Tunneling Nanotubes and Tumor Microtubes in Cancer



Abstract
1. Introduction
2. Structure of TNTs
3. Structure of TMs
4. Technical Challenges Regarding Membrane Tubes
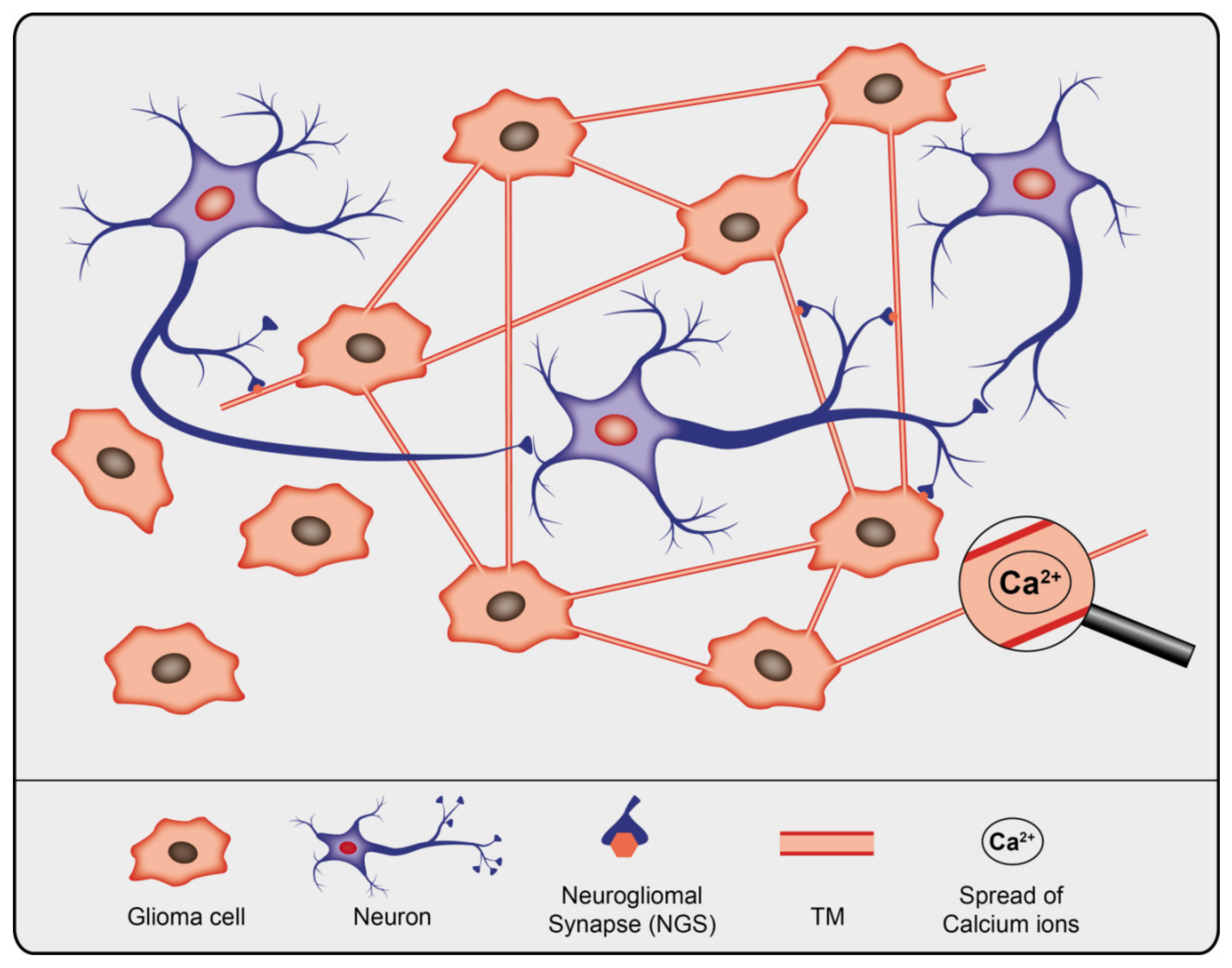
5. Occurrence of TNTs/TMs in Cancer Cells
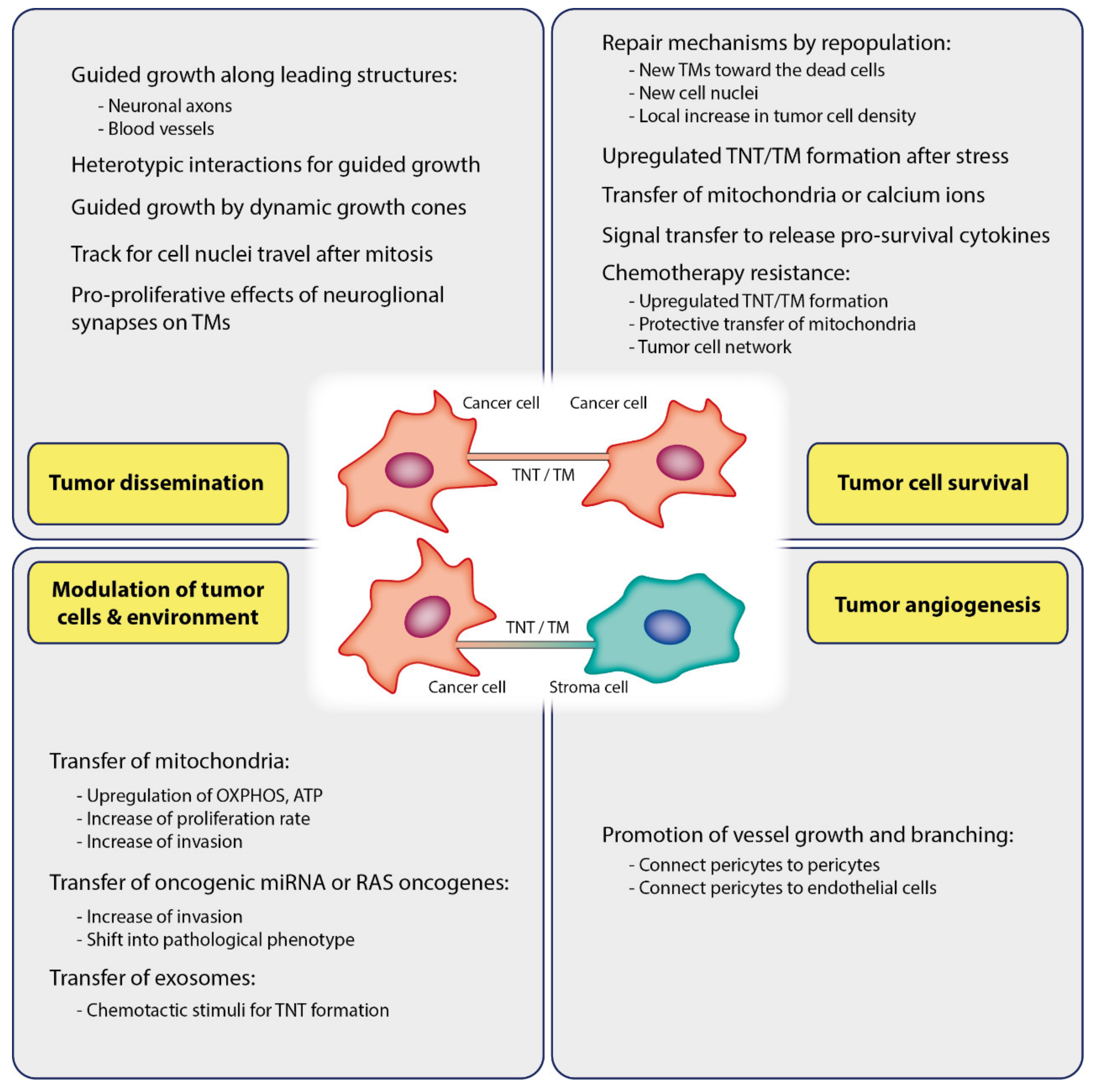
6. Functions of TNTs and TMs in Cancer
6.1. Cancer Growth and Invasion: Homotypic Interactions between Cancer Cells
6.2. Cancer Growth and Invasion: Heterotypic Interactions
6.3. Tumor Cell Survival
6.4. Modulating of Tumor Cells and Environment via TNT/TM Mediated Cargo
6.4.1. Transfer of Mitochondria
6.4.2. Transfer of microRNA
6.4.3. Transfer of Oncogenes
6.4.4. Transfer of Exosomes and Other Cargoes
6.5. Angiogenesis
6.6. Treatment Resistance
6.6.1. Resistance to Surgical Lesions
6.6.2. Resistance to Chemotherapy
6.6.3. Resistance to Radiotherapy
7. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALL | Acute lymphoblastic leukemia |
| AML | Acute myeloid leukemia |
| BTPC | Brain tumor propagating cells |
| Cx43 | Connexin 43 |
| EGF | Epidermal growth factor |
| EGFR | Epidermal growth factor receptor |
| GAP-43 | Growth associated protein-43 |
| HUVEC | Human umbilical vein endothelial cells |
| miRNA | MicroRNA |
| MHC II | Major histocompatibility complex II |
| MSC | Mesenchymal stem cells |
| NGS | Neurogliomal synapses |
| STED | Stimulated emission depletion |
| TNFaip2 | Tumor necrosis factor, alpha-induced protein 2 |
| TNTs | Tunneling nanotubes |
| TMs | Tumor microtubes |
| Ttyh-1 | Tweety-homolog 1 |
| VEGF | Vascular endothelial growth factor |
References
- Bissell, M.J.; Radisky, D. Putting Tumours in Context. Nat. Rev. Cancer 2001, 1, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Bard, M.P.; Hegmans, J.P.; Hemmes, A.; Luider, T.M.; Willemsen, R.; Severijnen, L.A.; van Meerbeeck, J.P.; Burgers, S.A.; Hoogsteden, H.C.; Lambrecht, B.N. Proteomic Analysis of Exosomes Isolated from Human Malignant Pleural Effusions. Am. J. Respir. Cell Mol. Biol. 2004, 31, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Hegmans, J.P.; Bard, M.P.; Hemmes, A.; Luider, T.M.; Kleijmeer, M.J.; Prins, J.B.; Zitvogel, L.; Burgers, S.A.; Hoogsteden, H.C.; Lambrecht, B.N. Proteomic Analysis of Exosomes Secreted by Human Mesothelioma Cells. Am. J. Pathol. 2004, 164, 1807–1815. [Google Scholar] [CrossRef]
- Cottin, S.; Ghani, K.; de Campos-Lima, P.O.; Caruso, M. Gemcitabine Intercellular Diffusion Mediated by Gap Junctions: New Implications for Cancer Therapy. Mol. Cancer 2010, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Naus, C.C.; Laird, D.W. Implications and Challenges of Connexin Connections to Cancer. Nat. Rev. Cancer 2010, 10, 435–441. [Google Scholar] [CrossRef]
- Bobrie, A.; Colombo, M.; Raposo, G.; Thery, C. Exosome Secretion: Molecular Mechanisms and Roles in Immune Responses. Traffic 2011, 12, 1659–1668. [Google Scholar] [CrossRef]
- Pap, E.; Pallinger, E.; Falus, A. The Role of Membrane Vesicles in Tumorigenesis. Crit. Rev. Oncol. Hematol. 2011, 79, 213–223. [Google Scholar] [CrossRef]
- Pap, E. The Role of Microvesicles in Malignancies. Adv. Exp. Med. Biol. 2011, 714, 183–199. [Google Scholar]
- Rustom, A. The Missing Link: Does Tunnelling Nanotube-Based Supercellularity Provide a New Understanding of Chronic and Lifestyle Diseases? Open Biol. 2016, 6, 160057. [Google Scholar] [CrossRef]
- Ariazi, J.; Benowitz, A.; de Biasi, V.; den Boer, M.L.; Cherqui, S.; Cui, H.; Douillet, N.; Eugenin, E.A.; Favre, D.; Goodman, S.; et al. Tunneling Nanotubes and Gap Junctions-Their Role in Long-Range Intercellular Communication During Development, Health, and Disease Conditions. Front. Mol. Neurosci. 2017, 10, 333. [Google Scholar] [CrossRef]
- Wang, X.; Veruki, M.L.; Bukoreshtliev, N.V.; Hartveit, E.; Gerdes, H.H. Animal Cells Connected by Nanotubes Can Be Electrically Coupled through Interposed Gap-Junction Channels. Proc. Natl. Acad. Sci. USA 2010, 107, 17194–17199. [Google Scholar] [CrossRef] [PubMed]
- Gousset, K.; Schiff, E.; Langevin, C.; Marijanovic, Z.; Caputo, A.; Browman, D.T.; Chenouard, N.; de Chaumont, F.; Martino, A.; Enninga, J.; et al. Prions Hijack Tunnelling Nanotubes for Intercellular Spread. Nat. Cell Biol. 2009, 11, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, H.H.; Bukoreshtliev, N.V.; Barroso, J.F. Tunneling Nanotubes: A New Route for the Exchange of Components between Animal Cells. FEBS Lett. 2007, 581, 2194–2201. [Google Scholar] [CrossRef] [PubMed]
- Eugenin, E.A.; Gaskill, P.J.; Berman, J.W. Tunneling Nanotubes (Tnt) Are Induced by Hiv-Infection of Macrophages: A Potential Mechanism for Intercellular Hiv Trafficking. Cell. Immunol. 2009, 254, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gerdes, H.H. Transfer of Mitochondria Via Tunneling Nanotubes Rescues Apoptotic Pc12 Cells. Cell Death Differ. 2015, 22, 1181–1191. [Google Scholar] [CrossRef]
- Cselenyak, A.; Pankotai, E.; Horvath, E.M.; Kiss, L.; Lacza, Z. Mesenchymal Stem Cells Rescue Cardiomyoblasts from Cell Death in an in Vitro Ischemia Model Via Direct Cell-to-Cell Connections. BMC Cell Biol. 2010, 11, 29. [Google Scholar] [CrossRef]
- Hase, K.; Kimura, S.; Takatsu, H.; Ohmae, M.; Kawano, S.; Kitamura, H.; Ito, M.; Watarai, H.; Hazelett, C.C.; Yeaman, C.; et al. M-Sec Promotes Membrane Nanotube Formation by Interacting with Ral and the Exocyst Complex. Nat. Cell Biol. 2009, 11, 1427–1432. [Google Scholar] [CrossRef]
