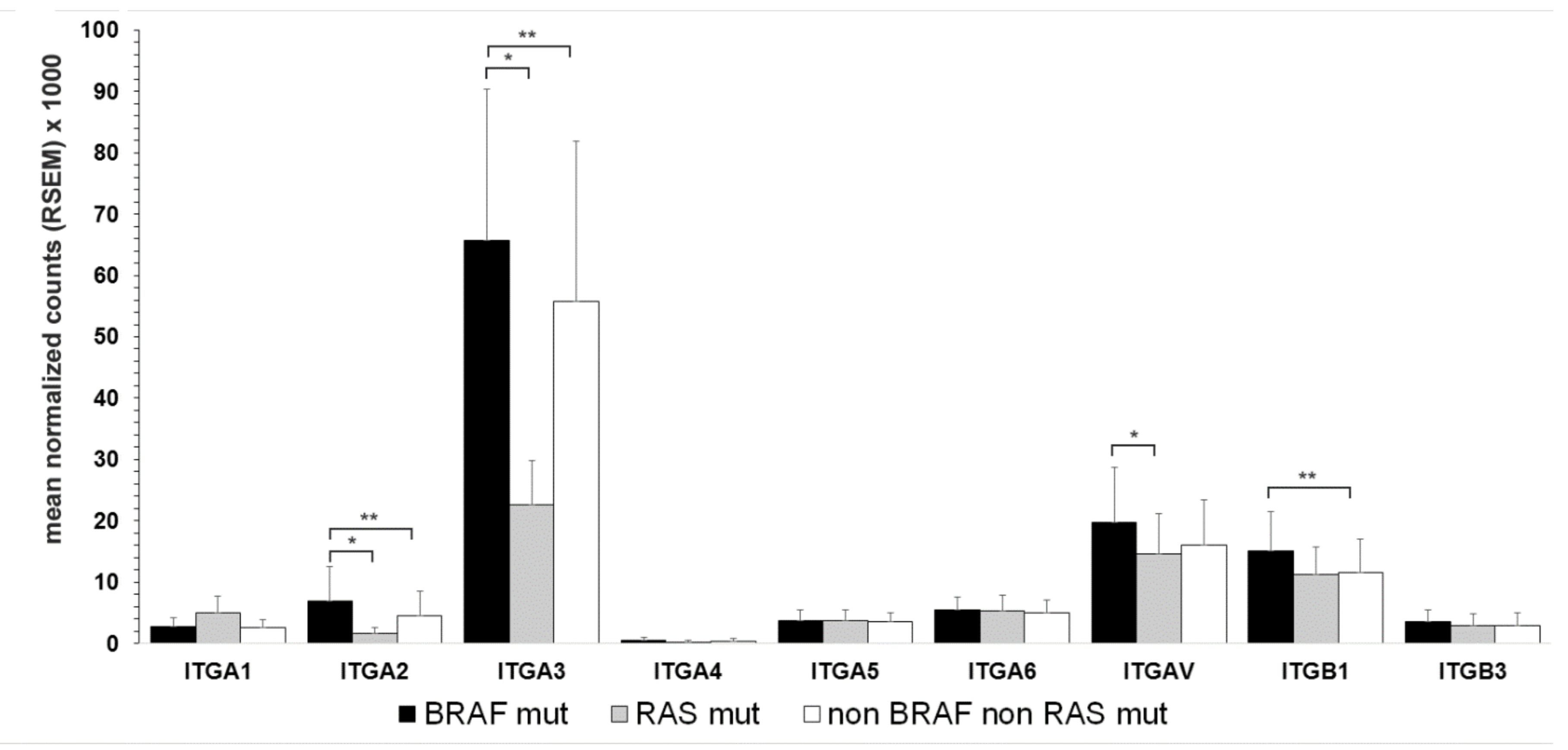
3.4. Association of Integrin Expression and Driver Gene Mutations
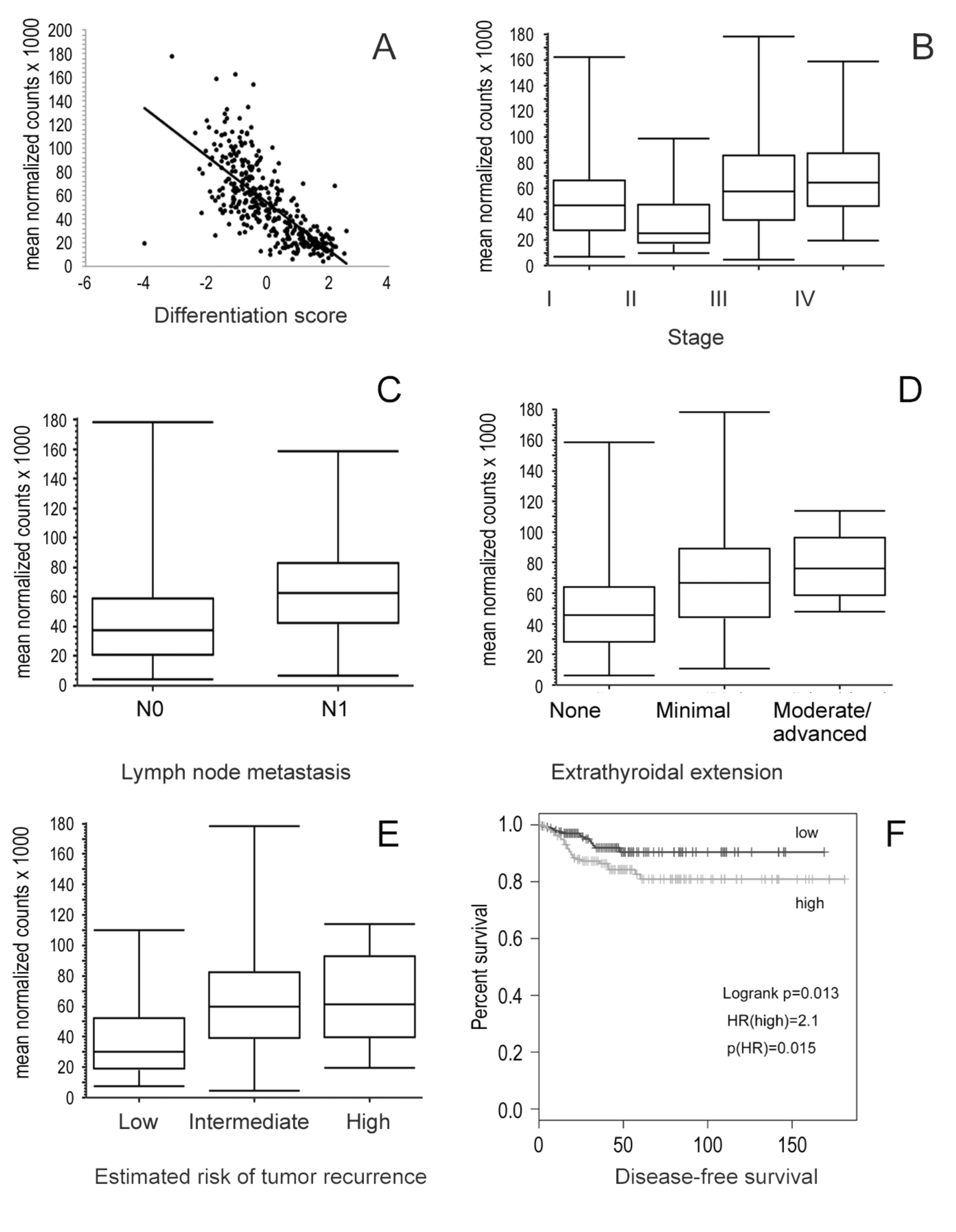
3.5. Association of Integrin Subunits Expression and Clinical Features
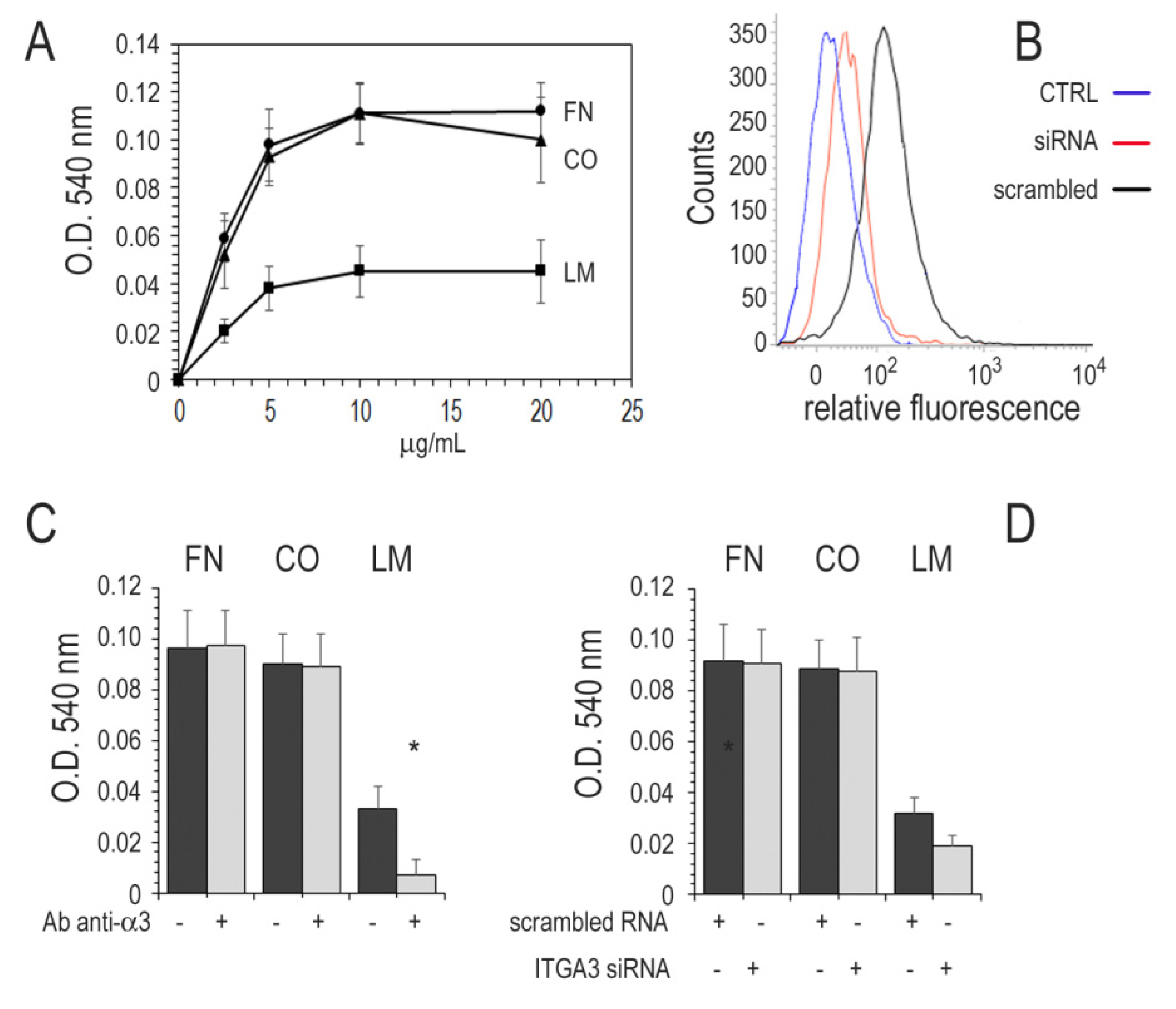
3.6. Adhesion of Cancer Cell Lines to ECM
3.7. Integrin α3β1 and Motility in Thyroid Cancer Cell Lines
3.8. Invasion Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamidi, H.; Ivaska, J. Every step of the way: Integrins in cancer progression and metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Geiger, B.; Bershadsky, A.; Pankov, R.; Yamada, K.M. Transmembrane crosstalk between the extracellular matrix—Cytoskeleton crosstalk. Nat. Rev. Mol. Cell Biol. 2001, 2, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma’ayan, A.; Iyengar, R.; Geiger, B. Functional atlas of the integrin adhesome. Nat. Cell Biol. 2007, 9, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef]
- Carlevato, M.T.; Trusolino, L.; Serini, G.; Valente, G.; Orlandi, F.; Angeli, A.; Cortesina, G.; Marchisio, P.C. Differential integrin expression in thyroid and laryngeal carcinomas. Anticancer Res. 1996, 16, 2379–2384. [Google Scholar]
- Hoffmann, S.; Maschuw, K.; Hassan, I.; Reckzeh, B.; Wunderlich, A.; Lingelbach, S.; Zielke, A. Differential pattern of integrin receptor expression in differentiated and anaplastic thyroid cancer cell lines. Thyroid 2005, 15, 1011–1020. [Google Scholar] [CrossRef]
- Vitale, M.; Bassi, V.; Fenzi, G.; Macchia, P.E.; Salzano, S.; Rossi, G. Integrin expression in thyroid cells from normal glands and nodular goiters. J. Clin. Endocrinol. Metab. 1993, 76, 1575–1579. [Google Scholar] [CrossRef]
- Vitale, M.; Di Matola, T.; Fenzi, G.; Illario, M.; Rossi, G. Fibronectin is required to prevent thyroid cell apoptosis through an integrin-mediated adhesion mechanism. J. Clin. Endocrinol. Metab. 1998, 83, 3673–3680. [Google Scholar] [CrossRef]
