Molecular Alterations in Dog Pheochromocytomas and Paragangliomas

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Clinical Findings
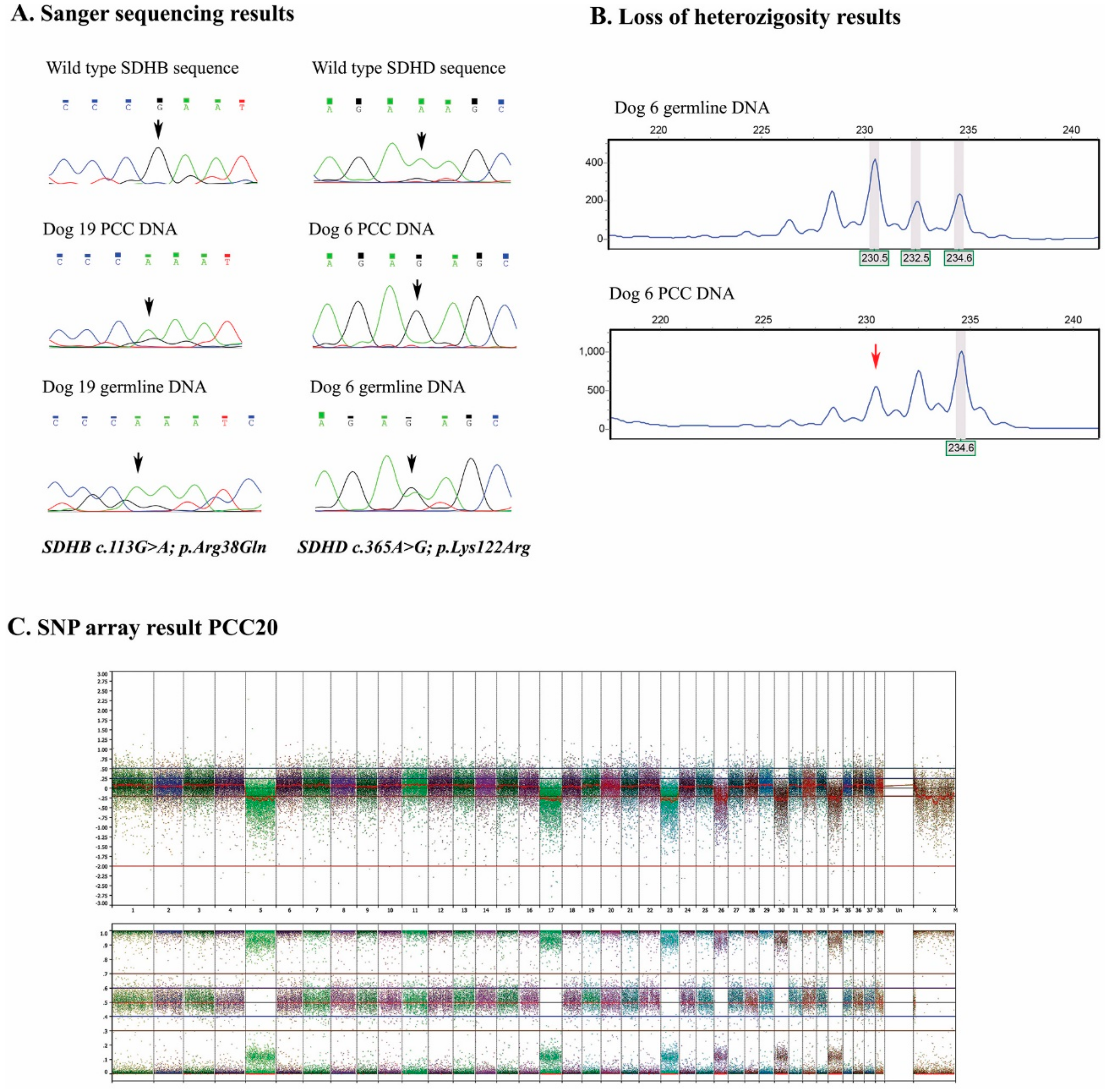
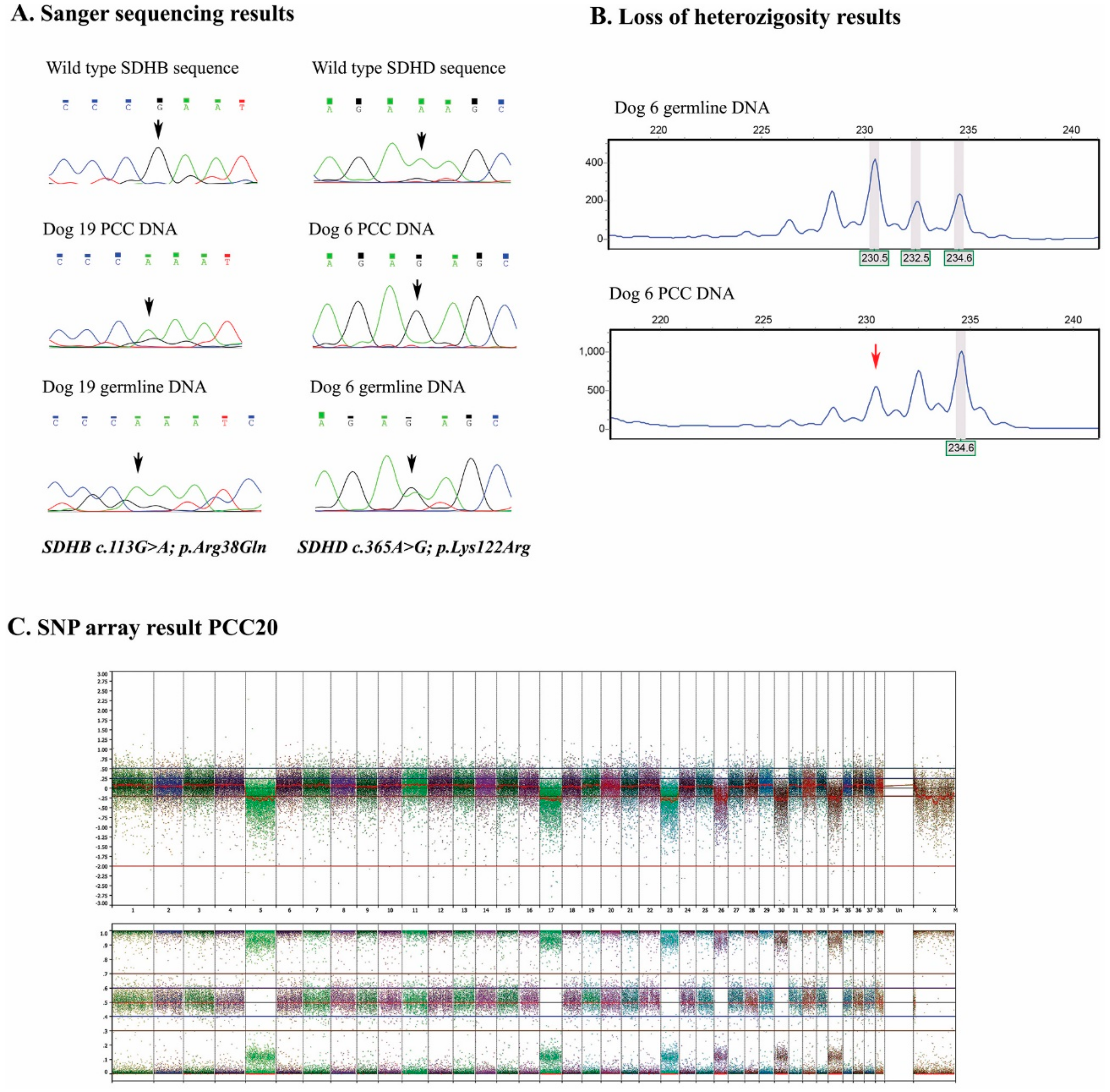
2.2. Genetic Analyses
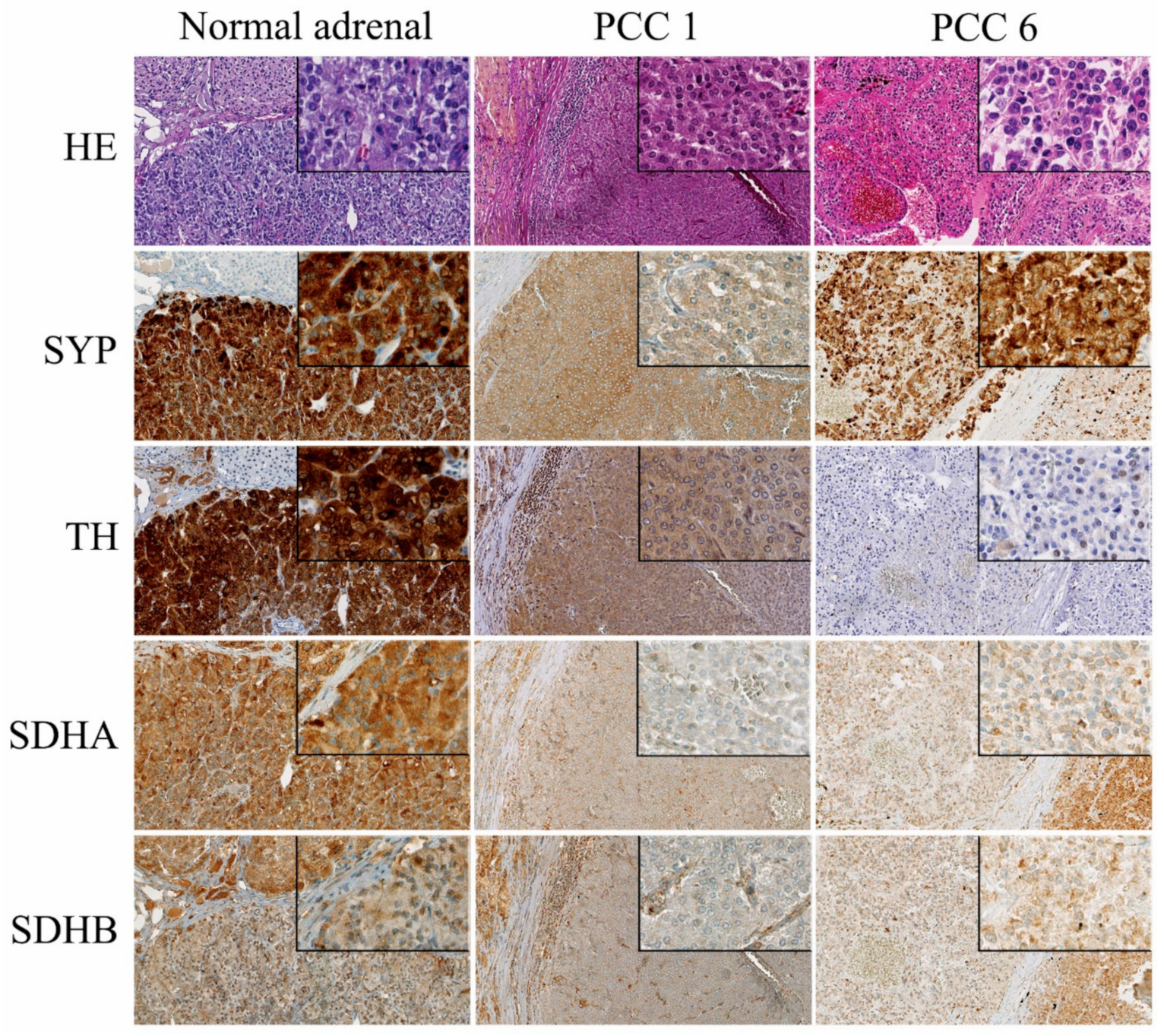
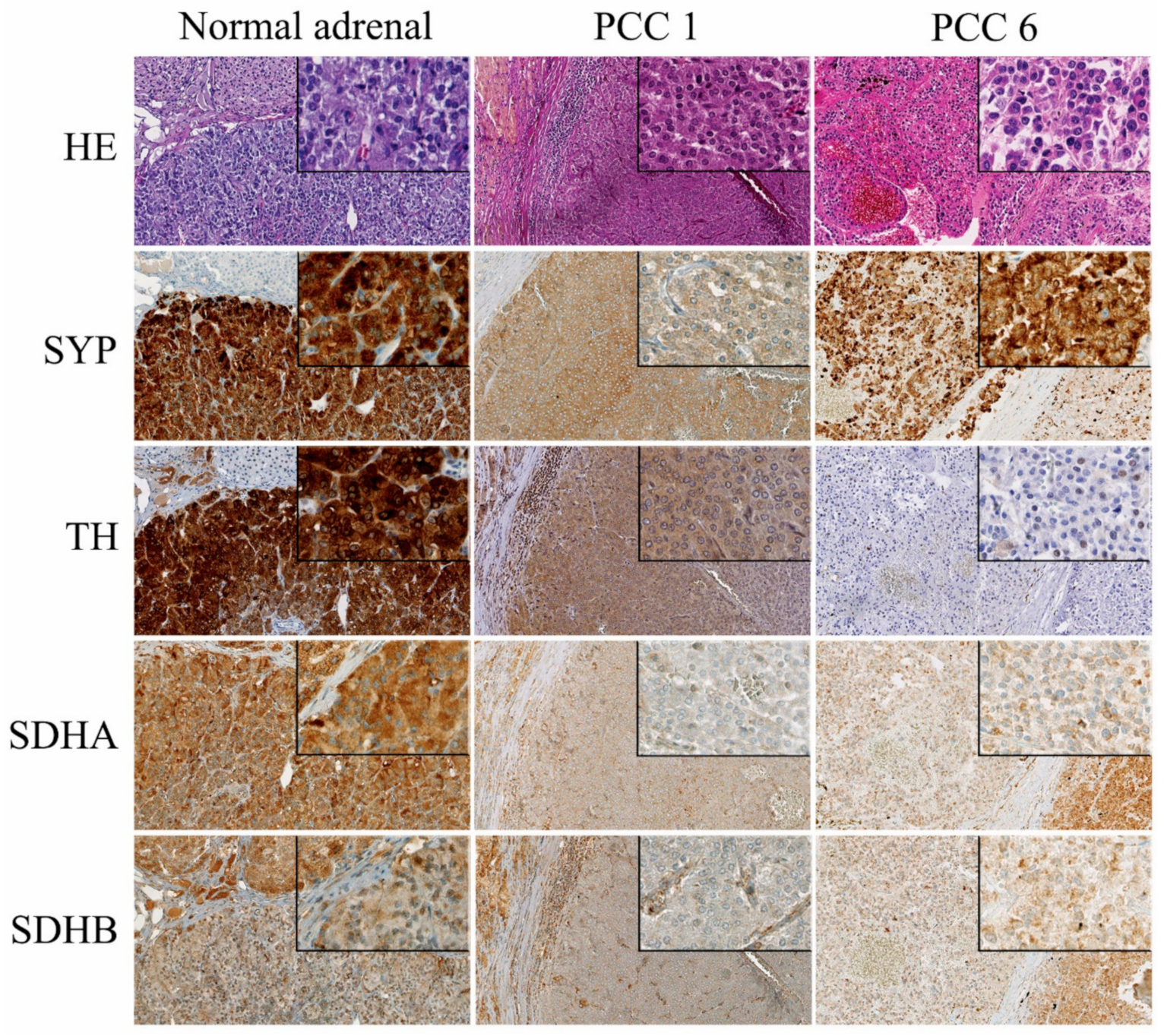
2.3. Immunohistochemistry
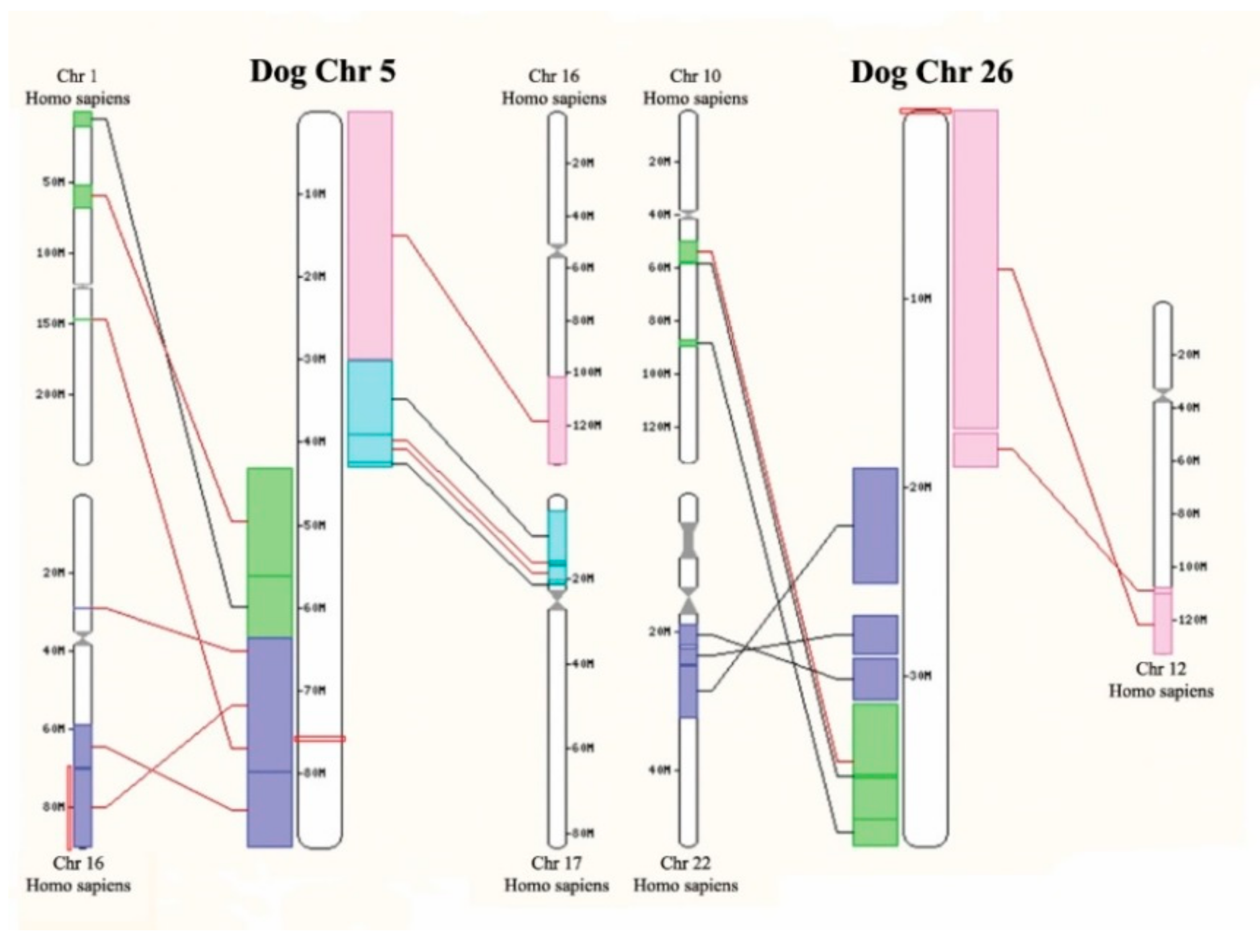
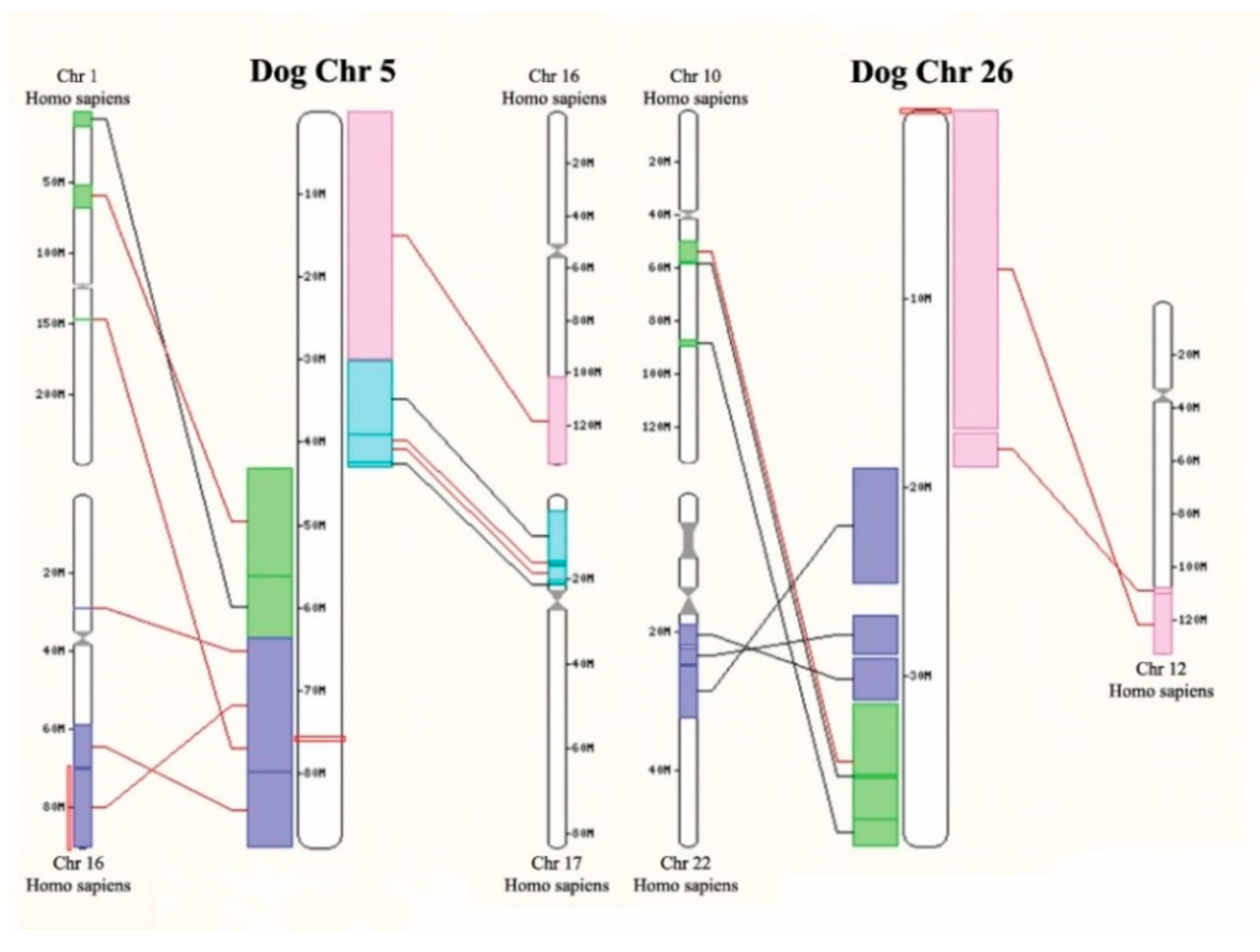
2.4. SNP Arrays
3. Discussion
4. Materials and Methods
4.1. Patients and Sample Selection
4.2. Immunohistochemistry
4.3. Genetic Screening
4.4. Loss of Heterozygosity
4.5. SNP Arrays:
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lenders, J.W.; Eisenhofer, G.; Mannelli, M.; Pacak, K. Phaeochromocytoma. Lancet 2005, 366, 665–675. [Google Scholar] [CrossRef]
- Dahia, P.L. Pheochromocytoma and paraganglioma pathogenesis: Learning from genetic heterogeneity. Nat. Rev. Cancer 2014, 14, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Fishbein, L.; Leshchiner, I.; Walter, V.; Danilova, L.; Robertson, A.G.; Johnson, A.R.; Lichtenberg, T.M.; Murray, B.A.; Ghayee, H.K.; Else, T.; et al. Comprehensive Molecular Characterization of Pheochromocytoma and Paraganglioma. Cancer Cell 2017, 31, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Lenders, J.W.; Duh, Q.Y.; Eisenhofer, G.; Gimenez-Roqueplo, A.P.; Grebe, S.K.; Murad, M.H.; Naruse, M.; Pacak, K.; Young, W.F., Jr. Pheochromocytoma and paraganglioma: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2014, 99, 1915–1942. [Google Scholar] [CrossRef]
- Timmers, H.J.; Kozupa, A.; Eisenhofer, G.; Raygada, M.; Adams, K.T.; Solis, D.; Lenders, J.W.; Pacak, K. Clinical presentations, biochemical phenotypes, and genotype-phenotype correlations in patients with succinate dehydrogenase subunit B-associated pheochromocytomas and paragangliomas. J. Clin. Endocrinol. Metab. 2007, 92, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Lepoutre-Lussey, C.; Thibault, C.; Buffet, A.; Morin, A.; Badoual, C.; Benit, P.; Rustin, P.; Ottolenghi, C.; Janin, M.; Castro-Vega, L.J.; et al. From Nf1 to Sdhb knockout: Successes and failures in the quest for animal models of pheochromocytoma. Mol. Cell. Endocrinol. 2016, 421, 40–48. [Google Scholar] [CrossRef]
- Korpershoek, E.; Pacak, K.; Martiniova, L. Murine models and cell lines for the investigation of pheochromocytoma: Applications for future therapies? Endocr. Pathol. 2012, 23, 43–54. [Google Scholar] [CrossRef]
