CD44s Induces miR-629-3p Expression in Association with Cisplatin Resistance in Head and Neck Cancer Cells

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
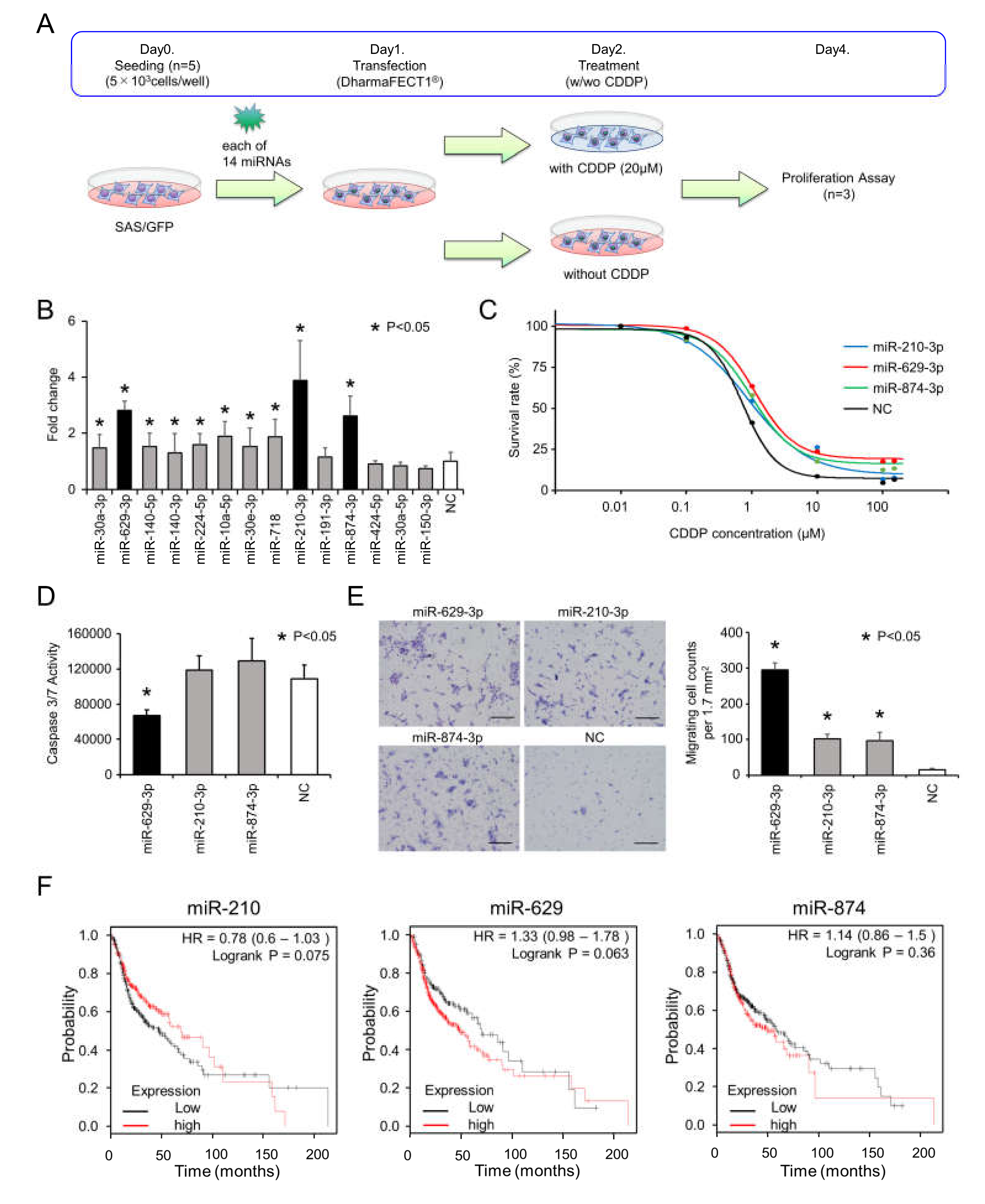
2.1. Search for miRNAs Responsible for CDDP Resistance in Head and Neck Cancer