- Yasuda, K.; HPark, C.; Ratliff, B.; Addabbo, F.; Hatzopoulos, A.K.; Chander, P.; Goligorsky, M.S. Adriamycin Nephropathy: A Failure of Endothelial Progenitor Cell-Induced Repair. Am. J. Pathol. 2010, 176, 1685–1695. [Google Scholar] [CrossRef]
- Plotnikov, E.Y.; Khryapenkova, T.G.; Galkina, S.I.; Sukhikh, G.T.; Zorov, D.B. Cytoplasm and Organelle Transfer between Mesenchymal Multipotent Stromal Cells and Renal Tubular Cells in Co-Culture. Exp. Cell Res. 2010, 316, 2447–2455. [Google Scholar] [CrossRef]
- Ranzinger, J.; Rustom, A.; Abel, M.; Leyh, J.; Kihm, L.; Witkowski, M.; Scheurich, P.; Zeier, M.; Schwenger, V. Nanotube Action between Human Mesothelial Cells Reveals Novel Aspects of Inflammatory Responses. PLoS ONE 2011, 6, e29537. [Google Scholar] [CrossRef]
- Lou, E.; Fujisawa, S.; Morozov, A.; Barlas, A.; Romin, Y.; Dogan, Y.; Gholami, S.; Moreira, A.L.; Manova-Todorova, K.; Moore, M.A. Tunneling Nanotubes Provide a Unique Conduit for Intercellular Transfer of Cellular Contents in Human Malignant Pleural Mesothelioma. PLoS ONE 2012, 7, e33093. [Google Scholar] [CrossRef] [PubMed]
- Watkins, S.C.; Salter, R.D. Functional Connectivity between Immune Cells Mediated by Tunneling Nanotubules. Immunity 2005, 23, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Salter, R.D.; Watkins, S.C. Dynamic Properties of Antigen Uptake and Communication between Dendritic Cells. Immunol. Res. 2006, 36, 211–220. [Google Scholar] [CrossRef]
- Wittig, D.; Wang, X.; Walter, C.; Gerdes, H.H.; Funk, R.H.; Roehlecke, C. Multi-Level Communication of Human Retinal Pigment Epithelial Cells Via Tunneling Nanotubes. PLoS ONE 2012, 7, e33195. [Google Scholar] [CrossRef] [PubMed]
- Sowinski, S.; Jolly, C.; Berninghausen, O.; Purbhoo, M.A.; Chauveau, A.; Kohler, K.; Oddos, S.; Eissmann, P.; Brodsky, F.M.; Hopkins, C.; et al. Membrane Nanotubes Physically Connect T Cells over Long Distances Presenting a Novel Route for Hiv-1 Transmission. Nat. Cell Biol. 2008, 10, 211–219. [Google Scholar] [CrossRef]
- Rudnicka, D.; Feldmann, J.; Porrot, F.; Wietgrefe, S.; Guadagnini, S.; Prevost, M.C.; Estaquier, J.; Haase, A.T.; Sol-Foulon, N.; Schwartz, O. Simultaneous Cell-to-Cell Transmission of Human Immunodeficiency Virus to Multiple Targets through Polysynapses. J. Virol. 2009, 83, 6234–6246. [Google Scholar] [CrossRef]
- Xu, W.; Santini, P.A.; Sullivan, J.S.; He, B.; Shan, M.; Ball, S.C.; Dyer, W.B.; Ketas, T.J.; Chadburn, A.; Cohen-Gould, L.; et al. Hiv-1 Evades Virus-Specific Igg2 and Iga Responses by Targeting Systemic and Intestinal B Cells Via Long-Range Intercellular Conduits. Nat. Immunol. 2009, 10, 1008–1017. [Google Scholar] [CrossRef]
- Galkina, S.I.; Stadnichuk, V.I.; Molotkovsky, J.G.; Romanova, J.M.; Sud’ina, G.F.; Klein, T. Microbial Alkaloid Staurosporine Induces Formation of Nanometer-Wide Membrane Tubular Extensions (Cytonemes, Membrane Tethers) in Human Neutrophils. Cell Adh. Migr. 2010, 4, 32–38. [Google Scholar] [CrossRef][Green Version]
- Koyanagi, M.; Brandes, R.P.; Haendeler, J.; Zeiher, A.M.; Dimmeler, S. Cell-to-Cell Connection of Endothelial Progenitor Cells with Cardiac Myocytes by Nanotubes: A Novel Mechanism for Cell Fate Changes? Circ. Res. 2005, 96, 1039–1041. [Google Scholar] [CrossRef]
- Domhan, S.; Ma, L.; Tai, A.; Anaya, Z.; Beheshti, A.; Zeier, M.; Hlatky, L.; Abdollahi, A. Intercellular Communication by Exchange of Cytoplasmic Material Via Tunneling Nano-Tube Like Structures in Primary Human Renal Epithelial Cells. PLoS ONE 2011, 6, e21283. [Google Scholar] [CrossRef]
- Gurke, S.; Barroso, J.F.; Hodneland, E.; Bukoreshtliev, N.V.; Schlicker, O.; Gerdes, H.H. Tunneling Nanotube (Tnt)-Like Structures Facilitate a Constitutive, Actomyosin-Dependent Exchange of Endocytic Organelles between Normal Rat Kidney Cells. Exp. Cell Res. 2008, 314, 3669–3683. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.C.; Stark, D.A.; Teddy, J.; Kulesa, P.M. Neural Crest Cell Communication Involves an Exchange of Cytoplasmic Material through Cellular Bridges Revealed by Photoconversion of Kikgr. Dev. Dyn. 2011, 240, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Gradilla, A.C.; Seijo, I.; Andres, G.; Rodriguez-Navas, C.; Gonzalez-Mendez, L.; Guerrero, I. Cytonemes Are Required for the Establishment of a Normal Hedgehog Morphogen Gradient in Drosophila Epithelia. Nat. Cell Biol. 2013, 15, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Weber, F.A.; Kornberg, T.B. Cytonemes: Cellular Processes That Project to the Principal Signaling Center in Drosophila Imaginal Discs. Cell 1999, 97, 599–607. [Google Scholar] [CrossRef]
- Inaba, M.; Buszczak, M.; Yamashita, Y.M. Nanotubes Mediate Niche-Stem-Cell Signalling in the Drosophila Testis. Nature 2015, 523, 329–332. [Google Scholar] [CrossRef]
- Chinnery, H.R.; Pearlman, E.; McMenamin, P.G. Cutting Edge: Membrane Nanotubes in Vivo: A Feature of Mhc Class Ii+ Cells in the Mouse Cornea. J. Immunol. 2008, 180, 5779–5783. [Google Scholar] [CrossRef]
- Seyed-Razavi, Y.; Hickey, M.J.; Kuffova, L.; McMenamin, P.G.; Chinnery, H.R. Membrane Nanotubes in Myeloid Cells in the Adult Mouse Cornea Represent a Novel Mode of Immune Cell Interaction. Immunol. Cell Biol. 2013, 91, 89–95. [Google Scholar] [CrossRef]