- Van Nederveen, F.H.; Perren, A.; Dannenberg, H.; Petri, B.J.; Dinjens, W.N.; Komminoth, P.; de Krijger, R.R. PTEN gene loss, but not mutation, in benign and malignant phaeochromocytomas. J. Pathol. 2006, 209, 274–280. [Google Scholar] [CrossRef]
- Burnichon, N.; Buffet, A.; Parfait, B.; Letouze, E.; Laurendeau, I.; Loriot, C.; Pasmant, E.; Abermil, N.; Valeyrie-Allanore, L.; Bertherat, J.; et al. Somatic NF1 inactivation is a frequent event in sporadic pheochromocytoma. Hum. Mol. Genet. 2012, 21, 5397–5405. [Google Scholar] [CrossRef]
- Holt, D.E.; Henthorn, P.; Howell, V.M.; Robinson, B.G.; Benn, D.E. Succinate dehydrogenase subunit D and succinate dehydrogenase subunit B mutation analysis in canine phaeochromocytoma and paraganglioma. J. Comp. Pathol. 2014, 151, 25–34. [Google Scholar] [CrossRef]
- Van Hulsteijn, L.T.; Dekkers, O.M.; Hes, F.J.; Smit, J.W.; Corssmit, E.P. Risk of malignant paraganglioma in SDHB-mutation and SDHD-mutation carriers: A systematic review and meta-analysis. J. Med. Genet. 2012, 49, 768–776. [Google Scholar] [CrossRef]
- Korpershoek, E.; Kloosterhof, N.K.; der Made, A.Zi.; Korsten, H.; Oudijk, L.; Trapman, J.; Dinjens, W.N.; de Krijger, R.R. Trp53 inactivation leads to earlier phaeochromocytoma formation in pten knockout mice. Endocr. Relat. Cancer 2012, 19, 731–740. [Google Scholar] [CrossRef]
