
Rintatolimod Induces Antiviral Activities in Human Pancreatic Cancer Cells: Opening for an Anti-COVID-19 Opportunity in Cancer Patients?

 ,
,  , ,
, ,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Treatment
2.2. RNA Extraction and Gene Expression Profile
2.3. Protein Extraction and Proteomic Profile
2.4. Data Analysis
3. Results
3.1. Rintatolimod Stimulated the Interferon Signaling Pathway in HPAC Cells
3.2. Rintatolimod Produced Immunomodulatory Activity in HPAC Cells
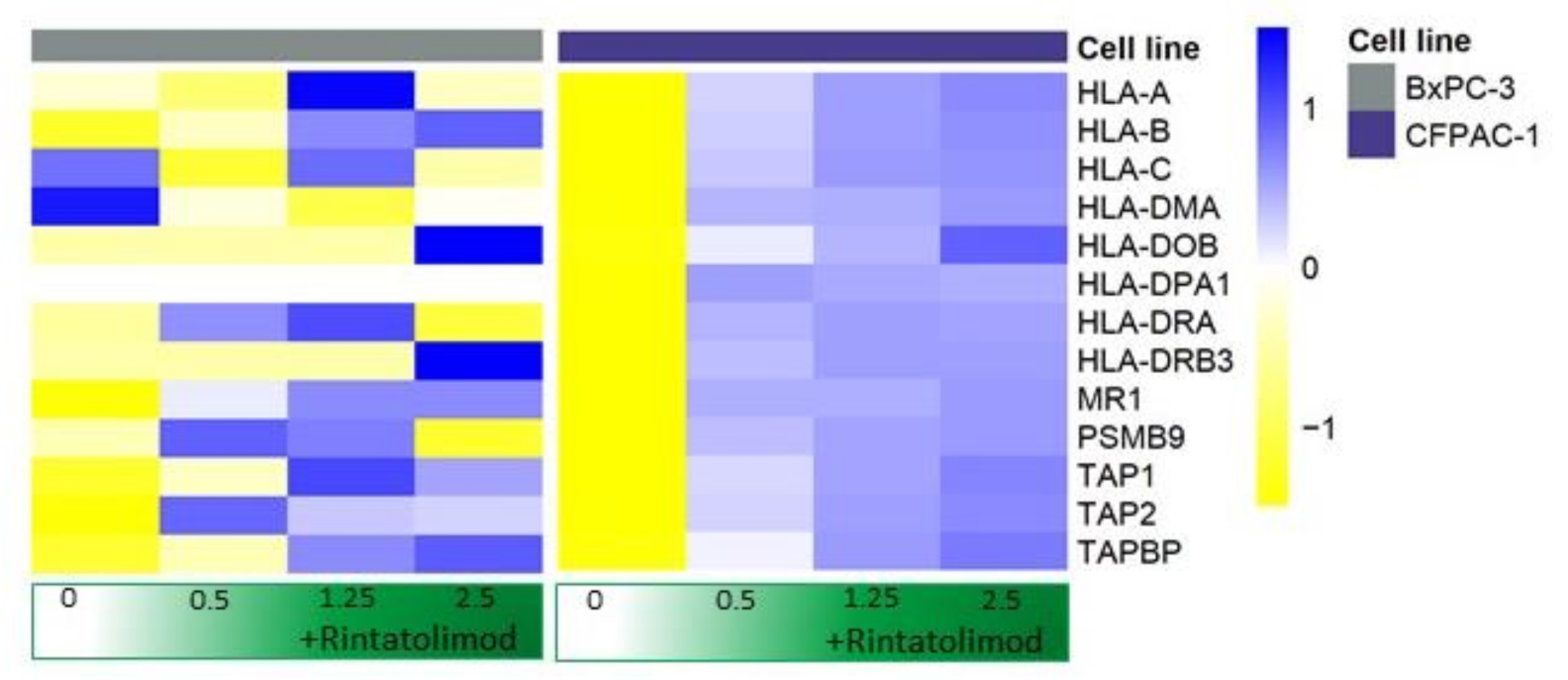
3.3. Rintatolimod Induced the Expression of MHC Class I and II Histocompatibility in HPAC Cells
3.4. Proteomics Measurements Validated the Antiviral Activities in HPAC Cells Treated with Rintatolimod
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coronavirus Disease (COVID-19) Outbreak World Health Organization. 2020. Available online: www.who.int/emergencies/diseases/novel-coronavirus-2019/events-as-they-happen (accessed on 5 May 2021).