- Osswald, M.; Jung, E.; Sahm, F.; Solecki, G.; Venkataramani, V.; Blaes, J.; Weil, S.; Horstmann, H.; Wiestler, B.; Syed, M.; et al. Brain Tumour Cells Interconnect to a Functional and Resistant Network. Nature 2015, 528, 93–98. [Google Scholar] [CrossRef]
- Rehberg, M.; Nekolla, K.; Sellner, S.; Praetner, M.; Mildner, K.; Zeuschner, D.; Krombach, F. Intercellular Transport of Nanomaterials Is Mediated by Membrane Nanotubes in Vivo. Small 2016, 12, 1882–1890. [Google Scholar] [CrossRef]
- Kenny, P.A.; Lee, G.Y.; Bissell, M.J. Targeting the Tumor Microenvironment. Front. Biosci. 2007, 12, 3468–3474. [Google Scholar] [CrossRef]
- Bissell, M.J.; Hines, W.C. Why Don’t We Get More Cancer? A Proposed Role of the Microenvironment in Restraining Cancer Progression. Nat. Med. 2011, 17, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Ruckert, F.; Grutzmann, R.; Pilarsky, C. Feedback within the Inter-Cellular Communication and Tumorigenesis in Carcinomas. PLoS ONE 2012, 7, e36719. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Aranda, V.; Bardelli, A.; Blanpain, C.; Bock, C.; Borowski, C.; Caldas, C.; Califano, A.; Doherty, M.; Elsner, M.; et al. Toward Understanding and Exploiting Tumor Heterogeneity. Nat. Med. 2015, 21, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Andor, N.; Graham, T.A.; Jansen, M.; Xia, L.C.; Aktipis, C.A.; Petritsch, C.; Ji, H.P.; Maley, C.C. Pan-Cancer Analysis of the Extent and Consequences of Intratumor Heterogeneity. Nat. Med. 2016, 22, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Altmann, C.; Keller, S.; Schmidt, M.H.H. The Role of Svz Stem Cells in Glioblastoma. Cancers 2019, 11, 448. [Google Scholar] [CrossRef] [PubMed]
- Fidler, I.J. The Pathogenesis of Cancer Metastasis: The ‘Seed and Soil’ Hypothesis Revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef]
- Fidler, I.J. The Biology of Brain Metastasis: Challenges for Therapy. Cancer J. 2015, 21, 284–293. [Google Scholar] [CrossRef]
- Alexander, S.; Friedl, P. Cancer Invasion and Resistance: Interconnected Processes of Disease Progression and Therapy Failure. Trends Mol. Med. 2012, 18, 13–26. [Google Scholar] [CrossRef]
- Axelrod, R.; Axelrod, D.E.; Pienta, K.J. Evolution of Cooperation among Tumor Cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13474–13479. [Google Scholar] [CrossRef]
- Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H.H. Nanotubular Highways for Intercellular Organelle Transport. Science 2004, 303, 1007–1010. [Google Scholar] [CrossRef]
- Bukoreshtliev, N.V.; Wang, X.; Hodneland, E.; Gurke, S.; Barroso, J.F.; Gerdes, H.H. Selective Block of Tunneling Nanotube (Tnt) Formation Inhibits Intercellular Organelle Transfer between Pc12 Cells. FEBS Lett. 2009, 583, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cui, J.; Sun, X.; Zhang, Y. Tunneling-Nanotube Development in Astrocytes Depends on P53 Activation. Cell Death Differ. 2011, 18, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Onfelt, B.; Purbhoo, M.A.; Nedvetzki, S.; Sowinski, S.; Davis, D.M. Long-Distance Calls between Cells Connected by Tunneling Nanotubules. Sci. STKE 2005, 2005, pe55. [Google Scholar] [CrossRef] [PubMed]
- Veranic, P.; Lokar, M.; Schutz, G.J.; Weghuber, J.; Wieser, S.; Hagerstrand, H.; Kralj-Iglic, V.; Iglic, A. Different Types of Cell-to-Cell Connections Mediated by Nanotubular Structures. Biophys. J. 2008, 95, 4416–4425. [Google Scholar] [CrossRef] [PubMed]
- Wiger, A.M.; Gerdes, H.-H.; Wang, X. Tunneling Nanotubes. Commun. Integr. Biol. 2014, 7, e27934. [Google Scholar]
- Ady, J.W.; Desir, S.; Thayanithy, V.; Vogel, R.I.; Moreira, A.L.; Downey, R.J.; Fong, Y.; Manova-Todorova, K.; Moore, M.A.; Lou, E. Intercellular Communication in Malignant Pleural Mesothelioma: Properties of Tunneling Nanotubes. Front. Physiol. 2014, 5, 400. [Google Scholar] [CrossRef]
- Thayanithy, V.; Dickson, E.L.; Steer, C.; Subramanian, S.; Lou, E. Tumor-Stromal Cross Talk: Direct Cell-to-Cell Transfer of Oncogenic Micrornas Via Tunneling Nanotubes. Transl. Res. 2014, 164, 359–365. [Google Scholar] [CrossRef]
- Pasquier, J.; Guerrouahen, B.S.; Al Thawadi, H.; Ghiabi, P.; Maleki, M.; Abu-Kaoud, N.; Jacob, A.; Mirshahi, M.; Galas, L.; Rafii, S.; et al. Preferential Transfer of Mitochondria from Endothelial to Cancer Cells through Tunneling Nanotubes Modulates Chemoresistance. J. Transl. Med. 2013, 11, 94. [Google Scholar] [CrossRef]
- Wang, X.; Gerdes, H.H. Long-Distance Electrical Coupling Via Tunneling Nanotubes. Biochim. Biophys. Acta 2012, 1818, 2082–2086. [Google Scholar] [CrossRef]
- Sherer, N.M.; Mothes, W. Cytonemes and Tunneling Nanotubules in Cell-Cell Communication and Viral Pathogenesis. Trends Cell Biol. 2008, 18, 414–420. [Google Scholar] [CrossRef]