- Korpershoek, E.; Loonen, A.J.; Corvers, S.; van Nederveen, F.H.; Jonkers, J.; Ma, X.; der Made, A.Zi.; Korsten, H.; Trapman, J.; Dinjens, W.N.; et al. Conditional Pten knock-out mice: A model for metastatic phaeochromocytoma. J. Pathol. 2009, 217, 597–604. [Google Scholar] [CrossRef]
- Bayley, J.P.; Grimbergen, A.E.; van Bunderen, P.A.; van der Wielen, M.; Kunst, H.P.; Lenders, J.W.; Jansen, J.C.; Dullaart, R.P.; Devilee, P.; Corssmit, E.P.; et al. The first Dutch SDHB founder deletion in paraganglioma-pheochromocytoma patients. BMC Med. Genet. 2009, 10, 34. [Google Scholar] [CrossRef]
- Cascon, A.; Montero-Conde, C.; Ruiz-Llorente, S.; Mercadillo, F.; Leton, R.; Rodriguez-Antona, C.; Martinez-Delgado, B.; Delgado, M.; Diez, A.; Rovira, A.; et al. Gross SDHB deletions in patients with paraganglioma detected by multiplex PCR: A possible hot spot? Genes Chromosomes Cancer 2006, 45, 213–219. [Google Scholar] [CrossRef]
- Richter, S.; Klink, B.; Nacke, B.; de Cubas, A.A.; Mangelis, A.; Rapizzi, E.; Meinhardt, M.; Skondra, C.; Mannelli, M.; Robledo, M.; et al. Epigenetic Mutation of the Succinate Dehydrogenase C Promoter in a Patient with Two Paragangliomas. J. Clin. Endocrinol. Metab. 2016, 101, 359–363. [Google Scholar] [CrossRef]