- Vitale, M.; Illario, M.; Di Matola, T.; Casamassima, A.; Fenzi, G.; Rossi, G. Integrin binding to immobilized collagen and fibronectin stimulates the proliferation of human thyroid cells in culture. Endocrinology 1997, 138, 1642–1648. [Google Scholar] [CrossRef]
- Vitale, M.; Casamassima, A.; Illario, M.; Bassi, V.; Fenzi, G.; Rossi, G. Cell-to-cell contact modulates the expression of the beta 1 family of integrins in primary cultures of thyroid cells. Exp. Cell Res. 1995, 220, 124–129. [Google Scholar] [CrossRef]
- Serini, G.; Trusolino, L.; Saggiorato, E.; Cremona, O.; De Rossi, M.; Angeli, A.; Orlandi, F.; Marchisio, P.C. Changes in integrin and E-cadherin expression in neoplastic versus normal thyroid tissue. J. Natl. Cancer Inst. 1996, 88, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, T.; Grimelius, L.; Wallin, G.; Rubin, K.; Westermark, K. Integrins in thyroid tissue: Upregulation of alpha2beta1 in anaplastic thyroid carcinoma. Eur. J. Endocrinol. 1998, 138, 104–112. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Celetti, A.; Garbi, C.; Consales, C.; Cerrato, A.; Greco, D.; Mele, E.; Nitsch, L.; Grieco, M. Analysis of cadherin/catenin complexes in transformed thyroid epithelial cells: Modulation by beta 1 integrin subunit. Eur. J. Cell Biol. 2000, 79, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Chernaya, G.; Mikhno, N.; Khabalova, T.; Svyatchenko, S.; Mostovich, L.; Shevchenko, S.; Gulyaeva, L. The expression profile of integrin receptors and osteopontin in thyroid malignancies varies depending on the tumor progression rate and presence of BRAF V600E mutation. Surg. Oncol. 