- Sherer, N.M.; Lehmann, M.J.; Jimenez-Soto, L.F.; Horensavitz, C.; Pypaert, M.; Mothes, W. Retroviruses Can Establish Filopodial Bridges for Efficient Cell-to-Cell Transmission. Nat. Cell Biol. 2007, 9, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.M.; Sowinski, S. Membrane Nanotubes: Dynamic Long-Distance Connections between Animal Cells. Nat. Rev. Mol. Cell Biol. 2008, 9, 431–436. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Shi, X.; Zhang, X.; Dang, S.; Ma, X.; Liu, F.; Xu, M.; Lv, Z.; Han, D.; Fang, X.; et al. Long-Distance Intercellular Connectivity between Cardiomyocytes and Cardiofibroblasts Mediated by Membrane Nanotubes. Cardiovasc. Res. 2011, 92, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Lou, E.; Zhai, E.; Sarkari, A.; Desir, S.; Wong, P.; Iizuka, Y.; Yang, J.; Subramanian, S.; McCarthy, J.; Bazzaro, M.; et al. Cellular and Molecular Networking within the Ecosystem of Cancer Cell Communication Via Tunneling Nanotubes. Front. Cell Dev. Biol. 2018, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Matejka, N.; Reindl, J. Perspectives of Cellular Communication through Tunneling Nanotubes in Cancer Cells and the Connection to Radiation Effects. Radiat. Oncol. 2019, 14, 218. [Google Scholar] [CrossRef]
- Hanna, S.J.; McCoy-Simandle, K.; Miskolci, V.; Guo, P.; Cammer, M.; Hodgson, L.; Cox, D. The Role of Rho-Gtpases and Actin Polymerization During Macrophage Tunneling Nanotube Biogenesis. Sci. Rep. 2017, 7, 8547. [Google Scholar] [CrossRef]
- Ohno, H.; Hase, K.; Kimura, S. M-Sec: Emerging Secrets of Tunneling Nanotube Formation. Commun. Integr. Biol. 2010, 3, 231–233. [Google Scholar] [CrossRef]
- D’Aloia, A.; Berruti, G.; Costa, B.; Schiller, C.; Ambrosini, R.; Pastori, V.; Martegani, E.; Ceriani, M. Ralgps2 Is Involved in Tunneling Nanotubes Formation in 5637 Bladder Cancer Cells. Exp. Cell Res. 2018, 362, 349–361. [Google Scholar] [CrossRef]
- Burtey, A.; Wagner, M.; Hodneland, E.; Skaftnesmo, K.O.; Schoelermann, J.; Mondragon, I.R.; Espedal, H.; Golebiewska, A.; Niclou, S.P.; Bjerkvig, R.; et al. Intercellular Transfer of Transferrin Receptor by a Contact-, Rab8-Dependent Mechanism Involving Tunneling Nanotubes. FASEB J. 2015, 29, 4695–4712. [Google Scholar] [CrossRef]
- Andresen, V.; Wang, X.; Ghimire, S.; Omsland, M.; Gjertsen, B.T.; Gerdes, H.H. Tunneling Nanotube (Tnt) Formation Is Independent of P53 Expression. Cell Death Differ. 2013, 20, 1124. [Google Scholar] [CrossRef]
- Mittal, R.; Karhu, E.; Wang, J.S.; Delgado, S.; Zukerman, R.; Mittal, J.; Jhaveri, V.M. Cell Communication by Tunneling Nanotubes: Implications in Disease and Therapeutic Applications. J. Cell Physiol. 2019, 234, 1130–1146. [Google Scholar] [CrossRef] [PubMed]
- Hsiung, F.; Ramirez-Weber, F.A.; Iwaki, D.D.; Kornberg, T.B. Dependence of Drosophila Wing Imaginal Disc Cytonemes on Decapentaplegic. Nature 2005, 437, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mendez, L.; AGradilla, C.; Guerrero, I. The Cytoneme Connection: Direct Long-Distance Signal Transfer During Development. Development 2019, 146, dev174607. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, S.; Kornberg, T.B. Glutamate Signaling at Cytoneme Synapses. Science 2019, 363, 948–955. [Google Scholar] [CrossRef]
- Venkataramani, V.; Tanev, D.I.; Strahle, C.; Studier-Fischer, A.; Fankhauser, L.; Kessler, T.; Korber, C.; Kardorff, M.; Ratliff, M.; Xie, R.; et al. Glutamatergic Synaptic Input to Glioma Cells Drives Brain Tumour Progression. Nature 2019, 573, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, H.S.; Morishita, W.; Geraghty, A.C.; Silverbush, D.; Gillespie, S.M.; Arzt, M.; Tam, L.T.; Espenel, C.; Ponnuswami, A.; Ni, L.; et al. Electrical and Synaptic Integration of Glioma into Neural Circuits. Nature 2019, 573, 539–545. [Google Scholar] [CrossRef]
- Matthias, O.; Solecki, G.; Wick, W.; Winkler, F. A Malignant Cellular Network in Gliomas: Potential Clinical Implications. Neuro Oncol. 2016, 18, 479–485. [Google Scholar]
- Pontes, B.; Viana, N.B.; Campanati, L.; Farina, M.; Neto, V.M.; Nussenzveig, H.M. Structure and Elastic Properties of Tunneling Nanotubes. Eur. Biophys. J. 2008, 37, 121–129. [Google Scholar] [CrossRef]
- Dekker, T.J.; van de Velde, C.J.; van Pelt, G.W.; Kroep, J.R.; Julien, J.P.; Smit, V.T.; Tollenaar, R.A.; Mesker, W.E. Prognostic Significance of the Tumor-Stroma Ratio: Validation Study in Node-Negative Premenopausal Breast Cancer Patients from the Eortc Perioperative Chemotherapy (Pop) Trial (10854). Breast Cancer Res. Treat. 2013, 139, 371–379. [Google Scholar] [CrossRef]
- Wu, J.; Liang, C.; Chen, M.; Su, W. Association between Tumor-Stroma Ratio and Prognosis in Solid Tumor Patients: A Systematic Review and Meta-Analysis. Oncotarget 2016, 7, 68954–68965. [Google Scholar] [CrossRef]
- Lou, E.; Gholami, S.; Romin, Y.; Thayanithy, V.; Fujisawa, S.; Desir, S.; Steer, C.J.; Subramanian, S.; Fong, Y.; Manova-Todorova, K.; et al. Imaging Tunneling Membrane Tubes Elucidates Cell Communication in Tumors. Trends Cancer 2017, 3, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.; Osswald, M.; Blaes, J.; Wiestler, B.; Sahm, F.; Schmenger, T.; Solecki, G.; Deumelandt, K.; Kurz, F.T.; Xie, R.; et al. Tweety-Homolog 1 Drives Brain Colonization of Gliomas. J. Neurosci. 2017, 37, 6837–6850. [Google Scholar] [CrossRef] [PubMed]
- Thayanithy, V.; Babatunde, V.; Dickson, E.L.; Wong, P.; Oh, S.; Ke, X.; Barlas, A.; Fujisawa, S.; Romin, Y.; Moreira, A.L.; et al. Tumor Exosomes Induce Tunneling Nanotubes in Lipid Raft-Enriched Regions of Human Mesothelioma Cells. Exp. Cell Res. 2014, 323, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Desir, S.; O’Hare, P.; Vogel, R.I.; Sperduto, W.; Sarkari, A.; Dickson, E.L.; Wong, P.; Nelson, A.C.; Fong, Y.; Steer, C.J.; et al. Chemotherapy-Induced Tunneling Nanotubes Mediate Intercellular Drug Efflux in Pancreatic Cancer. Sci. Rep. 2018, 8, 9484. [Google Scholar] [CrossRef]