- Korpershoek, E.; Favier, J.; Gaal, J.; Burnichon, N.; van Gessel, B.; Oudijk, L.; Badoual, C.; Gadessaud, N.; Venisse, A.; Bayley, J.P.; et al. SDHA immunohistochemistry detects germline SDHA gene mutations in apparently sporadic paragangliomas and pheochromocytomas. J. Clin. Endocrinol. Metab. 2011, 96, E1472–E1476. [Google Scholar] [CrossRef]
- Papathomas, T.G.; Oudijk, L.; Persu, A.; Gill, A.J.; van Nederveen, F.; Tischler, A.S.; Tissier, F.; Volante, M.; Matias-Guiu, X.; Smid, M.; et al. SDHB/SDHA immunohistochemistry in pheochromocytomas and paragangliomas: A multicenter interobserver variation analysis using virtual microscopy: A Multinational Study of the European Network for the Study of Adrenal Tumors (ENS@T). Mod. Pathol. 2015, 28, 807–821. [Google Scholar] [CrossRef]
- Castro-Vega, L.J.; Letouze, E.; Burnichon, N.; Buffet, A.; Disderot, P.H.; Khalifa, E.; Loriot, C.; Elarouci, N.; Morin, A.; Menara, M.; et al. Multi-omics analysis defines core genomic alterations in pheochromocytomas and paragangliomas. Nat. Commun. 2015, 6, 6044. [Google Scholar] [CrossRef]