2018, 27, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Di Matola, T.; Mueller, F.; Fenzi, G.; Rossi, G.; Bifulco, M.; Marzano, L.A.; Vitale, M. Serum withdrawal-induced apoptosis in thyroid cells is caused by loss of fibronectin-integrin interaction. J. Clin. Endocrinol. Metab. 2000, 85, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Illario, M.; Amideo, V.; Casamassima, A.; Andreucci, M.; di Matola, T.; Miele, C.; Rossi, G.; Fenzi, G.; Vitale, M. Integrin-dependent cell growth and survival are mediated by different signals in thyroid cells. J. Clin. Endocrinol. Metab. 2003, 88, 260–269. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Illario, M.; Cavallo, A.L.; Monaco, S.; Di Vito, E.; Mueller, F.; Marzano, L.A.; Troncone, G.; Fenzi, G.; Rossi, G.; Vitale, M. Fibronectin-Induced Proliferation in Thyroid Cells Is Mediated by {alpha}v{beta}3 Integrin through Ras/Raf-1/MEK/ERK and Calcium/CaMKII Signals. J. Clin. Endocrinol. Metab. 2005, 90, 2865–2873. [Google Scholar] [CrossRef]
- Kitajiri, S.; Hosaka, N.; Hiraumi, H.; Hirose, T.; Ikehara, S. Increased expression of integrin beta-4 in papillary thyroid carcinoma with gross lymph node metastasis. Pathol. Int. 2002, 52, 438–441. [Google Scholar] [CrossRef]
- Zhuang, Z.N.; Xu, Z.J.; Zhou, Q.; Xu, X.Z.; Tian, J.; Liu, Y.F.; Guo, S.; Wang, J.Y.; Xu, K.S. Clinical significance of integrin beta6 as a tumor recurrence factor in follicular thyroid carcinoma. Head Neck 2014, 37, 1439–1447. [Google Scholar] [CrossRef]
- Smit, J.W.; van der Pluijm, G.; Vloedgraven, H.J.; Lowik, C.W.; Goslings, B.M. Role of integrins in the attachment of metastatic follicular thyroid carcinoma cell lines to bone. Thyroid 1998, 8, 29–36. [Google Scholar] [CrossRef]
- Cockburn, J.G.; Richardson, D.S.; Gujral, T.S.; Mulligan, L.M. RET-mediated cell adhesion and migration require multiple integrin subunits. J. Clin. Endocrinol. Metab. 2010, 95, E342–E346. [Google Scholar] [CrossRef] [PubMed]