- Saenz-de-Santa-Maria, I.; Bernardo-Castineira, C.; Enciso, E.; Garcia-Moreno, I.; Chiara, J.L.; Suarez, C.; Chiara, M.D. Control of Long-Distance Cell-to-Cell Communication and Autophagosome Transfer in Squamous Cell Carcinoma Via Tunneling Nanotubes. Oncotarget 2017, 8, 20939–20960. [Google Scholar] [CrossRef]
- Antanaviciute, I.; Rysevaite, K.; Liutkevicius, V.; Marandykina, A.; Rimkute, L.; Sveikatiene, R.; Uloza, V.; Skeberdis, V.A. Long-Distance Communication between Laryngeal Carcinoma Cells. PLoS ONE 2014, 9, e99196. [Google Scholar] [CrossRef]
- Weil, S.; Osswald, M.; Solecki, G.; Grosch, J.; Jung, E.; Lemke, D.; Ratliff, M.; Hanggi, D.; Wick, W.; Winkler, F. Tumor Microtubes Convey Resistance to Surgical Lesions and Chemotherapy in Gliomas. Neuro Oncol. 2017, 19, 1316–1326. [Google Scholar] [CrossRef]
- Reindl, J.; Shevtsov, M.; Dollinger, G.; Stangl, S.; Multhoff, G. Membrane Hsp70-Supported Cell-to-Cell Connections Via Tunneling Nanotubes Revealed by Live-Cell Sted Nanoscopy. Cell Stress Chaperones 2019, 24, 213–221. [Google Scholar] [CrossRef]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of Human Brain Tumour Initiating Cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Linder, S. The Matrix Corroded: Podosomes and Invadopodia in Extracellular Matrix Degradation. Trends Cell Biol. 2007, 17, 107–117. [Google Scholar] [CrossRef]
- Murphy, D.A.; Courtneidge, S.A. The ‘Ins’ and ‘Outs’ of Podosomes and Invadopodia: Characteristics, Formation and Function. Nat. Rev. Mol. Cell Biol. 2011, 12, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zheng, X.; Li, F.; Yu, Y.; Chen, Z.; Liu, Z.; Wang, Z.; Xu, H.; Yang, W. Tunneling Nanotubes Promote Intercellular Mitochondria Transfer Followed by Increased Invasiveness in Bladder Cancer Cells. Oncotarget 2017, 8, 15539–15552. [Google Scholar] [CrossRef] [PubMed]
- Caicedo, A.; Fritz, V.; Brondello, J.M.; Ayala, M.; Dennemont, I.; Abdellaoui, N.; de Fraipont, F.; Moisan, A.; Prouteau, C.A.; Boukhaddaoui, H.; et al. Mitoception as a New Tool to Assess the Effects of Mesenchymal Stem/Stromal Cell Mitochondria on Cancer Cell Metabolism and Function. Sci. Rep. 2015, 5, 9073. [Google Scholar] [CrossRef] [PubMed]
- Hanna, S.J.; McCoy-Simandle, K.; Leung, E.; Genna, A.; Condeelis, J.; Cox, D. Tunneling Nanotubes, a Novel Mode of Tumor Cell-Macrophage Communication in Tumor Cell Invasion. J. Cell Sci. 2019, 132, jcs223321. [Google Scholar] [CrossRef]
- Connor, Y.; Tekleab, S.; Nandakumar, S.; Walls, C.; Tekleab, Y.; Husain, A.; Gadish, O.; Sabbisetti, V.; Kaushik, S.; Sehrawat, S.; et al. Physical Nanoscale Conduit-Mediated Communication between Tumour Cells and the Endothelium Modulates Endothelial Phenotype. Nat. Commun. 2015, 6, 8671. [Google Scholar] [CrossRef]
- Rimkute, L.; Jotautis, V.; Marandykina, A.; Sveikatiene, R.; Antanaviciute, I.; Skeberdis, V.A. The Role of Neural Connexins in Hela Cell Mobility and Intercellular Communication through Tunneling Tubes. BMC Cell Biol. 2016, 17, 3. [Google Scholar] [CrossRef]
- Desir, S.; Wong, P.; Turbyville, T.; Shetty, C.M.; Clark, C.; Zhai, E.; Romin, Y.; Manova-Todorova, K.; Starr, T.K.; Nissley, D.V.; et al. Intercellular Transfer of Oncogenic Kras Via Tunneling Nanotubes Introduces Intracellular Mutational Heterogeneity in Colon Cancer Cells. Cancers 2019, 11, 892. [Google Scholar] [CrossRef]
- Castro, M.A.; Grieneisen, V.A.; de Almeida, R.M. Disruption and De Novo Formation of Nanotubular Membrane Extensions in Sw620 Colon Carcinoma Cell Line During Cell Division. Cell Biol. Int. 2005, 29, 929–931. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y. Tunneling Nanotubes between Rat Primary Astrocytes and C6 Glioma Cells Alter Proliferation Potential of Glioma Cells. Neurosci. Bull. 2015, 31, 371–378. [Google Scholar] [CrossRef]
- Carone, C.; Genedani, S.; Leo, G.; Filaferro, M.; Fuxe, K.; Agnati, L.F. In Vitro Effects of Cocaine on Tunneling Nanotube Formation and Extracellular Vesicle Release in Glioblastoma Cell Cultures. J. Mol. Neurosci. 2015, 55, 42–50. [Google Scholar] [CrossRef]
- Ding, X.; Ma, M.; Teng, J.; Teng, R.K.; Zhou, S.; Yin, J.; Fonkem, E.; Huang, J.H.; Wu, E.; Wang, X. Exposure to Als-Ftd-Csf Generates Tdp-43 Aggregates in Glioblastoma Cells through Exosomes and Tnts-Like Structure. Oncotarget 2015, 6, 24178–24191. [Google Scholar] [CrossRef] [PubMed]
- Errede, M.; Mangieri, D.; Longo, G.; Girolamo, F.; de Trizio, I.; Vimercati, A.; Serio, G.; Frei, K.; Perris, R.; Virgintino, D. Tunneling Nanotubes Evoke Pericyte/Endothelial Communication During Normal and Tumoral Angiogenesis. Fluids Barriers CNS 2018, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Polak, R.; de Rooij, B.; Pieters, R.; den Boer, M.L. B-Cell Precursor Acute Lymphoblastic Leukemia Cells Use Tunneling Nanotubes to Orchestrate Their Microenvironment. Blood 2015, 126, 2404–2414. [Google Scholar] [CrossRef] [PubMed]