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.P.; Look, D.C.; Shi, L.; Hickey, M.; Pewe, L.; Netland, J.; Farzan, M.; Wohlford-Lenane, C.; Perlman, S.; Paul, B.M., Jr. ACE2 receptor expression and severe acute respiratory syndrome coronavirus infection depend on differentiation of human airway epithelia. J. Virol. 2005, 79, 14614–14621. [Google Scholar] [CrossRef]
- Shah, V.K.; Firmal, P.; Alam, A.; Ganguly, D.; Chattopadhyay, S. Overview of Immune Response During SARS-CoV-2 Infection: Lessons from the Past. Front. Immunol. 2020, 11, 1949. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yen, Y.T.; Singh, S.; Kao, C.L.; Wu Hsieh, B.A. SARS-CoV regulates immune function-related gene expression in human monocytic cells. Viral Immunol. 2012, 25, 277–288. [Google Scholar] [CrossRef]
- Wu, H.S.; Hsieh, Y.C.; Su, I.J.; Lin, T.H.; Chiu, S.C.; Hsu, Y.F.; Lin, J.H.; Wang, M.C.; Chen, J.Y.; Hsiao, P.W.; et al. Early detection of antibodies against various structural proteins of the SARS-associated coronavirus in SARS patients. J. Biomed. Sci. 2004, 11, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Chen, X.; Xu, A. Profile of specific antibodies to the SARS-associated coronavirus. N. Engl. J. Med. 2003, 349, 508–509. [Google Scholar] [CrossRef] [PubMed]
- Li, C.K.F.; Wu, H.; Yan, H.; Ma, S.; Wang, L.; Zhang, M.; Tang, X.; Temperton, N.J.; Weiss, R.A.; Brenchley, J.M.; et al. T cell responses to whole SARS coronavirus in humans. J. Immunol. 2008, 181, 5490–5500. [Google Scholar] [CrossRef]
- COVIDSurg Collaborative. Mortality and pulmonary complications in patients undergoing surgery with perioperative SARS-CoV-2 infection: An international cohort study. Lancet 2020, 396, 27–38. [Google Scholar] [CrossRef]
- Kim, D.H. Nutritional issues in patients with cancer. Intest. Res. 2019, 17, 455–462. [Google Scholar] [CrossRef]
- Dai, M.; Liu, D.; Liu, M.; Zhou, F.; Li, G.; Chen, Z.; Zhang, Z.; You, H.; Wu, M.; Zheng, Q.; et al. Patients with Cancer Appear More Vulnerable to SARS-CoV-2: A Multicenter Study during the COVID-19 Outbreak. Cancer Discov. 2020, 10, 783–791. [Google Scholar]
- Garassino, M.C.; Whisenant, J.G.; Huang, L.; Trama, A.; Torri, V.; Agustoni, F.; Baena, J.; Banna, G.; Berardi, R.; Bettini, A.C. COVID-19 in patients with thoracic malignancies (TERAVOLT): First results of an international, registry-based, cohort study. Lancet Oncol. 2020, 21, 914–922. [Google Scholar] [CrossRef]
- Lee, A.J.X.; Purshouse, K. COVID-19 and cancer registries: Learning from the first peak of the SARS-CoV-2 pandemic. Br. J. Cancer 2021, 124, 1777–1784. [Google Scholar] [CrossRef]
- Buijs, P.R.; van Eijck, C.H.; Hofland, L.J.; Fouchier, R.A.; van den Hoogen, B.G. Different responses of human pancreatic adenocarcinoma cell lines to oncolytic Newcastle disease virus infection. Cancer Gene Ther. 2014, 21, 24–30. [Google Scholar] [CrossRef]