- Noh, T.W.; Soung, Y.H.; Kim, H.I.; Gil, H.J.; Kim, J.M.; Lee, E.J.; Chung, J. Effect of {beta}4 integrin knockdown by RNA interference in anaplastic thyroid carcinoma. Anticancer Res. 2010, 30, 4485–4492. [Google Scholar] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Salvati, A.; Gigantino, V.; Nassa, G.; Giurato, G.; Alexandrova, E.; Rizzo, F.; Tarallo, R.; Weisz, A. The Histone Methyltransferase DOT1L Is a Functional Component of Estrogen Receptor Alpha Signaling in Ovarian Cancer Cells. Cancers 2019, 11, 1720. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Benjamini, Y.; Cohen, R. Weighted false discovery rate controlling procedures for clinical trials. Biostatistics 2017, 18, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, N.; Akbani, R.; Aksoy, B.A.; Ally, A.; Arachchi, H.; Asa, S.L.; Auman, J.T.; Balasundaram, M.; Balu, S.; Baylin, S.B.; et al. Integrated genomic characterization of papillary thyroid carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef]
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M.; et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid 2015, 26, 1–133. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, M.; Virtanen, I. Expression of laminin in thyroid gland and thyroid tumors: An immunohistologic study. Int. J. Cancer 1984, 34, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Mojares, E.; Del Rio Hernandez, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef]
- Kendall, C.H.; Sanderson, P.R.; Cope, J.; Talbot, I.C. Follicular thyroid tumours: A study of laminin and type IV collagen in basement membrane and endothelium. J. Clin. Pathol. 1985, 38, 1100–1105. [Google Scholar] [CrossRef]