- Burt, R.; Dey, A.; Aref, S.; Aguiar, M.; Akarca, A.; Bailey, K.; Day, W.; Hooper, S.; Kirkwood, A.; Kirschner, K.; et al. Activated Stromal Cells Transfer Mitochondria to Rescue Acute Lymphoblastic Leukaemia Cells from Oxidative Stress. Blood 2019, 134, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Marlein, C.R.; Zaitseva, L.; Piddock, R.E.; Robinson, S.D.; Edwards, D.R.; Shafat, M.S.; Zhou, Z.; Lawes, M.; Bowles, K.M.; Rushworth, S.A. Nadph Oxidase-2 Derived Superoxide Drives Mitochondrial Transfer from Bone Marrow Stromal Cells to Leukemic Blasts. Blood 2017, 130, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Qiu, Y.; Shi, Y.; Cai, J.; Wang, B.; Wei, X.; Ke, Q.; Sui, X.; Wang, Y.; et al. Cell Adhesion-Mediated Mitochondria Transfer Contributes to Mesenchymal Stem Cell-Induced Chemoresistance on T Cell Acute Lymphoblastic Leukemia Cells. J. Hematol. Oncol. 2018, 11, 11. [Google Scholar] [CrossRef]
- Ady, J.; Thayanithy, V.; Mojica, K.; Wong, P.; Carson, J.; Rao, P.; Fong, Y.; Lou, E. Tunneling Nanotubes: An Alternate Route for Propagation of the Bystander Effect Following Oncolytic Viral Infection. Mol. Ther. Oncolytics 2016, 3, 16029. [Google Scholar] [CrossRef]
- Desir, S.; Dickson, E.L.; Vogel, R.I.; Thayanithy, V.; Wong, P.; Teoh, D.; Geller, M.A.; Steer, C.J.; Subramanian, S.; Lou, E. Tunneling Nanotube Formation Is Stimulated by Hypoxia in Ovarian Cancer Cells. Oncotarget 2016, 7, 43150–43161. [Google Scholar] [CrossRef]
- Kretschmer, A.; Zhang, F.; Somasekharan, S.P.; Tse, C.; Leachman, L.; Gleave, A.; Li, B.; Asmaro, I.; Huang, T.; Kotula, L.; et al. Stress-Induced Tunneling Nanotubes Support Treatment Adaptation in Prostate Cancer. Sci. Rep. 2019, 9, 7826. [Google Scholar] [CrossRef]
- Fulga, T.A.; Rorth, P. Invasive Cell Migration Is Initiated by Guided Growth of Long Cellular Extensions. Nat. Cell Biol. 2002, 4, 715–719. [Google Scholar] [CrossRef]
- Yee, K.T.; Simon, H.H.; Tessier-Lavigne, M.; O’Leary, D.M. Extension of Long Leading Processes and Neuronal Migration in the Mammalian Brain Directed by the Chemoattractant Netrin-1. Neuron 1999, 24, 607–622. [Google Scholar] [CrossRef]
- Lowery, L.A.; van Vactor, D. The Trip of the Tip: Understanding the Growth Cone Machinery. Nat. Rev. Mol. Cell Biol. 2009, 10, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, C.; Poppleton, H.; Kocak, M.; Hogg, T.L.; Fuller, C.; Hamner, B.; Oh, E.Y.; Gaber, M.W.; Finklestein, D.; Allen, M.; et al. A Perivascular Niche for Brain Tumor Stem Cells. Cancer Cell 2007, 11, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Cuddapah, V.A.; Robel, S.; Watkins, S.; Sontheimer, H. A Neurocentric Perspective on Glioma Invasion. Nat. Rev. Neurosci. 2014, 15, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.; Miyoshi, H.; Sampetrean, O.; Shinozaki, M.; Morimoto, Y.; Iwasawa, C.; Fukaya, R.; Mine, Y.; Masuda, H.; Maruyama, T.; et al. Visualization of Spatiotemporal Dynamics of Human Glioma Stem Cell Invasion. Mol. Brain 2019, 12, 45. [Google Scholar] [CrossRef]
- Halleran, A.D.; Sehdev, M.; Rabe, B.A.; Huyck, R.W.; Williams, C.C.; Saha, M.S. Characterization of Tweety Gene (Ttyh1-3) Expression in Xenopus Laevis During Embryonic Development. Gene Expr. Patterns 2015, 17, 38–44. [Google Scholar] [CrossRef]
- Stefaniuk, M.; Swiech, L.; Dzwonek, J.; Lukasiuk, K. Expression of Ttyh1, a Member of the Tweety Family in Neurons in Vitro and in Vivo and Its Potential Role in Brain Pathology. J. Neurochem. 2010, 115, 1183–1194. [Google Scholar] [CrossRef]
- Cairncross, J.G.; Wang, M.; Jenkins, R.B.; Shaw, E.G.; Giannini, C.; Brachman, D.G.; Buckner, J.C.; Fink, K.L.; Souhami, L.; Laperriere, N.J.; et al. Benefit from Procarbazine, Lomustine, and Vincristine in Oligodendroglial Tumors Is Associated with Mutation of Idh. J. Clin. Oncol. 2014, 32, 783–790. [Google Scholar] [CrossRef]
- Goslin, K.; Schreyer, D.J.; Skene, J.H.; Banker, G. Development of Neuronal Polarity: Gap-43 Distinguishes Axonal from Dendritic Growth Cones. Nature 1988, 336, 672–674. [Google Scholar] [CrossRef]
- Skene, J.H.; Jacobson, R.D.; Snipes, G.J.; McGuire, C.B.; Norden, J.J.; Freeman, J.A. A Protein Induced During Nerve Growth (Gap-43) Is a Major Component of Growth-Cone Membranes. Science 1986, 233, 783–786. [Google Scholar] [CrossRef]
- Lavenius, E.; Gestblom, C.; Johansson, I.; Nanberg, E.; Pahlman, S. Transfection of Trk-a into Human Neuroblastoma Cells Restores Their Ability to Differentiate in Response to Nerve Growth Factor. Cell Growth Differ. 1995, 6, 727–736. [Google Scholar] [PubMed]
- Koponen, E.; Lakso, M.; Castren, E. Overexpression of the Full-Length Neurotrophin Receptor Trkb Regulates the Expression of Plasticity-Related Genes in Mouse Brain. Brain Res. Mol. Brain Res. 2004, 130, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Haag, D.; Zipper, P.; Westrich, V.; Karra, D.; Pfleger, K.; Toedt, G.; Blond, F.; Delhomme, N.; Hahn, M.; Reifenberger, J.; et al. Nos2 Inactivation Promotes the Development of Medulloblastoma in Ptch1(+/-) Mice by Deregulation of Gap43-Dependent Granule Cell Precursor Migration. PLoS Genet. 2012, 8, e1002572. [Google Scholar] [CrossRef] [PubMed]
- Zuber, M.X.; Goodman, D.W.; Karns, L.R.; Fishman, M.C. The Neuronal Growth-Associated Protein Gap-43 Induces Filopodia in Non-Neuronal Cells. Science 1989, 244, 1193–1195. [Google Scholar] [CrossRef]
- Aigner, L.; Arber, S.; Kapfhammer, J.P.; Laux, T.; Schneider, C.; Botteri, F.; Brenner, H.R.; Caroni, P. Overexpression of the Neural Growth-Associated Protein Gap-43 Induces Nerve Sprouting in the Adult Nervous System of Transgenic Mice. Cell 1995, 83, 269–278. [Google Scholar] [CrossRef]