- Magar, R.; Yadav, P.; Farimani, A.B. Potential Neutralizing Antibodies Discovered for Novel Corona Virus Using Machine Learning Rishikesh Magar. BioRxiv 2020, 11, 5261. [Google Scholar]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; van Haperen, R.; Osterhaus, A.D.M.E.; van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef]
- Colson, P.; Rolain, J.M.; Lagier, J.C.; Brouqui, P.; Raoult, D. Chloroquine and hydroxychloroquine as available weapons to fight COVID-19. Int. J. Antimicrob. Agents 2020, 55, 105932. [Google Scholar] [CrossRef]
- Grein, J.; Ohmagari, N.; Shin, D.; Diaz, G.; Asperges, E.; Castagna, A.; Feldt, T.; Green, G.; Green, M.L.; Lescure, F.X.; et al. Compassionate Use of Remdesivir for Patients with Severe Covid-19. N. Engl. J. Med. 2020, 382, 2327–2336. [Google Scholar] [CrossRef]
- Souto, S.C.; Reis, L.O.; Palma, T.; Palma, P.; Denardi, F. Prospective and randomized comparison of electrical stimulation of the posterior tibial nerve versus oxybutynin versus their combination for treatment of women with overactive bladder syndrome. World J. Urol. 2014, 32, 179–184. [Google Scholar] [CrossRef]
- Smith, M.; Martínez, E.G.; Pitter, M.R.; Fucikova, J.; Spisek, R.; Zitvogel, L.; Kroemer, G.; Galluzzi, L. Trial Watch: Toll-like receptor agonists in cancer immunotherapy. Oncoimmunology 2018, 7, e1526250. [Google Scholar] [CrossRef] [PubMed]
- Ts’o, P.O.; Alderfer, J.L.; Levy, J.; Marshall, L.W.; O’Malley, J.; Horoszewicz, J.S.; Carter, W.A. An integrated and comparative study of the antiviral effects and other biological properties of the polyinosinic acid-polycytidylic acid and its mismatched analogues. Mol. Pharmacol. 1976, 12, 299–312. [Google Scholar] [PubMed]
- Mitchell, W.M.; Nicodemus, C.F.; Carter, W.A.; Horvath, J.C.; Strayer, D.R. Discordant biological and toxicological species responses to TLR3 activation. Am. J. Pathol. 2014, 184, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.R.; Carter, W.A.; Stouch, B.C.; Stevens, S.R.; Bateman, L.; Cimoch, P.J.; Lapp, C.W.; Peterson, D.L.; Mitchell, W.M. Chronic Fatigue Syndrome AMP-516 Study Group. A double-blind, placebo-controlled, randomized, clinical trial of the TLR-3 agonist rintatolimod in severe cases of chronic fatigue syndrome. PLoS ONE 2012, 7, e31334. [Google Scholar] [CrossRef]
- Day, C.W.; Baric, R.; Cai, S.X.; Frieman, M.; Kumaki, Y.; Morrey, J.D.; Smee, D.F.; Barnarda, D.L. A new mouse-adapted strain of SARS-CoV as a lethal model for evaluating antiviral agents in vitro and in vivo. Virology 2009, 395, 210–222. [Google Scholar] [CrossRef]
- Overton, E.T.; Goepfert, P.A.; Cunningham, P.; Carter, W.A.; Horvath, J.; Young, D.; Strayer, D.R. Intranasal seasonal influenza vaccine and a TLR-3 agonist, rintatolimod, induced cross-reactive IgA antibody formation against avian H5N1 and H7N9 influenza HA in humans. Vaccine 2014, 32, 5490–5495. [Google Scholar] [CrossRef]
- Kozlowski, A.; Charles, S.A.; Harris, J.M. Development of pegylated interferons for the treatment of chronic hepatitis C. BioDrugs 2001, 15, 419–429. [Google Scholar] [CrossRef]