- Van Nederveen, F.; Korpershoek, E.; Deleeuw, R.; Verhofstad, A.A.; Lenders, J.W.; Dinjens, W.; Lam, W.; de Krijger, R. Array-CGH in sporadic benign pheochromocytomas. Endocr. Relat. Cancer 2009, 16, 505–513. [Google Scholar] [CrossRef]
- Gaal, J.; van Nederveen, F.H.; Erlic, Z.; Korpershoek, E.; Oldenburg, R.; Boedeker, C.C.; Kontny, U.; Neumann, H.P.; Dinjens, W.N.; de Krijger, R.R. Parasympathetic paragangliomas are part of the Von Hippel-Lindau syndrome. J. Clin. Endocrinol. Metab. 2009, 94, 4367–4371. [Google Scholar] [CrossRef] [PubMed]
- Galac, S.; Korpershoek, E. Pheochromocytomas and paragangliomas in humans and dogs. Vet. Comp. Oncol. 2017, 15, 1158–1170. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| PCC Number | SDHB Synonymous | SDHB Non-Syn | SDHD Syn | SDHD Non-Syn | Comment |
|---|---|---|---|---|---|
| PCC1 | p.Y50Y, p.Q164Q, p.A210A | WT | |||
| PCC2 | p.Y150Y, p.Q164Q, p.A210A | WT | |||
| PCC3 | WT | WT | |||
| PCC4 | p.A210A | WT | |||
| PCC5 | p.Y150Y, p.Q164Q, p.L188L, p.A210A | WT | |||
| PCC6 | p.Y150Y, p.Q164Q, p.A210A | p.A97A | p.K122RHO/HE | LOH confirmed | |
| PCC9 | p.Y150Y, p.Q164Q, p.A210A | WT | |||
| PCC10 | p.Y150Y, p.Q164Q, p.A210A | WT | |||
| PCC11 | p.L188L | WT | |||
| PCC13 | p.L188L | WT | |||
| PCC14 | p.L188L | WT | |||
| PCC15 | p.Y150Y, p.Q164Q, p.L188L, p.A210A | WT | |||