- Liotta, L.A.; Kohn, E.C. The Microenvironment of the Tumour-Host Interface. Nature 2001, 411, 375–379. [Google Scholar] [CrossRef]
- Moschoi, R.; Imbert, V.; Nebout, M.; Chiche, J.; Mary, D.; Prebet, T.; Saland, E.; Castellano, R.; Pouyet, L.; Collette, Y.; et al. Protective Mitochondrial Transfer from Bone Marrow Stromal Cells to Acute Myeloid Leukemic Cells During Chemotherapy. Blood 2016, 128, 253–264. [Google Scholar] [CrossRef]
- Griessinger, E.; Moschoi, R.; Biondani, G.; Peyron, J.F. Mitochondrial Transfer in the Leukemia Microenvironment. Trends Cancer 2017, 3, 828–839. [Google Scholar] [CrossRef]
- Patheja, P.; Sahu, K. Macrophage Conditioned Medium Induced Cellular Network Formation in Mcf-7 Cells through Enhanced Tunneling Nanotube Formation and Tunneling Nanotube Mediated Release of Viable Cytoplasmic Fragments. Exp. Cell Res. 2017, 355, 182–193. [Google Scholar] [CrossRef]
- Nzigou Mombo, B.; Gerbal-Chaloin, S.; Bokus, A.; Daujat-Chavanieu, M.; Jorgensen, C.; Hugnot, J.P.; Vignais, M.L. Mitoception: Transferring Isolated Human Msc Mitochondria to Glioblastoma Stem Cells. J. Vis. Exp. 2017, e55245. [Google Scholar] [CrossRef]
- Hekmatshoar, Y.; Nakhle, J.; Galloni, M.; Vignais, M.L. The Role of Metabolism and Tunneling Nanotube-Mediated Intercellular Mitochondria Exchange in Cancer Drug Resistance. Biochem. J. 2018, 475, 2305–2328. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.S.; Baty, J.W.; Dong, L.F.; Bezawork-Geleta, A.; Endaya, B.; Goodwin, J.; Bajzikova, M.; Kovarova, J.; Peterka, M.; Yan, B.; et al. Mitochondrial Genome Acquisition Restores Respiratory Function and Tumorigenic Potential of Cancer Cells without Mitochondrial DNA. Cell Metab. 2015, 21, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.F.; Kovarova, J.; Bajzikova, M.; Bezawork-Geleta, A.; Svec, D.; Endaya, B.; Sachaphibulkij, K.; Coelho, A.R.; Sebkova, N.; Ruzickova, A.; et al. Horizontal Transfer of Whole Mitochondria Restores Tumorigenic Potential in Mitochondrial DNA-Deficient Cancer Cells. Elife 2017, 6, e22187. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Weinberg, R.A. Micromanagers of Malignancy: Role of Micrornas in Regulating Metastasis. Trends Genet. 2008, 24, 448–456. [Google Scholar] [CrossRef]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour Invasion and Metastasis Initiated by Microrna-10b in Breast Cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Fearon, E.R. Molecular Genetics of Colorectal Cancer. Annu. Rev. Pathol. 2011, 6, 479–507. [Google Scholar] [CrossRef]
- De Roock, W.; de Vriendt, V.; Normanno, N.; Ciardiello, F.; Tejpar, S. Kras, Braf, Pik3ca, and Pten Mutations: Implications for Targeted Therapies in Metastatic Colorectal Cancer. Lancet Oncol. 2011, 12, 594–603. [Google Scholar] [CrossRef]
- Grabocka, E.; Pylayeva-Gupta, Y.; Jones, M.J.; Lubkov, V.; Yemanaberhan, E.; Taylor, L.; Jeng, H.H.; Bar-Sagi, D. Wild-Type H- and N-Ras Promote Mutant K-Ras-Driven Tumorigenesis by Modulating the DNA Damage Response. Cancer Cell 2014, 25, 243–256. [Google Scholar] [CrossRef]
- Dias Carvalho, P.; Guimaraes, C.F.; Cardoso, A.P.; Mendonca, S.; Costa, A.M.; Oliveira, M.J.; Velho, S. Kras Oncogenic Signaling Extends Beyond Cancer Cells to Orchestrate the Microenvironment. Cancer Res. 2018, 78, 7–14. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Bergers, G.; Benjamin, L.E. Tumorigenesis and the Angiogenic Switch. Nat. Rev. Cancer 2003, 3, 401–410. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Biziato, D.; Petrova, T.V. Microenvironmental Regulation of Tumour Angiogenesis. Nat. Rev. Cancer 2017, 17, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Welti, J.; Loges, S.; Dimmeler, S.; Carmeliet, P. Recent Molecular Discoveries in Angiogenesis and Antiangiogenic Therapies in Cancer. J. Clin. Investig. 2013, 123, 3190–3200. [Google Scholar] [CrossRef] [PubMed]
- Astanina, K.; Koch, M.; Jungst, C.; Zumbusch, A.; Kiemer, A.K. Lipid Droplets as a Novel Cargo of Tunnelling Nanotubes in Endothelial Cells. Sci. Rep. 2015, 5, 11453. [Google Scholar] [CrossRef]
- Weller, M.; Cloughesy, T.; Perry, J.R.; Wick, W. Standards of Care for Treatment of Recurrent Glioblastoma--Are We There Yet? Neuro Oncol. 2013, 15, 4–27. [Google Scholar] [CrossRef]
- Lin, Q.; Liu, Z.; Ling, F.; Xu, G. Astrocytes Protect Glioma Cells from Chemotherapy and Upregulate Survival Genes Via Gap Junctional Communication. Mol. Med. Rep. 2016, 13, 1329–1335. [Google Scholar] [CrossRef]
- Chen, W.; Wang, D.; Du, X.; He, Y.; Chen, S.; Shao, Q.; Ma, C.; Huang, B.; Chen, A.; Zhao, P.; et al. Glioma Cells Escaped from Cytotoxicity of Temozolomide and Vincristine by Communicating with Human Astrocytes. Med. Oncol. 2015, 32, 43. [Google Scholar] [CrossRef]
- Tombal, B.; Denmeade, S.R.; Gillis, J.M.; Isaacs, J.T. A Supramicromolar Elevation of Intracellular Free Calcium ([Ca(2+)](I)) Is Consistently Required to Induce the Execution Phase of Apoptosis. Cell Death Differ. 2002, 9, 561–573. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cancer Cells or Cancer-Derived Cell Lines | Cancer-Related Microenvironmental Cells | Cancer Model In Vivo | TNT/TM: Interaction | Ref. | |
|---|---|---|---|---|---|
| Bladder cancer | Human bladder carcinoma cell line 5637 (HTB-9) | CC–CC | [68] | ||
| Human urothelial carcinoma cell line: T24 | Nude mice: xenograft model (tumor growth) | CC–TC | [92] | ||