- Vitale, M.; Bassi, V.; Illario, M.; Fenzi, G.; Casamassima, A.; Rossi, G. Loss of polarity and de novo expression of the beta 1 family of integrins in thyroid tumors. Int. J. Cancer 1994, 59, 185–190. [Google Scholar] [CrossRef]
- Ibaragi, S.; Shimo, T.; Hassan, N.M.; Isowa, S.; Kurio, N.; Mandai, H.; Kodama, S.; Sasaki, A. Induction of MMP-13 expression in bone-metastasizing cancer cells by type I collagen through integrin alpha1beta1 and alpha2beta1-p38 MAPK signaling. Anticancer Res. 2011, 31, 1307–1313. [Google Scholar] [PubMed]
- Nones, K.; Waddell, N.; Song, S.; Patch, A.M.; Miller, D.; Johns, A.; Wu, J.; Kassahn, K.S.; Wood, D.; Bailey, P.; et al. Genome-wide DNA methylation patterns in pancreatic ductal adenocarcinoma reveal epigenetic deregulation of SLIT-ROBO, ITGA2 and MET signaling. Int. J. Cancer 2014, 135, 1110–1118. [Google Scholar] [CrossRef]
- Ding, W.; Fan, X.L.; Xu, X.; Huang, J.Z.; Xu, S.H.; Geng, Q.; Li, R.; Chen, D.; Yan, G.R. Epigenetic silencing of ITGA2 by MiR-373 promotes cell migration in breast cancer. PLoS ONE 2015, 10, e0135128. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Wu, H.Y.; Lin, Y.L.; Tzou, S.C.; Chuang, C.H.; Jian, T.Y.; Chen, P.R.; Chang, Y.C.; Lin, C.H.; Huang, T.H.; et al. Blockade of ITGA2 Induces Apoptosis and Inhibits Cell Migration in Gastric Cancer. Biol. Proced. Online 2018, 20. [Google Scholar] [CrossRef]
- Ren, D.; Zhao, J.; Sun, Y.; Li, D.; Meng, Z.; Wang, B.; Fan, P.; Liu, Z.; Jin, X.; Wu, H. Overexpressed ITGA2 promotes malignant tumor aggression by up-regulating PD-L1 expression through the activation of the STAT3 signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 485. [Google Scholar] [CrossRef]
- Qin, A.; Liu, Q.; Wang, J. Ropivacaine inhibits proliferation, invasion, migration and promotes apoptosis of papillary thyroid cancer cells via regulating ITGA2 expression. Drug Dev. Res. 2020, 81, 700–707. [Google Scholar] [CrossRef]