| PCC16 | p.L188L | WT | |||
| PCC17 | WT | WT | |||
| PCC18 | p.L188L | WT | |||
| PCC19 | p.Y150Y, p.Q164Q, p.A210A | p.R38QHO/HO | WT | loss Chr. 5 | |
| PCC20 | p.L188L | WT | loss Chr. 5 | ||
| PCC21 | p.Y150Y, p.Q164Q, p.L188L, p.A210A | WT | |||
| PCC22 | p.Y150Y, p.Q164Q, p.L188L, p.A210A | WT | |||
| PCC23 | p.Y150Y, p.Q164Q, p.L188L, p.A210A | p.A97A | p.K122RHE | ||
| PCC46 | NA | WT | p.K122RHO | loss Chr.5 |
| Chromosome | PCC19 | PCC20 | PCC36 * | PCC37 * | PCC43 * | PCC46 * |
|---|---|---|---|---|---|---|
| 2 | LOSS | |||||
| 3 | LOSS | |||||
| 5 | LOSS | LOSS | LOSS | LOSS | LOSS | LOSS |
| 7 | LOSS | LOSS | LOSS | |||
| 8 | LOSS | |||||
| 9 | GAIN | |||||
| 12 | LOSS | |||||
| 15 | LOSS | |||||
| 16 | LOSS | LOSS | ||||
| 17 | LOSS | LOSS | ||||
| 18 | LOSS | |||||
| 20 | LOSS | LOSS | ||||
| 21 | LOSS | |||||
| 22 | LOSS | |||||
| 23 | LOSS | |||||
| 25 | LOSS | |||||
| 26 | LOSS | LOSS | LOSS | LOSS | LOSS | |
| 27 | ||||||
| 28 | ||||||
| 29 | LOSS | |||||
| 30 | LOSS | LOSS | ||||
| 31 | LOSS | LOSS | ||||
| 32 | LOSS | LOSS | LOSS | |||
| 34 | LOSS | LOSS | ||||