| Non-malignant urinary papillary urothelial tumor cell line: RT4 | |||||
| Breast cancer | Human breast cancer cell line: MDA-MB231, MCF7 | Human MSCs from bone marrow aspirates of healthy donors | CC–CC | [58,93] | |
| CC–EC | |||||
| Human endothelial cells: E4ORF1-positive cells | CC–MSC | ||||
| Rat mammary adenocarcinoma MTLN3 cells | Murine RAW/LR 5 monocytes/macrophages | CC–macrophages | [94] | ||
| Mouse mammary carcinoma cells (4T1) | CC-CC | [88] | |||
| MDA-MB231, MDA-MB435, MCF7, MDA-MB468, SKBR3 | Human endothelial cells (HUVEC, primary dermal microvascular blood endothelial cells, primary dermal microvascular lymph endothelial cells) | CC-CC | [95] | ||
| CC-EC | |||||
| Human endothelial cell line: HMEC cells | (TNT-like structures: nanoscale intercellular membrane bridges) | ||||
| Cervix cancer | Human cervix carcinoma derived cell line: HeLa | Human NRK fibroblasts | CC–CC | [69,96] | |
| CC–SC | |||||
| Colon carcinoma | Human colon cancer cell lines: LoVo, HCT116, SW480, HT29, AAC1 | CC–CC | [97] | ||
| Human colon carcinoma cell line SW620 | CC–CC | [98] | |||
| Glioma | Human primary glioblastoma cell lines: patient-derived from resected glioblastomas (BTPCs): S24, T269, T325, T1, P3, T1, T269, WJ, BG5, E2 | Rat primary cultures of cortical neurons | NMRI nude mice and NSG mice (patient-derived xenograft models) | In vivo and in vitro: CC–CC | [38,75,82,87] |
| Human oligodendroglioma cell lines (BT088 and BT054) | Human tumor material from patients with glioblastoma | CC-N | |||
| Rat C6 glioma cells | Rat primary astrocytes | CC–SC | [99] | ||
| Human glioblastoma cell line: U87MG, U87 | CC–CC | [78,88,100] | |||
| Human glioma cell line U251 | CC–CC | [101] | |||
| Mouse glioma cell line GL261 | CC–CC | [88] | |||
| Human primary pericytes (HBVP) | Human fetal cerebral cortex, human glioblastoma tissue samples from primary tumors | SC–SC | [102] | ||
| Laryngeal cancer | Human primary culture of LSCC cells | Human LSCC tissue samples | CC–CC (CC–other cell types: ND) | [86] | |
| Leukemia | Human BCP-ALL cell line: NALM6 and REH | Primary MSCs: human bone marrow aspirates from patients with BCP-ALL | CC–MSC | [103] | |
| Human BCP-ALL cell line: REH, SD1, SEM, TOM1 | Human MSC cell line HS27a, Murine MSC MS5, Primary human MSC (from MNC) | Murine NSG model of disseminated SEM-derived ALL | CC–MSC | [104] | |
| CC–CAF | |||||
| Human primary AML blasts (from patient bone marrow) | BMSC: CD34-positive hematopoietic stem cells from human donor) | In vivo xenograft model (NSG mice) | CC–CC | [105] | |
| CC–BMSC | |||||
| Human T-ALL cell line Jurkat | Primary MSCs: bone marrow aspirates from human donor | CC–MSC | [106] | ||
| Human primary T-ALL cells | |||||
| Lung cancer | Lung adenocarcinoma cells from patients with pleural effusions | CC–CC | [21] | ||
| Tumor specimens from patients with poorly differentiated lung adenocarcinoma | |||||
| Mesothelioma | Human mesothelioma cell lines: MSTO-211H (CRL-2081), JMN, VAMT, H2052, H-Meso | Pleural mesothelioma cells from patients with pleural effusions | CC–CC | [21,56,83,107] | |
| Compared with: human immortalized normal (benign) mesothelium cell lines: LP9, Met5A | Tumor specimens from patients with malignant pleural mesothelioma and poorly differentiated lung adenocarcinoma | CC–TC | |||
| Osteo-sarcoma | Murine osteosarcoma cell line: K7M2 cells | Murine osteoblast cell line: MC3T3 | CC–CC | [57] | |
| CC–SC | |||||
| Ovarian cancer | Human ovarian cancer cell lines: SKOV3 (HTB-77), OVCAR3 (HTB-161) | Human MSCs | Human ovarian cancer explant | CC–CC | [58] |
| CC–EC | |||||
| Human endothelial cells: E4ORF1-positive cells | CC–MSC | ||||
| Human ovarian carcinoma cell line: A2780, C200 | Human immortalized cell line from normal ovarian epithelial cells: IOSE | CC–CC | [57,84,108] | ||
| Human ovarian adenocarcinoma cell line: SKOV3 | CC–SC | ||||
| Pancreatic cancer | Human pancreatic adenocarcinoma cell lines: MIA PaCa, S2013, CAPAN-1, CAPAN-2 | Human pancreatic duct epithelial cell line: HPDE cells | Tumor pieces from patients with pancreatic cancer | CC–CC | [84] |
| CC–SC | |||||
| Pheochromocytoma | Rat pheochromocytoma cells (PC12 cells) | CC–CC | [15,50,51] | ||
| Prostate cancer | Human prostate cancer cell lines: PC-3 and 22Rv1 cells | Human fetal osteoblastic cell line: hFOB | CC–CC | [109] | |
| Human androgen-sensitive prostate adenocarcinoma cells: LNCaP cells | CC–OB | ||||
| Squamous cell carcinoma (SCC) | Human SCC-derived cell lines: SCC2, 38, 40, 42B | Human CAF | Tumor pieces from patients with SCC | CC–CC | [85] |
| CC–CAF |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roehlecke, C.; Schmidt, M.H.H. Tunneling Nanotubes and Tumor Microtubes in Cancer. Cancers 2020, 12, 857. https://doi.org/10.3390/cancers12040857
Roehlecke C, Schmidt MHH. Tunneling Nanotubes and Tumor Microtubes in Cancer. Cancers. 2020; 12(4):857. https://doi.org/10.3390/cancers12040857
Chicago/Turabian StyleRoehlecke, Cora, and Mirko H. H. Schmidt. 2020. "Tunneling Nanotubes and Tumor Microtubes in Cancer" Cancers 12, no. 4: 857. https://doi.org/10.3390/cancers12040857
APA StyleRoehlecke, C., & Schmidt, M. H. H. (2020). Tunneling Nanotubes and Tumor Microtubes in Cancer. Cancers, 12(4), 857. https://doi.org/10.3390/cancers12040857