2.2. miR-629-3p, miR-210-3p, and miR-874-3p are Involved in CDDP Resistance
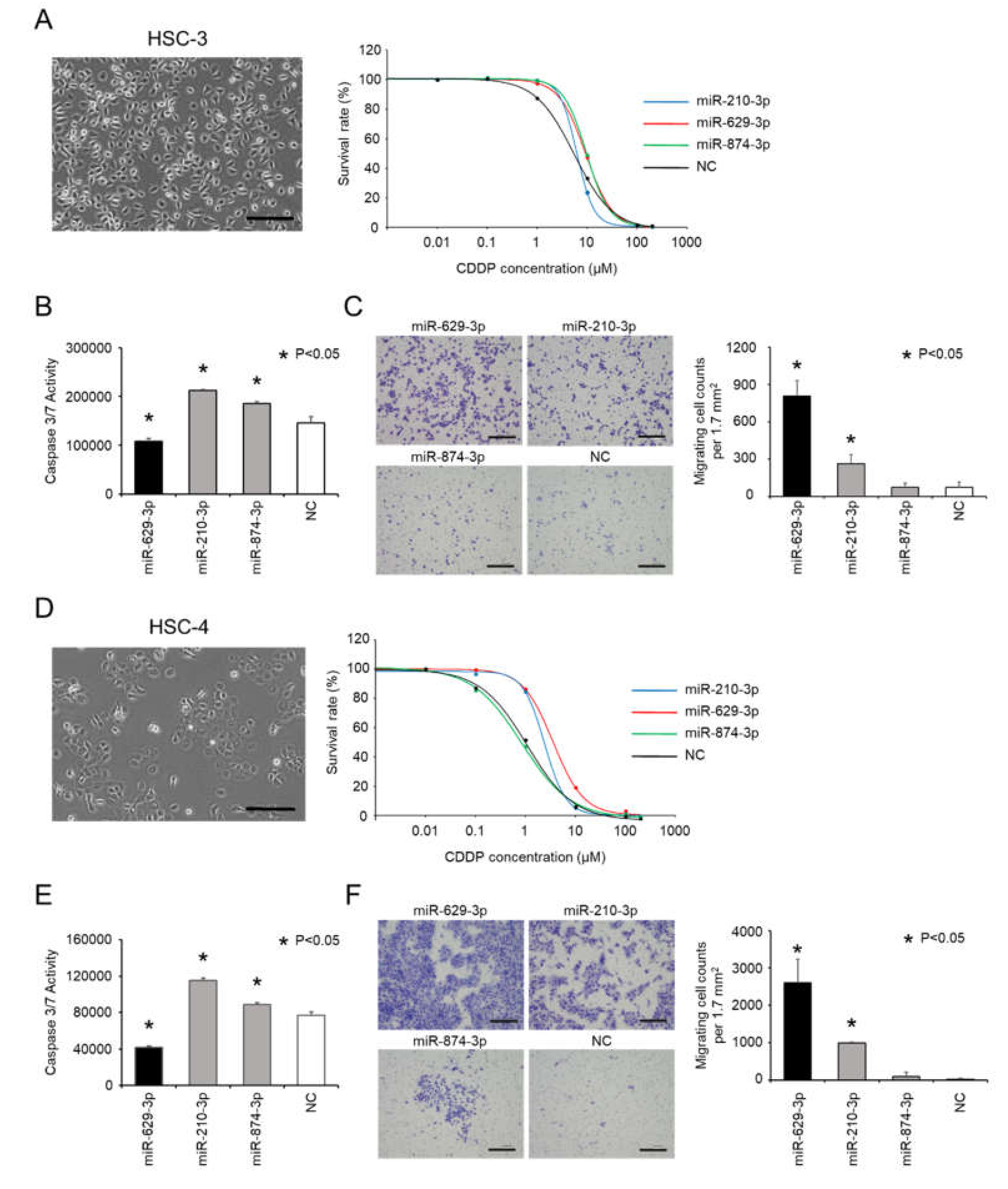
2.3. Confirmation of miR-629-3p Effects on CDDP Resistance in Other Head and Neck Cancer Cell Lines
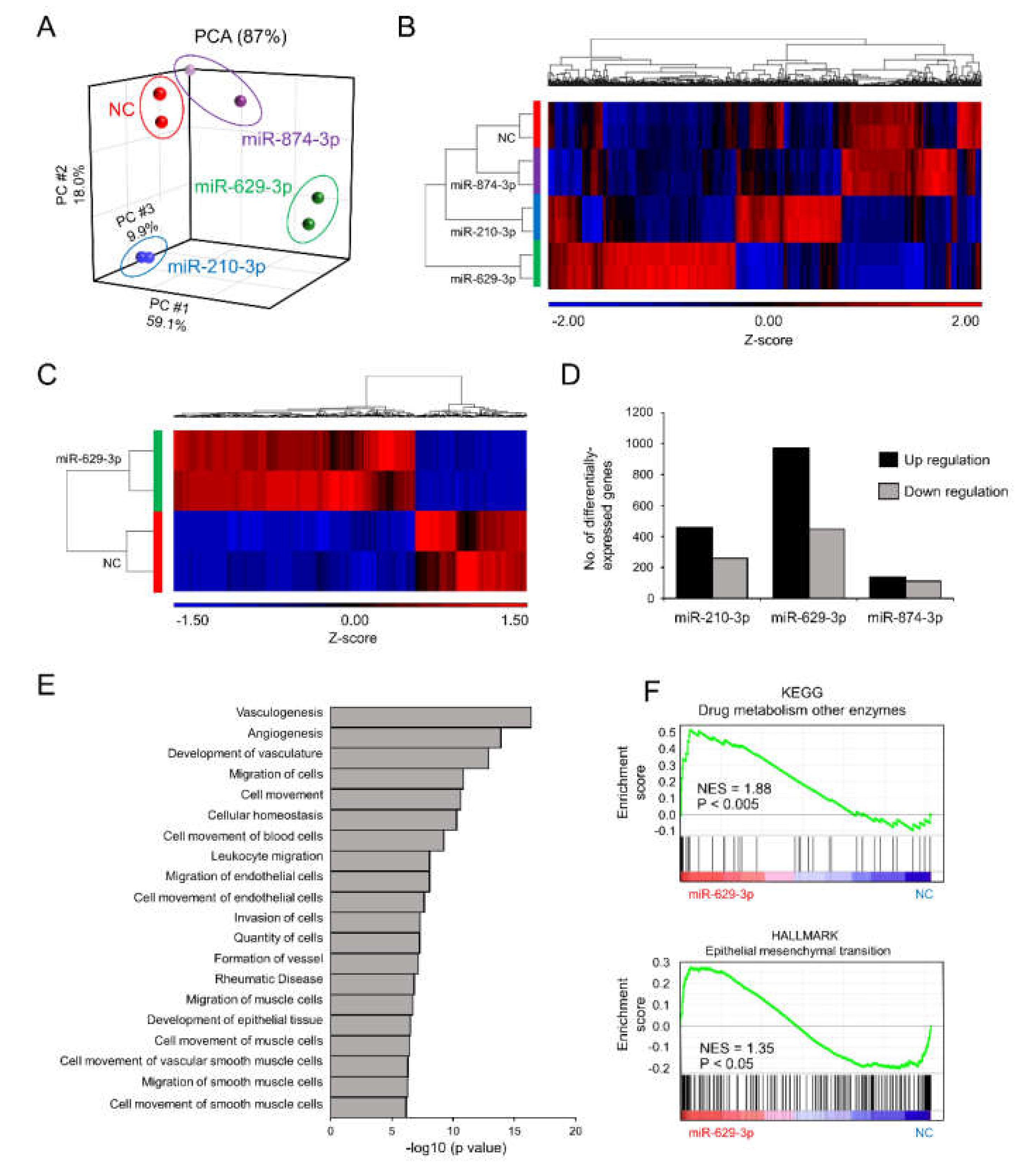
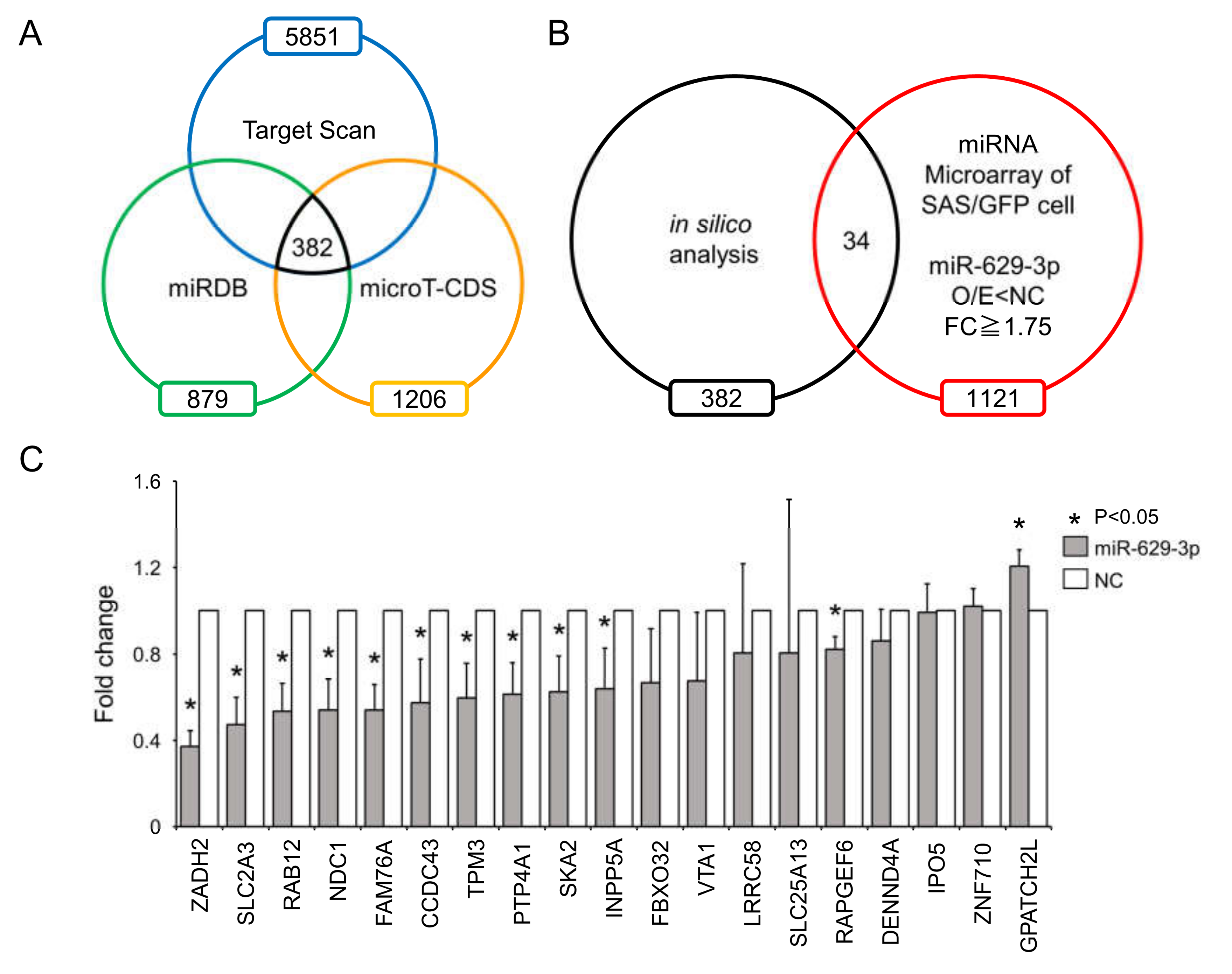