| 35 | LOSS | LOSS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korpershoek, E.; Dieduksman, D.A.E.R.; Grinwis, G.C.M.; Day, M.J.; Reusch, C.E.; Hilbe, M.; Fracassi, F.; Krol, N.M.G.; Uitterlinden, A.G.; de Klein, A.; et al. Molecular Alterations in Dog Pheochromocytomas and Paragangliomas. Cancers 2019, 11, 607. https://doi.org/10.3390/cancers11050607
Korpershoek E, Dieduksman DAER, Grinwis GCM, Day MJ, Reusch CE, Hilbe M, Fracassi F, Krol NMG, Uitterlinden AG, de Klein A, et al. Molecular Alterations in Dog Pheochromocytomas and Paragangliomas. Cancers. 2019; 11(5):607. https://doi.org/10.3390/cancers11050607
Chicago/Turabian StyleKorpershoek, Esther, Daphne A. E. R. Dieduksman, Guy C. M. Grinwis, Michael J. Day, Claudia E. Reusch, Monika Hilbe, Federico Fracassi, Niels M. G. Krol, André G. Uitterlinden, Annelies de Klein, and et al. 2019. "Molecular Alterations in Dog Pheochromocytomas and Paragangliomas" Cancers 11, no. 5: 607. https://doi.org/10.3390/cancers11050607
APA StyleKorpershoek, E., Dieduksman, D. A. E. R., Grinwis, G. C. M., Day, M. J., Reusch, C. E., Hilbe, M., Fracassi, F., Krol, N. M. G., Uitterlinden, A. G., de Klein, A., Eussen, B., Stoop, H., de Krijger, R. R., Galac, S., & Dinjens, W. N. M. (2019). Molecular Alterations in Dog Pheochromocytomas and Paragangliomas. Cancers, 11(5), 607. https://doi.org/10.3390/cancers11050607