- Jain, M.; Zhang, L.; Boufraqech, M.; Liu-Chittenden, Y.; Bussey, K.; Demeure, M.J.; Wu, X.; Su, L.; Pacak, K.; Stratakis, C.A.; et al. ZNF367 inhibits cancer progression and is targeted by miR-195. PLoS ONE 2014, 9, e101423. [Google Scholar] [CrossRef]
- Liu, H.; Chen, X.; Lin, T.; Yan, J.; Jiang, S. MicroRNA-524-5p suppresses the progression of papillary thyroid carcinoma cells via targeting on FOXE1 and ITGA3 in cell autophagy and cycling pathways. J. Cell Physiol. 2019, 234, 18382–18391. [Google Scholar] [CrossRef]
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef]
- Brooks, P.C.; Silletti, S.; von Schalscha, T.L.; Friedlander, M.; Cheresh, D.A. Disruption of angiogenesis by PEX, a noncatalytic metalloproteinase fragment with integrin binding activity. Cell 1998, 92, 391–400. [Google Scholar] [CrossRef]
- Dumin, J.A.; Dickeson, S.K.; Stricker, T.P.; Bhattacharyya-Pakrasi, M.; Roby, J.D.; Santoro, S.A.; Parks, W.C. Pro-COase-1 (matrix metalloproteinase-1) binds the alpha(2)beta(1) integrin upon release from keratinocytes migrating on type I CO. J. Biol. Chem. 2001, 276, 29368–29374. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subunit | R | p |
|---|---|---|
| ITGA1 | 0.51 | <0.0001 |
| ITGA2 | −0.50 | <0.0001 |
| ITGA3 | −0.71 | <0.0001 |
| ITGA4 | −0.17 | 0.0043 |
| ITGAV | −0.25 | 0.0024 |
| ITGB1 | −0.24 | 0.0034 |
| STAGE | |||||
|---|---|---|---|---|---|
| Subunit | I | II | III | IV | p |
| ITGA1 | 3437 | 4551 | 2917 | 2688 | <0.0001 |
| ITGA2 | 5179 | 3198 | 5695 | 5133 | 0.0417 |
| ITGA3 | 50620 | 35950 | 60690 | 68806 | <0.0001 |
| Subunit | N0 | N1 | p |
|---|---|---|---|
| ITGA1 | 3637 | 2894 | 0.0001 |
| ITGA2 | 4051 | 6251 | <0.0000 |
| ITGA3 | 44,777 | 64,078 | <0.0000 |
| ITGAV | 15,126 | 19,600 | <0.0000 |
| ITGB1 | 11,763 | 14,709 | <0.0000 |