- Nicodemus, C.F.; Berek, J.S. TLR3 agonists as immunotherapeutic agents. Immunotherapy 2010, 2, 137–140. [Google Scholar] [CrossRef]
- Thompson, K.A.; Strayer, D.R.; Salvato, P.D.; Thompson, C.E.; Klimas, N.; Molavi, A.; Hamill, A.K.; Zheng, Z.; Ventura, D.; Carter, W.A. Results of a double-blind placebo-controlled study of the double-stranded RNA drug polyI:polyC12U in the treatment of HIV infection. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 580–587. [Google Scholar] [CrossRef]
- Zarling, J.M.; Kung, P.C. Monoclonal antibodies which distinguish between human NK cells and cytotoxic T lymphocytes. Nature 1980, 288, 394–396. [Google Scholar] [CrossRef]
- Rosa, K. FDA Grants Orphan Drug Designation to Rintatolimod for Pancreatic Cancer: OncLive. 2020. Available online: https://www.onclive.com/view/fda-grants-orphan-drug-designation-to-rintatolimod-for-pancreatic-cancer (accessed on 5 May 2021).
- Rintatolimod and Pembrolizumab for the Treatment of Refractory Metastatic or Unresectable Colorectal Cancer: NIH library of Medicine. 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT04119830 (accessed on 5 May 2021).
- Vaccine Therapy in Combination with Rintatolimod and/or Sargramostim in Treating Patients with Stage II-IV HER2-Positive Breast Cancer: NIH Library of Medicine. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT01355393 (accessed on 5 May 2021).
- Monaco, G.; Lee, B.; Xu, W.; Mustafah, S.; Hwang, Y.Y.; Carré, C.; Burdin, N.; Visan, L.; Ceccarelli, M.; Poidinger, M.; et al. RNA-Seq Signatures Normalized by mRNA Abundance Allow Absolute Deconvolution of Human Immune Cell Types. Cell Rep. 2019, 26, 1627–1640.e7. [Google Scholar] [CrossRef]
- Kovach, M.A.; Standiford, T.J. Toll like receptors in diseases of the lung. Int. Immunopharmacol. 2011, 11, 1399–1406. [Google Scholar] [CrossRef]
- Chen, N.; Xia, P.; Li, S.; Zhang, T.; Wang, T.T.; Zhu, J. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life 2017, 69, 297–304. [Google Scholar] [CrossRef]
- Hewson, C.A.; Jardine, A.; Edwards, M.R.; Laza-Stanca, V.; Johnston, S.L. Toll-like receptor 3 is induced by and mediates antiviral activity against rhinovirus infection of human bronchial epithelial cells. J. Virol. 2005, 79, 12273–12279. [Google Scholar] [CrossRef]
- Uehara, A.; Fujimoto, Y.; Fukase, K.; Takada, H. Various human epithelial cells express functional Toll-like receptors, NOD1 and NOD2 to produce anti-microbial peptides, but not proinflammatory cytokines. Mol. Immunol. 2007, 44, 3100–3111. [Google Scholar] [CrossRef]
- Mustafa, D.A.M.; Dekker, L.J.; Stingl, C.; Kremer, A.; Stoop, M.; Smitt, P.A.E.S.; Kros, J.M.; Luider, T.M. A proteome comparison between physiological angiogenesis and angiogenesis in glioblastoma. Mol. Cell Proteom. 2012, 11, 008466. [Google Scholar] [CrossRef]