2.4. miR-629-3p Influences Multiple Pathways in Head and Neck Cancer Cells
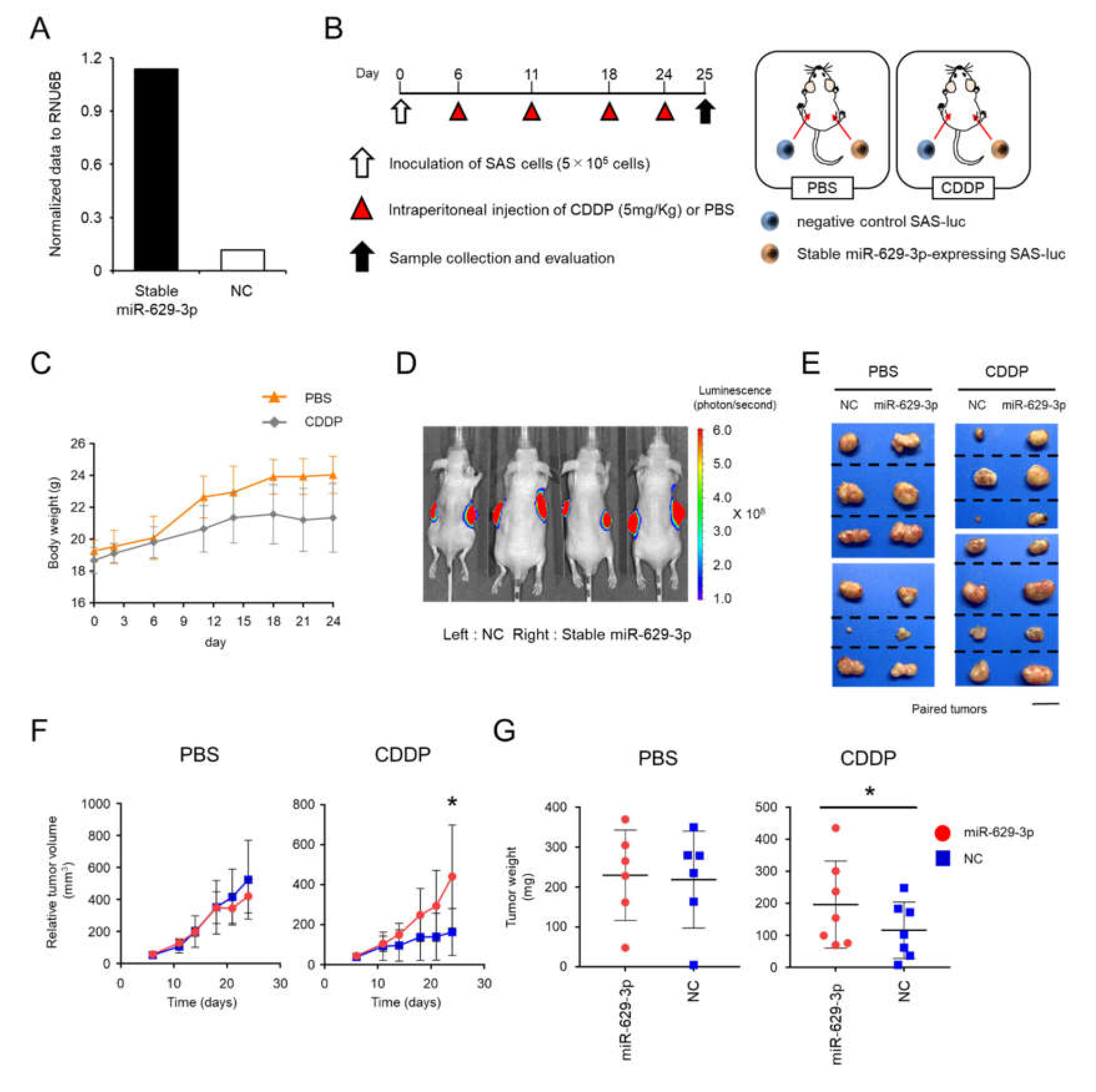
2.5. miR-629-3p Exerts a Functional Role in Drug Resistance in Vivo
3. Discussion
4. Materials and Methods
4.1. Plasmids and Establishment of Stable Cell Line Expressing miR-629-3p
4.2. Cell Lines and Cell Culture
4.3. miRNA Transfections
4.4. Microarray and Bioinformatics
4.5. Microarray Analsysis and Bioinformatics
4.6. Quantitative Reverse Transcription PCR (qRT-PCR)
4.7. Cell Proliferation Assay
4.8. Apoptosis Assay
4.9. Migration Assay
4.10. Animal Experiments
4.11. Kaplan–Meier Plot
4.12. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Editorial Board of the Cancer Statistics in Japan. Cancer Statistics in Japan 2018; Foundation for Promotion of Cancer Research: Tokyo, Japan, 2019; Available online: https://ganjoho.jp/data/reg_stat/statistics/brochure/2018/cancer_statistics_2018.pdf (accessed on 1 March 2019).