| Estrathyroidal Extension | ||||
|---|---|---|---|---|
| Subunit | None | Minimal | Moderate/ Advanced | p |
| ITGA1 | 3632 | 2770 | 2426 | <0.0001 |
| ITGA2 | 4284 | 7124 | 5963 | <0.0001 |
| ITGA3 | 46,027 | 69,104 | 77,692 | <0.0001 |
| ITGAV | 16,035 | 20,639 | 20,250 | <0.0001 |
| ITGB1 | 12,503 | 14,808 | 17,491 | <0.0001 |
| Risk | ||||
|---|---|---|---|---|
| Subunit | Low | Intermediete | High | p |
| ITGA1 | 4087 | 2929 | 2651 | <0.0001 |
| ITGA2 | 3321 | 6218 | 5497 | <0.0001 |
| ITGA3 | 37,455 | 62,487 | 65,943 | <0.0001 |
| ITGA4 | 342 | 446 | 269 | 0.02108 |
| ITGAV | 14,544 | 19,230 | 17,161 | <0.0001 |
| ITGB1 | 11,686 | 14,119 | 14,403 | 0.0003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mautone, L.; Ferravante, C.; Tortora, A.; Tarallo, R.; Giurato, G.; Weisz, A.; Vitale, M. Higher Integrin Alpha 3 Beta1 Expression in Papillary Thyroid Cancer Is Associated with Worst Outcome. Cancers 2021, 13, 2937. https://doi.org/10.3390/cancers13122937
Mautone L, Ferravante C, Tortora A, Tarallo R, Giurato G, Weisz A, Vitale M. Higher Integrin Alpha 3 Beta1 Expression in Papillary Thyroid Cancer Is Associated with Worst Outcome. Cancers. 2021; 13(12):2937. https://doi.org/10.3390/cancers13122937
Chicago/Turabian StyleMautone, Lorenza, Carlo Ferravante, Anna Tortora, Roberta Tarallo, Giorgio Giurato, Alessandro Weisz, and Mario Vitale. 2021. "Higher Integrin Alpha 3 Beta1 Expression in Papillary Thyroid Cancer Is Associated with Worst Outcome" Cancers 13, no. 12: 2937. https://doi.org/10.3390/cancers13122937
APA StyleMautone, L., Ferravante, C., Tortora, A., Tarallo, R., Giurato, G., Weisz, A., & Vitale, M. (2021). Higher Integrin Alpha 3 Beta1 Expression in Papillary Thyroid Cancer Is Associated with Worst Outcome. Cancers, 13(12), 2937. https://doi.org/10.3390/cancers13122937