- Riverol, Y.P.; Csordas, A.; Bai, J.; Llinares, M.B.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Khalil, N.; O’Connor, R.N.; Flanders, K.C.; Unruh, H. TGF-beta 1, but not TGF-beta 2 or TGF-beta 3, is differentially present in epithelial cells of advanced pulmonary fibrosis: An immunohistochemical study. Am. J. Respir Cell Mol. Biol. 1996, 14, 131–138. [Google Scholar] [CrossRef]
- Barnard, D.L.; Day, C.W.; Bailey, K.; Heiner, M.; Montgomery, R.; Lauridsen, L.; Chan, P.K.S.; Sidwell, R.W. Evaluation of immunomodulators, interferons and known in vitro SARS-coV inhibitors for inhibition of SARS-coV replication in BALB/c mice. Antivir. Chem. Chemother. 2006, 17, 275–284. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; López, C.T.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identified Proteins | Index Number | Associated Gene | Number of Peptides # | |||
|---|---|---|---|---|---|---|
| CN * | 2.5 ** | 1.25 ** | 0.5 ** | |||
| Probable ATP-dependent RNA helicase DDX60 | Q8IY21 | DDX60 | 0 | 27 | 21 | 11 |
| 2′–5′-oligoadenylate synthase-like protein | Q15646 | OASL | 0 | 7 | 4 | 3 |
| Single-stranded DNA-binding protein | P42566 | SSBP1 | 0 | 4 | 5 | 1 |
| Epidermal growth factor receptor substrate 15 | P42566 | EPS15L1 | 0 | 3 | 3 | 3 |
| Probable ATP-dependent RNA helicase DDX58 | O95786 | DDX58 | 0 | 3 | 3 | 1 |
| Transcription elongation factor SPT6 | Q7KZ85 | SUPT6H | 0 | 3 | 3 | 1 |
| HLA class I histocompatibility antigen, alpha chain E | P13747 | HLA-E | 0 | 3 | 2 | 2 |
| Interferon regulatory factor 3 | Q14653 | IRF3 | 0 | 1 | 0 | 2 |
| Interferon-induced protein with tetratricopeptide repeats 3 | O14879 | IFIT3 | 0 | 2 | 1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mustafa, D.A.M.; Saida, L.; Latifi, D.; Wismans, L.V.; de Koning, W.; Zeneyedpour, L.; Luider, T.M.; van den Hoogen, B.; van Eijck, C.H.J. Rintatolimod Induces Antiviral Activities in Human Pancreatic Cancer Cells: Opening for an Anti-COVID-19 Opportunity in Cancer Patients? Cancers 2021, 13, 2896. https://doi.org/10.3390/cancers13122896
Mustafa DAM, Saida L, Latifi D, Wismans LV, de Koning W, Zeneyedpour L, Luider TM, van den Hoogen B, van Eijck CHJ. Rintatolimod Induces Antiviral Activities in Human Pancreatic Cancer Cells: Opening for an Anti-COVID-19 Opportunity in Cancer Patients? Cancers. 2021; 13(12):2896. https://doi.org/10.3390/cancers13122896
Chicago/Turabian StyleMustafa, Dana A. M., Lawlaw Saida, Diba Latifi, Leonoor V. Wismans, Willem de Koning, Lona Zeneyedpour, Theo M. Luider, Bernadette van den Hoogen, and Casper H. J. van Eijck. 2021. "Rintatolimod Induces Antiviral Activities in Human Pancreatic Cancer Cells: Opening for an Anti-COVID-19 Opportunity in Cancer Patients?" Cancers 13, no. 12: 2896. https://doi.org/10.3390/cancers13122896
APA StyleMustafa, D. A. M., Saida, L., Latifi, D., Wismans, L. V., de Koning, W., Zeneyedpour, L., Luider, T. M., van den Hoogen, B., & van Eijck, C. H. J. (2021). Rintatolimod Induces Antiviral Activities in Human Pancreatic Cancer Cells: Opening for an Anti-COVID-19 Opportunity in Cancer Patients? Cancers, 13(12), 2896. https://doi.org/10.3390/cancers13122896