- Marcazzan, S.; Varoni, E.M.; Blanco, E.; Lodi, G.; Ferrari, M. Nanomedicine, an emerging therapeutic strategy for oral cancer therapy. Oral Oncol. 2018, 76, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kiyota, N.; Tahara, M.; Fujii, M. Adjuvant treatment for post-operative head and neck squamous cell carcinoma. Jpn. J. Clin. Oncol. 2015, 45, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Kreso, A.; Dick, J.E. Evolution of the Cancer Stem Cell Model. Cell Stem Cell 2014, 14, 275–291. [Google Scholar] [CrossRef]
- Prince, M.E.; Sivanandan, R.; Kaczorowski, A.; Wolf, G.T.; Kaplan, M.J.; Dalerba, P.; Weissman, I.L.; Clarke, M.F.; Ailles, L.E. Identification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinoma. Proc. Natl. Acad. Sci. USA 2007, 104, 973–978. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Boil. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Song, S.J.; Poliseno, L.; Song, M.S.; Ala, U.; Webster, K.; Ng, C.; Beringer, G.; Brikbak, N.J.; Yuan, X.; Cantley, L.C.; et al. MicroRNA-antagonism regulates breast cancer stemness and metastasis via TET-family-dependent chromatin remodeling. Cell 2013, 154, 311–324. [Google Scholar] [CrossRef]
- Takahashi, R.-U.; Miyazaki, H.; Takeshita, F.; Yamamoto, Y.; Minoura, K.; Ono, M.; Kodaira, M.; Tamura, K.; Mori, M.; Ochiya, T. Loss of microRNA-27b contributes to breast cancer stem cell generation by activating ENPP1. Nat. Commun. 2015, 6, 7318. [Google Scholar] [CrossRef]
- Tavazoie, S.F.; Alarcón, C.; Oskarsson, T.; Padua, D.; Wang, Q.; Bos, P.D.; Gerald, W.L.; Massagué, J. Endogenous human microRNAs that suppress breast cancer metastasis. Nature 2008, 451, 147–152. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Othman, N.; In, L.L.A.; Harikrishna, J.A.; Nagoor, N.H. Bcl-xL Silencing Induces Alterations in hsa-miR-608 Expression and Subsequent Cell Death in A549 and SK-LU1 Human Lung Adenocarcinoma Cells. PLoS ONE 2013, 8, e81735. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.S.; Yap, S.H.; Phuah, N.H.; In, L.L.A.; Nagoor, N.H. MicroRNAs associated with tumour migration, invasion and angiogenic properties in A549 and SK-Lu1 human lung adenocarcinoma cells. Lung Cancer 2014, 83, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, J.; Wang, H.; Wang, Y.; Li, Z.; Pan, Y.; Liu, Q.; Yang, M. Repression of the miR-93-enhanced sensitivity of bladder carcinoma to chemotherapy involves the regulation of LASS2. OncoTargets Ther. 2016, 9, 1813–1822. [Google Scholar] [CrossRef][Green Version]
- Yan, B.; Guo, Q.; Nan, X.-X.; Wang, Z.; Yin, Z.; Yi, L.; Wei, Y.; Gao, Y.-L.; Zhou, K.-Q.; Yang, J.-R. Micro-ribonucleic acid 29b inhibits cell proliferation and invasion and enhances cell apoptosis and chemotherapy effects of cisplatin via targeting of DNMT3b and AKT3 in prostate cancer. OncoTargets Ther. 2015, 8, 557–565. [Google Scholar]
- Miyazaki, H.; Takahashi, R.-U.; Prieto-Vila, M.; Kawamura, Y.; Kondo, S.; Shirota, T.; Ochiya, T. CD44 exerts a functional role during EMT induction in cisplatin-resistant head and neck cancer cells. Oncotarget 2018, 9, 10029–10041. [Google Scholar] [CrossRef]
- Phuah, N.H.; Azmi, M.N.; Awang, K.; Nagoor, N.H. Suppression of microRNA-629 enhances sensitivity of cervical cancer cells to 1′S-1′-acetoxychavicol acetate via regulating RSU1. Onco Targets Ther. 2017, 10, 1695–1705. [Google Scholar] [CrossRef]
- Li, B.; Meng, Y.-Q.; Li, Z.; Yin, C.; Lin, J.-P.; Zhu, D.-J.; Zhang, S.-B. MiR-629-3p-induced downregulation of SFTPC promotes cell proliferation and predicts poor survival in lung adenocarcinoma. Artif. Cells Nanomedicine Biotechnol. 2019, 47, 3286–3296. [Google Scholar] [CrossRef]
- Wang, J.; Song, C.; Tang, H.; Zhang, C.; Tang, J.; Li, X.; Chen, B.; Xie, X. miR-629-3p may serve as a novel biomarker and potential therapeutic target for lung metastases of triple-negative breast cancer. Breast Cancer Res. 2017, 19, 72. [Google Scholar] [CrossRef]
- Tanaka, N.; Kosaka, T.; Miyazaki, Y.; Mikami, S.; Niwa, N.; Otsuka, Y.; Minamishima, Y.A.; Mizuno, R.; Kikuchi, E.; Miyajima, A.; et al. Acquired platinum resistance involves epithelial to mesenchymal transition through ubiquitin ligase FBXO32 dysregulation. JCI Insight 2016, 1, e83654. [Google Scholar] [CrossRef][Green Version]
- Wang, C.; Li, X.; Zhang, J.; Ge, Z.; Chen, H.; Hu, J. EZH2 contributes to 5-FU resistance in gastric cancer by epigenetically suppressing FBXO32 expression. OncoTargets Ther. 2018, 11, 7853–7864. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chikuda, J.; Otsuka, K.; Shimomura, I.; Ito, K.; Miyazaki, H.; Takahashi, R.-u.; Nagasaki, M.; Mukudai, Y.; Ochiya, T.; Shimane, T.; et al. CD44s Induces miR-629-3p Expression in Association with Cisplatin Resistance in Head and Neck Cancer Cells. Cancers 2020, 12, 856. https://doi.org/10.3390/cancers12040856
Chikuda J, Otsuka K, Shimomura I, Ito K, Miyazaki H, Takahashi R-u, Nagasaki M, Mukudai Y, Ochiya T, Shimane T, et al. CD44s Induces miR-629-3p Expression in Association with Cisplatin Resistance in Head and Neck Cancer Cells. Cancers. 2020; 12(4):856. https://doi.org/10.3390/cancers12040856
Chicago/Turabian StyleChikuda, Junichiro, Kurataka Otsuka, Iwao Shimomura, Kagenori Ito, Hiroaki Miyazaki, Ryou-u Takahashi, Masahiro Nagasaki, Yoshiki Mukudai, Takahiro Ochiya, Toshikazu Shimane, and et al. 2020. "CD44s Induces miR-629-3p Expression in Association with Cisplatin Resistance in Head and Neck Cancer Cells" Cancers 12, no. 4: 856. https://doi.org/10.3390/cancers12040856
APA StyleChikuda, J., Otsuka, K., Shimomura, I., Ito, K., Miyazaki, H., Takahashi, R.-u., Nagasaki, M., Mukudai, Y., Ochiya, T., Shimane, T., Shirota, T., & Yamamoto, Y. (2020). CD44s Induces miR-629-3p Expression in Association with Cisplatin Resistance in Head and Neck Cancer Cells. Cancers, 12(4), 856. https://doi.org/10.3390/cancers12040856